
Method Article
발기인 캡처 안녕-c: 고해상도, 게놈 넓은 발기인 상호 작용의 프로 파일링
* 이 저자들은 동등하게 기여했습니다
요약
강화, 같은 DNA 규제 요소, 물리적으로 대상 유전자 발기인, 큰 게놈 거리에 걸친 장거리 염색체 상호 작용을 통해 자주 연락 하 여 유전자 발현을 제어 합니다. 발기인 캡처 안녕-C (PCHi-C) 그들의 표적 유전자를 잠재적인 규제 시퀀스의 지정을 활성화 발기인 및 원심 지역 사이 중요 한 상호 작용을 식별 합니다.
초록
게놈의 3 차원 조직 기능에 연결 된다. 예를 들어 transcriptional 강화 등 규제 요소 제어 신체 접촉, 종종 상당한 (경우에 따라 수백 kilobases의) 게놈 거리 브리징 및 우회를 통해 그들의 표적 유전자의 spatio 시간적 표현 근처는 유전자 인간 게놈 항구는 약된 1 백만 강화의 대다수는 알 수 없는 유전자 목표. 그들의 표적 유전자를 원심 규제 영역을 지정 하는 것은 따라서 유전자 식 제어를 이해 하는 중요 한 합니다. 우리는 단일 실험에서 모든 발기인에 대 한 발기인 캡처 안녕-C (PCHi-C) 원심 발기인-상호 작용 영역 (Pir)의 게놈 넓은 탐지를 사용 하도록 개발. PCHi-c, 매우 복잡 한이 C 라이브러리는 수천 biotinylated RNA baits 모든 발기인에 포함 된 금지 파편의 끝에 보완 솔루션에서 하이브리드 선택을 통해 발기인 시퀀스에 대 한 구체적으로 농축 된다. 목표를 다음 풀 다운 발기인 순서 및 그들의 빈번한 상호 작용 파트너 강화 등 다른 잠재적인 규제 요소입니다. 높은 처리량 쌍-엔드 시퀀싱, 후 통계 시험 제한 조각 수준에서 상당한 Pir를 식별 하기 위해 각 발기인 출혈 제한 부분에 적용 됩니다. 우리 인간의의 마우스 세포 유형 장거리 발기인 상호 작용의 아틀라스 생성 PCHi-C를 이용 했다. 이 발기인 위하여 지도 그들의 표적 유전자를 putative 규제 지역 지정 및 우대 공간 발기인-발기인 상호 작용 네트워크를 공개 하 여 포유류 유전자 식 컨트롤의 더 큰 이해에 공헌 했다. 이 정보는 또한 비 코딩 질병 관련 된 연결 하 여 잠재적인 질병 유전자의 식별 인간의 유전자 질환을 이해 하 고 관련성이 있다 변종에 또는 근처에 그들의 표적 유전자를 제어 시퀀스 시퀀스.
서문
게놈의 3 차원 조직 유전자 활성화1,2,3,4 억압을 포함 하 여 핵 프로세스의 범위에는 중요 한 기능 역할을 담당 하는 것을 제안 증거를 축적 ,,56,7,8, 재결합9,10, DNA 복구11, DNA 복제12,13, 그리고 세포 노화14. 먼 강화 그들은 적절 한 spatio 시간적 유전자 식 컨트롤에 대 한 필수적인15,,1617, 규제는 발기인에 공간 근접에서 발견 된다. 증강 삭제 표시 원심 강화 대상 유전자 전사18,19,20,,2122, 그리고 'chromatin 반복 강제'에 대 한 필수적입니다. 설계는 증강 및 Hbb 로커 스에서 그 대상 발기인 사이 tethering transcriptional 활성화23드라이브 충분 한지 보여 줍니다. 또한, 소성 증강의 통제 유전자를가지고 게놈 재배열 부적절 한 유전자 활성화 및 질병24,,2526발생할 수 있습니다. 함께, 이러한 예는 발기인 증강 상호 작용 유전자 제어에 필수적인 고 적절 한 유전자 발현 되도록 꽉 규제를 필요로 보여 줍니다. 인간의 마우스 genomes 각각 약 1 백만 강화 항구 추정. 이 강화의 대부분에 대 한 대상 유전자, 고 ' 교전 규칙 ' 발기인과 증강 사이 제대로 이해. 그들의 표적 유전자를 transcriptional 강화를 할당에 따라서 포유류 유전자 식 제어 해독에 큰 도전 남아 있다.
3 차원 게놈 아키텍처에 대 한 우리의 이해 3 C27 (염색체 구조 붙 잡음)의 소개 및 그것의 이체28,29,,3031 혁명 되었습니다. . 이러한 기법, 안녕-C (높은 처리량 염색체 구조 붙 잡음)의 가장 강력한는 세포 인구 내 염색체 상호 작용의 전체 앙상블을 식별 하도록 설계 되었습니다. 안녕하세요 C 라이브러리, 일반적으로 수백만의 세포에서에서 생성 되는 인간 게놈32~ 4 kb 조각 사이 약 1011 독립 결 찰 제품 매우 복잡 합니다. 결과, 개별 제한 간의 상호 작용의 안정적이 고 재현 가능한 식별 (예: 발기인 또는 증강 인자 포함)에서 조각으로 C 데이터 가능 하지 않습니다 하지 않는 한이 C 라이브러리 매우 깊은 시퀀싱을 복종 된다 정기적으로 C 라이브러리를 준비 하는 실험실에 대 한 경제적으로 실행 가능한 해결책 아니다. 이 단점을 우회 하 우리 발기인 캡처 안녕-C를 구체적으로 풍부이 C 라이브러리에서 결 찰 제품 발기인을 포함 하는 개발. 우리는 두 가지 이유로 발기인에 집중 했다. 첫째, 수많은 연구에서 (참조 위), 적절 한 유전자 식 수준에 대 한 중요 한 발기인-증강 연락처 표시 되었습니다 하 고 둘째, 발기인은 크게 변형 세포 유형 사이, 동일한 캡처 미끼 시스템 사용할 수 있습니다가 여러 세포 유형 및 조건에서 전기는 규제 회로 우리의 접근 방식은 수만 biotinylated RNA 120mers 발기인을 포함 하이-C 결 찰 제품 및 후속 캡처 streptavidin 입히는 자성 구슬에 보완이 C 라이브러리의 교 잡에서 솔루션에 의존합니다. PCHi C 라이브러리에이 결과 훨씬 상당히 높은 주파수에서 발기인에 출혈 조각의 식별에만 초점을 맞추고 원래이 C 라이브러리에 비해 복잡성 감소.
우리 인간의 숫자와 마우스 세포 유형 잠복 장거리 원심 발기인 상호 작용 영역 상 상속 규제 기능, 유전자 식 컨트롤의 더 나은 이해에 기여 하 PCHi C 뿐만 아니라 비 무작위 사용 핵의 3 차원 공간에서 발기인-발기인 연락처. 연구에서 수많은 셀 종류33,34,35,36,,3738, 발기인 증강 연락처의 수천 수백 매핑한 39, 식별 마우스 배아 줄기 세포7Polycomb 억압 적인 복잡 한 중재 공간 게놈 조직, 세포질 감 별 법37, 중 발기인 interactomes의 대규모 rewiring 시연 38 , 39, 그리고 연결 된 비 코딩 질병 관련 유전자 발기인35변종 시퀀스.
PCHi C DNA 순서 발기인 상호 작용의 게놈 넓은 앙상블을 지도 하는 이상적으로 적합 한 방법 이다. 관련된 접근, 같은 캡처 안녕-C를 연속 게놈 영역 ( 내용참조)의 고해상도 상호 작용 선택된 게놈 영역에 대 한 프로 파일을 얻기 위해 선택의 방법입니다. PCHi C와 캡처 안녕-C는 실험적인 관점 (유일한 차이점은 캡처 시스템의 선택)에서 매우 유사 하 있도록 조언과 지침을 제공 하는 우리 두 방법 모두에 적용 됩니다. 여기, 선물이 PCHi c.에 대 한 자세한 설명 우리 근거와 PCHi C 실험의 디자인, 단계별 PCHi C 라이브러리 생성 프로토콜을 제공 하 고 어떻게 PCHi C 라이브러리의 품질 높은-품질 데이터를 프로토콜의 다양 한 단계에서 모니터링할 수 있습니다 보여줍니다.
프로토콜
1. 포름알데히드 고정
-
셀 준비: 실험 당 2 × 107 셀의 최소와 함께 시작.
- 문화에서 성장 하는 셀에 대 한 문화 매체에 셀 resuspend. Ex vivo 세포에 대 한 1 x Dulbecco의 수정이 글 중간 (DMEM), 10% (vol/vol) 태아 둔감 한 혈 청 (FBS)와 보완에 resuspend.
- 부착 세포에 대 한 문화 매체를 제거 하 고 10% (vol/vol) FBS 실 온 (RT; 20-25 ° C)에서 신선한 매체의 30.625 mL를 추가 합니다.
- 현 탁 액 셀에 대 한 수집 및 셀 400 g x 3 분 제거 상쾌한 20 ° C에서 원심 하 고 다시 10% (vol/vol) 실시간에서 FBS와 매체의 30.625 ml에서 셀 펠 릿을 일시 중단
- 단단한 조직을 트립 신 (0.05% ~ 2.5% 최종 농도, 세포 유형에 따라) 또는 단일 세포 현 탁 액을 얻기 위해 조직 dounce를 사용 하 여. 이 추가 단계 후 세포 현 탁 액 셀 같은 취급.
-
2% (vol/vol)의 최종 농도를 16% 메탄올 무료 paraformaldehyde (앰 풀 오픈 직전 사용)의 4.375 mL를 추가 합니다. 로커에 부드러운 혼합과 RT에서 10 분에 대 한 수정.
주의: Paraformaldehyde은 유해 화학 물질. 적절 한 건강 및 안전 규정을 따릅니다. - 갓 1 M 차가운 글리신의 5 mL을 추가 하 여 반응을 끄다. 부드러운 락 RT에서와 5 분 동안 혼합 하 고 가끔 반전으로 15 분 동안 얼음에 품 어.
-
세척 하 고 고정된 셀을 수집.
- 부착 세포에 대 한 상쾌한을 제거 하 고 접시 벽에 PBS pH 7.4 x 얼음 1의 10 mL를 추가 하 고 그것을 제거. 1 mL의 PBS pH 7.4 x 얼음 1 추가, 세포 스 크레이 퍼와 50 mL 튜브로 이동을 사용 하 여 셀을 수집 합니다. 가능한 많은 세포 수집을 두 번 반복 합니다. 마지막 볼륨 50 mL에 얼음 처럼 차가운 PBS를 합계 한다.
- 현 탁 액 셀, 원심 분리기 세포 760 x g와 5 분, 4 ° C 상쾌한, 제거 하 고 다시 얼음 처럼 차가운 PBS pH 7.4의 50 mL에 셀 펠 릿을 일시 중단.
- G 와 10 분 동안 4 ° C x 400에 세포를 원심 하 고 상쾌한을 조심 스럽게 제거. 셀 펠 릿 스냅 액체 질소에서 동결 하 고 이후 몇 달 동안-80 ° C에 저장 될 수 있습니다.
2. 세포 세포의 용 해
- 갓 준비 얼음 세포의 용 해 버퍼의 50 mL에 셀 펠 릿 다시 일시 중단 (10 mM Tris HCl pH 8, 0.2% (vol/vol) Igepal CA-630, 10 mM NaCl, 그리고 1 개의 정제 프로 테아 제 억제 물 칵테일)와 섞어. 30 분 동안 얼음에 incubate, 가끔 반전으로. 760 x g 와 5 분 동안 4 ° C에서 핵 원심 고 상쾌한을 제거 합니다.
3. 힌dIII 소화
- 1.25 x 제한 버퍼 2 세포 핵을 씻어. 다시 차가운 1.25 x 제한 버퍼 2의 1 mL 펠 릿 셀과 1.5 mL 튜브에 전송 일시 중단 합니다. 760 x g와 5 분 동안 4 ° C에서 핵 스핀 하 고 상쾌한을 제거 합니다.
- 다시 1.25 x 제한 버퍼 2의 1790 µ L에서 셀 펠 릿을 일시 중단 합니다. 5 aliquots, 각 제한 버퍼 2 x 1.25의 358 µ L에 5-10 백만 셀을 확인 합니다.
- thermomixer에서 37 ° C에서 30 분 동안 950 혁명 당 분 (rpm)에서 약 수와 악수 당 10% (wt/vol) SDS의 11 µ L를 추가 합니다. 셀 덩어리 표시, 거품을 피하고, pipetting으로 해리.
- 추가 10%의 75 µ L 트라이 톤 X-100 (vol/vol) 약 수와 950 rpm을 thermomixer에 15 분 동안 37 ° C에서 흔들어. 셀 덩어리 표시, 거품을 피하고, pipetting으로 해리.
-
12 µ L 100 U / µ L 힌dIII 100의 (총에서 1200 단위) 당 약 수를 추가 하 고 37 ° C에서 하룻밤 (O/N)는 thermomixer에서 950 rpm에서 떨고 있는 동안 품 어.
- 소화 컨트롤에 대 한 효소 (소화 되지 않은 컨트롤)을 추가 하기 전에 새로운 튜브에서 샘플 (각 약 수에서 5 µ L)의 25 µ L를 전송 하 고 효소 (소화 컨트롤)을 추가한 후 동일한 절차를 반복 합니다. 이 C 라이브러리와 동일한 방식으로 두 튜브를 품 어.
- 다음 아침에 약 수 당 100 U / µ L 힌dIII (총에서 500 단위)의 5 µ L을 추가 하 고는 thermomixer에서 950 rpm에서 떨고 있는 동안 2 h 37 ° C에서 품 어.
-
소화 제어: (3.5.1 참조) 소화 및 소화 되지 않은 컨트롤에 대 한 수행 crosslink 반전 (6 단계), 페 놀: 클로 프롬 추출, 및 DNA 강수량 (7 단계).
- 힌dIII 사이트에 걸쳐 있는 뇌관 쌍을 디자인 합니다. 같은 지역에서 또 한 쌍의 뇌관 힌dIII 사이트를 하지 않습니다는 디자인. 정량 PCR (Q-PCR)를 위한 뇌관 디자인 Primer3를 사용 하 여 (http://bioinfo.ut.ee/primer3-0.4.0/) 및 다음 매개 변수:
뇌관 크기: 최적의 20 (분: 18, 최대: 27); 뇌관 Tm: 최적의 60 (분: 57, 최대: 63); 뇌관 CG % 콘텐츠: 분: 20, 최대: 80; Amplicon 크기: RT-PCR ~ 100 bp (기존의 PCR ~ 300에 대 한 bp); 라이브러리를 mispriming: 인간 (인간 뇌관) 또는 설치류 및 간단한 (마우스 뇌관). - Q-PCR 4 의미 Cts (임계값 주기)를 수행: Ct [D; H], [H]; 힌dIII 사이트에 걸쳐 있는 뇌관의 쌍 소화 샘플 [D]에서 얻은 Ct [D;-] 소화 샘플 [D] 힌dIII 사이트 [-]; 하지 않습니다는 뇌관의 쌍에서 얻은 Ct [U; H], 소화 되지 않은 샘플 [U] 힌dIII 사이트;는 뇌관의 쌍에서 얻은 Ct [U;-], 소화 되지 않은 샘플 [U] 힌dIII 사이트 [-] 하지는 뇌관의 쌍에서 얻은. 으로 소화의 비율 계산: % 소화 100-100/2 =(Ct[D,H]-Ct[D,-])-(Ct[U,H]-Ct[U,-]).
- 힌dIII 사이트에 걸쳐 있는 뇌관 쌍을 디자인 합니다. 같은 지역에서 또 한 쌍의 뇌관 힌dIII 사이트를 하지 않습니다는 디자인. 정량 PCR (Q-PCR)를 위한 뇌관 디자인 Primer3를 사용 하 여 (http://bioinfo.ut.ee/primer3-0.4.0/) 및 다음 매개 변수:
4입니다. 제한 조각 돌출부의 Biotinylation
- Biotinylation 마스터 믹스 준비: 30.6 µ L 제한 버퍼 2, H2O의 10.2 µ L x 10의 (분자 생물학 등급), 7.65 µ L의 10mm dCTP, 7.65 µ L 10 m m dGTP의, 10 m m dTTP의 7.65 µ L, 0.4 m m biotin-14-dATP의 191.25 µ L 및 5000 U/mL DNA 중 합 효소의 51 µ L 나 큰 ( Klenow) 단편입니다.
- Biotinylation 당 aliquot, 믹스 마스터 믹스의 60 µ L을 추가 하 고 1 시간 700 rpm (thermomixer) 5 떨고 37 ° C에서 품 어, 각 30 s. 1 시간 후 얼음에 aliquots를 배치 합니다.
5. 핵 결 찰
- 결 찰 마스터 믹스 준비: T4 DNA 리가 버퍼, 소 혈 청 알 부 민 10 mg/mL의 51 µ L x 10의 510 µ L (100 x BSA), 물 (분자 생물학 등급), 1754.4 µ L 그리고 1 U / µ L T4 DNA 리가의 127.5 µ L ( 재료의 표참조).
- 결 찰 당 aliquot 믹스 마스터 믹스의 479 µ L을 추가 하 고 4 h 5에 대 한 700 rpm에서 떨고 16 ° C에서 품 어 s는 thermomixer에 모든 2 분.
- 실시간에서 30 분을 품 어
6. Crosslink 반전
- (고속 원심 분리에 적합) 50 mL 원심 분리기 튜브에 모든 aliquots를 결합 한다.
- 10 mg/mL RNase A, 믹스의 62.5 µ L을 추가 하 고 37 ° c.에 30 분 동안 품 어
- 10 mg/mL 가수분해 K, 믹스의 300 µ L을 추가 하 고 37 ° c.에 30 분 동안 품 어
- 65 ° c.에 반응 O/N (또는 적어도 4 h)를 품 어 다음 아침에 10 mg/mL 가수분해 K, 믹스의 300 µ L을 추가 하 고 65 ° c.에 1 시간에 대 한 품 어
7. DNA 정화
- 4337.5 µ L TLE 버퍼 (10 mM Tris HCl pH 8.0, 0.1 m m EDTA pH 8.0)와 섞어를 추가 합니다.
- 새로운 50 mL 튜브에 RT와 위 (수성) 단계의 3 분 이전 9 ml 20000 x g 1 볼륨 (10 mL) 페 놀 pH 8.0 10 s, 그리고 원심 분리기에 대 한 소용돌이 추가 합니다.
주의: 페 놀은 유해 화학 물질. 적절 한 건강 및 안전 규정을 따릅니다. - 나머지 수성 단계로 TLE 버퍼의 2 개 mL를 추가, 소용돌이 위해 10 s와 RT 고 3 분에 대 한 20000 x g 에서 원심 전송 새로운 튜브에 수성 단계의 2.5 mL에서 단계 7.2, 최종 볼륨을 만드는 11.5 mL. 낮은 (유기) 단계를 포함 하는 튜브를 삭제 합니다.
- 새로운 50 mL 튜브에 RT와 20000 x g 3 분 위 (수성) 단계의 전송 11 ml에 페 놀: 클로 프롬: isoamyl 알콜 (25:24:1), 10, 및 원심 분리기에 대 한 소용돌이의 1 볼륨 (11.5 mL)를 추가 합니다. 7.3 단계를 반복 합니다. 총 샘플 볼륨 이제 13.5 mL 됩니다.
- 1.35 mL 3 M 나트륨 아세테이트 pH 5.2와 33.75 mL의 얼음 감기 100% 에탄올, 믹스의 추가 45 분-80 ° C에서 품 어 또는 또는-20 ° c.에서 하룻밤
- 4 ° C와 10 분 20000 x g 에서 원심 하 고, 상쾌한 제거 하 고 다시 갓 70% (vol/vol) 에탄올의 1 mL에 펠 릿을 일시 중단 한 새로운 튜브로 전송.
- 4 ° C에서 및 벤치탑 원심 분리기에서 3 분 최고 속도로 원심 다음 상쾌한을 제거 합니다.
- 다시 얼음 차가운 70% (vol/vol) 에탄올의 1 mL에 펠 릿을 일시 중단 하 고 반복 단계 7.7. 10 분 동안 37 ° C에서 펠 릿 건조 하 고 다시 TLE 버퍼의 650 µ L에서 일시 중단. 척도 이중 가닥 DNA를 형광 기반 분석 결과 사용 하 여 DNA 수율을 결정 합니다.
참고: 프로토콜 수 수 일시 중지 여기 스냅 동결 하 고 몇 달 동안-80 ° C에 또는 짧은 시간 동안-20 ° C에서 샘플을 저장 하 여.
8입니다. 품질 컨트롤
- DNA 전기 이동 법에 의해 도서관 무결성 및 결 찰을 모니터링 합니다. 0.8 %agarose / 1 x TBE 젤에 라이브러리의 실행 200 ng. DNA 밴드 10 kb 이상으로 실행 해야 합니다.
- 알려진된 셀 형 고정 짧은-및 장거리 상호 작용 기존의 PCR에 의해 감지. PCR 반응 당 DNA 템플릿의 사용 100 ng. 가까이 (참조 3.7.1) 위의 지침에 따라 제한 사이트 쪽으로 PCR 뇌관 디자인. 마우스와 인간의 C 라이브러리의 품질 관리에 대 한 입문 시퀀스는 표 1에 나열 됩니다.
-
채우기 및 결 찰 제어:에서 amplicons 포함 된 젤 밴드 밖으로 잘라 8.2 제어, 젤-추출 DNA, 및 동일한 뇌관 조합 4 개별 PCR 반응에 대 한 템플릿으로 DNA를 사용 하 여.
- Amplicons PCR 정화 키트를 사용 하 여 정화 하 고 DNA 농도 측정.
- 4 소화 반응 (힌dIII [a], NheI [b], 힌dIII + NheI [c]와 아무 효소 [d]) 각 amplicon 15 µ L의 최종 볼륨에 대 한 준비: 500 amplicon의, 제한 버퍼 2.1, 소 혈 청 알 부 민 10 mg/mL의 0.15 µ L x 10의 1.5 µ L (100 x BSA) 그리고 효소 (힌dIII [a], NheI [b], 힌dIII + NheI [c] 또는 물 [d])의 0.1 µ L (10 단위).
- 37 ° C에서 1 시간을 위한 digest 다음 1.5% (wt/vol) agarose/1 x TBE 젤에 소화 반응을 실행 합니다.
9. DNA 파편
- 새로운 튜브에서 샘플의 50.5 µ g를 전송 하 고 각 유리병에 라이브러리 (10 µ g)의 130 µ L을 추가 하 여 5 쥡니다 튜브 ( 재료의 표참조)에 655 µ L. 분할 샘플의 최종 볼륨을 TLE 버퍼를 추가 합니다. ~ 400의 크기에 전단 울트라 sonicator bp ( 재료의 표참조) 다음 매개 변수를 사용 하 여: 의무 비율: 10%; 피크 사고 전력 (w): 140; 버스트 사이클: 200; 시간: 55 s.
- 신선한 2 mL 튜브에서 sonicated 샘플을 수집 합니다.
10. 양면 SPRI-구슬 크기 선택
- 반전에 의해 잘 믹스 SPRI (단단한 단계 가역 Immobilization) 비드 솔루션 새로운 튜브 비드 솔루션의 1.85 mL 전송와 RT 15 분.
- 샘플을 물 (분자 생물학 학년) 350 µ L를 추가 (최종 볼륨 1 mL).
- SPRI 비드 솔루션의 600 µ L 샘플을 추가 (전체 용량 1.6 mL, dna SPRI 솔루션의 비율: 0.6 ~ 1), RT에서 5 분 동안 품 어 및 샘플 샘플 수집을 2-3 s 벤치탑 원심 분리기에서 회전.
- 뚜껑을 열고, 샘플 5 분, 분명 새로운 관으로 표면에 뜨는 전송에 대 한 자력 분리 스탠드에 놓고 구슬을 삭제 합니다.
- SPRI 구슬 두 번째 크기 선택 단계에 대 한 집중: 전송 930 µ L SPRI의 새로운 튜브로 비즈, 취소 표면에 뜨는 5 분 및 삭제에 대 한 자력 분리 스탠드에 배치. SPRI 비드 솔루션의 310 µ L에 다시 구슬을 일시 중단 합니다.
- 샘플을 집중된 SPRI 구슬 (단계 10.5)의 300 µ L를 추가 (전체 용량 1.9 mL, 비율 SPRI dna 솔루션은 이제 0.9 ~ 1), 5 분, RT에서 품 어와 스핀 샘플 2-3 미 신중 하 게 오픈에 대 한 벤치탑 원심 분리기에 뚜껑 5 분, 자력 분리 스탠드에 튜브를 배치 하 고 상쾌한 삭제.
- 30 s, 및 삭제 상쾌한 품 어, 자력 분리 스탠드에 샘플 튜브를 갓된 70% 에탄올 (vol/vol)의 1 mL를 추가 합니다. 두 번 반복 합니다.
- 그리고 더 이상 5 분 추가 300 µ L의 샘플, 믹스, TLE 버퍼에 대 한 thermomixer (튜브 뚜껑 오픈)에서 37 ° C에서 구슬 건조 실 온에서 10 분 동안 품 어.
- 샘플 2-3 s 벤치탑 원심 분리기에서 회전, 뚜껑 열고 장소 5 분 전송에 대 한 자력 분리에 관 서 상쾌한 새로운 튜브로 하 고 구슬 삭제 합니다.
11. 비오 틴/Streptavidin 풀 다운 결 찰 제품의
- 버퍼를 준비: 1 TB 버퍼 x (5 mM Tris HCl pH 8.0, 0.5 m m EDTA, 1 M NaCl, 0.05% 트윈 20); NTB 버퍼 (10 mM Tris HCl pH 8.0; 1 mM EDTA; 2 M NaCl); x 2 1 x NTB 버퍼 (5 mM Tris HCl pH 8.0, 0.5 m m EDTA, 1 M NaCl).
- 새로운 튜브로 자기 streptavidin 결합 구슬 ( 재료의 표참조)의 200 µ L을 추가 하 고 1 분에 대 한 자력 분리 스탠드에 상쾌한 제거.
-
구슬 1 x TB 버퍼의 500 µ L로 두 번 씻는 다.
- 각 세척 단계 동안 biotin 풀 다운, 최종 수리 및 DNA 끝 비 출혈, dATP 미행, 그리고 어댑터 결 찰 단계에서 biotin의 제거에 대 한 다시 해당 버퍼에 비즈를 일시 중단, RT와 3 분 15 rpm에 회전, 스핀, 벤치탑 원심 분리기 튜브 2-3 s 3 분 자력 분리 스탠드에 튜브를 배치 하 고 상쾌한을 제거 합니다.
- 다시 2 x NTB 버퍼의 300 µ L에 비즈를 일시 중단 합니다. 구슬 및 샘플 (600 µ L 총 볼륨)를 혼합 하 고 3 rpm에 회전 바퀴에 15 분 동안 RT에서 품 어.
- 3 분 자력 분리 스탠드에 구슬 회수 하 고 맑은 상쾌한을 제거 합니다. 첫 번째 그리고 1 x 결 찰 버퍼의 200 µ L에 구슬 1 x NTB 버퍼의 500 µ L에 두 번 세척. 다시 10 x 결 찰 버퍼의 50 µ L에 비즈를 일시 중단 합니다.
12. 최종 수리 및 비 출혈 DNA 끝에 Biotin의 제거
- 2.5 m m dNTP 믹스 (각 dNTP의 10 mM의 12.5 µ L), 18.1 µ L의 3000 U/mL T4 DNA 중 합 효소, 10000 U/mL T4 PNK의 18.1 µ L의 50 µ L 샘플 (총에서 50 µ L) 결합 5000 U/mL DNA 중 합 효소의 3.7 µ L 나 큰 (Klenow) 조각 그리고 360.1 µ L H2o.
- 혼합 및 20 ° C 5를 떨고 1 h에서 품 어 s 700 rpm는 thermomixer에서 매 2 분.
- 자력 분리 스탠드에 구슬 회수, 분명 상쾌한 제거 하 고 구슬을 TB 버퍼 x 1의 500 µ L에 두 번 세척.
- 500 µ L 1에의 한 세척 뒤 1 x NTB 버퍼의 500 µ L에 비즈를 씻어 x TLE.
- 자력 분리 스탠드에 구슬 회수, 분명 상쾌한을 제거 하 고 다시 1 x TLE 버퍼의 415 µ L에 비즈를 중단.
13. dATP 미행
- 제한 버퍼 2, 5 µ L의 10mm dATP, 그리고 30 µ L 5 U / µ L Klenow 엑 소-마이너스의 x 10의 50 µ L에 샘플 (415 µ L) 결합.
- 5을 흔들어 30 분 동안 37 ° C에서 품 어와 혼합 s 700 rpm는 thermomixer에서 매 2 분.
- 자력 분리 스탠드에 구슬 회수, 분명 상쾌한 제거 하 고 구슬을 TB 버퍼 x 1의 500 µ L에 두 번 세척.
- 1 x NTB 버퍼의 500 µ L에 비즈를 씻어.
14. 어댑터 결 찰
- 결 찰 반응 버퍼 x 1의 200 µ L에 비즈를 세척 ( 재료의 표참조).
- 다시 1 x 결 찰 반응 버퍼의 200 µ L에 비즈를 일시 중단 합니다. DNA 리가의 4 µ L 추가 ( 재료의 표참조) 및 15 µ M의 16 µ L 미리 단련 PE 어댑터 (동등한 양의 PE 어댑터 1 및 PE 어댑터 2 (둘 다 30 µ M)를 혼합 RT에서 몇 분 동안 배양 하 여 PE 어댑터 anneal 사전). 15 분 동안 RT에서 품 어.
- 자력 분리 스탠드에 구슬 회수, 분명 상쾌한 제거 하 고 구슬을 TB 버퍼 x 1의 500 µ L에 두 번 세척.
- 1 x NTB 버퍼의 500 µ L에 비즈를 씻어. 제한 버퍼 2, x 1의 100 µ L, 세척 구슬 다시 제한 버퍼 2, x 1의 50 µ L에 비즈를 일시 중단 하 고 새 튜브로 전송.
15. 안녕-C 라이브러리 확대
- PCR 마스터 믹스 준비: 100 µ L의 5 x Phusion 버퍼; 25 µ M PE PCR 뇌관 1.0;의 6 µ L 25 µ M PE PCR 뇌관 2.0;의 6 µ L 14 µ L dNTP 믹스 (10 mM 각); 6 µ L의 Phusion 중 합 효소; 318 µ L H2o.
- (총에서 500 µ L), 구슬과 혼합 PCR 마스터 믹스 50 µ L의 10 aliquots에서 분할 하 고 다음 조건을 사용 하 여 PCR에 의해 증폭:
98 ° C에서 30
7의 주기: 10 s 98 ° c; 30 s 65 ° c; 30 s 72 ° C에서
72 ° C에서 7 분 - 새로운 튜브로 PCR 반응 수집, 새로운 관으로 자력 분리 스탠드, 그리고 표면에 뜨는 전송에 구슬 (500 µ L)를 되 찾기.
-
SPRI 비즈를 사용 하 여 라이브러리 DNA를 정화.
- 믹스 SPRI 구슬, 새로운 튜브에 구슬의 460 µ L를 전송 하 고 rt PCR 반응 (최종 볼륨 950 µ L) SPRI 구슬의 15 분 추가 450 µ L에 대 한, RT에서 5 분 동안 품 어 가져오고 샘플 샘플 수집을 2-3 s 벤치탑 원심 분리기에서 회전.
- 뚜껑을 열고, 샘플 5 분, 자력 분리 스탠드에 놓고 상쾌한을 제거 합니다.
- 자력 분리 스탠드에 비즈를 유지, 구슬의 명확한 지역 튜브 샘플, 30 s 및 폐기 상쾌한 두고 70% 에탄올 (vol/vol)의 1 mL를 추가 합니다.
- 15.4.3 두 번 더 반복 합니다.
- 구슬 5 분 더 이상에 대 한 thermomixer (튜브 뚜껑 오픈)에서 37 ° C에서 드라이.
- 샘플, 믹스, TLE 버퍼의 51 µ L을 추가 하 고는 thermomixer에서 950 rpm에서 떨고 37 ° C에서 10 분 동안 품 어.
- 샘플 2-3 s 벤치탑 원심 분리기에서 회전, 뚜껑 열고 장소 5 분 전송에 대 한 자력 분리에 관 서 상쾌한 새로운 튜브로 하 고 구슬 삭제 합니다.
- 이 C 라이브러리의 농도 계량. 7 라운드 후 PCR 확대의, 우리는 정기적으로 C 라이브러리의 500-1500 ng 얻을.
16. 하이브리드 솔루션에서 캡처
참고: 차단기 및 버퍼 (SHS1-4)는 SureSelect에서 솔루션 아래는 키트 ( 재료의 표참조).
- 전송 500 ng 1 µ g의 새로운 튜브로이 C 라이브러리에 진공 집중 장치에 샘플을 증발 하 고 (참조 테이블의 자료, 45 ° C; 진공 압력: 30.0, 램프 5) 드라이까지.
- 3.6 µ L H2O의 (분자 생물학 등급), 차단기 1 차단 2, 2.5 µ L 및 사용자 지정 차단의 0.6 µ L의 2.5 µ L을 추가 하 여 다시 증발된이 C 라이브러리를 일시 중단 합니다.
- 새로운 PCR 튜브 스트립의 우물에 샘플을 전송 PCR 모자 스트립과 가까이 하 고 얼음에. (안녕하세요-C DNA)에 대 한 "D"으로 레이블.
- 교 잡 버퍼 준비: 12.5 µ L의 SHS1 버퍼; SHS2 버퍼;의 0.5 µ L SHS3 버퍼;의 5 µ L SHS4 버퍼의 6.5 µ L입니다.
- thermomixer에서 5 분 65 ° C에서 품 어. 새로운 PCR 튜브 스트립의 우물으로 전송, PCR 모자 스트립과 가까이 및 계속 실시간 라벨에 "h" (교 잡 버퍼).
- 새로운 PCR 튜브 스트립의 우물에 혼합 100 ng / µ L biotinylated의 5 µ L RNA 프로브 (얼음 사용 직전에-80 ° C 및 해 동에 게); SRNase B (RNase 억제제)의 0.5 µ L 및 1.5 µ L H2O의 (분자 생물학 등급).
- PCR 모자 스트립 및 얼음에 PCR 튜브 스트립을 닫습니다. "R" (RNA)에 대 한 라벨.
- PCR 기계는 다음과 같은 매개 변수를 사용 하 여 설정 합니다.
95 ° C에서 5 분 65 ° C에서 25 h 뚜껑 열; 29 µ L PCR 반응 볼륨입니다.
참고: PCR 기계 샘플 증발을 피하기 위하여 실행 되는 동안 모든 절차 동안 최대한 빨리 진행. - "D" PCR 튜브 스트립 PCR 기계에서 PCR 기계 뚜껑 닫아 놓고 PCR 반응을 시작 합니다. PCR 프로그램에 65 ° C에 도달 하면, PCR 기계 뚜껑을 열고 PCR 기계에 "H" PCR 튜브 스트립을 놓습니다. PCR 기계 뚜껑 닫고 3 분 오픈 PCR 기계 뚜껑, 장소 "R" PCR 튜브 PCR 기계에 제거 하 고 닫습니다 PCR 기계에 품 어.
- 2 분 후 PCR 기계 뚜껑 및 모든 PCR 튜브 스트립을 엽니다. 잘 "R"로 잘 "H의" 잘 "R"는, 다음 모든 볼륨에 잘 "D"의 13 µ L를 전송 합니다. 혼합 반응, 3 회 위아래로 피펫으로 닫습니다 PCR 튜브 스트립, 제거 "H"와 "D" PCR 튜브 스트립, PCR 기계 뚜껑을 닫습니다. 24 h 65 ° C에서 반응을 품 어.
17. 격리 조각을 포함 하는 발기인의 결 찰 제품
참고: 다음 단계는 SureSelect 어댑터 키트 및 라이브러리 ( Materals 표참조)와 함께 해야 할 것이 좋습니다.
- 미리 사전에 65 ° C에서 샘플 당 워시 버퍼 2의 1.5 mL를 따뜻한.
- 새로운 튜브로 결합 하는 streptavidin 자석 구슬 ( 재료의 표참조)의 60 µ L을 추가 하 고 1 분에 대 한 자력 분리 스탠드에 상쾌한 제거.
- 구슬 바인딩 버퍼 x 1의 200 µ L로 세 번 세척.
참고: 결 찰 제품 포함 하는 발기인의 포스트 캡처 격리 하는 동안 각 세척 단계에 다시 중단 구슬 해당 버퍼에, 3 분 RT와 회전 바퀴에 15 rpm 회전, 수집을 2-3 s 벤치탑 원심 분리기에 튜브를 부드럽게 회전 샘플, 튜브 자석 분리에 3 분, 및 제거 상쾌한 서. - 다시 바인딩 버퍼 x 1의 200 µ L에 비즈를 일시 중단 합니다. PCR 기계와 PCR 튜브 스트립 (PCR 프로그램은 여전히 실행) 하는 동안 열고 교 잡 반응 자석 구슬 가진 튜브로 전송 합니다. 3 rpm에 회전 바퀴에 30 분 동안 RT에서 품 어.
- 자력 분리 스탠드에 구슬 회수 하 고 맑은 상쾌한을 제거 합니다. 다시 워시 버퍼 1, 믹스의 500 µ L에 비즈를 일시 중단 하 고는 thermomixer에서 950 rpm에서 떨고 있는 동안 20 ° C에서 15 분 동안 품 어.
- 3 분 자력 분리 스탠드에 구슬 회수 하 고 맑은 상쾌한을 제거 합니다. 다시 구슬을 워시 버퍼 2의 500 µ L로 일시 중단, 혼합 하 고는 thermomixer에서 950 rpm에서 떨고 있는 동안 65 ° C에서 10 분을 품 어. 17.5 두 번 더 반복 합니다.
- 자력 분리 스탠드에 구슬 회수, 분명 상쾌한을 제거 하 고 다시 제한 버퍼 2 x 1의 200 µ L에 비즈를 중단. 자력 분리 스탠드에 구슬 회수, 상쾌한 제거 하 고 다시 제한 버퍼 2 x 1의 30 µ L에 비즈를 일시 중단.
18. PCHi C 라이브러리 확대
- PCR 마스터 믹스 준비: 5 x PCR 버퍼 (버퍼 Phusion)의 60 µ L, 3.6 µ L 25 µ M PE PCR 뇌관 1.0, 25 µ M PE PCR 뇌관 2.0, dNTP 믹스 (10 mM 각), Phusion 중 합 효소의 3.6 µ L 및 H2o.의 190.8 µ L의 8.4 µ L의 3.6 µ L의
- (총에서 300 µ L), 구슬과 혼합 PCR 마스터 믹스 50 µ L, 및 다음과 같은 조건을 사용 하 여 PCR 증폭 6 aliquots에 분할:
30 s 98 ° C에
4의 사이클: 10 s 98 ° C, 30에서 65 ° C, 30에서 s s 72 ° C에서
72 ° C에서 7 분 - 새로운 튜브에 모든 PCR 반응 수집, 자석에 구슬 회수 및 상쾌한 전송 (300 µ L; PCHi C 라이브러리를 포함)에 새로운 튜브에.
- SPRI 구슬, 아래 15.4 위에서 설명한 단계를 사용 하 여 PCHi C 라이브러리를 정화.
- 계량 PCHi C 라이브러리의 농도.
결과
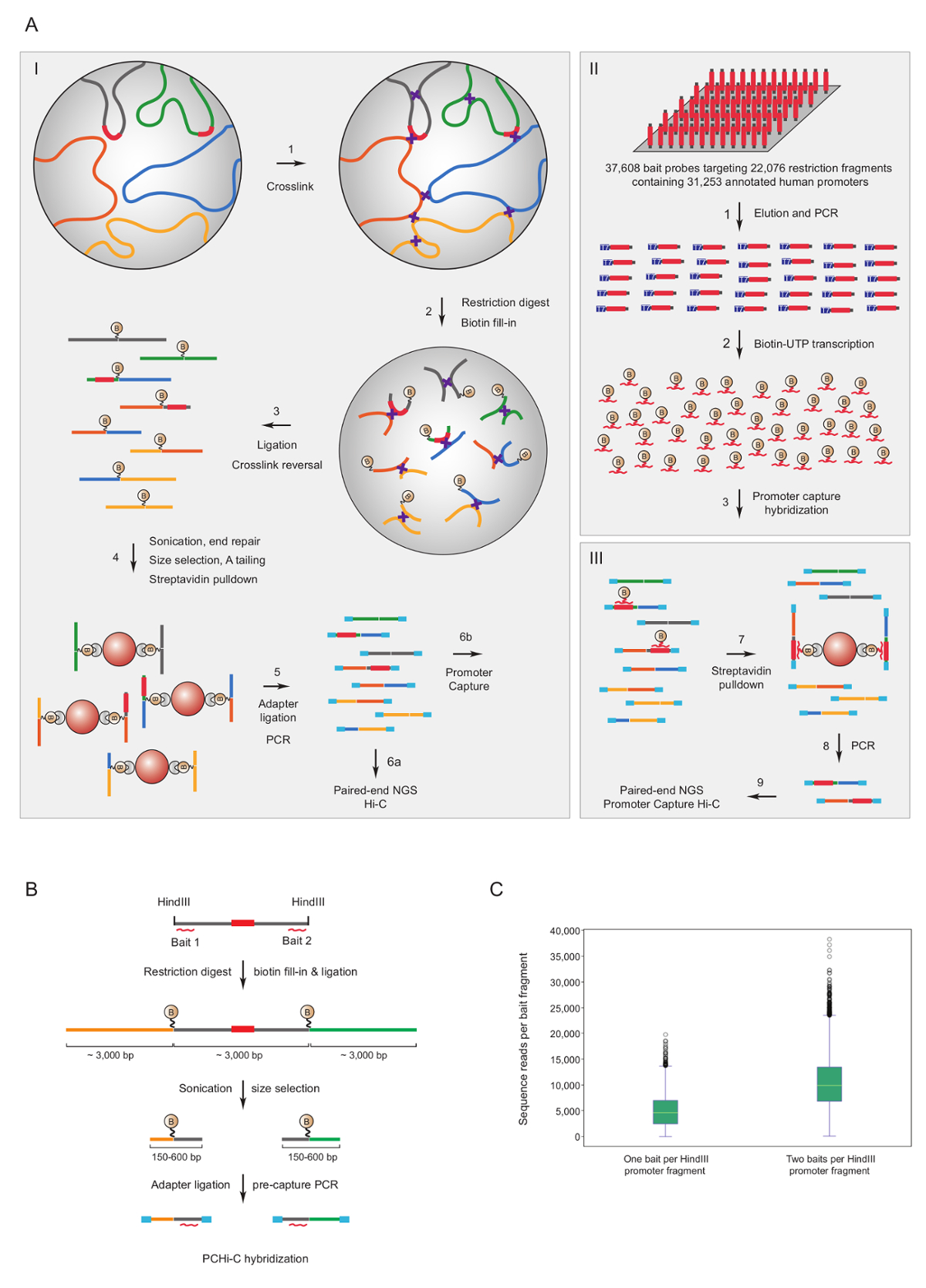
발기인 캡처 안녕-C 마우스7,34,,3639 및 인간의33,35,37,38 이 C 라이브러리를 풍부 하 게 사용 되었습니다. 발기인의 상호 유사한 프로토콜 (HiCap 라는) 샌드 그룹40에 의해 설명 하고있다. 그림 1A 보여주는 도식 워크플로 위한 발기인 캡처 안녕. 여기에 설명 된 프로토콜에서이 C 라이브러리에 핵 결 찰41, 스 퓨 리 어스 결 찰 제품42의 상당히 감소 번호에 어떤 결과 사용 하 여 생성 됩니다. PCHi-C에 대 한 매우 복잡 한 마우스 또는 인간의 C 라이브러리 솔루션에서 교 잡을 복종 되 고 39,021 biotinylated RNAs 22,225 마우스 발기인에 포함 된 HindIII 제한 조각을, 보완 또는 37,608 biotinylated RNAs를 사용 하 여 캡처 타겟팅 22,076 인간의 발기인에 포함 된 HindIII 제한 파편, 각각. 개별 biotinylated RNAs (그림 1B)에 의해 중 하나 또는 둘 다 끝에 금지 파편을 포함 하는 발기인을 대상 수 있습니다. 우리는 예상 대로 둘 다의 캡처의 개별 발기인 (그림 1C, 원시 시퀀스 읽기) 거의 2 배 향상 된 범위를 종료 발견. 따라서, 언제 든 지 가능한 (즉, 비-반복 영역에), 우리를 사용 하 여 biotinylated RNAs 제한 조각의 양쪽을 보완 수 조언.
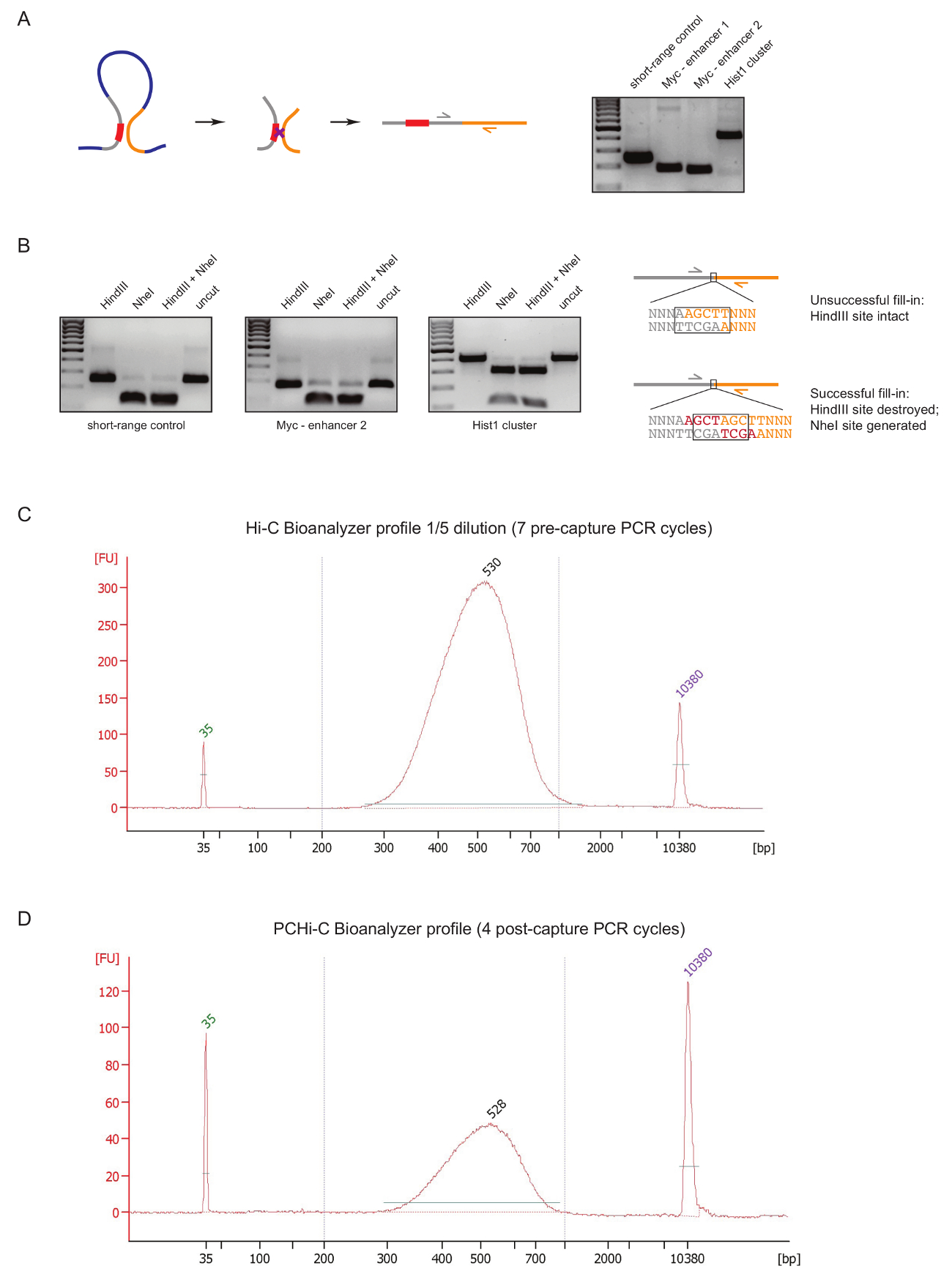
라이브러리 준비 하는 동안 초기 단계에서 PCHi C 라이브러리 품질을 평가, 우리 DNA 결 찰 및 정화, 앞에서 설명한31후 두 개의 컨트롤을 수행 합니다. 첫 번째 사용 하는 특정 뇌관 쌍 3 C27에서 결 찰 제품을 증폭입니다. 뇌관 쌍 (표 1)를 사용 하 여 셀 형 고정 장거리 결 찰 제품을 증폭 하는 우리, 같은: Myc 유전자와 그것의 알려진된 강화 약 2mb 거리 (그림 2A) 또는 Hist1 소재 시 (유전자 사이 구분 하 여 1.5 m b), 그리고 선형 근접 ('단거리 컨트롤')에 위치한 2 개의 지구 사이.
두 번째 품질 관리 biotin dATP와 제한 사이트 돌출부의 Klenow 중재 채우기 동안 biotin 결합의 효율성을 결정 하기 위해 수행 됩니다. 성공적인 Klenow 채우기 및 새로운 NheI 인식 사이트 (그림 2B 의 형성에서 DNA 분자의 결 찰 제품 및 HindIII의 경우 사이 원래 제한 사이트의 실종에서 이후 무뚝뚝한 끝 결 찰 결과 ). NheI 소화 결 찰 제품을 HindIII의 비율은 biotin 결합 효율의 직접 판독 합니다. 반면에 높은-품질 라이브러리는 결 찰 제품 (그림 2B)의 가까운 완전 한 NheI 소화 품질이 C 라이브러리 HindIII 소화의 높은 수준의 표시 됩니다.
준비 후에 C 라이브러리 (즉, biotin streptavidin 풀 다운 안녕 C 결 찰 제품 크기 선택, 어댑터 결 찰 및 캡처 전 PCR 후),이 C 라이브러리의 무결성 및 크기 분포는 Bioanalyzer (그림에 의해 평가 2 C). 동일한 컨트롤 PCHi C 라이브러리 준비 (즉, 결 찰 제품 발기인을 포함 하 고 포스트 캡처 PCR의 교 잡 캡처 후)의 끝에서 수행 됩니다. 안녕-C와 PCHi C Bioanalyzer 프로필의 비교 표시 예상 대로,이 C 라이브러리는 해당 PCHi C 라이브러리 보다 훨씬 더 집중 하지만 라이브러리의 크기 분포는 매우 유사한 캡처 단계에 있음을 나타내는 PCHi C 크기 바이어스 (그림 2C, D)를 도입 하지 않습니다.
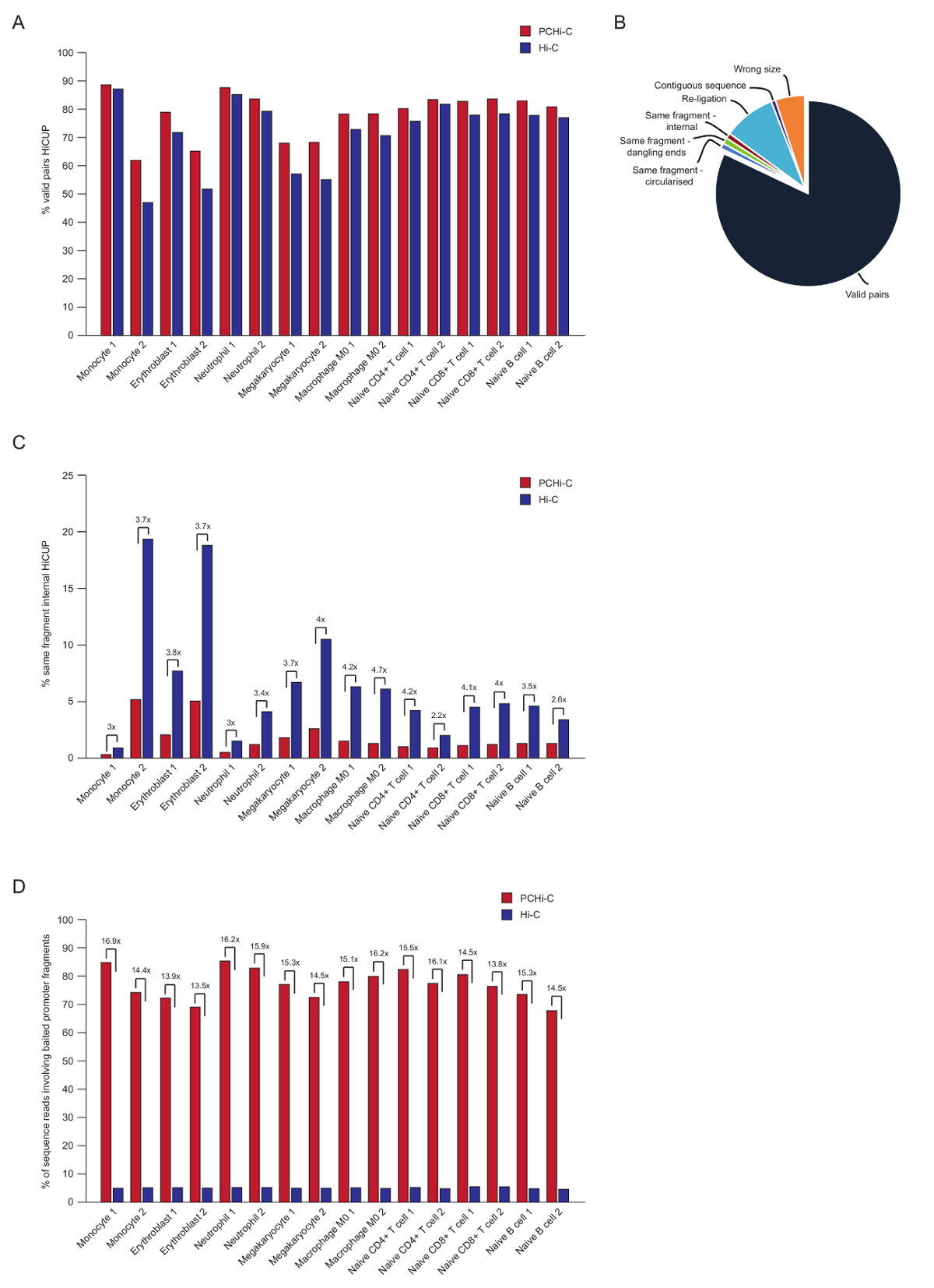
짝-엔드 시퀀싱, 후 PCHi C 읽기 매핑됩니다, 그리고 품질 제어 및 HiCUP 파이프라인43을 사용 하 여 필터링. 70-90% '유효한 쌍' 사이 포함 하는 높은-품질 PCHi C 라이브러리 (즉, 결합 하 여 최종 시퀀스 선형 게놈 지도;에 인접 하지 두 제한 조각 사이 읽습니다 그림 3A, B). 핵 결 찰 프로토콜41,42를 사용 하 여 트랜스 의 백분율 쌍 (즉, 결합 하 여 최종 시퀀스 다른 염색체에 있는 2 개의 제한 파편 사이 읽습니다)는 일반적으로 낮은 읽기 5와 사이 25%, 염색체 영토의 존재를 반영 하 고 품질 좋은 라이브러리를 나타내는. '유효한 쌍'이 C 라이브러리와 그들의 해당 PCHi C 라이브러리35사이 비율의 직접적인 비교, 쇼를 모든 경우에 유효한 쌍의 비율 (그림 3B) PCHi C 라이브러리에서 높은 수준 이다. 이 유효 하지 않은 '같은 조각 내부' 읽기 PCHi-c (그림 3C)의 비율에 있는 감소를 동반 된다. 이것은 예상, 캡처 단계 발기인 포함 된 결 찰 제품을 풍요롭게 하는 뿐만 아니라 또한 제한 조각 끝에 대 한 제한에 캡처 oligos의 위치 때문에 조각 ( 그림 1B를참조).
HiCUP 필터링, 후 우리는 캡처 효율을 결정 합니다. PCHi C 라이브러리 세 가지 유형의 HiCUP 필터링 후 유효한 시퀀스 읽기 포함:
1.) 발기인: 게놈 읽습니다 (캡처된 발기인 조각 및 게놈에서 발기인이 아닌 HindIII 제한 조각 사이즉, 읽기)
2.) 발기인: 발기인 읽습니다 (2 개의 캡처된 발기인 파편 사이 읽기)
3.) 게놈: 게놈 (배경 안녕 C 결 찰 제품 결 찰 제품 파트너의 캡처된 발기인 매핑됩니다)를 읽습니다. 이 다운스트림 분석 전에 삭제 됩니다.
높은-품질 PCHi C 라이브러리는 캡처 효율성 (카테고리 1과 2의 합) (그림 3D) 65-90% 사이. 이 C 라이브러리를 직접 비교를 보여줍니다 PCHi C에 결과 ~ 15 농축 제품에 대 한 발기인-포함 된 결 찰 (그림 3D), 일부의 경우 17-fold. 이 가상 최대 가깝습니다 (19.6-fold) PCHi-c, 게놈 금지 파편의 비율에 따라 농축 캡처 시스템에 의해 보호. 더 농축 캡처 시스템 적은 제한 조각44,,4546를 대상으로 디자인 하 여 구현할 수 있습니다.
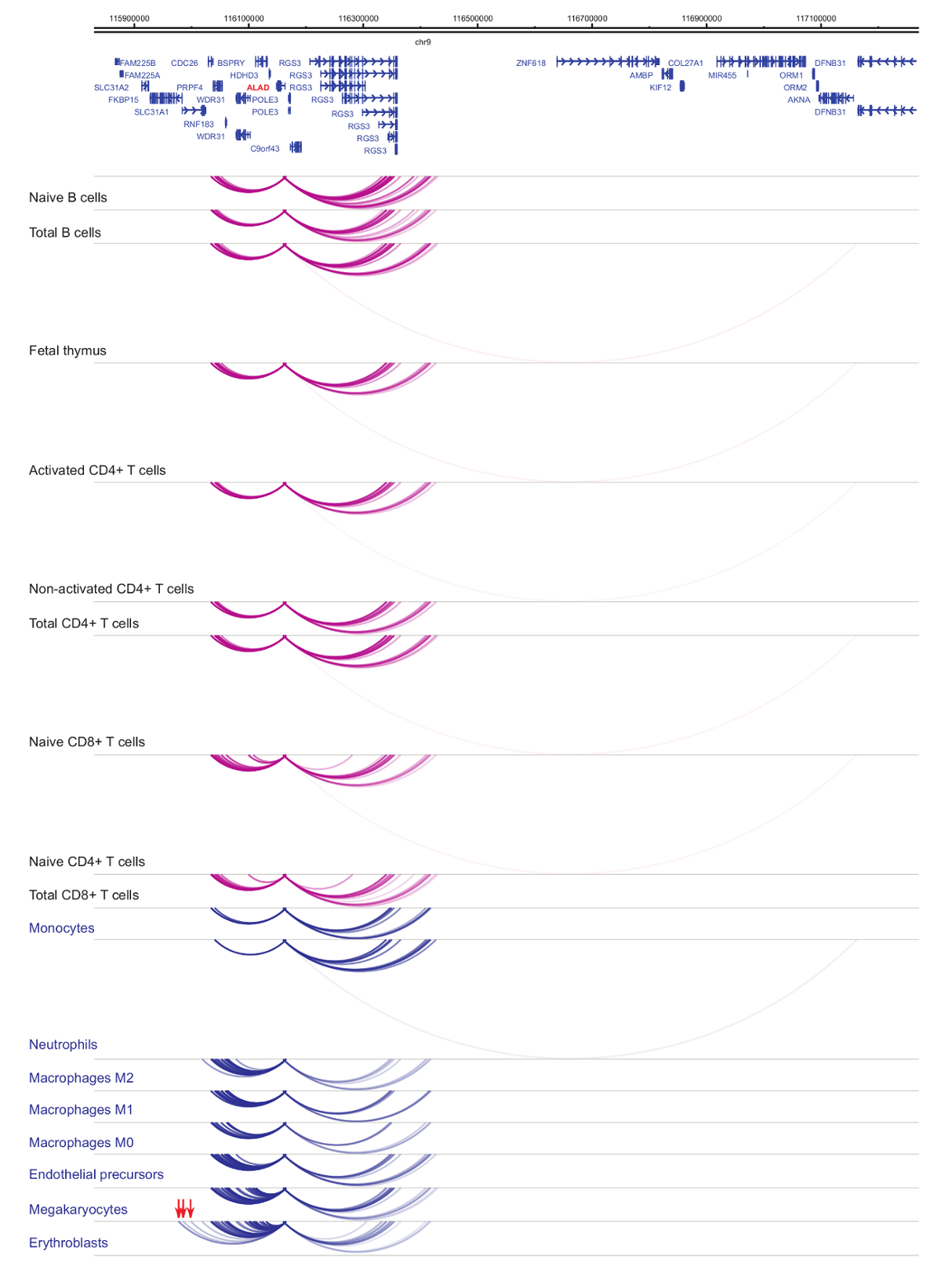
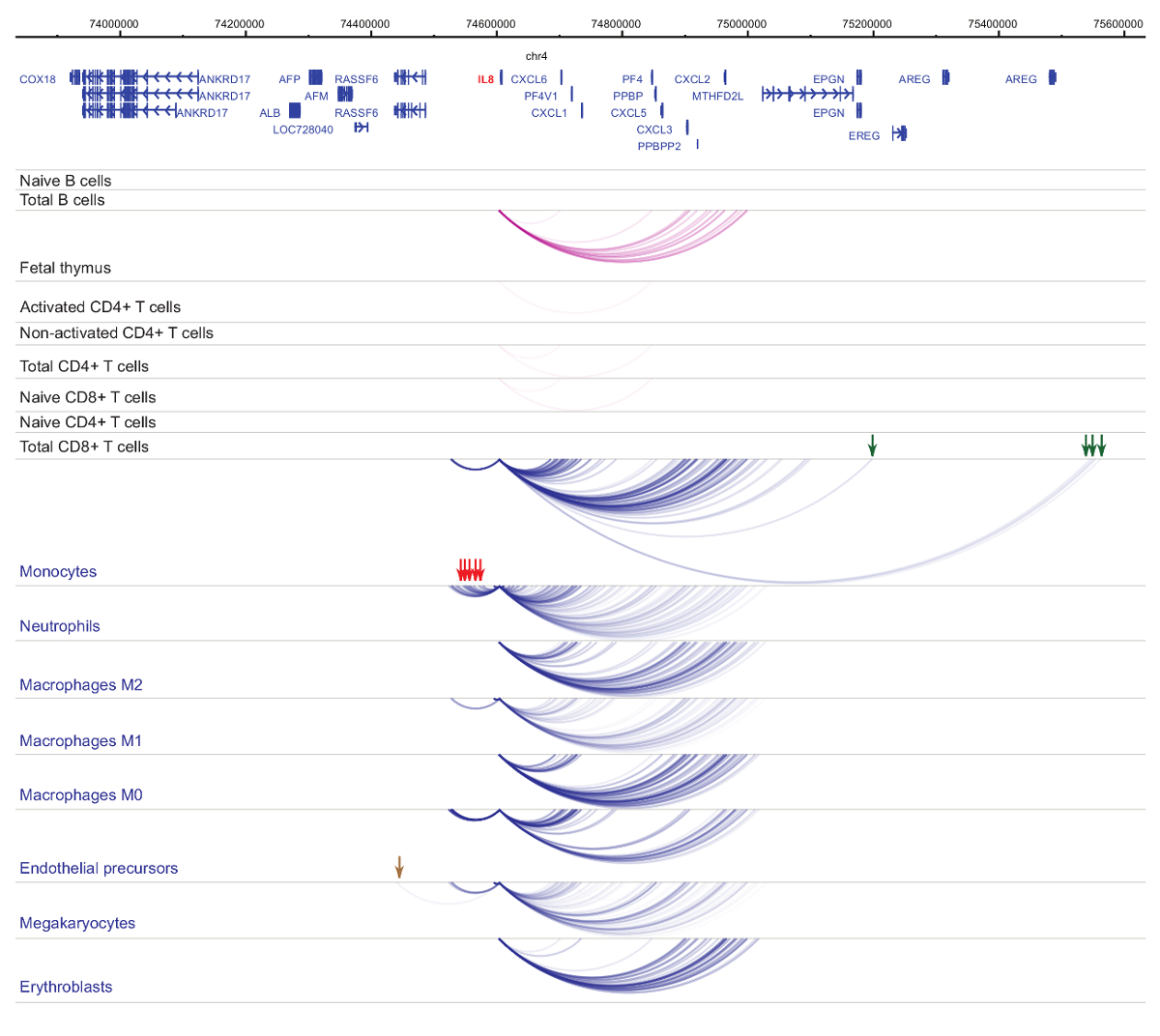
발기인 interactomes의 분석 세포 유형 및 혈통 특이성33,34,35, 세포질 감 별 법37,38,39 동안 발음 변화를 보여줍니다. . 그림 4 와 5 특정 발기인에 혈통 특이성 및 차별화 역학의 예를 표시합니다. 예를 들어 ALAD 는 constitutively 모든 셀에 표현 이지만 그 표현 upregulated erythroblasts47. ALAD 발기인 모든 조 혈 모 세포에서 여러 말 초 조각와 erythroblasts (그림 4)에 구체적으로 추가 상호 작용에 종사 합니다. IL-8 B 세포, T 세포, monocytes, 중 성구 및 없으며 (셀 타입 특정 상호 작용을 포함 하 여, 골수성 계보의 세포에서 상호 작용의 수십에서 거의 상호 작용에 아무 통계적으로 중요 한 상호 작용을 보여줍니다. 그림 5)입니다. 이러한 예 PCHi-C를 사용 하 여 셀 형식을 특정 interactomes를 푸는 규제 가능성으로 발기인 상호 작용 영역을 식별 하는 방법을 보여 줍니다.

그림 1 : 발기인 캡처 안녕-C 근거 및 캡처 미끼 디자인. (A) 회로도 워크플로 PCHi-c 핵 결 찰이 C41,42 (I) 뒤에 biotinylated RNA 미끼 (II)와 교 잡에서 솔루션 대상의 모든 인간의 제한 조각 (여기 묘사) 또는 마우스 유전자 발기인 (III). ((B)) 미끼 디자인 PCHi-c Biotinylated RNA 캡처 미끼 (빨간 곡선)는 발기인에 포함 된 제한 조각 (회색, 발기인 순서 (빨간색) 스스로 타겟으로하는 RNA 캡처 미끼에 의해 제한에 위치 하는 경우 노트의 끝에 대 한 설계 되었습니다. 조각 끝)입니다. 결 찰 발기인에 포함 된 제한 조각 (회색) 및 그들의 상호 작용 제한 조각 (황색 및 녹색)의 구성 된 제품은 미끼 RNA와 DNA 대상 간의 교 잡 시퀀스 complementarity 통해 격리 및 후속 biotin streptavidin 풀 다운, 발기인에 포함 된 제한 조각에 대 한 대답 (C) 비교 PCHi-C의 캡처 효율와 같이 대상 한 RNA 미끼 캡처 프로브 vs 두 RNA 미끼 캡처 프로브 (b에서 회로도 참조). 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 2 : PCHi C 사전 시퀀싱 품질 컨트롤. (A) 왼쪽, 발기인 및 PIR, 발기인에 포함 된 제한 조각 (회색, 빨간색으로 발기인 순서) 및 PIR 제한 조각 (황색)의 구성 된이 C 결 찰 제품 결과 사이 공간 나란히의 도식. 오른쪽, DNA 젤 전기 이동 법이 C 결 찰 제품 (로 왼쪽에 도식에 묘사 된) 특정 뇌관 쌍을 사용 하 여 증폭의 예를 보여주는. 안녕하세요 C 결 찰 제품 (PCR 제품 A에 표시 된)의 HindIII, NheI 및 HindIII/NheI 제한 다이제스트의 ((B)) 왼쪽, 대표적인 예. 바로, DNA의 설계도 C 결 찰 실패 다음 (위)에 후 시퀀스 또는 제한 접합 및 이후 결 찰의 성공 (아래) dNTP Klenow 채우기. (C) 대표가 C 라이브러리 bioanalyzer 프로 파일 (1/5 희석). (D) 대표 PCHi C 라이브러리 bioanalyzer 프로 파일 (희석). 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 3 : PCHi C 후 시퀀싱 품질 컨트롤. (A) 비율 유효한 시퀀스의 비교 HiCUP43 PCHi C vs이 C 라이브러리 (하비에르 외에서 데이터, 201635) 해당 처리 후 쌍을 읽기. (B) 대표 HiCUP PCHi-C 결과 유효한 보여주는 쌍, 및 다운스트림 분석 (하비에르 외에서 데이터, 201635) 전에 삭제는 다른 시퀀스 범주 읽기. (C) 비율 '같은 조각 내부'의 비교가 읽습니다 HiCUP 후 PCHi C vs이 C 라이브러리 (하비에르 외에서 데이터, 201635) 해당 처리. (D) 비율 시퀀스의 비교 PCHi C vs이 C 라이브러리 (하비에르 외., 201635에서 데이터)에 해당 관련 된 baited 발기인 조각 (캡처 효율)를 읽습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 4: ALAD 인간 조 혈 모 세포 프로 파일 PCHi C. 골수성 세포 유형의 상호 작용 발기인 블루 아치로 표시 되 고 림프 세포 유형의 상호 작용 발기인 보라색 아치로 표시 됩니다. Erythroblast-특정 상호 작용 (하비에르 외에서 데이터, 201635) 빨간색 화살표로 표시 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 5 : IL8 인간 조 혈 모 세포 프로 파일 PCHi C. 골수성 세포 유형의 상호 작용 발기인 블루 아치로 표시 되 고 림프 세포 유형의 상호 작용 발기인 보라색 아치로 표시 됩니다. Monocyte 특정 상호 작용 녹색 화살표로 표시 됩니다, neutrophil 특정 상호 작용 빨간색 화살표로 표시 됩니다 그리고 megakaryocyte 특정 상호 작용을 (하비에르 외에서 데이터, 201635) 갈색 화살표로 표시 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}
| 인간의 | ||||||||
| 이름 | 시퀀스 | 염색체 | 스트랜드 | 시작 GRCh38/hg38 | 끝 GRCh38/hg38 | 3 C 상호 작용 및 biotin 합동 테스트 뇌관 조합 | ||
| hs AHF64 데커 | GCATGCATTAGCCTCTGCTGTTCTCTGAAATC | 11 | + | 116803960 | 116803991 | hs AHF66 데커와 함께 사용 | ||
| hs AHF66 데커 | CTGTCCAAGTACATTCCTGTTCACAAACCC | 11 | + | 116810219 | 116810248 | hs AHF64 데커와 함께 사용 | ||
| hs MYC 로커 스 | GGAGAACCGGTAATGGCAAA | 8 | - | 127733814 | 127733833 | hs MYC +1820와 함께 사용 하 여 hs MYC-538 | ||
| hs MYC +1820 | AAAATGCCCATTTCCTTCTCC | 8 | + | 129554527 | 129554547 | hs MYC 로커 스와 함께 사용 | ||
| hs MYC-538 | TGCCTGATGGATAGTGCTTTC | 8 | - | 127195696 | 127195716 | hs MYC 로커 스와 함께 사용 | ||
| hs HIST1 F | AAGCAGGAAAAGGCATAGCA | 6 | + | 26207174 | 26207193 | hs HIST1 R와 함께 사용 | ||
| hs HIST1 R | TCTTGGGTTGTGGGACTTTC | 6 | + | 27771575 | 27771594 | hs HIST1 F와 함께 사용 | ||
| 마우스 | ||||||||
| 시퀀스 | 염색체 | 스트랜드 | 시작 GRCm38/mm10 | 끝 GRCm38/mm10 | 3 C 상호 작용 및 biotin 합동 테스트 뇌관 조합 | |||
| TCATGAGTTCCCCACATCTTTG | 8 | + | 84841090 | 84841111 | mm Calr2와 함께 사용 | |||
| CTGTGGGCACCAGATGTGTAAAT | 8 | + | 84848519 | 84848541 | mm Calr1와 함께 사용 | |||
| TATCAAGGGTGCCCGTCACCTTCAGC | 6 | + | 125163098 | 125163123 | Gapdh4 데커와 함께 사용 | |||
| GGGCTTTTATAGCACGGTTATAAAGT | 6 | + | 125163774 | 125163799 | Gapdh3 데커와 함께 사용 | |||
| GGAGGAGGGAAAAGGAGTGATT | 6 | + | 52212829 | 52212850 | mm Hoxa13와 함께 사용 | |||
| CAGGCATTATTTGCTGAGAACG | 6 | - | 52253490 | 52253511 | mm Hoxa7와 함께 사용 | |||
| GGGTAATGGTGTCACTAACTGG | 13 | + | 23571284 | 23571305 | m Hist1h3e m 또는 mm Hist1h4i와 함께 사용 | |||
| GGGTTTGATGAGTTGGTGAAG | 13 | + | 23566541 | 23566561 | mm Hist1h2ae와 함께 사용 | |||
| TTGGGCCAAAGCCTATATGA | 13 | + | 22043085 | 22043104 | mm Hist1h2ae와 함께 사용 | |||
표 1: 인간의 품질 관리에 대 한 입문 시퀀스 및 마우스가 C 라이브러리.
토론
모듈형 디자인의 발기인 캡처 안녕-C
발기인 캡처 안녕-C는 구체적으로 발기인을 포함 하는 상호 작용에 대 한이 C 라이브러리를 풍부 하 게 설계 되었습니다. 이러한 상호 작용만이 C 라이브러리에 결 찰 제품의 하위 집합으로 구성 됩니다.
캡처 안녕-C 쉽게 캡처 시스템을 변경 하 여 모든 게놈 지역 또는 관심 영역에 대 한이 C 라이브러리를 풍부 하 게 수정할 수 있습니다. 캡쳐 영역 연속 게놈 세그먼트44,45,,4648, 강화 된 수 나 지나치게 사이트49 PCHi-C (' 리버스 캡처 안녕-C'35), 또는 DNase에서 확인 . 실험 범위에 따라 캡처 시스템의 크기를 조정할 수 있습니다. 예를 들어 드 라이덴은 외. 유 방 암44와 관련 된 3 개의 유전자 사막에서 519 미끼 조각을 대상. 마틴 외캡처 시스템. 두 연속 게놈 세그먼트를 대상 (' 지역 캡처 ': 총에서 211 게놈 영역; 2,131 제한 조각) 발기인 (3,857 유전자 발기인)45를 선택.
SureSelect 라이브러리는 다양 한 크기 범위에서 사용할 수: 499 kb (5190-4,806) 2.9 메가바이트 (5190-4,816), 500 킬로바이트 1 kb와 3 5.9 mb (5190-4,831). 각 개별 캡처 biotin RNA 120 뉴클레오티드 긴으로, 이러한 캡처 시스템 4,158의 최대 수용, 24,166 및 49,166 개별 캡처 프로브, 각각. 이 각각 2,079, 12,083, 및 24,583 대상된 제한 조각에 해당 (참고 제한 조각에 대 한 숫자는 경계값 모든 제한에 대 한 두 개의 개별 캡처 프로브를 디자인 될 수 있다 가정에 따라 조각-반복 시퀀스 인해 현실에서이 되지 것입니다 모든 제한 경우 조각 ( 그림 1B, C참조), 대상 제한 조각 일정 수가 사용 가능한 캡처 프로브에 대 한 높은 수 인 ).
여기에 설명 된 프로토콜을 장거리 상호 작용을 밝히기 위해서 6 bp 인식 사이트와 제한 효소의 사용을 기반으로 합니다. 4 혈압 인식 사이트와 더 인접 상호 작용의 높은 해상도 대 한 제한 효소를 사용 하 여 가능한40,49또한 이다.
PCHi C의 한계
모든 염색체 구조 붙 잡음 분석 실험의 한 제한을입니다 해상도 라이브러리 생성을 위해 사용 하는 제한 효소에 의해 결정 됩니다. 동일한 제한 조각에 있는 DNA 요소 간에 발생 하는 상호 작용 ' C 형 ' 분석에 표시 되지 않습니다. 또한, PCHi-c, 어떤 경우에 더 이상의 전사 시작 사이트 동일한 발기인에 포함 된 제한 부분에 있을 수 있습니다 및 일부 경우에서 Pir 항구 모두 적극적이 고 억압 적인 히스톤 부호 어렵게 그것을 정확 하 게 규정 요소는 상호 작용을 중재 발기인 상호 작용의 규정 출력을 예측 하 고. 4 혈압 인식 사이트와 제한 효소를 사용 하 여이 문제를 완화 하지만 대폭 증가 C 라이브러리 복잡성의 비용 (4 혈압 인식 사이트 제한 효소 생성이 C 라이브러리는이 C 보다 적어도 100 배는 더 복잡 한 6 혈압 인식 사이트 제한 효소를 사용 하 여 생성 하는 라이브러리의 경우) 다음 세대 시퀀싱 관련 비용.
또 다른 한계는 현재 PCHi C 프로토콜 자료, 희귀 종류 상호 작용 발기인의 분석 제외를 시작으로 세포의 수백만 요구. 100000 10000 셀 (예를 들어 초기 배아 개발 또는 조 혈 줄기 세포)와 세포 인구에서 발기인 연락처의 심문 수 있도록 PCHi C의 수정된 된 버전 캡처에 귀중 한 추가 될 것 이다 안녕하세요 C 도구 상자입니다.
마지막으로, 포름알데히드 고정에 사용 하는 방법에 대 한 모든, 같은 PCHi C만 기록 '냉동' 상호 작용의 고정 시간 시점에서. 따라서, 공부 하 고 활동 및 발기인 상호 작용의 역동성, 슈퍼 해상도 라이브 셀 현미경 같은 방법 필요 PCHi C와 함께
높은 해상도에서 공간 염색체 조직 해 부 방법
염색체 상호 작용 라이브러리의 광대 한 복잡 통계적 의미와 두 가지 특정 제한 조각 사이 상호 작용 제품의 신뢰할 수 있는 식별을 금지합니다. 이 문제를 회피, 시퀀스 캡처 안녕 C33,34,,4044 또는 특정 상호 작용에 대 한 3 C50,51 라이브러리를 풍부 하 게 사용 되었습니다. 농축 단계에 대 한이 C 라이브러리 이상 3 C 라이브러리를 사용 하 여의 주요 장점은 그이-C, 3 C, 달리 포함 정품 결 찰 제품에 대 한 농축 단계입니다. 결과적으로, PCHi C 라이브러리에서 유효한 읽기 비율 약 10 배 보다 높습니다 캡처-c에서 라이브러리50을 포함 약 5-8% 유효한 HiCUP 필터링 후 읽습니다. Sahlen 외. 직접 HiCap PCHi c 3 C 라이브러리를 사용 하 여 캡처-C 달리 캡처 농축이 C 라이브러리를 사용 하는 캡처-C를 비교 했습니다. 우리의 결과와 일치, 그들은 캡처 C 라이브러리는 주로 유엔 합자 조각40이루어진 발견. 또한, HiCap 라이브러리 캡처 C 라이브러리40보다 높은 복잡을 했다.
캡처-C, 다음-세대 캡처 C52 (NG 캡처-C) 라는 변종 PCHi C33,34, 겹치는 원본에 사용 되는 프로브 대신에 이전 설립 제한 조각 끝 당 하나 올리고 사용 캡처-C 프로토콜50. 이 겸손, 캡처-C를 비교 하는 유효한 읽기의 비율을 증가 하지만 NG 캡처-C 사용 하 여 캡처 농축의 두 순차 라운드 및 주기 PCR의 상대적으로 높은 수 (총, 20 ~ 24 사이클에 비해 11 주기 일반적으로 PCHi-c)는 필연적으로 결과 시퀀스 중복 및 낮은 라이브러리 복잡성의 높은 숫자에. 그러나 우리 고유 (즉, 중복)의 비율 쌍만 약 15% 이었다 19 PCR 주기 사용 했을 때 읽기 발견 PCHi C의 최적화 하는 동안 평가 실험, (13 사이클 미리 캡처 + 6 주기 후 캡처; 데이터 표시 되지 않음), PCR 주기, 낮은 수를 최적화 일반적으로 75-90% 고유 읽기 쌍 생성 합니다. 따라서, 실질적으로 PCR 사이클의 수를 줄이고 유익한 시퀀스 데이터 양을 증가 합니다.
최근 방법 안녕-C (HiChIP53)의 특정 단백질에 의해 중재 염색체 상호 작용에 초점을 가진 칩을 결합 합니다. ChIA 애완 동물54, 유사한 이론적 기반에 비해 HiChIP 데이터 정보 순서 읽기,53를 호출 하는 높은 신뢰 상호 작용에 대 한 허용 더 높은 수가 포함 되어 있습니다. 그것은 매우 직접 해당 HiChIP를 비교 하는 흥미로운 되며 안녕-C를 캡처 데이터 세트 한 번 그들이 사용할 수 있게 (예: HiChIP cohesin 단위 Smc1a에 대 한 항 체를 사용 하 여53 와 캡처 안녕-C 모든 Smc1a에 대 한 바인딩 제한 파편) 나란히입니다. 이러한 두 방법의 고유의 차이입니다 캡처 안녕-C chromatin immunoprecipitation에 의존 하지 않는 및 따라서 염색체 상호 작용 단백질 인에 관계 없이 질문은. 특정 요소 바인딩, PRC1 마우스 ESC 공간 게놈 건축7의 키 레 귤 레이 터로 식별 하는 데 사용 되었습니다가의 유무에서 3D 게놈 조직의 비교 수 있습니다.
PCHi C와 GWAS
그 보다 큰 95%의 질병-관련 된 게놈 넓은 협회 연구 결과 (GWAS) 밝혀졌다 시퀀스 변종 단백질 코딩 유전자55먼 거리에서 종종 게놈의 비 코딩 영역에 있습니다. GWAS 변종 들은 근접 DNase 나 지나치게 사이트 발견, 시퀀스를 잠재적인 규제 활동의 특징은. PCHi C와 캡처 안녕-C 링크 발기인 GWAS 위험 loci 유 방 암44,48대 장 암 및 자가 면역 질환35,,4546에 연루를 광범위 하 게 사용 되어 왔습니다. PCHi C 종류 발견 자가 면역 질환과 관련 된 Snp 림프 세포에서 Pir에 농축 된 시퀀스 변종 혈소판 및 적혈구 특정 특색과 관련 된 했다에서 주로 발견 되지만 17 다른 인간의 조 혈 세포에 관한 연구 대 식 세포 그리고 erythroblasts, 각각35,56. 따라서, PCHi-C에 의해 발견 하는 interactomes 비 코딩 질병 관련의 기능을 이해를 도움이 될 수 하는 조직 형 특정 발기인 이체 순서 하 고 치료 적 개입에 대 한 새로운 잠재적인 질병 유전자를 식별 합니다.
발기인 상호 작용 영역의 특성
여러 줄 증거의 발기인 interactomes 유전자 식 제어 링크. 첫째, 여러 PCHi C 학문은 게놈 영역 (높은) 표현한 유전자의 발기인 상호 작용 H3K27 acetylation 등 p300 바인딩33,34 증강 활동과 관련 된 표시에 농축은 설명 했다 , 37. 우리 진 식 수준 상호 작용 강화, 강화 결과 증가 유전자 발현에서의 첨가제 효과34,35레벨 제안 수 사이 긍정적인 상관 관계를 발견. 둘째, 자연스럽 게 식 양적 특성 loci (eQTLs) 누구의 식 eQTLs35에 의해 영향을 같은 유전자에 연결 되어 있는 Pir에서 풍성 하 게 발생 합니다. 셋째, 여행57 및 PCHi C 데이터 통합, 케언즈 연구진이 발견 그 여행 리포터 유전자 마우스 ESCs에서 Pir를 보여 강한 기자 통합 사이트에서 리포터 유전자 보다 유전자 발현 발기인 상호 작용 영역에서 58, Pir transcriptional 규제 활동 소유를 나타내는. 함께, 이러한 결과 다양 한 마우스 및 인간의 세포 유형 PCHi-C에 의해 발견 하는 발기인 interactomes 유전자 식 컨트롤에 대 한 주요 규제 모듈 포함 것이 좋습니다.
그것은 PCHi C33,34에 의해 발견 하는 모든 Pir의 강화만 작은 분수 (~ 20%)을 나타내는 지적 가치가 있다. 다른 Pir 직접 transcriptional 규정 하는 기능 보다는 오히려 구조 또는 토폴로지 역할 있다. 그러나, 증거가 PCHi C 클래식 증강 마크 항구 하지 않는 규제 기능을 가진 DNA 요소를 밝히기 수 있습니다 또한 있다. 인간 림프 세포 라인에서 BRD7 발기인 보유 취재 원 유전자 분석 실험33증강 활동 표시 했다 증강 마크 없는 지역 상호 작용 발견 됐다. 유사한 특성을 가진 규제 요소는 현재 감사 보다 더 풍부한 있을 수 있습니다. 예를 들어 CRISPR 기반 화면 요소에 대 한 규제 DNA 요소 식별 표시 규제 (UREs) 유전자 발현을 제어 하지만 증강 없는59를표시 합니다.
다른 경우에, Pir chromatin 마크 transcriptional 억제와 관련 된 항구 표시 되었습니다. Pir 및 상호 작용 발기인 PRC1 마우스 ESCs에 의해 구속에 종사 했다 억압된 유전자 베어링의 광범위 한 공간 네트워크는 억압 적인 표시 H3K27me37. 인간의 lymphoblastoid 세포에서 BCL6 발기인 상호 작용 요소를 먼 transgene 기자 진 식33, 네이티브 컨텍스트에서 BCL6 전사를 억 누르기 위해 작동 수 있습니다 제안을 억 눌렀다.
Pir 농축 대 인 인간의 ESCs와 자문위37 chromatin 절연체 단백질 CTCF의 Pir의 또 다른 클래스를 나타낼 수 있습니다. 샨 다, 이러한 결과 Pir 유전자 기능 특징을 아직 규제 활동의 컬렉션 항구 것이 좋습니다.
공개
저자는 공개 없다.
감사의 말
우리는 원고 및 그림 1 전문가 도움의 중요 한 독서에 대 한 Valeriya Malysheva 감사합니다. 이 작품은 의료 연구 위원회, 영국 (미스터/L007150/1)와 영국 생명 공학 및 생물 과학 연구 협의회, 영국 (BB/J004480/1)에 의해 지원 되었다.
자료
| Name | Company | Catalog Number | Comments |
| 16% (vol/vol) paraformaldehyde solution | Agar Scientific | R1026 | |
| Dulbecco's Modified Eagle Medium (DMEM) 1x | Life Technologies | 41965-039 | |
| Fetal bovine serum (FBS) sterile filtered | Sigma | F9665 | |
| Low-retention filter tips | Starlab | S1180-3810, S1180-1810, S1180-8810 and S1182-1830 | |
| 10x PBS pH 7.4 | Life Technologies | 70011-036 | |
| Molecular biology grade water | Sigma-Aldrich | W4502 | |
| 1 M Tris-HCl pH 8.0 | Life Technologies | 15568-025 | |
| IGEPAL CA-630 | Sigma-Aldrich | I8896 | |
| 5 M NaCl | Life Technologies | 24740-011 | |
| Protease inhibitor cocktail (EDTA-free) | Roche Diagnostics | 11873580001 | |
| Restriction buffer 2 (10x NEBuffer 2) | New England Biolabs | B7002 | |
| DNA LoBind tube, 1.5 mL | Eppendorf | 0030 108.051 | |
| DNA LoBind tube, 2 mL | Eppendorf | 30108078 | |
| 20% (wt/vol) SDS | Bio-Rad Laboratories | 161-0418 | |
| 20% (vol/vol) Triton X-100 | Sigma-Aldrich | T8787 | |
| HindIII, 100 U/uL | New England Biolabs | R0104 | |
| 10 mM dCTP | Life Technologies | 18253-013 | |
| 10 mM dGTP | Life Technologies | 18254-011 | |
| 10 mM dTTP | Life Technologies | 18255-018 | |
| 0.4 mM Biotin-14-dATP | Life Technologies | 19524-016 | |
| DNA polymerase I large (Klenow) fragment 5000 units/mL | New England Biolabs | M0210 | |
| 10x T4 DNA ligase reaction buffer | New England Biolabs | B0202 | |
| 100x 10mg/ml Bovine Serum Albumin | New England Biolabs | B9001 | |
| T4 DNA ligase, 1 U/μL | Invitrogen | 15224-025 | |
| RNase A | Roche | 10109142001 | |
| Proteinase K, recombinant, PCR grade | Roche | 3115836001 | |
| 20 000×g 50 ml centrifuge tube | VWR | 525-0156 | |
| 0.5 M EDTA pH 8.0 | Life Technologies | 15575-020 | |
| Phenol pH 8.0 | Sigma | P4557 | |
| Phenol: Chloroform: Isoamyl Alcohol 25:24:1 | Sigma | P3803 | |
| Sodium acetate pH 5.2 | Sigma | S7899 | |
| Quant-iT PicoGreen | Invitrogen | P7589 | |
| QIAquick Gel Extraction Kit | Qiagen | 28704 | |
| QIAquick PCR Purification Kit | Qiagen | 28104 | |
| Restriction buffer 2.1 (10x NEBuffer 2.1) | New England Biolabs | B7202 | |
| NheI, 100U/uL | New England Biolabs | R0131 | |
| Micro TUBE AFA Fiber Pre-slit snap cap 6x16mm vials | Covaris | 520045 | For sonication |
| SPRI beads (Agencourt AMPure XP) | Beckman Coulter | A63881 | |
| Dynabeads MyOne Streptavidin C1 beads | Invitrogen | 65001 | |
| Tween 20 | Sigma | P9416 | |
| 10 mM dATP | Life Technologies | 18252-015 | |
| T4 DNA polymerase 3000 units/mL | New England Biolabs | M0203 | |
| T4 PNK 10000 units/mL | New England Biolabs | M0201 | |
| Klenow exo minus 5000 units/mL | New England Biolabs | M0212 | |
| Quick ligation reaction buffer | New England Biolabs | B6058 | |
| NEB DNA Quick ligase | New England Biolabs | M2200 | |
| PE adapter 1.0 (5'-P-GATCGGAAGAGCGGTTCAGC AGGAATGCCGAG-3') | Illumina | ||
| PE adapter 2.0 (5'-ACACTCTTTCCCTACACGACGCT CTTCCGATCT-3') | Illumina | ||
| NEB Phusion PCR kit | New England Biolabs | M0530 | |
| PE PCR primer 1.0 (5'-AATGATACGGCGACCACCGA GATCTACACTCTTTCCCTAC ACGACGCTCTTCCGATCT-3') | Illumina | ||
| PE PCR primer 2.0 (5'-CAAGCAGAAGACGGCATACGA GATCGGTCTCGGCATTCCT GCTGAACCGCTCTTCCGATCT-3') | Illumina | ||
| PCR strips | Agilent Technologies | 410022 and 401425 | |
| SureSelect SSEL TE Reagent ILM PE full adaptor kit | Agilent Technologies | 931108 | |
| SureSelect custom 3-5.9 Mb library | Agilent Technologies | 5190-4831 | custom design mouse or human PCHi-C system |
| Dynabeads MyOne Streptavidin T1 beads | Invitrogen | 65601 | |
| E220 high-performance focused ultra-sonicator | Corvaris | E220 |
참고문헌
- Osborne, C. S., et al. Active genes dynamically colocalize to shared sites of ongoing transcription. Nature Genetics. 36, 1065-1071 (2004).
- Schoenfelder, S., et al. Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells. Nature Genetics. 42, 53-61 (2010).
- de Wit, E., et al. The pluripotent genome in three dimensions is shaped around pluripotency factors. Nature. 501, 227-231 (2013).
- Bantignies, F., et al. Polycomb-dependent regulatory contacts between distant Hox loci in Drosophila. Cell. 144, 214-226 (2011).
- Engreitz, J. M., et al. The Xist lncRNA exploits three-dimensional genome architecture to spread across the X chromosome. Science. 341, 1237973(2013).
- Denholtz, M., et al. Long-range chromatin contacts in embryonic stem cells reveal a role for pluripotency factors and polycomb proteins in genome organization. Cell Stem Cell. 13, 602-616 (2013).
- Schoenfelder, S., et al. Polycomb repressive complex PRC1 spatially constrains the mouse embryonic stem cell genome. Nature Genetics. 47, 1179-1186 (2015).
- Kundu, S., et al. Polycomb Repressive Complex 1 generates discrete compacted domains that change during differentiation. Molecular Cell. 65, 432-446 (2017).
- Skok, J. A., Gisler, R., Novatchkova, M., Farmer, D., de Laat, W., Busslinger, M. Reversible contraction by looping of the Tcra and Tcrb loci in rearranging thymocytes. Nature Immunology. 8, 378-387 (2007).
- Zhang, Y., et al. Spatial organization of the mouse genome and its role in recurrent chromosomal translocations. Cell. 148, 908-921 (2012).
- Aymard, F., et al. Genome-wide mapping of long-range contacts unveils clustering of DNA double-strand breaks at damaged active genes. Nature Structural & Molecular Biology. 24, 353-361 (2017).
- Ryba, T., et al. Evolutionarily conserved replication timing profiles predict long-range chromatin interactions and distinguish closely related cell types. Genome Research. 20, 761-770 (2010).
- Pope, B. D., et al. Topologically associating domains are stable units of replication-timing regulation. Nature. 515, 402-405 (2014).
- Chandra, T., et al. Global reorganization of the nuclear landscape in senescent cells. Cell Reports. 10, 471-483 (2015).
- Carter, D., Chakalova, L., Osborne, C. S., Dai, Y. F., Fraser, P. Long-range chromatin regulatory interactions in vivo. Nature Genetics. 32, 623-626 (2002).
- Tolhuis, B., Palstra, R. J., Splinter, E., Grosveld, F., de Laat, W. Looping and interaction between hypersensitive sites in the active beta-globin locus. Molecular Cell. 10, 1453-1465 (2002).
- Amano, T., Sagai, T., Tanabe, H., Mizushina, Y., Nakazawa, H., Shiroishi, T. Chromosomal dynamics at the Shh locus: limb bud-specific differential regulation of competence and active transcription. Developmental Cell. 16, 47-57 (2009).
- Zuniga, A., et al. Mouse limb deformity mutations disrupt a global control region within the large regulatory landscape required for Gremlin expression. Genes & Development. 18, 1553-1564 (2004).
- Sagai, T., Hosoya, M., Mizushina, Y., Tamura, M., Shiroishi, T. Elimination of a long-range cis-regulatory module causes complete loss of limb-specific Shh expression and truncation of the mouse limb. Development. 132, 797-803 (2005).
- D'Haene, B., et al. Disease-causing 7.4 kb cis-regulatory deletion disrupting conserved non-coding sequences and their interaction with the FOXL2 promotor: implications for mutation screening. PLoS Genet. 5, e1000522(2009).
- Sur, I. K., et al. Mice lacking a Myc enhancer that includes human SNP rs6983267 are resistant to intestinal tumors. Science. 338, 1360-1363 (2012).
- Herranz, D., et al. A NOTCH1-driven MYC enhancer promotes T cell development, transformation and acute lymphoblastic leukemia. Nature Medicine. 20, 1130-1137 (2014).
- Deng, W., et al. Controlling long-range genomic interactions at a native locus by targeted tethering of a looping factor. Cell. 149, 1233-1244 (2012).
- Groschel, S., et al. A single oncogenic enhancer rearrangement causes concomitant EVI1 and GATA2 deregulation in leukemia. Cell. 157, 369-381 (2014).
- Lupianez, D. G., et al. Disruptions of topological chromatin domains cause pathogenic rewiring of gene-enhancer interactions. Cell. 161, 1012-1025 (2015).
- Franke, M., et al. Formation of new chromatin domains determines pathogenicity of genomic duplications. Nature. 538, 265-269 (2016).
- Dekker, J., Rippe, K., Dekker, M., Kleckner, N. Capturing chromosome conformation. Science. 295, 1306-1311 (2002).
- Simonis, M., et al. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C). Nature Genetics. 38, 1348-1354 (2006).
- Zhao, Z., et al. Circular chromosome conformation capture (4C) uncovers extensive networks of epigenetically regulated intra- and interchromosomal interactions. Nature Genetics. 38, 1341-1347 (2006).
- Dostie, J., et al. Chromosome Conformation Capture Carbon Copy (5C): A massively parallel solution for mapping interactions between genomic elements. Genome Research. 16, 1299-1309 (2006).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326, 289-293 (2009).
- Belton, J. M., McCord, R. P., Gibcus, J. H., Naumova, N., Zhan, Y., Dekker, J. Hi-C: a comprehensive technique to capture the conformation of genomes. Methods. 58, 268-276 (2012).
- Mifsud, B., et al. Mapping long-range promoter contacts in human cells with high-resolution capture Hi-C. Nature Genetics. 47, 598-606 (2015).
- Schoenfelder, S., et al. The pluripotent regulatory circuitry connecting promoters to their long-range interacting elements. Genome Res. 25, 582-597 (2015).
- Javierre, B. M., et al. Lineage-specific genome architecture links enhancers and non-coding disease variants to target gene promoters. Cell. 167, 1369-1384 (2016).
- Wilson, N. K., et al. Integrated genome-scale analysis of the transcriptional regulatory landscape in a blood stem/progenitor cell model. Blood. 127, e12-e23 (2016).
- Freire-Pritchett, P., et al. Global reorganisation of cis-regulatory units upon lineage commitment of human embryonic stem cells. Elife. 6, (2017).
- Rubin, A. J., et al. Lineage-specific dynamic and pre-established enhancer-promoter contacts cooperate in terminal differentiation. Nature Genetics. 49, 1522-1528 (2017).
- Siersbaek, R., et al. Dynamic rewiring of promoter-anchored chromatin loops during adipocyte differentiation. Molecular Cell. 66, 420-435 (2017).
- Sahlen, P., et al. Genome-wide mapping of promoter-anchored interactions with close to single-enhancer resolution. Genome Biology. 16, 156(2015).
- Nagano, T., et al. Single-cell Hi-C reveals cell-to-cell variability in chromosome structure. Nature. 502, 59-64 (2013).
- Nagano, T., Varnai, C., Schoenfelder, S., Javierre, B. M., Wingett, S. W., Fraser, P. Comparison of Hi-C results using in-solution versus in-nucleus ligation. Genome Biology. 16, 175(2015).
- Wingett, S., et al. HiCUP: pipeline for mapping and processing Hi-C data. F1000 Res. 4, 1310(2015).
- Dryden, N. H., et al. Unbiased analysis of potential targets of breast cancer susceptibility loci by Capture Hi-C. Genome Research. 24, 1854-1868 (2014).
- Martin, P., et al. Capture Hi-C reveals novel candidate genes and complex long-range interactions with related autoimmune risk loci. Nature Communications. 6, 10069(2015).
- McGovern, A., et al. Capture Hi-C identifies a novel causal gene, IL20RA, in the pan-autoimmune genetic susceptibility region 6q23. Genome Biol.ogy. 17, 212(2016).
- Hodge, D., et al. A global role for EKLF in definitive and primitive erythropoiesis. Blood. 107, 3359-3370 (2006).
- Jager, R., et al. Capture Hi-C identifies the chromatin interactome of colorectal cancer risk loci. Nature Communications. 6, 6178(2015).
- Joshi, O., et al. Dynamic reorganization of extremely long-range promoter-promoter Interactions between two states of pluripotency. Cell Stem Cell. 17, 748-757 (2015).
- Hughes, J. R., et al. Analysis of hundreds of cis-regulatory landscapes at high resolution in a single, high-throughput experiment. Nature Genetics. 46, 205-212 (2014).
- Kolovos, P., et al. Targeted Chromatin Capture (T2C): A novel high-resolution high-throughput method to detect genomic interactions and regulatory elements. Epigenetics Chromatin. 7, 10(2014).
- Davies, J. O., et al. Multiplexed analysis of chromosome conformation at vastly improved sensitivity. Nature Methods. 13, 74-80 (2016).
- Mumbach, M. R., et al. HiChIP: efficient and sensitive analysis of protein-directed genome architecture. Nature Methods. 13, 919-922 (2016).
- Fullwood, M. J., et al. An oestrogen-receptor-alpha-bound human chromatin interactome. Nature. 462, 58-64 (2009).
- Maurano, M. T., et al. Systematic localization of common disease-associated variation in regulatory DNA. Science. 337, 1190-1195 (2012).
- Petersen, R., et al. Platelet function is modified by common sequence variation in megakaryocyte super enhancers. Nat. Commun. 8, 16058(2017).
- Akhtar, W., et al. Chromatin position effects assayed by thousands of reporters integrated in parallel. Cell. 154, 914-927 (2013).
- Cairns, J., et al. CHiCAGO: Robust detection of DNA looping interactions in Capture Hi-C data. Genome Biology. 17, 127(2016).
- Rajagopal, N., et al. High-throughput mapping of regulatory DNA. Nature Biotechnology. 34, 167-174 (2016).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유