
Aby wyświetlić tę treść, wymagana jest subskrypcja JoVE. Zaloguj się lub rozpocznij bezpłatny okres próbny.
Method Article
Analysis of Minerals Produced by hFOB 1.19 and Saos-2 Cells Using Transmission Electron Microscopy with Energy Dispersive X-ray Microanalysis
W tym Artykule
Podsumowanie
We present a protocol to compare the state of minerals in vesicles released by two human bone cell lines: hFOB 1.19 and Saos-2. Their mineralization profiles were analyzed by Alizarin Red-S (AR-S) staining, ultraviolet (UV) light visualization, transmission electron microscopy (TEM) imaging and energy dispersive X-ray microanalysis (EDX).
Streszczenie
This video presents the use of transmission electron microscopy with energy dispersive X-ray microanalysis (TEM-EDX) to compare the state of minerals in vesicles released by two human bone cell lines: hFOB 1.19 and Saos-2. These cell lines, after treatment with ascorbic acid (AA) and β-glycerophosphate (β-GP), undergo complete osteogenic transdifferentiation from proliferation to mineralization and produce matrix vesicles (MVs) that trigger apatite nucleation in the extracellular matrix (ECM).
Based on Alizarin Red-S (AR-S) staining and analysis of the composition of minerals in cell lysates using ultraviolet (UV) light or in vesicles using TEM imaging followed by EDX quantitation and ion mapping, we can infer that osteosarcoma Saos-2 and osteoblastic hFOB 1.19 cells reveal distinct mineralization profiles. Saos-2 cells mineralize more efficiently than hFOB 1.19 cells and produce larger mineral deposits that are not visible under UV light but are similar to hydroxyapatite (HA) in that they have more Ca and F substitutions.
The results obtained using these techniques allow us to conclude that the process of mineralization differs depending on the cell type. We propose that, at the cellular level, the origin and properties of vesicles predetermine the type of minerals.
Wprowadzenie
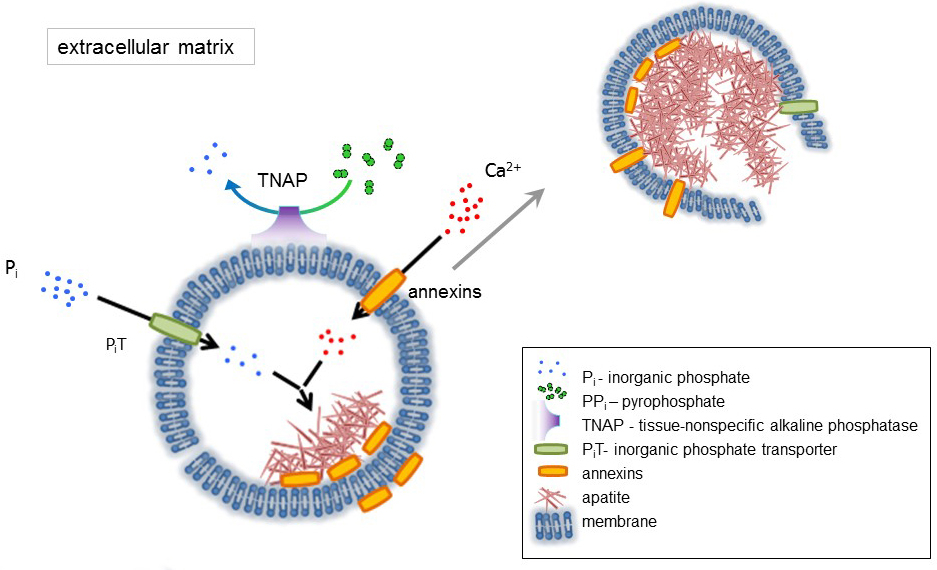
Bone is a type of connective tissue composed of two parts: organic (cells and collagen fibers) and mineral (calcium and phosphate compounds). The main mineral components in bones are apatites1. Different types of mineralization-competent cells in bone (osteoblasts), in teeth (odontoblasts) and in cartilage (chondrocytes) regulate the initial steps of mineralization by producing proteins of the extracellular matrix (ECM) and releasing matrix vesicles (MVs) (Figure 1). MVs are 100-300 nm diameter vesicles that accumulate calcium and phosphate facilitating apatite nucleation and subsequently bind to collagen2,3. Then, MVs disintegrate to release apatites to the extracellular medium. The apatites continue to grow in contact with collagen fibers and form the bone matrix. The mineralization is sustained by the constant supply of Pi and Ca2+ in the extracellular medium. Some recently published data support our model4,5. Soft tissues do not mineralize under physiological conditions. However, ectopic calcification may occur under pathological conditions such as vascular calcification3. Vascular cells that acquire the osteoblast phenotype can produce MVs that induce nucleation of apatites and initiate mineralization in the medial and intimal layers of the wall of blood vessels. Since ectopic calcification resemble normal endochondral mineralization3, understanding the molecular mechanisms of mineralization of osseous cells and chondrocytes should provide some clues on ectopic calcification of soft tissues that are formed.
The development of skeletal tissues is regulated by various enzymes, growth factors, and promoters or inhibitors of mineralization. The antagonistic action of tissue-nonspecific alkaline phosphatase (TNAP) (Figure 1) and ectonucleotide pyrophosphatase/phosphodiesterase I (NPP1), together with ankyrin (ANK), controls inorganic pyrophosphate (PPi) concentration6. PPi, a potent inhibitor of HA formation, is hydrolyzed by TNAP; NPP1 hydrolyzes nucleotide triphosphates to form PPi while ANK exports PPi from the cell to the ECM. The Pi/PPi ratio may regulate apatite formation7,8 with possible pathological consequences9.
The MV membrane is enriched in ion transport proteins that facilitate the initial precipitation of calcium and phosphate inside the MVs during the nucleation process (Figure 1). The phosphate transporter 1 (PiT) helps to incorporate Pi generated in the perivesicular space into the MVs10,11. Annexins may be involved in the binding and transport of Ca2+ and in the biophysical process that initiates mineralization in the MV lumen12,13. We favor the hypothesis, suggested earlier, for mineralization within intracytoplasmic vesicles of internal nucleation of apatite inside the MV before its propagation in the ECM14,15. In vitro modeling confirmed the induction of Ca2+/Pi complexes formation in proteoliposomes made from PS and AnxA516. This may indicate that accumulation of Ca2+, Pi, AnxA5 and PS complexes in lipid rafts of microvilli-like membranesrepresent the nucleation core (NC) of apatite within MVs. Annexins and TNAP also possess collagen-binding capacities that may be helpful in placing MVs along collagen fibers and, in stimulating the propagation of mineralization in the ECM. Fetuin A and osteopontin (OPN)17, are known as inhibitors of apatite formation that may slow down the propagation of mineralization on the collagenous scaffold. Nucleation and propagation are distinct events, the former preceding the latter, and both may be relevant for the process of pathological mineralization.
To discover how the chemistry of calcium phosphate complexes may change physiological mineralization and ectopic calcification, it is necessary to identify the minerals produced by cells. Apatites are a group of calcium and phosphate containing minerals with the general crystal unit cell formula Ca10(PO4)6X2, where X = Cl, F, OH. They are classified as follows18: fluorapatite (FA) Ca10(PO4)6F2, chlorapatite (CA) Ca10(PO4)6Cl2 and hydroxyapatite (HA) Ca10(PO4)6(OH)2.
The choice of osteoblast cell lines to induce mineral formation is crucial, since each cell line exhibits a distinct profile of mineralization. In this report, we compared the nucleation of minerals by two selected human cell models of mineralization: osteoblastic hFOB 1.19 cells and osteosarcoma Saos-2 cells. Osteosarcoma-derived cells are commonly used as osteoblastic models and Saos-2 cells have preserved the most mature osteoblastic character19 while undifferentiated human fetal hFOB cells are widely used as a model for normal osteoblastic differentiation20. Their mineralization profiles were analyzed by different methods: Alizarin Red-S (AR-S) staining, ultraviolet (UV) light visualization, transmission electron microscopy (TEM) imaging, energy dispersive X-ray microanalysis (EDX) quantitation, and ion mapping. The advantage of TEM-EDX over alternative techniques used in previous studies is that it gives quantitative and qualitative results of ion replacement in apatite crystals4,5,21. The overall goal of using TEM-EDX was to find a simple method for imaging and quantification of the distribution of Ca, F and Cl ions in various minerals from different types of cells during distinct stages of the mineralization process. This method has been successfully used, for example, for monitoring the interaction of zinc nanoparticles with coexisting chemicals and their combined effects on aquatic organisms22. In another study, a copper photocatalyst on titanium materials in aqueous solution was extensively characterized by means of inductively coupled plasma optical emission spectrometry (ICP-OES), N2 physisorption (BET), XRD, UV-vis DRS, FT-IR, Raman spectroscopy, TEM-EDX, and photoelectrochemical measurements23. Our aim was to compare the origin and properties of vesicles and minerals in two cell lines to understand the mechanism that controls mineralization during osseous differentiation.

Figure 1. Scheme of the initial steps of mineralization in osseous cells involving the synthesis of extracellular matrix (ECM) proteins and release of matrix vesicles (MVs) from the membrane. MVs accumulate calcium through the action of calcium binding proteins, annexins and phosphate, through the action of an inorganic phosphate transporter (PiT) followed by the activity of tissue non-specific alkaline phosphatase (TNAP), which dephosphorylates PPi to Pi, thereby facilitating apatite nucleation. Then, MVs disintegrate and release apatites to the extracellular medium. The mineralization is sustained by the constant supply of Pi and Ca2+ in the extracellular medium4,5. Please click here to view a larger version of this figure.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
Protokół
1. Cell Culture and Treatment
- Put all the necessary materials under the laminar flow hood and sterilize them under UV light. The culture media are: a 1:1 mixture of Ham's F12 and DMEM media with 2.5 mM L-glutamine supplemented with 100 U/mL penicillin, 100 U/mL streptomycin, 0.3 mg/mL G418 and 10% Fetal Bovine Serum (FBS) (v/v) for human fetus hFOB 1.19 SV40 large T antigen transfected osteoblasts, and McCoy's 5A medium with 1.5 mM L-glutamine supplemented with 100 U/mL penicillin, 100 U/mL streptomycin and 15% FBS (v/v) for human osteosarcoma Saos-2 cells.
- Culture hFOB 1.19 cells at 34 °C in an atmosphere of 5% CO2 and Saos-2 cells at 37 °C in an atmosphere of 5% CO2. Transfer cell cultures, either hFOB 1.19 or Saos-2 cells, from the incubator to the laminar flow hood and change the medium to 10 mL of fresh culture medium with FBS.
- Incubate cells without stimulators (resting cells) or stimulate them with 50 µg/mL Ascorbic Acid (AA) followed by 7.5 mM β-Glycerophosphate (β-GP), prepared earlier in ultrapure water and filtered through 0.22 µm syringe filters. Add the stimulators from plastic microcentrifuge tubes onto the surface of the culture medium. Gently stir the culture dish and incubate for 7 days.
2. Detection of Calcium Minerals
- Wash the cell cultures with Phosphate Buffer Saline (PBS: 125 mM NaCl, 5 mM KCl, 10 mM Na2HPO4, 1 mM KH2PO4, pH 7.0).
- Add 5 mL of 2% (g:100 mL) AR-S in PBS, pH 5.0 and incubate the plates for 30 min to stain the minerals.
- Wash 3 times with PBS. Carefully add PBS to the dish wall, try not to destroy the minerals.
- Observe calcium minerals under an inverted light microscope and take images.
3. Visualization of Probes under UV Light
- For cell lysates, treat cell cultures, either resting or stimulated for 7 days, according to the collagenase digestion protocol24.
- Collect the medium from cell cultures and wash the cells with PBS.
- Digest the cells with 3 mL of 500 U/mL collagenase in a solution of 0.25 M sucrose, 0.12 M NaCl, 0.01 M KCl, and 0.02 M Tris-HCl buffer, pH 7.45, at 37 °C for 3 h.
- Mechanically scrape the cells, transfer them to plastic microcentrifuge tubes, and pass them 10 times through a 1 mL 40 U syringe with 0.5 × 16 needle.
- Centrifuge the samples at 500 × g for 5 min.
- Discard the supernatant and suspend the cell pellet in 500 µL of Synthetic Cartilage Lymph (SCL, 100 mM NaCl, 12.7 mM KCl, 0.57 mM MgCl2, 1.83 mM NaHCO3, 0.57 mM NaH2PO4, 5.5 mM D-glucose, 63 mM sucrose, 16.5 mM Hepes, pH 7.4).
- Transfer the hydroxyapatite (HA), fluorapatite (FA) and chlorapatite (CA) powders from the bottles on the UV transilluminator with a spatula and use as controls.
- Transfer the cell lysates from the plastic tubes with plastic tips and place carefully on the UV transilluminator.
- Take images under visible and UV light.
4. Preparation of Probes for TEM-EDX
- Preparation of minerals for negative staining
- Suspend 2.5 mg of synthetically produced HA, CA and FA minerals25 in 500 µL of deionized water and incubate at 37 °C in an atmosphere of 5% CO2 for 1 h.
- Take Formvar/Carbon 300 Mesh Ni grids from the box with antistatic forceps, place on a porcelain multi-well plate and drop 10 µL of HA, CA and FA suspensions on the grids.
- Dry the samples for 30 min at room temperature.
- Preparation of resting and stimulated cells for embedding 26
- Collect medium from the cell cultures and wash the cells with Physiological Desensitization (PD) medium (125 mM NaCl, 5 mM KCL, 10 mM NaHCO3, 1 mM KH2PO4, 10 mM glucose, 20 mM HEPES, pH 7.4).
- Fix the cells with 5 mL of a mixture of 3% (g:100 mL) paraformaldehyde/1% (g:100 mL) glutaraldehyde in 100 mM sodium phosphate buffer, pH 7.2, for 1 h at room temperature under the fume hood.
- Wash the cells with 5 mL of 100 mM sodium phosphate buffer and gently remove the buffer after washing.
- In the dark room, postfix the samples with 2 mL of 1% (g:100 mL) osmium tetroxide in 100 mM sodium phosphate buffer, pH 7.2, for 20 min at room temperature under the fume hood.
- Remove osmium tetroxide and utilize it.
- Wash the cells with 5 mL of 100 mM sodium phosphate buffer.
- Then, dehydrate the samples in 5 mL aliquots of a graded ethanol solution series at room temperature: 25% (by vol.) for 5 min, 50% (by vol.) for 10 min, 75% (by vol.) for 15 min, 90% (by vol.) for 20 min. Finally use absolute ethanol twice and incubate for 30 min and 12 h.
- Mechanically scrape the cells from the plastic Petri culture dishes, collect into plastic microcentrifuge tubes and centrifuge the samples at 130 x g for 1 min.
- Remove the supernatants and suspend the cells in 1 mL of a mixture of the LR White resin and absolute ethanol at a volume ratio of 1:2.
- Mix well the content of the glass tubes before use and incubate for 30 min at room temperature.
- Centrifuge the samples at 130 x g for 1 min.
- Remove the supernatants and repeat the previous step using 1 mL of a 1:1 mixture of LR White resin and absolute ethanol.
- Mix well and incubate for 30 min at room temperature.
- Centrifuge the samples at 130 x g for 1 min.
- Remove the supernatants.
- Finally, add 1 mL of pure LR White resin to the samples twice and incubate for 1 h at room temperature in plastic tubes.
- Place 500 µL of each sample into gelatin capsules.
NOTE: The samples are labeled using a small sheet of paper and a pencil so that the resin does not destroy the labels. - Close the gelatin capsules, put them into plastic microcentrifuge tubes and centrifuge at 130 x g for 1 min in a swing-out rotor.
- Remove the capsules from the plastic tubes using a vacuum pump.
- Move the samples to the oven and polymerize at 56 °C for 48 h.
- Prepare the blocks by mounting them into the holder and trimming them to the pyramid.
- Put the holder into the ultramicrotome, attach the diamond Ultra 45° knife and fill it with deionized water; remember to clean the blade from incidental air bubbles.
- Then cut sections (700 Å) using the diamond knife onto the deionized water bath.
- Set the scraps using bovine eyelash and place them on the shiny side of the Formvar/Carbon 300 mesh Ni grid and dry them.
- Prepare 2.5% (by vol.) uranyl acetate in absolute ethanol in the dark under a fume hood. Keep the uranyl acetate in a lead container and remember not to pick up the sediment.
- In the dark room, counterstain the grids of synthetic apatites and cell samples with 2.5% (by vol.) uranyl acetate in ethanol for 20 min at room temperature under the fume hood.
- Wash the grids in 50% ethanol (by vol.), then in deionized water and dry at room temperature for 24 h. Finally, put the grids into the box.
5. TEM-EDX Analysis
- TEM imaging by a transmission electron microscope (TEM) equipped with full range Energy Dispersive X-ray microanalysis (EDX) System and 11 Megapixel camera
- Prepare a beryllium holder for the observation of minerals and cells. Use antistatic tools.
- Remove the pair of screws and lift the beryllium plate and beryllium washer away from the rest of the retainer.
- Mount the grid, shiny side up, on the holder.
- Carefully place the beryllium washer and beryllium plate and screw the fastening screws tightly.
- Put the holder into the vacuum chamber and turn on the vacuum pump.
- Once a vacuum is achieved, gently insert the holder into the imaging chamber and turn on the beam.
- On the fluorescent monitor, set the aperture parameters of the microscope. Perform image astigmatism correction; set the zoom, focus, and frame; and take TEM images at a magnification of 50,000X.
- STEM imaging and X-ray microanalysis for spectral and compositional analysis
- Insert the energy dispersive X-ray (EDX) detector into the scanning transmission electron Microscopy (STEM) imaging chamber.
- Adjust the sharpness of the image in the focus mode.
- Take STEM images at a magnification of 15,000X.
- Select a point in the sample for X-ray microanalysis and collect spectra.
- Obtain data by summing all atomic weights for all elements of the Periodic Table in the sample (as 100%) and signifying the content of selected elements: Ca, F, Cl, and P (as atomic%). Then, calculate the ratios of Ca, F or Cl to P for each sample.
- Ion mapping
- Select elements such as calcium, fluorine, chlorine and phosphorus to make ion mapping and perform EDX maps of the selected elements in the samples.
- Obtain data by indicating the localization of analyzed elements: Ca, F, Cl, and P (as atomic %) and calculate the co-localization (in %) of Ca, F or Cl with P for each sample.
Access restricted. Please log in or start a trial to view this content.
Wyniki
TEM-EDX allows for the in vitro imaging of matrix vesicles (MVs) released by mineralizing cells and of minerals produced by MVs. The results obtained using this technique demonstrate that the process of mineralization may proceed differently in various types of cells. The two cell lines received the same osteoblastic transdifferentiation treatment, yet stimulated Saos-2 cells mineralized more efficiently than hFOB 1.19 osteoblasts, as evidenced by AR-S staining (
Access restricted. Please log in or start a trial to view this content.
Dyskusje
In the current paper, we described the protocols for AR-S staining, UV light identification of fluorapatite and TEM-EDX in vitro imaging of MVs released by mineralizing cells and of minerals produced by MVs. It is possible to address all methods mentioned above by following some common troubleshooting steps. In order to obtain optimal results, several critical steps should be performed carefully. First, it is better to add AA (which is acidic) followed by β-GP (which is alkaline) to preserve the pH...
Access restricted. Please log in or start a trial to view this content.
Ujawnienia
The authors declare that they have no conflict of interest.
Podziękowania
MK and ASK performed manual operations and LB prepared drawings and made the movie. ASK wrote the manuscript, LB wrote the script and MK prepared the table. SM, RB and SP critically read the table, the script and the manuscript. The authors would like to thank Hanna Chomontowska for her excellent assistance with ultramicrotomy as well as Szymon Suski and Henryk Bilski for their excellent assistance with TEM-EDX analysis. The authors would like to thank dr Patrick Groves for professional English language correction and Barbara Sobiak for recording the instructions.
This work was supported by grant N N401 140639 from the Polish Ministry of Science and Higher Education to ASK, by grants from the National Science Centre, Poland 2016/23/N/NZ4/03313 to LB and 2016/23/N/NZ1/02449 to MK, EU FP7 Project BIOIMAGINE: BIO-IMAGing in research INnovation and Education, GA No. 264173, and by the statutory funds of the Nencki Institute of Experimental Biology, Polish Academy of Sciences.
Access restricted. Please log in or start a trial to view this content.
Materiały
| Name | Company | Catalog Number | Comments |
| Reagent | |||
| Ham’s DMEM/F12 media mixture | PAA | E15-813 | 1:1, for human fetus hFOB 1.19 SV40 large T antigen transfected osteoblasts (ATCC CRL-11372) |
| McCoy’s 5A medium | PAA | E82312-0025 | for human osteosarcoma Saos-2 cells (ATCC HTB-85) |
| Antibiotics mixture (penicillin/streptomycin) | Sigma | P0781-100ML | 100 U/mL each |
| G-418 | Sigma | 68168 | 0.3 mg/mL |
| FBS | Gibco | 10270 | 10% for hFOB 1.19 and 15% for Saos-2 |
| AA | Sigma | A-5960 | 50 µg/mL |
| ß-GP | Sigma | G9422-100G | 7.5 mM |
| Bio-Gel HTP Gel | Bio-Rad | 130-0420 | for HA |
| FA | synthesized by us | ||
| CA | synthesized by us | ||
| Sodium phosphate buffer Na2HPO4/NaH2PO4 mixture | Sigma | S7907/S8282 | 0.1 M, pH 7.2 |
| PBS | pH 7.0, prepared by us | ||
| AR-S in PBS | Sigma | A5533-25G | 0.5 g/100 mL, pH 5.0 |
| Collagenase type IA | Sigma | C2674 | 500 U/mL |
| SCL buffer | prepared by us | ||
| Deionized wather | produced by us | ||
| Ethanol | POCh | BA6480111 | absolut 99.8% and solutions 25, 50, 75, 90% |
| Uranyl acetate in 50% ethanol | Polysciences Inc. | 21447-25 | 0.25 g/10 mL |
| PD medium | pH 7.4, prepared by us | ||
| Fixation mixture (paraformaldehyde/glutaraldehyde) | Sigma | 158127/G-6257 | 3%:1% |
| Post-fixation OsO4 | Sigma | 75633 | 1% |
| LR White resin in ethanol | Polysciences Inc. | 17411-MUNC 500g | 1:2, 1:1, 100% |
| Acetone | CHEMPUR | 111024800 | pure |
| Tool | |||
| Cryogenic vials | Corning Inc. | 430487 | 1.2 mL |
| Plastic Petri culture dishes | Falcon | 353003 | 100 mm |
| Plastic tubes | Falcon | 352096 and 352070 | 15 and 50 mL |
| Serological pipettes | Falcon and VWR | 357521 and 612-3700 | 1 and 10 mL |
| Plastic microcentrifuge tubes | Sigma | Z688312 and Z628034 | 1.5 mL black and 2 mL transparent |
| Plastic tips | VWR | 613-0364, 613-0239 and 613-1050 | 0.1-10 µL natural, 1-200 µL yellow and 200-1000 µL blue |
| Plastic racks | Light Labs | A-7055-Z, A-7053-C | green for tubes, orange for micro tubes and blue for TEM probes |
| Laminar Hera Save | Thermo Scientific Co. | KS12 | HEPA filter (H14 according to DIN EN 1822) |
| Incubators Hera Cell | Thermo Scientific Co. | 150 | 34°C for hFOB 1.19 and 37°C for Saos-2 |
| Fume hood | POLON | WCS-2 | for TEM stainings |
| Glass bottles | SIMAX | 1632414501050 and 1632414501100 | 50 and 100 mL |
| Quartz glass tubes | SIMAX | 638422010100 | Ø 10 mm, L 100 mm |
| Pump | IBS Integra Biosciences | VACUSAFE comfort | for vacuum |
| Oven | Memmert | UNE 400 | 56°C |
| Porcelain multi-well plate | Rosenthal technik | 229/12 | 12 wells |
| Glass beakers | SIMAX | 632417010025 | 25 mL |
| Glass bottles | Pocord | DIN22 | 10 mL |
| Plastic box | Agar Scientific Ltd. | for darkness | |
| Snap Fit Gelatin Capsules | Agar Scientific Ltd. | G3741 | size 1 |
| Formvar/Carbon 300 Mesh Ni grids in box | Agar Scientific Ltd. | S162N3 | film on the shiny side |
| Silicon cell scraper | Sigma | SIAL0010-100EA | size 1.8/25 cm |
| Syringe with needle | BogMark | 007 | syringe 1 mL 40 U, needle 0.5 x 16 |
| Syringe | Chirana | CH005L | 5 mL |
| Centrifuge | MPW Medical Instruments | MPW-350R | 130 x g and 500 x g |
| UV transluminator | UVP | M-20 | for visible and UV light |
| Ultramicrotome | LKB | NOVA | 700Å sections |
| Block holder | LKB | E6711 | round shape |
| Diamond knife | DiATOME | Ultra 45° | size 3 |
| Eyelash holder | bovine, prepared by us | ||
| Forceps | ROTH | 2855.1 | antistatic for grids |
| Spatulas set | ROTH | E286.1 | antistatic for powders |
| Imaging | |||
| Inverted Light Microscope | Zeiss with Canon | AxioObserver Z1 equipped with PowerShot G9 | Phase contrast, Transmitted light, 20 x objective, RGB filters |
| Transmission Electron Microscope | TEM Jeol Co. with Oxford Instruments and SiS-Olympus | JEM-1400 TEM equipped with full range INCA Energy Dispersive X-ray microanalysis (EDX) System and 11 Megapixel MORADA G2 camera | magnification 50,000X for TEM and 15,000X for STEM and EDX |
| Camera body and lenses | Nikon | Nikon D7100 Nikkor AF Micro 105 mm f/2.8D Nikkor AF-S 50 mm f/1.8G Nikkor AF 28 mm f/2.8D | for movie recordings |
| Microphone | MXL Mics | Tempo | for voice recordings |
Odniesienia
- Buckwalter, J. A., Cooper, R. R. Bone structure and function. Instr. Course Lect. 36, 27-28 (1987).
- Anderson, H. C. Molecular biology of matrix vesicles. Clin. Orthop. Relat. Res. 314, 266-280 (1995).
- Anderson, H. C. Matrix vesicles and calcification. Curr Rheumatol. 5 (3), 222-226 (2003).
- Bolean, M., Simão, A. M. S., Barioni, M. B., Favarin, B. Z., Sebinelli, H. G., Veschi, E. A., Janku, T. A. B., Bottini, M., Hoylaerts, M. F., Itri, R., Millán, J. L., Ciancaglini, P. Biophysical aspects of biomineralization. Biophys Rev. 9 (5), 747-760 (2017).
- Bottini, M., Mebarek, S., Anderson, K. L., Strzelecka-Kiliszek, A., Bozycki, L., Simão, A. M. S., Bolean, M., Ciancaglini, P., Bandorowicz Pikula, J., Pikula, S., Magne, D., Volkmann, N., Hanein, D., Millán, J. L., Buchet, R. Matrix vesicles from chondrocytes and osteoblasts: Their biogenesis, properties, functions and biomimetic models. Biochim Biophys Acta. 1862 (3), 532-546 (2018).
- Hessle, L., Johnson, K. A., Anderson, H. C., Narisawa, S., Sali, A., Goding, J. W., Terkeltaub, R., Millan, J. L. Tissue-nonspecific alkaline phosphatase and plasma cell membrane glycoprotein-1 are central antagonistic regulators of bone mineralization. Proc. Natl. Acad. Sci. U. S. A. 99 (14), 9445-9449 (2002).
- Garimella, R., Bi, X., Anderson, H. C., Camacho, N. P. Nature of phosphate substrate as a major determinant of mineral type formed in matrix vesicle-mediated in vitro mineralization: An FTIR imaging study. Bone. 38 (6), 811-817 (2006).
- Thouverey, C., Bechkoff, G., Pikula, S., Buchet, R. Inorganic pyrophosphate as a regulator of hydroxyapatite or calcium pyrophosphate dihydrate mineral deposition by matrix vesicles. Osteoarthr. Cartil. 17, 64-72 (2009).
- Terkeltaub, R. A. Inorganic pyrophosphate generation and disposition in pathophysiology. Am. J. Phys. 281 (1), 1-11 (2001).
- Guicheux, J., Palmer, G., Shukunami, C., Hiraki, Y., Bonjour, J. P., Caverzasio, J. A novel in vitro culture system for analysis of functional role of phosphate transport in endochondral ossification. Bone. 27 (1), 69-74 (2000).
- Yadav, M. C., Bottini, M., Cory, E., Bhattacharya, K., Kuss, P., Narisawa, S., Sah, R. L., Beck, L., Fadeel, B., Farquharson, C., Millán, J. L. Skeletal mineralization deficits and impaired biogenesis and function of chondrocyte-derived matrix vesicles in Phospho1(-/-) and Phospho1/Pi t1 double-knockout mice. J. Bone Miner. Res. 31 (6), 1275-1286 (2016).
- Thouverey, C., Malinowska, A., Balcerzak, M., Strzelecka-Kiliszek, A., Buchet, R., Dadlez, M., Pikula, S. Proteomic characterization of biogenesis and functions of matrix vesicles released from mineralizing human osteoblast-like cells. J. Proteome. 74 (7), 1123-1134 (2011).
- Wang, W., Xu, J., Kirsch, T. Annexin-mediated Ca2+ influx regulates growth plate chondrocyte maturation and apoptosis. J. Biol. Chem. 278 (6), 3762-3769 (2003).
- Nollet, M., Santucci-Darmanin, S., Breuil, V., et al. Autophagy in osteoblasts is involved in mineralization and bone homeostasis. Autophagy. 10 (11), 1965-1977 (2014).
- Boonrungsiman, S., Gentleman, E., Carzaniga, R., Evans, N. D., McComb, D. W., Porter, A. E., Stevens, M. M. The role of intracellular calcium phosphate in osteoblast-mediated bone apatite formation. Proc. Natl. Acad. Sci. U. S. A. 109 (35), 14170-14175 (2012).
- Genge, B. R., Wu, L. N., Wuthier, R. E. In vitro modeling of matrix vesicle nucleation: synergistic stimulation of mineral formation by annexin A5 and phosphatidylserine. J. Biol. Chem. 282 (36), 26035-26045 (2007).
- Jahnen-Dechent, W., Schäfer, B., Ketteler, M., McKee, M. D. Mineral chaperones: a role for fetuin-A and osteopontin in the inhibition and regression of pathologic calcification. J. Mol. Med. (Berl). 86 (4), 379-389 (2008).
- Suchanek, W., Yoshimura, M. Processing and properties of hydroxyapatite-based biomaterials for use as hard tissue replacement implants. J. Miner. Res. 13 (1), 94-117 (1998).
- Pautke, C., Schieker, M., Tischer, T., Kolk, A., Neth, P., Mutschler, W., Milz, S. Characterization of osteosarcoma cell lines MG-63, Saos-2 and U-2 OS in comparison to human osteoblasts. Anticancer Res. 24 (6), 3743-3748 (2004).
- Yen, M. -L., Chien, C. -C., Chiu, I. -M., Huang, H. -I., Chen, Y. -C., Hu, H. -I., Yen, B. L. Multilineage differentiation and characterization of the human fetal osteoblastic 1.19 cell line: a possible in vitro model of human mesenchymal progenitors. Stem Cells. 25 (1), 125-131 (2007).
- Brittle, S. W., Foose, D. P., O'Neil, K. A., Sikon, J. M., Johnson, J. K., Stahler, A. C., Ryan, J. D., Higgins, S. R., Sizemore, I. E. A raman-based imaging method for characterizing the molecular adsorption and spatial distribution of silver nanoparticles to hydrated mineral surfaces. Environ Sci Technol. , (2018).
- Liu, N., Wang, Y., Ge, F., Liu, S., Xiao, H. Antagonistic effect of nano-ZnO and cetyltrimethyl ammonium chloride on the growth of Chlorella vulgaris: Dissolution and accumulation of nano-ZnO. Chemosphere. 196, 566-574 (2018).
- Tasbihi, M., Kočì, K., Troppová, I., Edelmannová, M., Reli, M., Čapek, L., Schomäcker, R. Photocatalytic reduction of carbon dioxide over Cu/TiO2 photocatalysts. Environ Sci Pollut Res Int. , (2017).
- Chen, N. X., O'Neill, K. D., Chen, X., Moe, S. M. Annexin-Mediated Matrix Vesicle Calcification in Vascular Smooth Muscle Cells. J. Bone Miner. Res. 23 (11), 1798-1805 (2008).
- Strzelecka-Kiliszek, A., Bozycki, L., Mebarek, S., Buchet, R., Pikula, S. Characteristics of minerals in vesicles produced by human osteoblasts hFOB 1.19 and osteosarcoma Saos-2 cells stimulated for mineralization. J. Inorg. Bioch. 171, 100-107 (2017).
- Thouverey, C., Strzelecka-Kiliszek, A., Balcerzak, M., Buchet, R., Pikula, S. Matrix vesicles originate from apical membranę microvilli of mineralizing osteoblast-like Saos-2 cells. J. Cell. Biochem. 106 (1), 127-138 (2009).
- Cazalbou, S., Eichert, D., Ranz, X., Drouet, C., Combes, C., Harmand, M. F., Rey, C. Ion exchanges in apatites for biomedical application. J. Mater. Sci. Mater. Med. 16 (5), 405-409 (2005).
- Kraus, D. Consolidated data analysis and presentation using an open-source add-in for the Microsoft Excel® spreadsheet software. Med. Writ. 23 (1), 25-28 (2014).
- Kawasaki, K., Buchanan, A. V., Weiss, K. M. Biomineralization in humans: making the hard choices in life. Annu. Rev. Genet. 43, 119-142 (2009).
- Bonucci, E. Bone mineralization. Front. Biosci. 17, 100-128 (2012).
- Veis, A., Dorvee, J. R. Biomineralization mechanisms: A new paradigm for crystal nucleation in organic matrices. Calcif. Tissue Int. 93 (4), 307-315 (2013).
- Nudelman, F., Lausch, A. J., Sommerdijk, N. A., Sone, E. D. In vitro models of collagen biomineralization. J. Struct. Biol. 183 (2), 258-269 (2013).
- Alliston, T. Biological regulation of bone quality. Curr. Osteoporos. Rep. 12 (3), 366-375 (2014).
- Wang, W., Kirsch, T. Retinoic acid stimulates annexin-mediated growth plate chondrocyte mineralization. J. Cell Biol. 157 (6), 1061-1069 (2002).
- Wang, W., Xu, J., Kirsh, T. Annexin V and terminal differentiation of growth plate chondrocytes. Exp. Cell Res. 305 (1), 156-165 (2005).
Access restricted. Please log in or start a trial to view this content.
Przedruki i uprawnienia
Zapytaj o uprawnienia na użycie tekstu lub obrazów z tego artykułu JoVE
Zapytaj o uprawnieniaThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Wszelkie prawa zastrzeżone