
Method Article
Microscopia crioelettronica a particella singola: dal campione alla struttura
In questo articolo
Riepilogo
La determinazione della struttura dei complessi macromolecolari utilizzando il crioEM è diventata di routine per alcune classi di proteine e complessi. Qui, questa pipeline è riassunta (preparazione del campione, screening, acquisizione ed elaborazione dei dati) e i lettori sono indirizzati verso ulteriori risorse dettagliate e variabili che possono essere modificate nel caso di campioni più impegnativi.
Abstract
La crioscopia elettronica (cryoEM) è una potente tecnica per la determinazione della struttura di complessi macromolecolari, tramite analisi di singole particelle (SPA). Il processo complessivo prevede i) la vetrificazione del campione in un film sottile supportato su una griglia crioEM; ii) screening del campione per valutare la distribuzione delle particelle e la qualità del ghiaccio; iii) se la griglia è idonea, raccogliendo un singolo dataset di particelle per l'analisi; e iv) elaborazione delle immagini per produrre una mappa di densità EM. In questo protocollo viene fornita una panoramica per ciascuno di questi passaggi, con particolare attenzione alle variabili che un utente può modificare durante il flusso di lavoro e alla risoluzione dei problemi comuni. Con il funzionamento del microscopio remoto che sta diventando standard in molte strutture, verranno descritte le variazioni sui protocolli di imaging per assistere gli utenti in un funzionamento efficiente e l'imaging quando l'accesso fisico al microscopio è limitato.
Introduzione
CryoEM a singola particella
Per indagare la vita a livello molecolare dobbiamo capire la struttura. Sono disponibili molte tecniche per sondare la struttura delle proteine, come NMR, cristallografia a raggi X, spettrometria di massa e microscopia elettronica (EM). Ad oggi, la maggior parte delle strutture depositate nella Protein Databank (PDB) sono state risolte utilizzando la cristallografia a raggi X. Tuttavia, dal ~ 2012 in poi, la microscopia crioelettronica (cryoEM) è diventata una tecnica tradizionale per la determinazione della struttura proteica e il suo uso è aumentato drasticamente. Il numero totale di mappe EM depositate presso la Banca dati di microscopia elettronica (EMDB) (a dicembre 2020) era di 13.421 rispetto alle 1.566 del 2012 (Figura 1, www.ebi.co.uk). Nel 2012 il numero di coordinate atomiche modellate nelle mappe di densità crioEM, depositate nel PDB era solo 67 ma a dicembre 2020 sono state depositate finora 2.309 strutture, un aumento di 35 volte. Questa crescita di fondo della qualità e della quantità delle mappe di densità crioEM prodotte, a volte indicata come la "rivoluzione della risoluzione"1, è stata causata da una coalescenza di progressi in più aree: lo sviluppo di nuove telecamere per l'imaging note come rivelatori di elettroni diretti; nuovo software; e microscopi più stabili2,3,4.

Figura 1: Invii cumulativi all'EMDB dal 2012 a dicembre 2020. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
L'analisi a singola particella (SPA) è un potente strumento per generare informazioni biologiche in un'ampia varietà di tipi di campioni chiarendo strutture ad alta risoluzione di complessi isolati5,6 tra cui virus7,8, proteine di membrana9,10, assemblaggi elicoidali11 e altri complessi macromolecolari dinamici ed eterogenei12,13, le cui dimensioni variano in ordine di grandezza (da 39 kDa Da 14,15 a decine di megadaltoni). Qui viene descritto un protocollo per una pipeline standard per cryoEM SPA dal campione alla struttura.
Prima di intraprendere questa pipeline, un campione purificato dovrebbe essere sottoposto ad analisi biochimiche per valutarne le possibilità di successo a valle. La preparazione di un campione adatto è probabilmente la più grande barriera alla SPA, in particolare per i complessi transitori ed eterogenei (sia compositivi che conformazionali). La preparazione del complesso macromolecolare dovrebbe contenere il minor numero possibile di contaminanti, a concentrazione sufficiente per produrre molte particelle in ogni micrografia crioEM e in una composizione tampone adatta all'analisi crioEM. Alcuni costituenti tampone, tra cui saccarosio, glicerolo e alte concentrazioni di sali (~> 350 mM, a seconda delle dimensioni del campione, delle proprietà e di altri componenti tampone) possono interferire con il processo di vitrificazione o ridurre il rapporto segnale-rumore nelle immagini, ostacolando la determinazione della struttura16.
Tipicamente, come minimo, la cromatografia di esclusione dimensionale (SEC) e l'analisi del gel SDS-PAGE dovrebbero essere utilizzate per valutare la purezza del campione17,18, ma il dicroismo circolare, i saggi funzionali, la SEC accoppiata con la diffusione della luce multi-angolo e i saggi di stabilità termica sono tutti strumenti utili per l'analisi qualitativa dei preparati complessi macromolecolari prima dell'analisi crioEM. Tuttavia, i risultati di queste analisi biochimiche possono fornire poche informazioni sull'eterogeneità strutturale del campione e sul suo comportamento su una griglia crioEM. Per questo motivo, la macchia negativa EM viene abitualmente utilizzata come strumento rapido, economico e potente per valutare l'eterogeneità compositiva e conformazionale, e quindi un buon modo per accertare quale frazione di eluizione da una purificazione è più promettente, o per lo screening di diverse composizioni tampone19,20. Una volta identificato un campione promettente, possiamo procedere alla pipeline CRYOEM SPA. La macchia negativa non sempre si allinea con i risultati successivi osservati in crioEM; a volte un campione sembra povero di macchia negativa ma migliora se visto nel ghiaccio vitreo in crioEM. Al contrario, a volte i campioni sembrano eccellenti durante le fasi di colorazione negativa, ma richiedono un'ulteriore ottimizzazione significativa quando si passa al crioEM. Tuttavia, nella maggior parte dei casi la macchia negativa fornisce un'utile fase di controllo della qualità.
Vitrificazione
L'ambiente ostile all'interno del sistema a vuoto del microscopio elettronico provoca sia disidratazione che danni da radiazioni a campioni biologici non fissati21. Pertanto, per visualizzare il campione in uno stato nativo, il campione biologico deve essere conservato prima dell'imaging. Per i preparati purificati di complessi macromolecolari, la vitrificazione è il metodo di scelta per consentirne la visualizzazione tramite crioEM preservando i dettagli atomici del complesso. La scoperta della vitrificazione come metodo di preparazione del campione è stato un progresso fondamentale nella microscopia elettronica di campioni biologici, per il quale Dubochet è stato riconosciuto nel Premio Nobel per la Chimica 2017. La vitrificazione del campione comporta la creazione di un sottile strato di soluzione contenente il campione di interesse, tipicamente di decine di nm di spessore, sospeso su un supporto a griglia crioEM. Il film sottile viene quindi congelato estremamente rapidamente in un criogeno come l'etano liquido a ~ -175 ° C. La velocità di congelamento è di ~ 106 ° C / s, abbastanza veloce da formarsi ghiaccio amorfo o vitreo, sospendendo il campione in un film sottile e solido22.
La variabile iniziale da considerare è il supporto della griglia cryoEM scelto23. Una griglia EM è tipicamente costituita da un film di carbonio amorfo con perforazioni (regolari o irregolari), su una struttura di supporto. La struttura di supporto è tipicamente una griglia metallica circolare di 3,05 mm di diametro, solitamente realizzata in rame, ma possono essere utilizzati altri metalli come l'oro o il molibdeno (che ha proprietà di espansione termica preferite24). A volte, un ulteriore supporto sottile e continuo viene applicato attraverso la griglia, come grafene, ossido di grafene o uno strato di carbonio amorfo sottile (~ 1-2 nm). Mentre le griglie crioEM standard (più comunemente rame a maglie 400-200 con un supporto in carbonio perforato (fori rotondi da 1,2 μm separati da 1,3 μm (r1,2 / 1,3) o 2 μm separati da 2 μm di carbonio (r2/2)) - sebbene siano disponibili molti modelli diversi) sono state utilizzate nella stragrande maggioranza delle strutture segnalate fino ad oggi, sono state segnalate nuove tecnologie a griglia con conduttività migliorata e ridotto movimento del campione25 . Le griglie selezionate sono sottoposte a un trattamento di pulizia a scarica di bagliore/plasma per renderle idrofile e suscettibili di applicazione su campione26.
Dopo la scarica di bagliore, la fase successiva è la formazione di film sottili. Questo film sottile è più comunemente formato utilizzando carta da filtro per rimuovere il liquido in eccesso dalla griglia. Mentre questo può essere eseguito manualmente, sono disponibili in commercio numerosi dispositivi di congelamento a tuffo, tra cui Vitrobot Mk IV (Thermo Fisher Scientific), EM GP II (Leica) e CP3 (Gatan). Con questi dispositivi, ~ 3-5 μL di campione in soluzione viene applicato alla griglia EM, seguito da un'eliminazione della soluzione in eccesso utilizzando carta da filtro. La griglia, con un film sottile sospeso su di essa, viene quindi immersa in etano liquido raffreddato da azoto liquido (LN2) a ~ -175 ° C. Una volta congelata, la griglia viene mantenuta a una temperatura inferiore al punto di devitrificazione (-137 °C) prima e durante l'imaging.
Screening dei campioni e raccolta dei dati
Dopo la vetrificazione di una griglia crioEM, la fase successiva consiste nello schermare la griglia per valutarne la qualità e determinare se la griglia è adatta per procedere alla raccolta di dati ad alta risoluzione. Una griglia crioEM ideale ha ghiaccio vitreo (opposto al ghiaccio cristallino) con lo spessore del ghiaccio appena sufficiente per ospitare la dimensione più lunga del campione, assicurando che il ghiaccio circostante contribuisca il minor rumore possibile all'immagine risultante. Le particelle all'interno del ghiaccio dovrebbero avere una dimensione e (se nota) forma coerente con la biochimica, e idealmente essere monodisperse con una distribuzione casuale degli orientamenti delle particelle. Infine, la griglia dovrebbe avere abbastanza aree di qualità sufficiente per soddisfare la lunghezza di raccolta dei dati desiderata. A seconda del campione, questo può richiedere molte iterazioni di vetrificazione e screening fino a quando non vengono prodotte griglie ottimali. Sia fortunatamente che sfortunatamente, ci sono una vasta gamma di variabili che possono essere testate empiricamente per alterare la distribuzione delle particelle sulle griglie crioEM (recensito in16,27). In questo manoscritto sono mostrati i risultati rappresentativi per un progetto di proteina di membrana10.
Una volta identificata una griglia adatta, la raccolta dei dati può procedere. Diversi modelli di microscopi elettronici a criotrasmissione per campioni biologici sono ottimizzati per raccogliere dati ad alta risoluzione in modo automatizzato. In genere, i dati vengono raccolti su sistemi a 300 kV o 200 kV. La raccolta automatizzata dei dati può essere ottenuta utilizzando software tra cui EPU (Thermo Fisher Scientific)28, Leginon29, JADAS30 e SerialEM31,32. Una raccolta automatizzata di dati con rilevatori moderni in genere si traduce in terabyte (TB) di dati grezzi in un periodo di 24 ore (i set di dati medi sono ~ 4 TB di dimensione).
A causa delle restrizioni COVID-19 in vigore su gran parte del mondo (momento della scrittura dicembre 2020), molte strutture di microscopia sono passate a offrire l'accesso remoto. Una volta che le griglie sono state caricate nel caricatore automatico di un microscopio, l'acquisizione dei dati può essere condotta da remoto.
Elaborazione delle immagini e costruzione di modelli
Laddove una sessione di raccolta dati può essere in genere di 0,5-4 giorni, l'elaborazione successiva delle immagini può richiedere molte settimane e mesi, a seconda della disponibilità di risorse di elaborazione. È standard per le fasi iniziali di elaborazione delle immagini, vale a dire la correzione del movimento e la stima della funzione di trasferimento del contrasto (CTF) che avvengono "al volo" 33,34. Per l'elaborazione a valle, ci sono una pletora di suite software disponibili. Le particelle vengono "raccolte" ed estratte dalle micrografie35,36. Una volta estratte le particelle, un protocollo standard sarebbe quello di elaborare le particelle attraverso diversi cicli di classificazione (sia in due dimensioni (2D) che in tre dimensioni (3D) e / o focalizzate su specifiche regioni di interesse) per raggiungere un sottoinsieme omogeneo di particelle. Questo sottoinsieme omogeneo di particelle viene quindi mediato insieme per produrre una ricostruzione 3D. A questo punto i dati vengono spesso corretti ulteriormente per produrre la mappa di massima qualità possibile, ad esempio attraverso il perfezionamento CTF, le correzioni di distorsione37 e la lucidatura bayesiana38 . Il risultato di questa elaborazione delle immagini è una mappa crioEM 3D del campione biologico di interesse. L'intervallo di risoluzione raggiunto in un esperimento automatizzato "standard" di singola particella da una griglia di qualità sufficiente, con dati raccolti su un sistema di microscopio a 300 kV è in genere compreso tra 10 Å e 2 Å a seconda delle dimensioni e della flessibilità del complesso proteico. Con un campione ideale, risoluzioni di ~ 1,2 Å sono state raggiunte utilizzando i flussi di lavoro SPA5. Mentre questo protocollo descrive in dettaglio i passi verso l'ottenimento di una mappa di densità EM, una volta che questo è in mano può essere ulteriormente interpretato attraverso l'adattamento e la raffinazione di un modello proteico (se la risoluzione è < 3,5 Å) o la costruzione de novo39. I dati associati agli esperimenti di determinazione della struttura possono essere depositati in archivi pubblici online, tra cui mappe di densità EM (Electron Microscopy Data Bank)40, coordinate atomiche risultanti (Protein Data Bank)41 e set di dati grezzi (Electron Microscopy Public Image Archive)42.
In questo protocollo, il complesso proteico di membrana esterna RagAB (~340 kDa) di Porphyromonas gingivalis viene utilizzato come esempio di complesso macromolecolare10 (EMPIAR-10543). Per coloro che sono nuovi a cryoEM, il supporto per i campioni attraverso questa pipeline dal campione alla struttura è disponibile, soggetto a revisione tra pari, attraverso schemi di accesso finanziati come iNEXT Discovery e Instruct.
Protocollo
1. Vetrificazione della griglia
NOTA: per tutte le fasi dei passaggi 1 e 2, assicurarsi che tutti gli utensili siano puliti, asciutti e a temperatura ambiente prima di raffreddarli a temperatura LN2 , utilizzando LN2 appena travasato per ridurre la contaminazione da ghiaccio. Ove possibile, lavorare all'interno di un ambiente a umidità controllata con < 20% di umidità relativa. Assicurarsi che i dispositivi di protezione individuale appropriati e la documentazione H & S siano in atto prima di iniziare il lavoro.
- Assicurarsi che il campione di interesse sia pronto per la preparazione del campione.
- Scegli le griglie crioEM appropriate e assicurati che siano state rese idrofile utilizzando la scarica di bagliore o il trattamento al plasma. È disponibile un'ampia varietà di sistemi e varianti al protocollo, ma tutti comportano il posizionamento delle griglie in un sistema di scarico a bagliore / pulizia al plasma e l'esecuzione di un programma che pomperà la camera al livello di vuoto desiderato, prima di introdurre una specifica miscela di gas / vapore chimico o aria nel sistema. Una corrente elettrica viene fatta passare attraverso il sistema, ionizzando le particelle di gas e inducendo la superficie delle griglie ad essere resa più idrofila.
- Avviare il dispositivo di congelamento a tuffo per la vetrificazione della rete accendendo il sistema utilizzando l'interruttore di alimentazione sul retro e attendere il caricamento del touch screen.
- Utilizzando lo stilo o le dita in dotazione, in Console, impostare la temperatura di lavoro desiderata della camera (l'intervallo disponibile è 4-60 °C, consigliato per la maggior parte delle macromolecole 4-6 °C).
- Riempire l'umidificatore con 50 ml di acqua di laboratorio di tipo II utilizzando una siringa, tramite il tubo di gomma nella parte inferiore dell'umidificatore. Assicurarsi di rimuovere l'aria intrappolata nella siringa prima del riempimento. Fare attenzione a non riempire eccessivamente l'umidificatore o l'acqua trasuderà nella camera. Una volta riempito l'umidificatore, estrarre lo stantuffo della siringa di 5-10 ml per creare un sigillo sottovuoto.
- In Console, impostare l'umidità relativa desiderata per la camera (l'intervallo disponibile è 0-100%, l'umidità del 95-100% viene in genere utilizzata). Lasciare l'umidità impostata su "off" fino a immediatamente prima della creazione della griglia in modo che la camera non si bagni troppo.
- Ottenere le pinzette del dispositivo di congelamento a tuffo e la carta da filtro tagliata alla dimensione corretta per adattarsi ai pad, acquistati o utilizzando un timbro per tagliare un'apertura della dimensione appropriata.
- Preparare il criogeno per il congelamento a tuffo.
- Posizionare il supporto della scatola a criogriglia metallica, la tazza di criogeno e le zampe del ragno metallico nel contenitore del refrigerante.
- Raffreddare il contenitore riempiendo la camera esterna con LN2. Tenere la camera esterna rabboccata per coprire la parte superiore del supporto della scatola crio-griglia. Aggiungere ~ 1 cm di LN2 aggiuntivo alla tazza criogena per facilitare l'equilibrio del sistema alla temperatura LN2 .
NOTA: L'anello anti-contaminazione può essere utilizzato per limitare la condensazione dell'aria umida attorno alla tazza di criogeno e portare alla contaminazione da crio-refrigerante / etano. Questo non è generalmente richiesto in un ambiente a umidità controllata. Se si utilizza l'anello anti-contaminazione, fare attenzione a non riempire eccessivamente il contenitore con LN2 o potrebbe fuoriuscire quando l'anello viene premuto nel contenitore più avanti nel processo. - Attendere 3-5 minuti per osservare l'ebollizione delle zampe di ragno, quindi attendere altri 3 minuti per assicurarsi che la tazza di criogeno sia sufficientemente fredda da condensare il mezzo di vetrificazione.
- Liquefare il criogeno (etano liquido) nella tazza di criogeno.
- Prendi il tubo del cilindro di etano con tubi sottili e un ugello per erogare il gas. Una punta della pipetta P200 con l'apertura aperta tagliando la punta stessa usando una lama di rasoio è l'ideale qui. È necessaria un'apertura più ampia per evitare che l'etano si solidifichi sulla punta e blocchi il flusso di gas.
- Assicurandosi che la tazza di criogeno non contenga LN2 rimanente, prendere l'ugello del gas etano e posizionarlo all'interno della tazza di criogeno. Utilizzando il regolatore della bombola del gas, avviare un flusso basso ed erogare gas criogeno nella tazza di criogeno per condensare il gas. Tenere la punta da cui scorre il gas premuto direttamente contro la parete della coppa criogenica, ma spostarla delicatamente avanti e indietro con un movimento di picchiettamento contro la superficie. Regolare il flusso del gas per consentire a un flusso basso e costante di iniziare a condensare / liquefare in modo controllato all'interno della coppa criogena.
- Riempire la tazza appena sotto il bordo del ragno d'argento e interrompere il flusso di gas, quindi rimuovere con attenzione la linea del gas per evitare di contaminare l'LN2 circostante con etano.
- Riempire il contenitore del refrigerante con LN2, facendo molta attenzione a non versarne alcuno nell'etano liquido.
- Lasciare le zampe dei ragni in posizione per ~ 3-5 minuti per garantire che l'etano liquido sia bilanciato a una temperatura sufficientemente fredda. Il criogeno inizierà ad apparire torbido/ leggermente opaco. Ciò indica che è vicino al suo punto di congelamento. In questa fase, usa una pinzetta per rimuovere il ragno. Finché l'LN2 viene conservato all'interno del contenitore che circonda la tazza criogenica, l'etano rimarrà liquefatto e adatto alla vetrificazione per 1-2 ore. Tuttavia, mirare a completare la procedura il più rapidamente possibile, soprattutto in ambienti non controllati dall'umidità, per ridurre la contaminazione da ghiaccio.
NOTA: se il ragno sembra essere "bloccato", utilizzare un oggetto metallico come un dado e tenerle contro le zampe del ragno per riscaldarle leggermente, quindi rimuovere le gambe.
- Preparare il dispositivo di congelamento a tuffo e gli accessori per la vetrificazione del campione.
- Aggiungere le scatole di stoccaggio della griglia al supporto della scatola della crio-griglia metallica e durante tutta la procedura assicurarsi che LN2 sia mantenuto rabboccato appena sopra il livello delle scatole della griglia (di solito ogni ~ 5 minuti).
- Nella schermata del dispositivo di congelamento a tuffo, nella casella Parametri di processo immettere i parametri scelti, tra cui: tempo di macchia (il tempo in cui i cuscinetti del dispositivo di congelamento a tuffo si uniranno), forza (la distanza dei tamponi di blotting dalla griglia, che altera il gradiente di formazione del ghiaccio) e totale (numero di volte in cui i pad di blotting entreranno per incontrarsi). Scegli questi parametri in base al singolo dispositivo di congelamento a tuffo e al comportamento della macromolecola. I valori tipici sono una forza di macchia compresa tra 0 e 5, un tempo di macchia da 1 a 6 s e un totale di macchia di 1. Il tempo di attesa tipico (tempo tra l'avvio della macchia e l'inizio della macchia) e il tempo di drenaggio (tempo dopo il blotting prima di immergersi) è 0-2 s.
NOTA: a seconda delle preferenze dell'utente, è possibile selezionare opzioni aggiuntive nella sezione Opzioni, Varie , tra cui Usa pedale per passare al passaggio successivo su ogni pressione, Salta trasferimento griglia (salta il passaggio finale in cui il braccio della pinzetta viene leggermente sollevato), Disattiva umidità durante il processo (mentre il campione viene applicato, interrompe l'umidificazione attiva della camera che può rendere più difficile vedere la griglia) e Autoraise Ethanelift (combina il passo delle pinzette che vengono sollevate nella camera e sollevano il contenitore del refrigerante - salta Solleva il passaggio del contenitore di etano ). Qui, tutte queste opzioni sono attivate. - Posizionare saldamente il contenitore del refrigerante sul braccio della piattaforma mobile sotto la camera
- Inserire carta assorbente fresca su ciascun braccio blotter assicurandosi che le clip ad anello di plastica siano fissate. Ogni carta da filtro consentirà 16 macchie (le braccia ruotano la carta assorbente). Premere il pulsante Reimposta carta macchia nella sezione Controlli .
- Eseguire 1 ciclo completo del processo di vetrificazione del dispositivo di congelamento a tuffo per garantire che ogni parte mobile si comporti come previsto.
- Premere (o utilizzare il pedale) per posizionare una nuova griglia, quindi Avviare il processo, quindi Elaborare e quindi Continuare. In questa fase, fai attenzione per assicurarti che i bracci di blotting si contattino l'un l'altro come previsto.
- Accendere l'umidificatore. Verrà prodotto vapore acqueo (purché l'umidità impostata sia superiore a quella attualmente presente nella camera).
- L'esemplare di interesse può ora essere vetrificato. Usa il pedale o Posiziona nuova griglia e l'asta di immersione scenderà fuori dalla camera consentendo alle pinzette di essere fissate al supporto.
- Utilizzando le pinzette del dispositivo di congelamento a tuffo, raccogliere la griglia cryoEM a scarica di bagliore / plasma pulita desiderata, avendo cura di notare quale lato è il lato corretto da utilizzare per l'applicazione del campione in base al produttore della griglia. Prelevare la griglia dal cerchio, avendo cura di evitare un contatto eccessivo/non necessario con le pinzette in quanto ciò danneggerebbe il supporto. Fissare la griglia nelle pinzette spostando la clip nera verso il basso nella parte increspata delle pinzette. La griglia deve essere tenuta saldamente, ma la clip non deve essere troppo in basso in quanto contatterà i tamponi di blotting, portando a un blotting irriproducibile e, in seguito, le pinzette dovranno essere tenute al di sotto di questo punto quando si rilascia la clip.
- Posizionare le pinzette del dispositivo di congelamento a tuffo tenendo la griglia crioEM sul braccio pneumatico con il lato corretto rivolto verso la mano dominante. Il design delle pinzette e della camera del dispositivo di congelamento a tuffo sono tali che il campione può essere applicato attraverso il lato destro o sinistro della camera, in base alla maneggevolezza dell'utente.
NOTA: l'applicazione del campione su lati diversi con gli stessi parametri di blotting raramente si traduce in risultati comparabili, quindi i ricercatori mancini potrebbero dover regolare i loro parametri di blotting indipendentemente dai loro colleghi destrimani. - Premere Start Process e la griglia tenuta nelle pinzette verrà portata nella camera e il contenitore del refrigerante verrà sollevato.
- Premere Processo e le pinzette sposteranno la griglia nella posizione in cui è possibile utilizzare una pipetta per applicare il campione alla griglia. Aprire la porta laterale rivolta verso il lato corretto della griglia e applicare il campione mediante pipettaggio, assicurandosi che la punta della pipetta non tocchi la griglia in quanto potrebbe causare danni al supporto della griglia / piegatura della griglia, ma erogare il liquido abbastanza vicino in modo che la goccia eroghi sulla griglia. In genere, vengono applicati 3-5 μL.
- Premere Continua e i parametri predefiniti dell'utente cancelleranno la griglia e quindi immergeranno le pinzette con la griglia montata nella tazza del liquido di raffreddamento per la vetrificazione del campione. Le pinzette scenderanno in combinazione con il braccio che tiene il contenitore del refrigerante e il refrigerante, mantenendo la griglia immersa nel criogeno.
- Trasferire la griglia dalla tazza di criogeno alla scatola di stoccaggio della griglia immersa in LN2.
- Staccare le pinzette dal braccio della pinzetta, facendo molta attenzione a non contattare la griglia vetrificata con i lati della coppa criogenica. Regolare l'impugnatura in modo che le pinzette siano tenute comodamente. Il più rapidamente e attentamente possibile, spostare la griglia dal criogeno all'LN2. Con una mano, tieni le pinzette chiuse usando le dita e con l'altra mano, fai scorrere la clip nera verso l'alto fuori mano, tenendo le pinzette chiuse. Regola nuovamente l'impugnatura e manipola la griglia nella scatola di stoccaggio della griglia.
- Ripeti i passaggi 1.10-1.19 fino a quando non vengono create tutte le griglie (una sessione tipica comporterà la creazione di 4-12 griglie). Conservare tutte le scatole di stoccaggio della griglia contenenti griglie in LN2 dewar fino alle fasi successive.
2. Griglie di ritaglio per il caricamento in un microscopio autoloader
- Ritaglia le griglie nell'assemblaggio autogrid secondo il protocollo descritto in precedenza 28.
3. Accesso remoto sicuro ai microscopi
NOTA: con i controlli COVID-19 al momento della scrittura, ma anche con le preoccupazioni ambientali associate ai viaggi internazionali, più strutture di microscopia hanno offerto servizi in cui l'utente opera da remoto. Il metodo di implementazione per questo varierà in base alla configurazione IT locale di ciascuna struttura e alle esigenze della sua comunità di utenti interni ed esterni. Qui viene descritto il processo per accedere da remoto ai crioEM presso eBIC e controllare il microscopio tramite il software EPU.
- Accedi in remoto a cryoEM. L'accesso remoto è mediato tramite il software NoMachine per accedere al PC di supporto del microscopio ed è configurato per consentire l'accesso solo agli utenti registrati durante una visita tramite le credenziali di accesso FedID degli utenti. L'accesso rimane attivo solo per la durata della sessione.
- Aprire NoMachine e avviare una nuova connessione NX per nx-cloud.diamond.ac.uk con autenticazione tramite password.
- Apri la connessione e accedi con il nome utente fedid@fed.cclrc.ac.uk e la password FedID. Fare doppio clic sull'icona corrispondente al microscopio pertinente tra le opzioni disponibili per aprire una connessione al PC di supporto pertinente.
- Immettere nome utente clrc\FedID e password nella schermata di accesso di Windows.
- Apri il software TeamViewer dall'icona del desktop e connettiti a PartnerID: TEM con la password fornita. Questo stabilisce la connessione dal PC di supporto al PC TEM. Il pulsante Next Monitor nella barra multifunzione TeamViewer può essere utilizzato per passare dall'interfaccia utente del microscopio alla finestra EPU.
- Le funzioni del microscopio possono quindi essere controllate dagli utenti direttamente tramite l'interfaccia EPU.
4. Caricamento dei campioni in un microscopio autoloader e screening per la qualità del ghiaccio e del campione
NOTA: In questa sezione viene utilizzato un microscopio con caricatore automatico e software EPU per lo screening dei campioni, ma ciò può essere ottenuto utilizzando altri software e un sistema di ingresso laterale e crioEM di altri produttori.
- Caricare le griglie ritagliate nel caricatore automatico del microscopio come descritto in precedenza28.
- Nella scheda caricatore automatico dell'interfaccia utente del microscopio, spegnere la finestra di dialogo Opzioni utilizzando la freccia e premere il pulsante Inventario . Questo controllerà sequenzialmente ogni posizione nella cassetta per determinare se è presente una cartuccia. Gli slot occupati saranno etichettati in blu. Se tutti gli slot occupati sono stati mappati, premere nuovamente il pulsante Inventario per fermarsi dopo la posizione corrente, altrimenti lasciare in esecuzione fino a quando tutti gli slot occupati non sono stati mappati. Etichettare tutti gli slot occupati con i dettagli del campione nelle caselle fornite.
- Evidenziare la griglia da trasferire nella colonna del microscopio e fare clic su Carica. L'etichetta dello slot passerà dal blu al giallo una volta che la griglia sarà stata caricata correttamente sul palco. Procedere allo screening delle griglie.
- Aprire il software EPU. Nella pagina Preparazione , selezionare Ottica di acquisizione e impostazioni , quindi selezionare il predefinito Atlas dal menu a discesa. Scegliere i preset di impostazione del fascio appropriati (ad esempio, 64x mag nominale, dimensione spot 5, Microsonda, con un'area illuminata nella gamma parallela per il rilevatore Falcon - per ulteriori informazioni scegliere i preset di impostazione del fascio vedi28). Premere Imposta per spingere i parametri al microscopio.
- Premere Apri valvole a colonna e inserire FluScreen. Verificare che un raggio sia visibile e sufficientemente diffuso e centrato per coprire il rilevatore. Se necessario, passare a una regione più sottile della griglia utilizzando il joystick o il menu dello stage per controllare i movimenti dello stage in X e Y.
- Solleva FluScreen e scatta un'immagine usando il pulsante Anteprima in EPU. Sulla base dell'immagine acquisita, la dose può essere aumentata passando a un numero inferiore di punti e viceversa.
- In EPU, vai alla pagina Atlas e premi Nuova sessione. Selezionare il formato immagine MRC e immettere un nome e un percorso di cartella appropriati per il salvataggio della sessione di screening, quindi fare clic su Applica.
- Seleziona Screening dal menu a sinistra. Seleziona le caselle di controllo accanto a ciascuna griglia per acquisire un montaggio dell'atlante. Avviare la sessione di screening in EPU. Verrà acquisito un atlante per ogni griglia controllata, con un numero di quadrati di griglia disponibili elencati al completamento. Ogni atlante può essere visualizzato evidenziandolo nella pagina di screening, completo di un mark-up che mostra i quadrati della griglia con uno spessore di ghiaccio previsto simile raggruppato per colore.
- Al termine, rivedere gli atlanti raccolti e identificare le griglie adatte a valutare la qualità del campione a ingrandimenti più elevati (cioè quelli con un numero appropriato di quadrati della griglia che non sono né asciutti né oscurati da ghiaccio spesso). Evidenziare la griglia scelta nel menu di screening EPU e fare clic su Carica campione.
- Utilizzare i preset di impostazione del fascio (vedere 28 per la spiegazione dei preset di impostazione del fascio desiderati per ogni fase) e la funzione di anteprima per esaminare i quadrati della griglia desiderati in modo più dettagliato.
- Dal menu di screening Atlante, selezionare la griglia attualmente caricata e spostare lo stage in un quadrato della griglia contenente fori riempiti facendo clic con il pulsante destro del mouse sulla posizione desiderata sull'immagine della griglia e selezionando Sposta nel quadrato della griglia.
- Tornare a EPU, pagina Preparazione e selezionare il predefinito GridSquare .
- Aprire la pagina EPU, Funzioni automatiche ed eseguire Auto-eucentrico per inclinazione dello stadio con il preset GridSquare per spostare il campione all'altezza eucentrica.
NOTA: è disponibile anche l'inclinazione auto-eucentrica del fascio, che è più veloce ma in genere meno accurata dell'auto-eucentrico per inclinazione dello stadio. - In EPU, Preparazione, acquisire una nuova immagine di anteprima GridSquare. Notate i diversi valori di grigio tra i diversi fori che indicano diversi spessori di ghiaccio. Spostate lo stage su un foro facendo clic con il pulsante destro del mouse > sposta lo stage qui. Selezionate il predefinito Foro/Altezza eucentrica (Hole/Eucentric Height ) e Anteprima (Preview).
NOTA: A seconda del peso molecolare e della forma della particella di interesse, potrebbe essere possibile identificarla con l'ingrandimento Foro/Altezza Eucentrica. - Selezionare il preset acquisizione dati e impostare un ingrandimento che consenta una facile identificazione delle particelle (corrispondente a un campionamento di oggetti generalmente <2 Å/pixel). Impostare l'offset di sfocatura su ~-3 a -5 μm con una dose di elettroni di esposizione di ~40-80 e-/ Å2.
- Scorrere i passaggi in 4.4 per valutare una gamma di spessori di ghiaccio per la distribuzione, l'orientamento e la contaminazione delle particelle attraverso la griglia. La distribuzione delle particelle può variare vicino ai bordi rispetto al centro del foro e quindi è importante esaminare diverse posizioni con il foro.
- Esamina tutte le griglie che mostrano promesse dagli atlanti come aventi quadrati di griglia sufficienti. Tenerli nel microscopio e procedere all'acquisizione dei dati utilizzando l'EPU, oppure scaricare i campioni dal microscopio e conservarli sotto LN2 fino a quando non viene programmata la raccolta dei dati.
5. Raccolta di dati crioEM a singola particella (con particolare attenzione al funzionamento remoto)
NOTA: un protocollo dettagliato per l'acquisizione dei dati con EPU è descritto nel manuale del produttore e altrove28. Qui vengono evidenziate le modifiche di questo protocollo per il funzionamento remoto (vale a dire la riduzione dell'uso dei pannelli manuali per svolgere attività e l'utilizzo di alternative basate su software).
- A meno che non ne sia già stato raccolto uno durante la sessione, raccogli un Atlante per la griglia.
- Definire ciascuno dei preset di impostazione del fascio in base alle esigenze sperimentali del progetto.
- Eseguire calibrazioni di spostamento dell'immagine28.
- Configurare la sessione EPU.
- In EPU, selezionare la pagina EPU , quindi Configurazione sessione, selezionare Nuova sessione, quindi Nuovo dalle preferenze.
- Seleziona Nuova sessione apparirà un pop-up che fornisce un'opzione per utilizzare le impostazioni precedenti. Yes caricherà automaticamente le impostazioni dall'EPU precedente (ad esempio, portacampione, intervallo di sfocatura, impostazioni di messa a fuoco automatica, tipo di griglia) nella sessione EPU corrente. La selezione di Nuovo dalle preferenze consente all'utente di scegliere un file con le preferenze salvate (ad esempio, intervallo di sfocatura, impostazioni di messa a fuoco automatica, tipo di griglia) e queste informazioni verranno precaricate in EPU.
- Inserisci il nome della sessione con qualcosa di informativo. La struttura locale può suggerire una convenzione di denominazione.
- In Tipo selezionare Manuale.
- Per la modalità di acquisizione, selezionare un centraggio accurato del foro o un'acquisizione più rapida.
- In Formato immagine, selezionare il formato desiderato.
- Selezionare una cartella di archiviazione appropriata e EPU creerà una directory con il nome della sessione.
- Selezionare il portacampioni appropriato in base al tipo di griglia e alla spaziatura dei fori utilizzati (ad esempio, Quantifoil 1.2/1.3) e premere Applica. Questo protocollo descrive il processo di generazione di una maschera per una matrice regolare di fori
- Selezionare un quadrato della griglia iniziale e impostare un modello di acquisizione.
- Vai selezione quadrata, se tutti i quadrati sono verdi, fai clic su deseleziona tutto in alto a sinistra.
- Apri riquadri (fai clic con il pulsante destro > apri riquadro). Selezionare un quadrato (fare clic con il pulsante destro del mouse > aggiungere, fare clic con il pulsante destro del mouse > sposta lo stage nel quadrato della griglia).
- Andare a Selezione fori e premere Auto Eucentric. Attendi fino a quando questo è completo e un'immagine grid square è stata scattata. Se la funzione automatica fallisce, ciò potrebbe essere dovuto al fatto che l'altezza è significativamente disattivata; in tal caso può essere regolato manualmente utilizzando l'ingrandimento FluScreen at Grid Square.
- Misurare le dimensioni del foro. Sposta e regola i cerchi gialli in modo che siano sopra i fori con dimensioni e spaziatura corrette.
- Premere Trova fori. Controllare che i fori siano stati trovati correttamente. In caso contrario, modificare la dimensione del foro e trovare nuovamente i fori. Ripeti questa operazione finché non trova correttamente il foro. Se fallisce costantemente, considera di passare a un numero inferiore (più luminoso) di dimensioni dello spot all'ingrandimento quadrato della griglia.
- Utilizzare l'istogramma Filtra qualità ghiaccio sulla destra per regolare la selezione dei fori. Questo può essere utile per escludere aree con ghiaccio spesso e ghiaccio sottile. Questo sarà ricordato per i futuri quadrati della griglia selezionati durante questa sessione.
- Ottimizzate la selezione dei fori con gli strumenti disponibili nel menu Seleziona (Select ) in alto. Ad esempio, fate clic su Rimuovi fori vicino alla barra della griglia.
- Vai a Definizione modello e premi Acquisisci.
- Fate clic su Trova (Find) e foro centrale (Center Hole). Ora ci sarà un'immagine di un buco con un cerchio giallo attorno al buco.
NOTA: se fatica a trovare il foro, inserire l'apertura dell'obiettivo. Se ancora non riesce a trovare il foro, provate ad aumentare il tempo di esposizione per il foro/altezza eucentrica preimpostata o ad aumentare la sfocatura per questo preset o a bloccare l'immagine. Una grande modifica della sfocatura può alterare l'allineamento dello spostamento dell'immagine. - Modificare i tempi delay after stage shift e delay after image shift a 1-5 s.
- Selezionare Il valore massimo di spostamento dell'immagine (se l'opzione è disponibile) è come desiderato. Se viene utilizzata la raccolta di spostamento dell'immagine libera da aberrazione, questo valore viene definito nel file di configurazione EPU, altrimenti 5 μm è un valore standard.
- Fai clic su Aggiungi area di acquisizione, quindi fai clic in un punto qualsiasi dell'immagine. Spostare l'area di acquisizione nella posizione desiderata (cioè sul bordo di un foro) in modo che le aree di acquisizione non siano doppiamente esposte con il raggio (il quadrato nel cerchio verde rappresenta l'area del rilevatore, il cerchio verde è il diametro del fascio).
- In alto a destra, aggiungi la gamma di sfocatura. Quindi aggiungi altre aree di acquisizione. Un tipico elenco di defocus per un progetto di proteina di membrana è da -0,8 a -3 μm di defocalizzazione.
- Fare clic su Aggiungi area di messa a fuoco automatica e fare clic in un punto qualsiasi dell'immagine. Spostate l'area di messa a fuoco automatica sul carbonio che circonda un foro. La pratica standard è quella di mettere a fuoco automaticamente dopo la centratura quando si utilizza AFIS, o ogni 5-15 μm, a seconda della variazione di altezza z attraverso il quadrato.
- Fare clic su Aggiungi area di misurazione della deriva, la misurazione della deriva eseguita una volta per quadrato della griglia, con una soglia impostata di 0,05 nm/s è un'impostazione standard. L'area di misurazione della deriva può (ed è una buona idea) sovrapporsi direttamente all'area di messa a fuoco automatica. Assicurati che né la deriva né l'area di messa a fuoco automatica si sovrappongano a un'area di acquisizione.
NOTA: il modello può essere controllato utilizzando la funzione di esecuzione del modello. Questa è una buona idea per vedere se le aree di acquisizione devono essere spostate (ad esempio, troppo / non abbastanza carbonio nelle immagini), ma non è necessario. - Torna alla selezione quadrata e sulla griglia seleziona i quadrati per l'acquisizione. Utilizzare il numero di aree di acquisizione e la velocità di acquisizione dati prevista (dalla struttura basata su rilevatori e configurazione sperimentale) per prevedere quante aree di acquisizione sono necessarie.
- Quando tutti i quadrati desiderati sono selezionati, premere Prepara tutti i quadrati.
- Una volta raccolto ogni quadrato, naviga tra i quadrati della griglia e perfeziona i fori usando il pennello di selezione.
- Spostati in una posizione dello stage sopra il campione e utilizza le funzioni automatiche per impostare l'altezza eucentrica. Eseguire gli allineamenti al microscopio come descritto in precedenza28, ma invece di eseguire manualmente l'allineamento senza coma e correggere manualmente l'astigmatismo oggettivo, utilizzare gli strumenti di allineamento all'interno del software. In breve, impostare le condizioni del fascio di acquisizione, assicurarsi che l'apertura dell'obiettivo (OA) venga rimossa e che lo stadio sia posizionato su un'area stabile del fascio di campione ad altezza eucentrica. Eseguire l'allineamento senza coma all'interno delle funzioni automatiche prima di reinserire e centrare l'OA e correggere l'astigmatismo della lente dell'obiettivo con EPU. Assicurarsi che entrambi gli allineamenti convergano su valori adeguati (<150 nm di coma e astigmatismo vicino allo zero.
- Prima di iniziare l'acquisizione automatica, assicurarsi che la turbopompa del caricatore automatico sia spenta e che l'apertura dell'obiettivo sia inserita.
- In Acquisizione automatica premere Avvia esecuzione per avviare l'acquisizione automatica dei dati.
6. Elaborazione delle immagini per produrre la mappa di densità EM
NOTA: la maggior parte delle strutture crioEMA offre la pre-elaborazione di filmati micrografici "al volo". Ci sono una vasta gamma di pacchetti software e approcci disponibili per questo, tra cui le pipeline RELION28,33, cryoSPARC43, Scipion34 e WarpEM44. Una pipeline basata su RELION è descritta qui e si presume che l'utente abbia spostato i filmati micrografici in una posizione di archiviazione appropriata con accesso alle risorse di elaborazione. Viene fornita una panoramica del processo e dei risultati rappresentativi per un progetto di proteine di membrana, una descrizione dettagliata e un tutorial passo passo sono disponibili sulla homepage di RELION: https://www3.mrc-lmb.cam.ac.uk/relion.
- Eseguire analisi "al volo" della correzione del movimento micrografico e della stima CTF. Avviare RELION all'interno della directory del progetto. Pianifica i processi di importazione, correzione del movimento e stima CTF in loop in modo che siano simultanei con la raccolta e il trasferimento dei dati. Uno script di analisi micrografica28 fornisce un feedback visivo in tempo reale sull'astigmatismo e sui valori di sfocatura stimati (vedere risultati rappresentativi).
- Scegli le particelle dalle micrografie pre-elaborate. Ci sono una serie di pacchetti software automatizzati per la raccolta delle particelle tra cui scegliere. Le opzioni di prelievo senza riferimento e basate su modello sono disponibili nella scheda Prelievo automatico di RELION37. Altri programmi possono essere utilizzati per vari passaggi, ad esempio utilizzando crYOLO per il prelievo di particelle35.
- Estrarre particelle dalle micrografie corrette CTF.
NOTA: per ridurre il tempo di calcolo necessario per le fasi iniziali di "pulizia", elaborazione, ridimensionamento/bin delle particelle al momento dell'estrazione. I dettagli su come eseguire il processo di estrazione sono disponibili nell'esercitazione RELION 3.1. Per questo progetto, le particelle sono state inizialmente legate da un fattore di 2. - Eseguire la media delle classi 2D. La classificazione tra 100-200 classi funziona bene per la maggior parte dei set di dati contenenti ≥ 100.000 particelle. Non è consigliabile utilizzare molte più di 200 classi o meno di 50 classi, anche quando i set di dati sono piccoli a meno che il campione non abbia un'elevata simmetria (ad esempio, virus icosaedrico), nel qual caso meno di 50 classi potrebbero comunque dare un buon risultato. Impostare il diametro della maschera abbastanza grande da ospitare la dimensione più lunga della particella, ma abbastanza stretto da escludere eventuali particelle vicine (ciò potrebbe richiedere alcuni tentativi ed errori).
- Seleziona le buone classi (ad esempio, quelle con dettagli strutturali) usando il lavoro di selezione del sottoinsieme. Esempi di medie di classe 2D buone e cattive possono essere trovati nella sezione dei risultati rappresentativi.
- Generare un modello iniziale de novo dai dati utilizzando il processo di modello iniziale 3D in RELION.
NOTA: pile di particelle meno pulite possono trarre vantaggio dal perfezionamento multi-riferimento ab initio SGD (discesa del gradiente stocastico) poiché ciò offre un'ulteriore opportunità per setacciare le particelle spazzatura / sub-ottimali. Selezionare un diametro della maschera che possa contenere la particella di interesse e lasciare i valori predefiniti per i campi nella scheda "SGD" poiché questi funzionano regolarmente bene. Assicurarsi che il modello iniziale appaia ragionevole in Chimera (o in un altro programma di visualizzazione appropriato) (vedere i risultati rappresentativi). - Eseguire la classificazione 3D per affrontare l'eterogeneità dei dati utilizzando l'output del passaggio 6.6 come modello di riferimento. Valuta le mappe risultanti in Chimera. Elabora le pile di particelle corrispondenti a stati conformazionali univoci in modo indipendente. Utilizzare il processo di selezione dei sottoinsiemi per selezionare una classe/classi di interesse e generare file particles.star per le pile di particelle associate.
- Esegui il perfezionamento automatico 3D. Utilizzare le medie di classe 3D ottenute nel passaggio precedente come riferimenti per il perfezionamento delle pile di particelle corrispondenti. Se la risoluzione del raffinamento si avvicina al limite di Nyquist dei dati, estrai le particelle senza ridimensionare. Dopo la ri-estrazione, ripetere il processo di auto-raffinazione 3D con la pila di particelle non accoppiate. In questo caso, i modelli di riferimento 3D devono essere riscalati in modo tale che le dimensioni dei pixel e delle caselle siano coerenti con quelle delle immagini delle particelle riestratte. Utilizzare lo strumento da riga di comando relion_image_handler per eseguire questa operazione.
- Utilizzare la simmetria nel raffinamento, se appropriato. Se una mappa ricostruita possiede simmetria, allineare la mappa sull'asse di simmetria appropriato utilizzando lo strumento da riga di comando relion_align_symmetry. Utilizzare la mappa allineata risultante come riferimento in un nuovo processo di auto-perfezionamento 3D con l'operatore di simmetria appropriato specificato nella scheda di riferimento.
- Affina le mappe dal perfezionamento automatico 3D. Questo viene fatto usando il lavoro di post-elaborazione in RELION, ma prima deve essere creata una maschera adatta dalla mappa raffinata. I passaggi della creazione e della post-elaborazione della maschera sono dettagliati nel tutorial RELION (vedere anche i risultati rappresentativi).
NOTA: La risoluzione di molte ricostruzioni può essere ulteriormente migliorata utilizzando le funzionalità di lucidatura bayesiana e raffinamento CTF in RELION. Utilizzare il tipo di lavoro di raffinamento CTF per stimare e correggere le aberrazioni di ordine superiore (inclinazione del fascio, aberrazioni trilobate e aberrazioni di 4° ordine) e, come lavori separati, l'ingrandimento anisotropo e la sfocatura per particella. Successivamente, utilizzare il lavoro di lucidatura bayesiana (addestrato o con valori predefiniti) per affrontare il movimento indotto dal fascio su base per particella. Come affrontato nel tutorial RELION 3.1, questi lavori trarranno probabilmente beneficio da un approccio iterativo (perfezionamento CTF → lucidatura bayesiana → auto-raffinamento 3D → post-elaborazione → ... loop) poiché entrambi beneficiano di modelli a risoluzione più elevata. - Correggere la maneggevolezza delle mappe di densità EM, se necessario. Esaminare le mappe per determinare se la maneggevolezza è corretta tentando di adattarsi a un modello atomico esistente o valutando la maneggevolezza delle regioni alfa elicoidali. Dove richiesto, capovolgere la mappa lungo l'asse z in UCSF Chimera45 utilizzando il comando 'vop zflip'.
Risultati
Durante lo screening, le griglie possono essere scartate nella fase dell'atlante, dove le caratteristiche risolte a basso ingrandimento contrassegnano la griglia come non adatta per l'acquisizione dei dati. Ad esempio, se una griglia è stata soggetta a danni meccanici significativi con la maggior parte dei quadrati della griglia rotti (Figura 2A) o dove la griglia sembra essere "asciutta", senza ghiaccio vitreo (Figura 2B). Tali griglie sono in genere identificabili in quanto i bordi dei quadrati della griglia appaiono nitidi e distinti. Nella maggior parte delle griglie realizzate utilizzando il dispositivo di congelamento a tuffo, si osserva un gradiente di ghiaccio (Figura 2C, D). La distribuzione delle particelle, a seconda del campione di interesse, può variare notevolmente con lo spessore del ghiaccio e quindi si consiglia di vagliare una gamma di quadrati della griglia per valutare la distribuzione delle particelle. Gli strumenti sono stati implementati all'interno del software EPU durante la fase di screening dell'atlante per aiutare l'utente a identificare quadrati di griglia di spessore di ghiaccio simile o diverso, che possono essere particolarmente utili per gli utenti che sono nuovi all'esame delle griglie crioEM (Figura 2E, F).

Figura 2: Esempi di montaggi "atlante" a basso ingrandimento da sessioni di screening. A) Una griglia che ha subito danni significativi con la maggior parte dei quadrati della griglia rotti - inadatti alla raccolta. B) Una griglia asciutta senza ghiaccio vitreo - inadatta alla raccolta. C) Una griglia che dimostra un gradiente di ghiaccio con ~ 50% della griglia utilizzabile. D) Un gradiente di ghiaccio con ~ 33% della griglia utilizzabile. Sia C che D, sono adatti per la raccolta dei dati se i quadrati della griglia utilizzabili hanno uno spessore del ghiaccio appropriato per la raccolta e ci sono abbastanza aree di acquisizione per soddisfare la durata minima di una raccolta (ad esempio, 24 h) E) Un atlante di esempio con gamma di spessori di ghiaccio. F) Lo stesso atlante presentato in E ma con, quadrati di griglia classificati e colorati dal software EPU in base allo spessore del ghiaccio. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Quando si esegue lo screening della distribuzione delle particelle, assicurarsi che i parametri di imaging, come l'ingrandimento e la dose totale di elettroni, siano simili a quelli che si prevede di utilizzare durante l'acquisizione dei dati al fine di fornire un quadro accurato dei risultati attesi. Durante lo screening, una distribuzione ideale delle particelle è monodispersa con una gamma di orientamenti delle particelle visibili (a seconda del campione e delle conoscenze esistenti della morfologia della particella, questo può essere difficile da accertare) (Figura 3A). Il ghiaccio dovrebbe essere il più sottile possibile mentre ospita le particelle più grandi, se il ghiaccio è troppo sottile può sciogliersi quando illuminato con il fascio di elettroni. Ciò causa un movimento eccessivo nella micrografia e le aree che mostrano questa caratteristica dovrebbero essere evitate (Figura 3B). Dall'esperienza collettiva, questo effetto è più comunemente osservato quando c'è detersivo nel tampone. Ciò può causare ghiaccio molto sottile al centro del foro e quindi le particelle possono essere fisicamente escluse e forzate verso il bordo. Questo effetto è osservato nella Figura 3C, ma in questo caso non è un esempio estremo e queste immagini contribuirebbero comunque utilmente a un set di dati. Infine, il ghiaccio deve essere vitreo; escludere dall'acquisizione dei dati qualsiasi area della griglia (o delle griglie) in cui la maggior parte o tutte le immagini scattate mostrano ghiaccio cristallino (Figura 3D). Spesso, il ghiaccio non vitreo viene osservato ai margini dei quadrati della griglia. Per ulteriori informazioni, i lettori rimandano a revisioni dettagliate delle variabili che possono essere modificate durante la vitrificazione della griglia16 e alle descrizioni del comportamento delle particelle nell'ambiente a film sottile46,47.

Figura 3: Micrografie rappresentative che mostrano diverse distribuzioni di particelle. A) Una distribuzione "ideale" di particelle monodisperse che adottano una gamma di orientamenti. B) Ghiaccio eccessivamente sottile nel mezzo del foro che si deforma dopo l'esposizione al fascio di elettroni causando un movimento eccessivo nella micrografia. Questo effetto è più spesso osservato quando il detergente è presente nel tampone C) Dove il ghiaccio è più sottile al centro del foro, questo esclude fisicamente le particelle dal centro, causando l'affollamento delle particelle verso il bordo del foro. In questo caso non è abbastanza estremo da impedire che queste immagini siano utili, ma suggerisce che vale la pena schermare aree leggermente più spesse. D) Il ghiaccio non è vitreo, i dati non dovrebbero essere raccolti su aree che assomigliano a questa micrografia di esempio. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
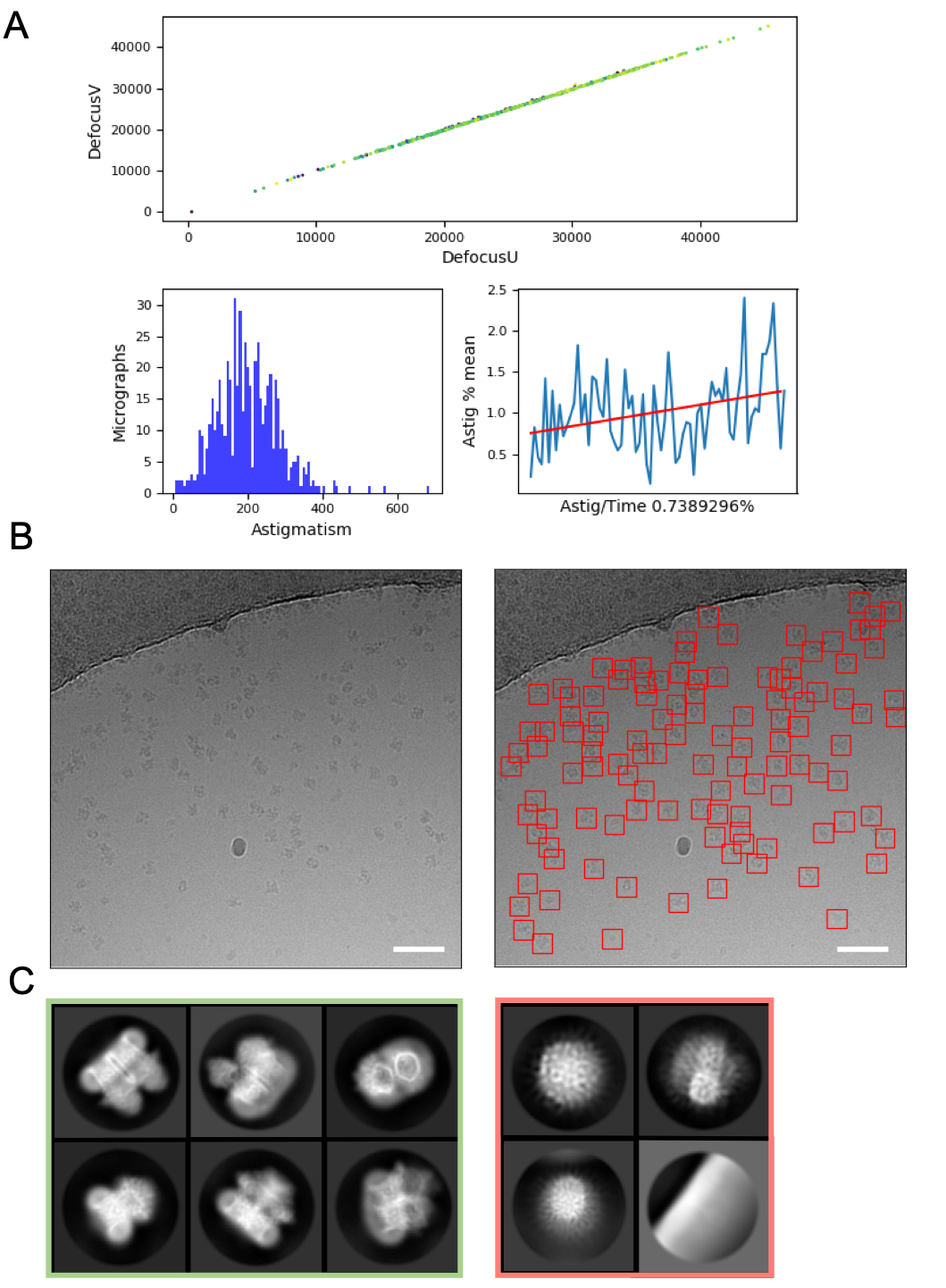
L'elaborazione delle immagini al volo può aiutare a rilevare errori e problemi con l'acquisizione dei dati e quindi è sempre consigliata ove possibile. Ad esempio, un movimento eccessivo all'interno dei micrografi può indicare che la turbopompa del caricatore automatico è attiva o che i dati vengono raccolti su un quadrato della griglia incrinato in cui il ghiaccio si muove in modo significativo nel fascio di elettroni, indicando che il quadrato della griglia dovrebbe essere saltato. Al volo la stima CTF può rivelare circostanze in cui viene applicato un punto di messa a fuoco positivo (piuttosto che defocus) (in cui vengono utilizzati programmi di stima CTF e parametri per trovare questi punti) e determinare lo sfasamento in cui viene utilizzata una piastra di fase Volta48. Le pipeline di elaborazione delle immagini al volo spesso includono un riepilogo grafico dei dati (Figura 4A) per rendere più facile per gli utenti valutare rapidamente la qualità delle micrografie e decidere se sono necessarie modifiche alla raccolta dei dati.
La selezione delle particelle dalle micrografie, evitando al contempo "falsi positivi" come la contaminazione o la pellicola di supporto della griglia può richiedere un'ottimizzazione. Tuttavia, i raccoglitori di particelle come crYOLO spesso funzionano sufficientemente bene utilizzando parametri predefiniti per un "primo passaggio" dei dati (Figura 4B), consentendo la progressione verso la media di classe 2D dove può essere più facile valutare la qualità dei dati e la probabilità di successo a valle. Per la maggior parte dei progetti, la classificazione 2D di ~ > particelle 10k dovrebbe iniziare a rivelare classi che hanno dettagli di struttura secondaria. Per passare al 3D, la fase di classificazione 2D dovrebbe in genere rivelare classi che rappresentano una gamma di orientamenti delle particelle. Se viene rivelato un orientamento preferito, potrebbero essere necessarie più iterazioni di preparazione del campione16 o un'ulteriore acquisizione di dati con il campione inclinato49. Tutte le classi che mostrano dettagli della struttura secondaria dovrebbero essere scelte per portare avanti l'analisi 3D, mentre le particelle "spazzatura" vengono scartate (Figura 4C).

Figura 4: Fasi iniziali di elaborazione delle immagini. A) Output da uno script di elaborazione delle immagini "al volo". B) Micrografia di esempio (a sinistra) con particelle opportunamente auto-selezionate identificate utilizzando il modello generale crYOLO (a destra, con particelle delimitate da quadrati rossi) Le barre di scala (bianche) sono 50 nm. C) Risultati della classificazione 2D che mostrano le classi che sono state scartate nel quadrato rosso e le classi da cui sono state selezionate le particelle per un'ulteriore elaborazione in verde. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
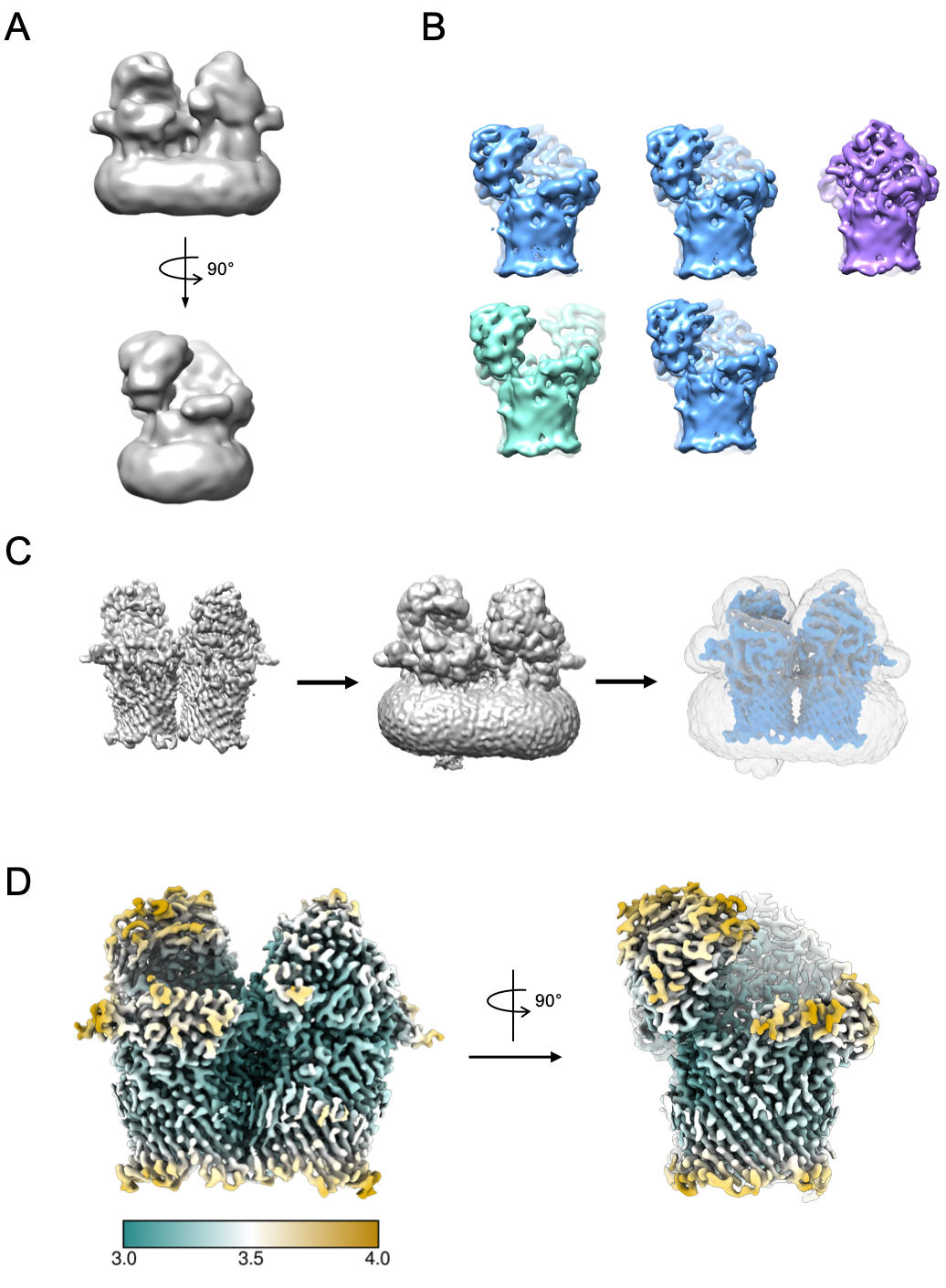
Un piccolo sottoinsieme di particelle può essere utilizzato per generare un modello iniziale (Figura 5A). Questo modello iniziale può quindi essere utilizzato come modello di partenza nella classificazione e nel perfezionamento 3D. Nel caso di RagAB, il set di dati conteneva tre conformisti distinti che possono essere separati durante la classificazione 3D (Figura 5B). Le particelle che contribuiscono a ciascuna di queste classi possono quindi essere trattate in modo indipendente e utilizzate per perfezionare una mappa di densità EM che può quindi essere soggetta a ulteriori interpretazioni e costruzione di modelli.

Figura 5: Generazione di una mappa di densità EM 3D. A) Tipico modello iniziale generato utilizzando RELION. B) Classificazione 3D su 5 classi che mostrano la separazione delle particelle in tre distinti stati conformazionali: aperto-aperto (verde), aperto-chiuso (blu), chiuso-chiuso (viola). C) Processo di creazione della maschera. La mappa dal perfezionamento 3D (a sinistra) dovrebbe essere visualizzata in chimera. Il visualizzatore di volume può quindi essere utilizzato per identificare la soglia più bassa alla quale la mappa è priva di densità disarticolata e rumorosa (al centro). Questo valore di soglia viene immesso come soglia di binarizzazione iniziale nel processo di creazione della maschera RELION. Un esempio di output della maschera è mostrato in grigio (a destra). D) Mappa di densità EM ad alta risoluzione dello stato aperto-chiuso di RagAB (EMD-10245), filtrata e colorata per risoluzione locale (Å). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
In questo protocollo abbiamo descritto una pipeline di base applicabile ai campioni suscettibili di SPA di routine. Mentre questo metodo di blotting della carta da filtro per la formazione e la vetrificazione di film sottili ha indubbiamente successo dato il suo uso nella stragrande maggioranza dei progetti SPA fino ad oggi, presenta una serie di svantaggi. Questi includono lo spreco del campione, i tempi lenti (secondi) necessari per formare il film sottile e congelare il campione, l'irriproducibilità segnalata27 e gli effetti negativi segnalati dell'uso della carta da filtro per cancellare il liquido in eccesso50. Recentemente sono state sviluppate nuove tecnologie per migliorare la riproducibilità della produzione a film sottile51,52. Sono state sviluppate altre tecnologie che riducono il tempo tra l'applicazione del campione e la vetrificazione53,54,55. Mentre i metodi basati su carta da filtro per la formazione di film sottili rimangono il metodo più onnipresente di preparazione del campione SPA cryoEM al momento della scrittura, queste nuove tecnologie possono portare una serie di vantaggi in termini di efficienza e riproducibilità della vitrificazione della griglia, oltre a creare nuove opportunità per introdurre ulteriori dimensioni sperimentali, come la risoluzione temporale e la miscelazione rapida prima della vetrificazione.
Il processo di screening della griglia per la maggior parte degli utenti è attualmente un processo qualitativo che prevede l'acquisizione di atlanti a basso ingrandimento seguiti dall'acquisizione di immagini ad alto ingrandimento attraverso la griglia per valutare la distribuzione delle particelle. Mentre questo è un approccio sufficientemente robusto per alcuni tipi di campioni, può essere difficile valutare a occhio se il campione è effettivamente ciò che il ricercatore spera di immaginare o ha un orientamento preferito, ad esempio con campioni piccoli (<200 kDa) o dove la morfologia a bassa risoluzione rende difficile identificare ad occhio se è presente una gamma di distribuzioni di particelle. Per alcuni progetti, è impossibile determinare se il campione è come desiderato, ad esempio dove un ligando è legato o dove il campione viene sottoposto a screening per valutare se una piccola subunità (ad esempio, 10 kDa) è ancora presente in associazione con un complesso. Per questi progetti, pipeline completamente automatizzate per l'analisi dei dati combinate con raccolte "brevi" da 0,5 a 1 ora, che possono procedere attraverso fasi di elaborazione delle immagini fino alla classificazione 2D o persino alla classificazione e al perfezionamento 3D, aiuterebbero a determinare in modo efficiente se è necessaria una raccolta più lunga. Queste pipeline sono ancora in fase di sviluppo e non sono attualmente ampiamente implementate, ma hanno il potenziale per migliorare l'efficienza dello screening della griglia crioEM, specialmente per i campioni difficili.
I miglioramenti nei rivelatori di elettroni diretti, così come le modifiche nella microscopia combinate con i progressi nell'elaborazione delle immagini come la raccolta dei dati di spostamento delle immagini, hanno aumentato il throughput e la qualità delle immagini prodotte durante l'acquisizione dei dati. Questo aumento del tasso di raccolta dei dati evidenzia la necessità di uno screening approfondito delle griglie crioEM prima dell'acquisizione di molti TB di dati.
CryoEM SPA è diventata una tecnica di biologia strutturale veramente mainstream e in molti casi l'approccio "go to" per alcune classi di campioni, come complessi macromolecolari eterogenei e labili. Mentre il protocollo qui descrive una panoramica di base della pipeline SPA, ogni sezione trattata qui (vetrificazione e screening della griglia, crioEM ed elaborazione delle immagini) è un argomento a sé stante e degno di essere esplorato durante lo sviluppo di un progetto SPA. Man mano che le tecnologie di preparazione dei campioni e microscopia progrediscono e nuovi algoritmi e approcci di elaborazione delle immagini entrano in linea, SPA continuerà a svilupparsi come una pipeline, aiutando i ricercatori a ottenere informazioni su sistemi biologici complessi.
Divulgazioni
Non vengono segnalati conflitti di interesse.
Riconoscimenti
Questo lavoro è stato supportato dall'iNEXT-Discovery (Grant 871037) finanziato dal programma Horizon 2020 della Commissione Europea. J B. R. White è finanziato dal Wellcome Trust (215064/Z/18/Z). I microscopi FEI Titan Krios sono stati finanziati dall'Università di Leeds (premio UoL ABSL) e wellcome Trust (108466/Z/15/Z). Ringraziamo M Iadanza per l'uso del suo script di analisi micrografica. Riconosciamo Diamond Light Source per l'accesso e il supporto delle strutture crio-EM presso l'Electron Bio-imaging Centre (eBIC) nazionale del Regno Unito finanziato dal Wellcome Trust, MRC e BBRSC.
Materiali
| Name | Company | Catalog Number | Comments |
| Blunt tweezers | Agar Scientific | AGT5022 | |
| Cryo EM round storage box | Agar Scientific | AGG3736 | |
| CryoEM autogrid boxes | ThermoFisher Scientific | 1084591 | |
| CryoEM grids | Quantifoil | N1-C14nCu30-01 | |
| Ethane gas | Boc | 270595-F | |
| LN2 foam dewar | Agar Scientific | AG81760-500 | |
| LN2 storage dewar | Worthington industries | HC 34 | |
| Pipette | Gilson | 10082012 | |
| Pipette tips | Star labs | s1111-1706 | |
| Syringe | BD | BD 300869 | |
| Type II lab water | Suez | select fusion | |
| Vitrobot | ThermoFisher Scientific | 1086439 | |
| Vitrobot filter paper | Whatman | 1001-055 | |
| Vitrobot styrophome container assembly | ThermoFisher Scientific | 1086439 | |
| Vitrobot tweesers | ThermoFisher Scientific | 72882-D | |
| Software | |||
| EPU | ThermoFisher Scientific | 2.8.1.10REL | |
| TEM server | ThermoFisher Scientific | 6.15.3.22415REL | |
| Tia | ThermoFisher Scientific | 5.0.0.2896REL | |
| Titan krios microscope | ThermoFisher Scientific | Titan Krios G2 |
Riferimenti
- Kuehlbrandt, W. The Resolution Revolution. Science. 343 (6178), 1443-1444 (2014).
- McMullan, G., Faruqi, A. R., Henderson, R. Direct Electron Detectors. Methods in Enzymology. , (2016).
- Elmlund, D., Le, S. N., Elmlund, H. High-resolution cryo-EM: the nuts and bolts. Current Opinion in Structural Biology. , (2017).
- Lyumkis, D. Challenges and opportunities in cryo-EM single-particle analysis. Journal of Biological Chemistry. , (2019).
- Nakane, T., et al. Single-particle cryo-EM at atomic resolution. Nature. , (2020).
- Yip, K. M., Fischer, N., Paknia, E., Chari, A., Stark, H. Atomic-resolution protein structure determination by cryo-EM. Nature. , (2020).
- Conley, M. J., et al. Calicivirus VP2 forms a portal-like assembly following receptor engagement. Nature. 565 (7739), 377-381 (2019).
- Hesketh, E. L., et al. The 3.3 Å structure of a plant geminivirus using cryo-EM. Nature communications. 9 (1), 2369 (2018).
- Malone, L. A., et al. Cryo-EM structure of the spinach cytochrome b6 f complex at 3.6 A resolution. Nature. 575 (7783), 535-539 (2019).
- Madej, M., et al. Structural and functional insights into oligopeptide acquisition by the RagAB transporter from Porphyromonas gingivalis. Nature Microbiology. , (2020).
- Gallardo, R., et al. Fibril structures of diabetes-related amylin variants reveal a basis for surface-templated assembly. Nature Structural and Molecular Biology. , (2020).
- Scarff, C., et al. Structure of the shutdown state of myosin-2. Nature. , (2020).
- Scarff, C. A., et al. Structure of the protective nematode protease complex H-gal-GP and its conservation across roundworm parasites. PLoS Pathogens. 16 (4), 1008465 (2020).
- Wu, M., Lander, G. C. How low can we go? Structure determination of small biological complexes using single-particle cryo-EM. Current Opinion in Structural Biology. , (2020).
- Khoshouei, M., Radjainia, M., Baumeister, W., Danev, R. Cryo-EM structure of haemoglobin at 3.2 Å determined with the Volta phase plate. Nature Communications. , (2017).
- Drulyte, I., et al. Approaches to altering particle distributions in cryo-electron microscopy sample preparation. Acta crystallographica. Section D, Structural biology. 74, 560-571 (2018).
- Thompson, R. F., Walker, M., Siebert, C. A., Muench, S. P., Ranson, N. A. An introduction to sample preparation and imaging by cryo-electron microscopy for structural biology. Methods. 100, 3-15 (2016).
- Cheng, Y., Grigorieff, N., Penczek, P. A., Walz, T. A primer to single-particle cryo-electron microscopy. Cell. 161 (3), 438-449 (2015).
- Scarff, C. A., Fuller, M. J. G., Thompson, R. F., Iadanza, M. G. Variations on negative stain electron microscopy methods: tools for tackling challenging systems. Journal of Visualized Experiments. (132), e57199 (2018).
- Ohi, M., Li, Y., Cheng, Y., Walz, T. Negative Staining and Image Classification - Powerful Tools in Modern Electron Microscopy. Biological procedures online. 6, 23-34 (2004).
- Baker, L. A., Rubinstein, J. L. Radiation Damage in Electron Cryomicroscopy. Methods in enzymology. 481, 371-388 (2010).
- Dubochet, J., et al. Cryo-electron microscopy of vitrified specimens. Quarterly Reviews of Biophysics. 21 (02), 129-228 (1988).
- Passmore, L. A., Russo, C. J. Specimen Preparation for High-Resolution Cryo-EM. Methods in enzymology. 579, 51-86 (2016).
- Carragher, B., et al. Current outcomes when optimizing ‘standard’ sample preparation for single-particle cryo-EM. Journal of Microscopy. , (2019).
- Naydenova, K., Jia, P., Russo, C. J. Cryo-EM with sub-1 Å specimen movement. Science. , (2020).
- Passmore, L. A., Russo, C. J. Specimen Preparation for High-Resolution Cryo-EM. Methods in Enzymology. , (2016).
- Thompson, R. F., Iadanza, M. G., Hesketh, E. L., Rawson, S., Ranson, N. A. Collection, pre-processing and on-the-fly analysis of data for high-resolution, single-particle cryo-electron microscopy. Nature protocols. 14 (1), 100-118 (2019).
- Suloway, C., et al. Automated molecular microscopy: the new Leginon system. Journal of Structural Biology. 151 (1), 41-60 (2005).
- Zhang, J., et al. JADAS: A customizable automated data acquisition system and its application to ice-embedded single particles. Journal of Structural Biology. , (2009).
- Mastronarde, D. N. SerialEM: A program for automated tilt series acquisition on Tecnai microscopes using prediction of specimen position. Microscopy and Microanalysis. , (2003).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. , (2019).
- Fernandez-Leiro, R., Scheres, S. H. W. A pipeline approach to single-particle processing in RELION. Acta crystallographica. Section D, Structural biology. 73, 496-502 (2017).
- Gómez-Blanco, J., et al. Using Scipion for stream image processing at Cryo-EM facilities. Journal of Structural Biology. , (2018).
- Wagner, T., et al. SPHIRE-crYOLO is a fast and accurate fully automated particle picker for cryo-EM. Communications biology. 2 (1), 213-218 (2019).
- Bepler, T., et al. TOPAZ: A Positive-Unlabeled Convolutional Neural Network CryoEM Particle Picker that can Pick Any Size and Shape Particle. Microscopy and Microanalysis. , (2019).
- Zivanov, J., et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife. 7, 163 (2018).
- Zivanov, J., Nakane, T., Scheres, S. H. W. A Bayesian approach to beam-induced motion correction in cryo-EM single-particle analysis. IUCrJ. , (2019).
- Cianfrocco, M. A., Kellogg, E. H. What Could Go Wrong? A Practical Guide to Single-Particle Cryo-EM: From Biochemistry to Atomic Models. Journal of Chemical Information and Modeling. , (2020).
- Tagari, M., Newman, R., Chagoyen, M., Carazo, J. M., Henrick, K. New electron microscopy database and deposition system. Trends in Biochemical Sciences. , (2002).
- Berman, H. M., et al. The Protein Data Bank. Nucleic Acids Research. , (2000).
- Iudin, A., Korir, P. K., Salavert-Torres, J., Kleywegt, G. J., Patwardhan, A. EMPIAR: A public archive for raw electron microscopy image data. Nature Methods. , (2016).
- Punjani, A., Rubinstein, J. L., Fleet, D. J., Brubaker, M. A. CryoSPARC: Algorithms for rapid unsupervised cryo-EM structure determination. Nature Methods. , (2017).
- Tegunov, D., Cramer, P. Real-time cryo-electron microscopy data preprocessing with Warp. Nature Methods. , (2019).
- Goddard, T. D., et al. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Science. 27 (1), 14-25 (2018).
- Klebl, D. P., et al. Need for Speed: Examining Protein Behavior during CryoEM Grid Preparation at Different Timescales. Structure. , (2020).
- Noble, A. J., et al. Routine single particle CryoEM sample and grid characterization by tomography. eLife. 7, 32 (2018).
- Danev, R., Buijsse, B., Khoshouei, M., Plitzko, J. M., Baumeister, W. Volta potential phase plate for in-focus phase contrast transmission electron microscopy. Proceedings of the National Academy of Sciences. , (2014).
- Zi Tan, Y., et al. Addressing preferred specimen orientation in single-particle cryo-EMthrough tilting. Nature Methods. , (2017).
- Armstrong, M., Han, B. -. G., Gomez, S., Turner, J., Fletcher, D. A., Glaeser, R. M. Microscale Fluid Behavior during Cryo-EM Sample Blotting. Biophysical Journal. 118 (3), 708-719 (2020).
- Arnold, S. A., et al. Blotting-free and lossless cryo-electron microscopy grid preparation from nanoliter-sized protein samples and single-cell extracts. Journal of Structural Biology. , (2017).
- Dandey, V. P., et al. Spotiton: New Features and Applications. Journal of Structural Biology. , (2018).
- Rubinstein, J. L., et al. Shake-it-off: a simple ultrasonic cryo-EM specimen-preparation device. Acta crystallographica. Section D, Structural biology. 75, 1063-1070 (2019).
- Tan, Y. Z., Rubinstein, J. L. Through-grid wicking enables high-speed cryoEM specimen preparation. bioRxiv. , (2020).
- Klebl, D. P., et al. Sample deposition onto cryo-EM grids: From sprays to jets and back. Acta Crystallographica Section D: Structural Biology. , (2020).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati