
Method Article
Formação espontânea e rearranjo das redes de nanotubo lipídica Artificial como um modelo de baixo para cima para o retículo endoplasmático
Neste Artigo
Resumo
Com suporte sólido, isento de proteína, dobro fosfolipídeo BICAMADA membranas (DLBM) podem ser transformadas em redes de nanotubo de lipídios complexos e dinâmicos e podem servir como modelos 2D de baixo para cima do retículo endoplasmático.
Resumo
Apresentamos um método conveniente para formar um modelo estrutural organela ascendente para o retículo endoplasmático (ER). O modelo consiste de nanotubos lipídico altamente densos que são, em termos de morfologia e dinâmica, uma reminiscência de ER. As redes são derivadas de patches de membrana dupla BICAMADA fosfolipídicas aderindo ao substrato transparente Al2O3 . A adesão é mediada por Ca2 + no buffer de ambiente. Subsequente depleção de Ca2 + através de BAPTA/EDTA causa retração da membrana, resultando na formação de rede de nanotubo de lipídios espontânea. O método só é composto por fosfolipídios e microfabricated superfícies simples formação de um modelo ER e não requer a adição de proteínas ou energia química (por exemplo, GTP ou ATP). Em contraste com a morfologia 3D do retículo endoplasmático celular, o modelo é bidimensional (embora o nanotubo dimensões, geometria, estrutura e dinâmica é mantida). Este modelo exclusivo em vitro ER consiste apenas de alguns componentes, é fácil de construir e pode ser observada sob um microscópio de luz. A estrutura resultante pode ser ainda mais decorada para obter funcionalidades adicionais, tais como a adição de proteínas associadas a ER ou partículas para estudar os fenômenos de transporte entre os tubos. As redes artificiais descritas aqui são modelos estruturais adequados para o celular de ER, cuja morfologia característica única mostrou estar relacionada com a sua função biológica, Considerando que detalhes sobre a formação do domínio tubular e rearranjos dentro são ainda não completamente entendidos. Notamos que esse método usa o Al2O3 lamelas microscopia fino-revestido por película, que estão comercialmente disponíveis mas exigem encomendas especiais. Portanto, é aconselhável ter acesso a uma instalação microfabrication para preparação.
Introdução
O ER executa tarefas cruciais na célula biológica, incluindo o enrolamento de proteínas, síntese de lipídios e cálcio Regulamento1,2. A morfologia do ER é intrínseca para as funções que ele executa. Ele combina pilhas planares e domínios tubulares denso-dinâmico, que continuamente interagem com o citoesqueleto e sofrem rearranjo e movimento constante. Alguns da remodelação que se submeter a estruturas ER incluem a transformação contínua entre folhas planas e tubos, formação de vesículas de ou fusão ao lúmen do ER, alongamento dos tubos pré-existentes, retração de tubo, fusão e ruptura3. A estrutura peculiar das redes tubulares é energeticamente desfavorável. Os caminhos e os mecanismos pelos quais o ER gera e mantém esta organização, bem como a forma como isso se relaciona com a sua função não é ainda totalmente compreendido4,5.
É sabido que o ER avarias quando perde seu estado homeostático, resultando em estresse de ER, uma condição causada por um aumento na síntese proteica, acúmulo de misfolded proteínas ou mudanças de Ca2 + e equilíbrio oxidativo. ER stress por sua vez provoca a deformação da morfologia natural da organela, especificamente, perturbar a organização de rede6,7. Como resposta, a célula ativa um mecanismo de reparação para retornar a um estado homeostático. Falha no reparo pode levar à apoptose celular induzida por ER, que contribui para várias doenças metabólicas e degenerativas como a doença de Alzheimer, diabetes tipo 2, doença de Parkinson, esclerose lateral amiotrófica e vários outros7, 8. Pesquisa atual visa a organização das redes tubulares ER e vários estudos estão se concentrando em reconstituir o ER em vitro2. Existindo alguns modelos2,9,10 necessitam de proteínas para iniciar e manter a membrana curvatura3,11 e ajudar a organela atingir sua forma. Claramente, os sistemas modelo que espelham alguns dos principais recursos estruturais e organizacionais do PS e fornecem acesso a avançados estudos experimentais estão em grande demanda.
Apresentamos aqui os procedimentos para a preparação de um facile, proteína química/energia livre, dinâmico em vitro modelo para o ER, fornecendo uma plataforma básica para estudar a morfologia de ER e funções associadas4. Neste método, um modelo ER é fabricado com uma abordagem de baixo para cima, usando apenas alguns elementos, em que as moléculas de interesse podem ser integradas para adicionar complexidade. A rede representa a dinâmica e a estrutura de ER. Além disso, transformação reversível entre a membrana planar e os tubos, formação de vesículas de tubos, tubo de fusão, deslizando e retração todos observam. Além de servir como um modelo de baixo para cima para o ER celular incompletamente compreendido, a rota de lipídios para redes de nanotubos descritos no presente protocolo pode ser aplicável para pesquisadores estudando auto-montagem, nanofluidics, único-molécula e coloide fenômenos de transporte, fluxo de Marangoni e outros campos relacionados. Os blocos de construção moleculares somente usados em nosso método são fosfolipídios. O protocolo exige laboratório pouco trabalho e equipamento básico e está acessível para a incorporação de elementos adicionais.
Protocolo
1. preparação da suspensão de vesículas fosfolipídicas
Nota: Para todos os materiais referidos como "limpa" no presente protocolo, completamente lavá-los com isopropanol, seguido de água desionizada e seque-os com nitrogênio. Observe que um tratamento de substratos de vidro com fortemente oxidantes agentes ácidos (solução de Piranha), que é normalmente aplicado em protocolos de preparação para filmes com suporte lipídico em substratos sólidos, não deve ser realizado em Al2O3-revestido porta-aviões.
- Para um balão de fundo redondo ou invertido pera vidro limpo 10ml: soja L-α fosfatidil colina (PC, 69% w/w), 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine (droga, 30% w/w) e um fluoróforo lipídios conjugados de escolha [por exemplo, Texas Red 1,2 - dihexadecanoyl-sn-glicero-3-phosphoethanolamine trietilamônio sal (TR-DHPE, 1% w/w)] no clorofórmio; para uma quantidade total de 3000 µ g de lipídeos em 300 µ l de clorofórmio, resultando em uma concentração final de 10 mg/mL.
Nota: Use limpos, vidro, seringas hermética com êmbolos de politetrafluoroetileno ao manusear compostos contendo clorofórmio.
Cuidado: Clorofórmio é tóxico e altamente volátil e deve sempre ser tratado sob uma coifa com equipamento de proteção pessoal associado. - Ligar o balão ao evaporador rotativo, posição com uma inclinação de 45° e gire lentamente a 24 rpm dentro de um banho de água a 23 ° C, durante 6 h com pressão de ar reduzida para remover completamente o clorofórmio. Começa a reduzir o direito de pressão após iniciar a rotação por etapas de 20 kPa cada 2 min até atingir 20 kPa (150 Torr, 80% de vácuo).
Nota: A formação de um filme lipídico homogênea de espessura uniforme no recipiente de preparação é o requisito mais importante para o procedimento de rotavap. Preparações de lipídios são sensíveis à velocidade de rotação, mudanças rápidas de pressão e pressão final valor; Portanto, segui rigorosamente as etapas de redução lenta, bem como a velocidade final de pressão e rotação. Posicione o balão com uma inclinação de 45° para garantir que o bolo de lipídios desidratado é formado uniformemente como um filme na parede do frasco. Muito rápido de uma rotação leva a turbulências e lenta de uma rotação leva (devido à gravidade) à acumulação de uma espessa camada de fluido na parte inferior do frasco. Durante o pernoite subsequente inchaço processo, um lipídio muito inhomogenous é que não responde bem à etapa final sonication produzidos em massa, e as frações resultantes são de composições diferentes. Pressão e tempo dentro do intervalo especificado no método garante desolvation lento. Com clorofórmio como solvente, muito rápida de uma queda na pressão esfria a mistura, resultando em aumento da viscosidade e formação de filme agregada e desigual. Recomenda-se o período de tempo de 6 h a fim de remover o solvente na maior medida possível, como as partições de solventes orgânicas no material lipídico após reidratação. - Após 6 h, parar a rotação e aumentar a pressão de ar novamente, aos poucos, em etapas de 20 kPa cada 2 min até atingir 100 kPa. Retirar o frasco do evaporador rotativo e adicionar 3 mL de PBS e 30 µ l de glicerol. Agite suavemente o frasco para dissolver o glicerol. Use uma rolha de vidro hermético para selar o frasco contendo os lipídios.
Nota: O glicerol é usado para prevenir a desidratação completa do filme lipídico e permite a separação de BICAMADA12. Ele deve ser aquecido antes do uso para diminuir sua viscosidade, que facilita o manuseio deste composto. O glicerol aquecido ainda não misture com o tampão PBS. Rodando suave é necessária até que o glicerol é completamente dissolvido. - Armazene o frasco na geladeira a 4 ° C durante a noite para reidratação e inchaço dos filmes de lipídios.
- No dia seguinte, proceda à sonicação lipídeos com um banho ultra-sônico de água à temperatura ambiente (RT, ~ 21 ° C) e a frequência de 35 kHz até alcançar uma suspensão lipídico uniforme, ligeiramente turva.
Nota: Sonication pode demorar em torno de 10-30 s. prolongada sonication (~ 1 min) produz calor e é prejudicial para a formação de vesículas. - Passos 1.1-1.5 rendimento uma suspensão contendo dois tipos de estruturas vesiculares: vesículas multilamellar (MLV) e vesículas unilamellar gigante (GUV) (Figura 1A-1F).
- Para armazenamento, divida a suspensão lipídico em 100 alíquotas µ l, usando um total de 30 tubos microcentrifuga e armazená-los em um congelador a-20 ° C.
Nota: Flash congelamento com nitrogênio líquido não é necessário e não é usado antes do armazenamento. O protocolo pode ser pausado aqui. Deixando as suspensões de lipídios na geladeira a 4 ° C para prolongada vezes causas lipídico Lise, que afeta a composição da membrana.
2. preparação de substratos
Nota: O seguinte protocolo é realizado em uma sala limpa classificada como ISO 8 na especificação padrão ISO 14644-1. Deposição de camada atômica (ALD) é usada para fabricar Al2O3 substratos. Os parâmetros de processo especificado são dependentes do instrumento e podem variar entre diferentes modelos de equipamentos. Eles podem ser usados como parâmetros iniciais para desenvolver o processo.
- Regule a temperatura do reator ALD a 200 ° C.
- Carrega as superfícies de vidro (por exemplo, as lamelas de vidro) na câmara de amostra juntamente com uma bolacha de silicone, que será usada mais tarde como uma superfície de referência para determinar a espessura da deposição por elipsometria.
Nota: Substratos de vidro foram usados para fora-de-caixa e não foram limpos de solvente antes da deposição. Eles só foram liberados com gás de nitrogênio para remover partículas. - Evacuar a câmara de carga para 400 Pa (3 torr) e transferir as amostras para a câmara de reação principal e evacuar a 200 PA.
Nota: A temperatura do reator deve ser mantida a 200 ° C para deposição adequada. Flutuações de temperatura após o carregamento da amostra deve ser equilibrado, portanto, antes de iniciar a deposição. Pressão da câmara é definido para ser superior a pressão do reator para evitar qualquer precursores que se espalhou para fora da câmara. - Começa a depositar o filme atômico. Um ciclo consiste de uma exposição alumínio 150 ms pulso trimetil, seguida por um 1 expurgo de s e posteriormente, uma exposição de2O H de duração de 200 ms, seguida por um 1 s expurgo.
Nota: Todas as configurações, incluindo a câmara e o reator de pressão, comprimento dos ciclos e expurgos são automatizados para alcançar uma taxa definida de deposição. Esses parâmetros podem variar entre os vários modelos de equipamentos. As receitas pré-configurado, muitas vezes, são preparadas pelo fornecedor ou ferramenta responsável em sala limpa e comunicadas ao usuário, como a espessura de película depositada por unidade de tempo. - Para chegar a 10 nm de Al2O3 no substrato, repita o processo para 100 ciclos. O número de ciclos depende da taxa de deposição, que pode variar entre diferentes receitas ou equipamento.
- Para remover as amostras do reator, primeira ventilação da câmara até que sua pressão atingir a pressão atmosférica, em seguida, retire as amostras.
- Armazene as amostras em recipientes de ar comprimido em RT até o uso.
Nota: Não mais limpeza é recomendado antes da utilização. O protocolo pode ser pausado aqui.
Nota: As amostras idealmente devem ser usadas imediatamente após a deposição. O armazenamento ideal exige posicionamento as superfícies dentro portadores de polipropileno da bolacha, seguidos por envolvendo as transportadoras em sala limpa-compatível com sacos de plástico, que são para ser nitrogênio liberado antes da selagem do vácuo. O objetivo é evitar expor a superfície para contaminantes pelo ar. Se necessário, as superfícies podem ser mantidas em recipientes de ar comprimido em RT no máximo por 5 dias. Armazenamento mais longo não é recomendado. Para usuários que não têm fácil acesso a uma sala limpa nas proximidades e comprar ou obter as superfícies do exterior, re-oxidação os substratos através de tratamento com oxigênio ozônio ou plasma pode ser uma solução alternativa13.
3. transformação de filmes Phospholipid Molecular para redes tubulares
- Descongelar a suspensão lipídico e transferir 4 gotas µ l da suspensão para uma slide de microscópio limpa vidro/lamela.
- Desidratar a gota por 20 min. O droplet entrará em colapso em uma película plana circular de lipídios após a dessecação, que é visível ao olho.
- Re-hidratar o filme lipídico com 1 mL de tampão HEPES (ver Tabela de materiais) por 3 min.
Nota: O volume de reidratação tampão afeta a densidade de suspensão de vesículas (o número de vesículas por unidade de volume), que posteriormente é transferido para a câmara de observação. Dependendo do volume da câmara de observação e a densidade desejada vesículas, o volume de reidratação pode ser ajustado para 0,5-1 mL. Slides de borosilicato limpo tendem a apoiar as gotas de vários cem microlitros até 1,5 mL sem problemas. Desde que o lamela não precisa ser movido, isso não implica que um problema técnico. Em superfícies mais hidrofóbicas, como SU-8 slides de polímero-revestido, até 1,5 mL pode ser depositado12.
Nota: Lipídios devem ser preparados na hora, como a exposição do filme lipídico re-hidratado por mais de 20 min no RT leva a evaporação do buffer e desidratação parcial das vesículas anteriormente re-hidratadas, que leva a um mal definida composição. - Preparar a câmara de observação: para permitir a troca de reserva por meio de uma pipeta automática, que é necessário para iniciar a transformação de ER, utilizou-se uma câmara de observação com uma parte superior aberta. Esta câmara consiste em um frame do polydimethylsiloxane (PDMS) com dimensões de 1,5 x 1,5 x 0,5 cm, aderiu em Al2O3 depositado lamela. Um esquema da câmara de observação montado é representado na Figura 1G. As etapas a seguir foram realizadas para fabricar o quadro PDMS e montar a câmara de observação:
- Prepare uma solução KOH misturando-se 100 g de KOH com 100 mL de isopropanol em um béquer em um banho de gelo. Mexa por 10 h ou mais, até que o KOH é dissolvido completamente, utilizando um agitador magnético e placa de agitação magnética.
Cuidado: Solução KOH é corrosiva e pode levar a queimaduras de pele, então sempre lidar com equipamento de proteção pessoal apropriado.
Nota: A solubilidade do KOH em isopropanol não é tão elevada como na água. A dissolução é exotérmica. Esmagar as pelotas KOH antes da dissolução e agitação contínua é aconselhável. - Mergulhe um placa de Petri de vidro (d = 6 cm) para a solução KOH no RT e mantê-lo durante a noite.
- No dia seguinte, retire o prato de vidro da solução, mergulhe-a em um recipiente com água desionizada para 5 min, enxágue várias vezes com água e coloque-a dentro de um forno de secagem a 80 ° C, durante 1 h. golpe a superfície brevemente com um fluxo de nitrogênio para garantir essa parte Icles são removidos.
- Para passivate da superfície e impedir a adesão de PDMS, transferi 200 µ l de dimetildiclorossilano com uma seringa de plástico em um recipiente de plástico limpo como um barco de ponderação.
- Armazene o vidro de Petri juntamente com silano por 1h em um dessecador evacuado (baixo vácuo, ~ 20 kPa).
Cuidado: Dimetildiclorossilano é tóxico e deve sempre ser tratado sob uma coifa com equipamento de proteção pessoal associado. - Espere 15 min antes de coletar o prato de Petri para que o vapor de dimetildiclorossilano restantes se dissipar. O prato de Petri é agora silanizadas, e a superfície é hidrofóbica.
Nota: Uma maneira rápida para testar o sucesso desta etapa é colocar uma gota de água sobre a prato de Petri silanizadas. O ângulo de contato da gota com a superfície visivelmente deve aumentar em relação ao vidro não tratado. - Em um recipiente plástico de 250 mL (transparente plástico copo fresco do pacote), misture 10 g de base de elastômero de silicone com 1 g de silicone elastômero do agente de cura (10:1). Misture usando uma espátula de plástico agitador/plástico por 5 min.
Nota: As bolhas de ar são formadas em cima de agitação, e o PDMS vai olhar pálido-branco. - Seque completamente a mistura para desgaseificar a < 20 kPa até todas as bolhas de ar em expansão tem desmoronado (maior vácuo acelera o processo). Despeje a mistura libertos na prato de Petri silanizadas.
- Curar a 65 ° C por 2 h no forno.
Nota: É possível dobrar a velocidade de cura, aumentando a temperatura > 95 ° c. A temperatura de cura crescente resulta em um aumento na rigidez do material. - Arrefecer a placa de Petri preenchida com o PDMS curado para RT e remover a laje PDMS com uma espátula.
- Com um bisturi, corte o quadro nas dimensões e geometria apropriada para a abertura disponível na fase de microscópio. Dimensões de 1,5 (comprimento) x 1,5 (largura) x 0,5 cm (altura) são adequadas para a maioria das configurações.
- Trazer o lado liso (lado inferior que estava em contato com o prato de Petri) do PDMS frame em contacto com o lado ativo da superfície onde reside o Al2O3 filme e suavemente aplique pressão para empurrar o quadro e de superfície contra o outro para fazê-los aderir.
Nota: A adesão entre o quadro PDMS e Al2O3 substrato é fraca. A presença de bolhas de ar na interface de contato pode causar de fixação e, consequentemente, o escapamento do tampão e conteúdo relacionado. O quadro PDMS pode ser usado várias vezes, se imediatamente após e antes de cada uso, ele é lavado com isopropanol, seguido por lavagem com água e secar com nitrogênio. A prato de Petri silanizada também pode ser reutilizada.
- Prepare uma solução KOH misturando-se 100 g de KOH com 100 mL de isopropanol em um béquer em um banho de gelo. Mexa por 10 h ou mais, até que o KOH é dissolvido completamente, utilizando um agitador magnético e placa de agitação magnética.
- Encha a câmara de observação com Ca2 +- HEPES buffer (ver Tabela de materiais).
Nota: A superfície deve ser usada imediatamente após a retirada do selo do pacote. Contacto com o ar leva a adsorção de contaminantes, o que diminui a atividade da superfície gradualmente. A câmara deve ser preenchida com buffer imediatamente após a montagem. Não encha o volume da câmara inteira para permitir a adição de lipídeos re-hidratados na etapa subsequente. - Coloque a câmara no palco do microscópio confocal. Transferi o material lipídico rehidratado, agora uma suspensão contendo vesículas gigantes, para a câmara com uma pipeta Pasteur (Figura 1A-1 G) de plástico.
- Espere 10-20 min para deixar as vesículas aderir sobre o substrato e se espalhou por toda a superfície (Figura 1H-1J).
Nota: A divulgação começa imediatamente após a deposição de lipídios na superfície. A taxa de propagação pode variar ligeiramente dependendo da composição lipídica, a técnica de deposição de Al2O3 (ADL, depósito de vapor RF-sputtering, química, etc.), frescura do substrato e concentração de cátion divalente na Reserva. Certifique-se de que a troca de reserva é executada antes de romper a repartição dos patches14. - Depois de observar várias propagações de lipídios, remova lentamente o buffer de ambiente através de uma pipeta automática, tal que só um filme fino reserva permanece na parte inferior.
Nota: A rápida remoção do buffer perturba as estruturas lipídicas na superfície. - Proceder à troca ambiente reserva, lentamente, preenchendo a câmara de observação com tampão HEPES-quelante (ver Tabela de materiais) usando uma pipeta automática (Figura 1-K).
Nota: A adição abrupta do buffer perturba as estruturas lipídicas na superfície. - Esta etapa final produz redes nanotubular dinâmico, formadas como resultado do quelante-induzida depinning e retração do DLBM para o MLV4 (Figura 1L-1Y).
4. microscopia observação
- Adquirir as imagens com um laser invertido microscópio confocal usando um 40 X objetivo de imersão de óleo (NA 1.3) com uma frequência de varredura de 400 Hz. empregar uma fonte de laser de luz branca para excitar o Texas Red DHPE em 595 nm. Coletar a emissão de 605 a 700 nm, usando um detector de fótons híbrido.
Nota: Como alternativa, um microscópio de fluorescência-epi pode ser usado para a imagem latente. Dependendo das fontes de luz disponíveis, selecione um apropriado lipid-tingem o conjugado.
Resultados
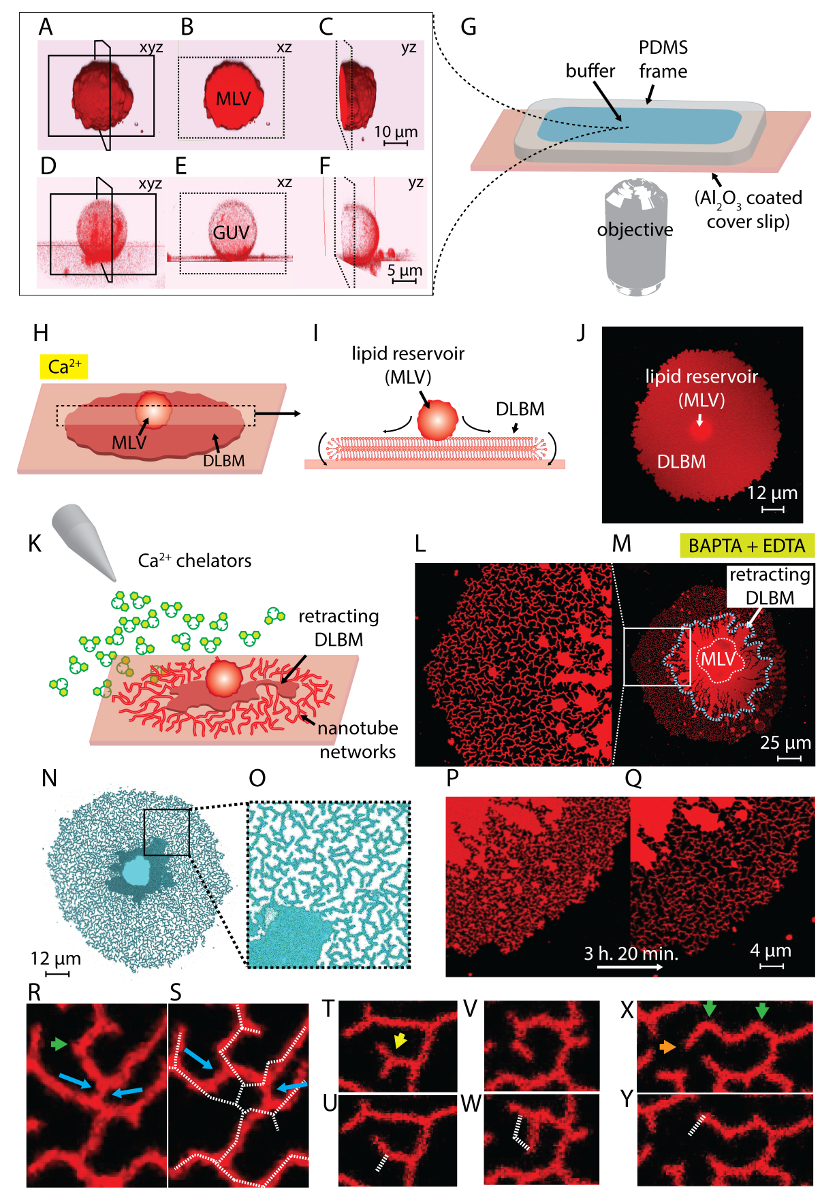
A suspensão lipídico obtido na etapa 1 do protocolo e usado durante os experimentos contém dois tipos principais de vesículas: MLVs e GUVs. Figura 1 A-1F mostra micrografias confocal das vesículas na amostra inicial construído em 3D de exploração do laser. Figura 1 A-1_C mostra um MLV (depósito de lipídios) no xyz, xz e yz aviões, respectivamente. Figura 1 D-1F mostra opiniões semelhantes de uma vesícula unilamellar gigante (chefe). A parte interna dos GUVs, que falta de multilamellarity, é oca; Portanto, o material lipídico para espalhar é significativamente limitado. Portanto, os reservatórios de lipídios só é útil para este método são MLVs.
Quando a suspensão lipídico é transferida para a câmara de observação contendo o Ca2 +- tampão HEPES (Figura 1G), os MLVs (Figura 1A-1_C) começam a acertar na superfície Al2O3 . Em caso de contacto, as vesículas aderem à superfície, e uma membrana de bicamada lipídica plana duplo circular (DLBM) começa a se espalhar de cada MLV sobre o suporte sólido (Figura 1H-1J). O MLV age como um reservatório que fornece os lipídios para a DLBM continuamente em expansão. A membrana de BICAMADA (superior) distal, no que diz respeito ao apoio sólido, está ligada à membrana de BICAMADA proximal (inferior) ao longo da circunferência da borda circular, realizando um movimento de rolamento (Figura 1eu). Os dois bilayers são posicionados categoricamente em cima do outro, tendo apenas uma fina película líquida encapsulada entre eles. Durante a propagação, a membrana proximal continuamente adere à superfície de apoio por baixo, enquanto a membrana distal é puxada lateralmente nas bordas pela borda proximal da membrana em expansão. A difusão da DLBM é mediada por Ca2 +, que atua como um agente de fusogenic entre grupos de cabeça de lipídios de membrana proximal e substrato sólido4.
Se a propagação continua por períodos prolongados, a membrana de tensão aumenta, levando a rupturas (Figura 2 e 2B). Após esse ponto, retração de membrana, que é necessária para criar a morfologia tubular ER, já não pode ser induzida. Portanto, é importante reconhecer as ruptura de membranas. Desde que os lipídios em nossos experimentos fluorescente são rotulados, a ruptura pode ser diretamente observado. Um indicador-chave de ruptura é a queda significativa na intensidade de fluorescência na região de ruptura (Figura 2 e 2B)14. A ruptura é um resultado do aumento da tensão e formação subsequente dos poros na membrana distal. Sob um microscópio fluorescente, as regiões de ruptura, portanto, irão expor metade a emissão intensidade (única BICAMADA proximal) das regiões unruptured (dupla BICAMADA)14 (Figura 2B). Durante a formação de poros, o material da membrana, que foi inicialmente posicionado nas regiões rompido, migra para as bordas do patch do espalhamento. Este por sua vez provoca um crescimento da área global de remendo. Portanto, uma expansão rápida do contorno do patch circular também pode ser observada durante a ruptura.
Para evitar o crescimento extensivo, levando a ruptura dos patches, imediatamente após a área de remendo circular atinge 100-200 µm, Ca2 +- HEPES buffer é removido suavemente com uma pipeta automática até uma fina película de líquido permanece na superfície. O tampão HEPES-quelante suavemente, em seguida, é adicionado à câmara para iniciar a retração (Figura 1-K). Uma desidratação completa da amostra (Figura 2C) ou troca rápida de buffers (Figura 2D e 2E) faz com que a perturbação, ruptura ou deformação dos patches. A adição de quelantes gradualmente remove Ca2 + o espaço entre a superfície e a membrana. O tampão HEPES-quelante gradualmente acessa o espaço inter-bilayer-substrato, a partir da periferia do remendo da membrana lipídica. Portanto, a remoção dos sites fixação começa a partir das bordas do patch circular e se propaga para o interior (Figura 1K-1T). Como resultado de depinning, a membrana lipídica começa a separar e retrai-se desde o interior de bordas, progredindo em direção o MLV no centro. O processo de retração conduz a uma nova interface de dinamicamente desenvolver redes tubulares lipídico4 (Figura 1L-1Y). As regiões persistentes da fixação, que não permitem a membrana separar completamente, permanecem sobre a superfície e nucleada tabulação, levando a longa cadeia ramificada rede de nanotubos. (Figura 1L-1Y). Contínua de fixação e mais retração de lipídios nanotubos são observados ao longo do tempo como resultado do processo de quelação gradual. Esta aspereza e rearranjo dos ramos rede desempenha um papel fundamental no comportamento dinâmico das redes tubulares, que se assemelham o re liso.
Figura 1 L-1Y mostra as micrografias das redes nanotubo obtidas no protocolo. Figura 1 L é que um close-up da região na Figura 1M marcado no quadro branco. As regiões de vermelho-contínuas na Figura 1L e 1 M são a fração de retracção da DLBM (marcado com uma linha tracejada azul na Figura 1M). Micrografia de uma rede tubular em Figura 1N e 1O é invertida para aumentar o contraste. Figura 1 P e 1T retrata a redução da densidade tubular em uma região de membrana ao longo de 3 h e 20 min. A diminuição da densidade tubular ocorre devido a depinning gradual seguido de retração do DLBM da superfície durante o período experimental. Ao longo do tempo, o número de pontos, liberados da fixação aumenta, levando a rearranjos e uma redução da área coberta por tubos (Figura 1P e 1T). Os rearranjos tubulares são motivados pela minimização de energia de superfície livre de um nanotubo de lipídios suspendido entre dois pontos fixos. É bem estabelecido que a maneira mais eficiente de minimizar a energia de superfície de um nanotubo é reduzir seu comprimento15. Portanto, quando as regiões de fusogenic, que inicialmente os nanotubos na superfície, são de fixado, os nanotubos deslize e organizar-se espontaneamente, adotando um comprimento mínimo. Estes rearranjos causam uma gradualmente reduzida cobertura da superfície pelos nanotubos (Figura 1P e 1T).
Não visualizamos o Ca2 +-mediada pontos de fixação, mas podemos estabelecer seus locais como os pontos onde os tubos têm voltas afiadas ou terminais. Curvas são referidas como V-junções15 ou pontos de viragem devido a mudança súbita na direção do alinhamento do tubo (setas verdes na Figura 1R e 1 X). O ponto de extremidade representa o término do tubo, que impede que o tubo retrai (setas laranja na Figura 1X). Durante a re-organização, energeticamente favorável disposição dos tubos, identificado como "Y-junções" ou "3-way junções", aparecem. A junção-Y conecta três tubos com aproximadamente 120° ângulos entre cada tubo, onde o comprimento total do tubo mais curto pode ser garantido. As junções de Y, que não possuem um ponto de extremidade e em vez disso, estão posicionadas entre múltiplos nanotubos, não estão fixadas. Este é o único tipo de junção-Y que pode executar o deslizamento (setas azuis, Figura 1R). Como mostrado na Figura 1R e 1S, deslizando de uma junção-Y ao longo de um resultados de interseção altamente instável na formação de duas individuais Y-junções (setas azuis na Figura 1S). Linha branca tracejada sobreposta em Figura 1S representa o contorno do fragmento de rede tubular no Figura 1R. Uma fração das Y-entroncamentos possui terminais da extremidade (seta amarela na Figura 1T) que, ao longo do tempo, eventualmente retraem (Figura 1-U). A transformação de uma junção-V para uma única, tubo reto pela depinning do ponto de interseção de segmentos de dois linha e pela retração de um dos tubos formando o V pode ser observado na Figura 1V e 1W e Figura 1X e 1Y, respectivamente.

Figura 1 : Transformação de lipídios depósitos para redes tubulares, como ER. (A-F) Laser scanning confocal micrografias das vesículas na amostra inicial, construído em 3D. (A-C) Aviões de vesículas lipídicas multilamellar (MLV, depósito de lipídios) em xyz, xz e yz, respectivamente. (D-F) Opiniões semelhantes de uma vesícula unilamellar gigante (chefe). A parte interna das GUVs é oca, o que torna o material lipídico para espalhar significativamente limitada. Os reservatórios de lipídios útil para este método são, portanto, MLVs. (G) ilustração da câmara de observação, montada em um microscópio invertido, no qual são depositados os buffers e lipídios. A câmara é composta por um PDMS quadro aderiu em uma Al2O3 revestido da lamela, fornecendo um alto volume aberto. (H-J) Ilustração dos fenômenos propagação do MLV na presença de Ca2 +. (H) em caso de contacto com Al2O3, o MLV espontaneamente se espalha na forma de uma circular, dupla BICAMADA membrana lipídica (DLBM). (I) vista esquemática lateral da DLBM no plano xz, onde as periferias executam o movimento de rolamento. MLV (d = 5-15 µm) e DLBM (espessura = 10 nm) não são desenhadas em escala. (J) uma micrografia confocal de uma propagação DLBM de vista superior. (K) descreve a etapa principal do presente protocolo, onde o buffer é trocado para um que contém Ca2 + agentes quelantes, inibindo a propagação, causando retração do DLBM para o MLV e levando à formação das nanotubos de lipídios. (L-Y) Micrografias das redes nanotubo obtidas com o método descrito. (L) close-up da região em (M) marcado em um quadro. As regiões vermelho-contínuas (L e M) representam o DLBM (também marcado com uma linha tracejada azul em M). Micrografia de uma rede tubular (N e O) é invertida para aumentar o contraste. (P e Q) Representação da redução da densidade tubular em uma região de membrana ao longo de 3 h e 20 min. (R-Y) representante tubular reorganizações. (R e S) Deslizamento, (T e U) fazendo a transição de uma junção-Y para V-junção por retração de um ponto de extremidade e (V e W) de fixação de um ponto de viragem, resultando na erradicação de uma junção-V. (X e Y) Retração de um ponto de extremidade. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2 : Resultados negativos potenciais. (A e B) Ruptura da membrana distal devido a um longo tempo de espera antes da troca dos buffers. As áreas de ruptura, onde a membrana proximal torna-se visível, aparecem como regiões escuras em comparação com as regiões de unruptured membrana distal. A inserção de painel B mostra a intensidade da luz ao longo da seta azul na Micrografia. A intensidade das regiões de ruptura da membrana (BICAMADA proximal/individual torna-se visível) corresponde a metade da intensidade da membrana unruptured (BICAMADA de duplo/distal). (C) a aparência de um patch de lipídios seco formado como resultado da remoção de todo líquido da câmara de observação. (D e E) Rompimento da membrana de troca rápida de amortecedores através de pipeta automática. Em (D), o MLV tem dividido em 2 MLVs, levando a um patch de lipídios não circular, deformado. Na alínea E, o padrão do fluxo (setas), criado por forte injeção do buffer quelante-HEPES, reflecte-se sobre a estrutura da membrana tubulated. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
A discussão a seguir, são descritas as etapas críticas, possíveis modificações e limitações do protocolo. O primeiro passo crítico é a correcta montagem da câmara de observação, dado que a adesão do quadro PDMS à superfície Al2O3 é intrinsecamente fraca. No caso onde o quadro não adere ao substrato corretamente, o conteúdo irá vazar da câmara de observação e o experimento vai chegar a um impasse. Os principais fatores que impedem a adequada vedação da superfície e quadro são 1) a falta de limpeza completa do quadro PDMS (reutilizado) e 2) ar bolhas que ocasionalmente obter aprisionadas entre o quadro e o substrato. Deve ser usado um quadro PDMS recém preparado ou completamente limpo. O quadro deve ser lavado antes e após cada utilização com isopropanol, seguido por lavagem com água e secar com nitrogênio. As Al2O3 superfícies não requerem qualquer limpeza prévia, uma vez que eles são fabricados em um ambiente de sala limpa e mantidos em recipientes fechados até o uso. Devido a natureza anfótera da Al2O3, não devem ser exposta a soluções fortemente ácidas ou básicas. Outros projetos para a câmara de observação podem ser empregados, dependendo da acessibilidade da instalação do individual. Características importantes desta câmara são acesso gratuito para a amostra de líquido da parte superior aberta e a inércia do material quadro no que diz respeito a soluções usadas e amostras. As dimensões da Câmara também são um fator significativo, como eles devem acomodar um volume de 0,5 a 1 mL. Desde que as superfícies utilizadas são normalmente tamanho padrão lamelas (24 x 60 mm), o volume da câmara é determinado principalmente pela espessura da moldura. A nosso conhecimento, espaçadores com o tamanho e profundidade que pode acomodar os volumes de amostra normalmente tratados neste protocolo não estão disponíveis comercialmente. Por conseguinte, dedicamos uma seção no protocolo para detalhes de fabricação e montagem de um quadro de câmara de amostra.
O outro passo crítico neste protocolo é a troca de reserva. Um desafio nesta etapa é o tempo necessário para realizar esta troca. A propagação do MLV em caso de contacto com o Al2O3 substrato é instantânea, e da contínua expansão do DLBM leva à sua ruptura, encerrando o experimento (Figura 2A, B). Portanto, a divulgação deve ser constantemente monitorizada, e a troca de reserva deve ser realizada em tempo hábil. A troca não deve ser realizada muito rapidamente após a inicialização da divulgação, a fim de permitir que os patches de membrana atingir um tamanho ideal (100-200 µm de diâmetro). Por outro lado, adesão contínua na superfície faz com que tensão de membrana de alta, que leva à ruptura. Assim, a membrana todos patches eventualmente ruptura se espalhar não é interrompida. O momento da ruptura difere para cada patch, uma vez que depende do tamanho e estrutura interna do MLV e acessibilidade dos lipídeos nele. Portanto, o momento da troca deve ser providenciado para um Commit na qual os patches não-ruptura com tamanhos ideais representam a maioria da população inteira. Outro desafio na etapa de troca de amortecedor é a taxa de remoção e adição de buffers. Realizar esta substituição muito rapidamente tem uma influência prejudicial sobre as estruturas de membrana final (Figura 2C-E). A extração excessiva de Ca2 +- HEPES buffer sem deixar uma película fina de líquido sobre o substrato, resulta em manchas secas e irreversivelmente deformado de membrana (Figura 2C). Mesmo que uma quantidade adequada de líquido é mantida na superfície, adição abrupta do buffer quelante-HEPES também causa perturbação das estruturas de membrana. Figura 2 D,E mostra a aparência típica de patches de membrana direccionado rompidas. O rompimento total morfológico não necessariamente influencia as propriedades dinâmicas das estruturas finais (i.e., os rearranjos tubulares nas restantes áreas ainda irão ocorrer). No entanto, ele se tornará difícil de observar a transformação material nas estruturas deformadas. Por exemplo, na Figura 2D, seria difícil determinar a direção para a qual se retrai MLV a DLBM.
Uma eventual modificação do protocolo é a composição de lipídios utilizada. O foco principal tem sido na fosfolipídios que dominam a composição de ER em mamíferos e16 do fermento (e. g., fosfatidilcolina (PC), fosfatidiletanolamina (PE) e fosfatidilinositol (PI). Os experimentos originais foram realizados usando misturas de PC e PI4. Nos resultados apresentados, foi usadas uma mistura de PC e droga e um derivado do PE. No entanto, nem todas as composições de lipídios arbitrárias foram encontradas para criar as estruturas tubulares, obtidas através do presente protocolo. Algumas das outras misturas lipídico experimentalmente investigada envolvem extrato total coração, polar de extracto de feijão de soja, extrato de e. coli polar, misturas de PC com stearoyl-2-hydroxy-sn-glycero-3-phosphoinositol (Super-PI) em razões diferentes e misturas de PC-PE-PI-Posphatidyl serina (PS), em proporções variáveis. Desde estruturas tubulares da membrana possuem altas curvaturas e requerem um arranjo especial de moléculas lipídicas individuais, espera-se que o fenômeno observado é lipídico composição específica.
Outra modificação aplicada neste protocolo é o método para a fabricação de superfície. Aqui, o ALD foi usado para fabricar as Al2O3 revestido lamelas. Isto difere do método de deposição originalmente relatado, reativo4que sputtering. Enquanto isso indica que um método alternativo de fabricação superfície ainda pode levar a ER, como tubulation, uma limitação importante parece ser a especificidade do material superficial. O modo de propagação e a força de adesão são altamente dependentes de propriedades do material de superfície, que influenciam fatores como interações eletrostáticas, molhabilidade, hidrofobicidade e rugosidade da superfície. Al2O3 superfícies fornecem a força de adesão ideal, e filmes de lipídios podem ambos anexar fortemente o suficiente para se espalhou como uma membrana de bicamada lipídica dupla e desanexar a redes tubulares de forma após a remoção de íons de Ca2 + . Anteriormente, nós testamos o mesmo experimento com SiO2, em que as vesículas multilamellar se espalhou como uma membrana de bicamada lipídica dupla, mas sem formação de rede tubular foi observada após a adição de quelantes17. De fixação e formação do tubo são observados somente em Al2O3 ou plasma gravado Al18. Nossas investigações revelaram que o parâmetro contribuindo levando a tal fenômeno era o zeta potencial das superfícies, para que Al e Al2O3 foram perto de zero (mV) e SiO2 significativamente negativos. O potencial zeta de borosilicato é semelhante ao SiO219; Portanto, a adesão de filmes de lipídios em borosilicato é igualmente forte e irreversível. Na verdade, lipídios multilamellar reservatório de contato com superfícies de borosilicato normalmente leva a ruptura imediata e formação de único lipid bilayers20. As Al2O3 superfícies necessárias para este protocolo não são prontamente ou comercialmente disponíveis. Podem, no entanto, ser ordenada personalizada de fabricantes de vidro e o substrato de especialidade. Acesso às instalações de salas limpas com equipamento de fabricação de película fina é altamente recomendado.
Os outros métodos ascendente existentes para fabricar redes tubulares, como ER2,10 envolvem proteínas, bem como a entrada de energia química (ex., GTP e ATP). Rapoport e colegas2 relatou a formação de redes-ER vidro lamelas em vitro misturando as proteínas de membrana-flexão presentes na sala de emergência, com fosfolipídios e GTP. O trabalho de Bachand et al 10 mostra como essas redes tubulares dinâmicas podem ser criadas usando motores moleculares e ATP como fonte de energia. Este protocolo apresentado não requer proteínas de membrana ou hidrólise de compostos orgânicos para a energia. Os componentes essenciais só são o substrato sólido e os fosfolipídios. Purificação e extração de proteínas não são necessários. Este protocolo fornece, em termos de simplicidade das moléculas constitutivas, o modelo mais básico de ER.
Com este modelo básico, com base em lipídios ER estabelecido, construir a complexidade, adicionando componentes ER-associada é de interesse, uma vez que permite a investigação dos impactos individuais do sistema. Semelhante às redes ER reais, tubos no modelo são dinâmicos. A conjugação de e a migração das proteínas de membrana rotulada ou partículas fluorescentes em toda a rede tubular, podem dar informações sobre a direção do movimento da membrana. Encapsulamento e monitoramento dos líquidos dentro do DLBM e tubos fluorescentes durante a transformação e um possível mapeamento do transporte intratubular conteúdo podem servir como outro foco. Finalmente, pode adoptar-se uma transição do modelo ER 2D resultantes deste protocolo para um modelo 3D de ER suave através de encapsulamento das redes em arquiteturas de hidrogel.
Divulgações
Os autores não têm nada para divulgar.
Agradecimentos
Agradecemos o Prof Aldo Jesorka partir da Chalmers University of Technology na Suécia por suas inestimáveis comentários sobre o manuscrito. Este trabalho foi tornado possível através do apoio financeiro obtido a partir da concessão do projeto de Conselho de pesquisa da Noruega (Forskningsrådet) 274433, UiO: ambiente de convergência das Ciências da vida, o Conselho Sueco de pesquisa (Vetenskapsrådet) Project Grant 2015-04561, bem como o financiamento de start-up fornecido pelo centro de Medicina Molecular Noruega & faculdade de matemática e ciências naturais na Universidade de Oslo.
Materiais
| Name | Company | Catalog Number | Comments |
| Pear-shape flask 10 mL | Lenz Laborglasinstrumente | 3.0314.13 | In which the lipid mixture is prepared |
| Hamilton 5 mL glass syringe (P/N) | Hamilton | P/N81520 | For transfer of the chloroform to beaker |
| Custom large hub needle Gauge 22 S | Hamilton | 7748-18 | Removable needle for syringe specified in row 3 |
| Hamilton 250 µL glass syringe | Hamilton | 7639-01 | Used for transfer of lipids in chloroform to the flask |
| Large hub Gauge 22 S | Hamilton | 7780-03 | Removable needle for syringe specified in row 5 |
| Hamilton 50 µL glass syringe | Hamilton | 7637-01 | Used for transfer of fluorophore-conjugated lipids to the flask |
| Small hub Gauge 22 S | Hamilton | 7770-01 | Removable needle for syringe specified in row 7 |
| Chloroform anhydrous (≥99%) | Sigma-Aldrich | 288306 | Used to complete the lipid mixture to a total of 300 µL |
| Soy L-α Phosphatidyl choline lipid (Soy PC) | Avanti Polar Lipids Inc | 441601 | phospholipid species contributing to 69% of the total composition/mixture |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine (DOPE) | Avanti Polar Lipids Inc | 850725 | phospholipid species contributing to 30% of the lipid composition/mixture |
| L-α Phosphatidyl inositol lipid (Soy PI) | Avanti Polar Lipids Inc | 840044 | alterative phospholipid species contributing to 30% of the lipid composition/mixture (from the original article Bilal and Gözen, Biomaterials Science, 2017) |
| Texas Red 1,2-dihexadecanoyl-sn-glycero-3-phosphoethanolamine,triethylammonium salt (Texas Red DHPE) | Invitrogen (Thermo Fisher Scientific) | T1395MP | Fluorescent-lipid conjugate, 1% of the lipid composition/mixture |
| Digital Dry Baths/Block Heaters | Thermo Fischer | 88870006 | To warm glycerol in order to decrease its viscosity |
| Glycerol for molecular biology (≥99%) | Sigma Life Science | G5516 | For lipid preparation |
| PBS buffer (pH=7.8); ingredients below in rows 17-21 | Used to prepare the lipid suspension | ||
| TRIZMA base, primary standard and buffer (≥99%) | Sigma Life Science | T1503 | Used to prepare PBS buffer |
| Potassium phosphate tribasic, reagent grade (≥98%) (K3PO4) | Sigma-Aldrich | P5629 | PBS buffer ingredient |
| Magnesium sulfate heptahydrate, BioUltra (≥99,5%) KT (MgSO47H2O) | Sigma Life Science | 63138 | PBS buffer ingredient |
| Potassium phosphate monobasic, anhydrous, free flowing, Redi-Dri, ACS (KH2PO4) | Sigma-Aldrich | 795488 | PBS buffer ingredient |
| Ethylenediaminetetraacetic acid disodium salt dihydrate ACS reagent, 99.0-101.0% (Na2EDTA) | Sigma-Aldrich | E4884 | PBS and Chelator-HEPES buffer ingredient |
| Ultrasonic cleaner USC-TH | VWR | 142-0084 | Ultrasonication of rehydrated lipids |
| Rotary evaporator - Büchi rotary evaporator Model R-200 | Sigma | Z626797 | For evaporation of chloroform |
| Pressure meter - Vacuum regulator IRV-100 | SMC | IRV10/20 | For controlling the pressure value during lipid dehydration |
| HEPES-buffer (pH=7.8); ingredients below in rows 26-27 | Used for rehydration of lipids. Content: 10 mM HEPES with 100 m NaCl diluted in ultrapure deionized water | ||
| HEPES ≥99.5% (titration) | Sigma Life Science | H3375 | HEPES-buffer ingredient |
| Sodium chloride for molecular biology, DNase, RNase, and protease, none detected, ≥98% (titration) (NaCl) | Sigma Life Science | S3014 | HEPES-buffer ingredient |
| Calcium-HEPES buffer (pH=7.8); effective ingredient below in row 29 | Used for spreading of lipids. Content: 10 mM HEPES, 100 mM NaCl, 4 mM CaCl2 diluted in ultrapure deionized water | ||
| Calcium chloride anhydrous, BioReagent, suitable for insect cell culture, suitable for plant cell culture, ≥96.0% (CaCl2) | Sigma Life Science | C5670 | To prepare Calcium-HEPES buffer |
| Chelator-HEPES buffer (pH=7.8); effective ingredient below in row 31 | Used to promote the formation of tubular networks. Content: 10 mM HEPES, 100 mM NaCl, 10 mM EDTA and 7 mM BAPTA diluted in ultrapure deionized water | ||
| 1,2-Bis(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid tetrasodium salt ≥95% (HPLC) (BAPTA-Na4) | Sigma Life Science | 14513 | Chelator-HEPES buffer ingredient |
| Sodium Hydroxide | Sigma | 30620 | Basic solution used to adjust the pH of the buffers |
| pH meter accumet™ AE150 pH | Fisher Scientific | 1544693 | Used to measure the pH of all buffers |
| Glass petri dish | VWR | HECH41042012 | 6 cm, used for making the PDMS sheet |
| Potassium hydroxide ACS reagent, ≥85%, pellets (KOH) | Sigma-Aldrich | 221473 | To make the KOH solution for cleaning glass petri dish for the fabrication of the PDMS sheet |
| Isopropanol prima ren 99.5% | Antibac AS | 600079 | KOH solution ingredient |
| Heating and drying oven - venticell | MMM Medcenter Einrichtungen GmbH | MC000714 | For drying of the glass petri dish after silanization and to cure PDMS |
| Dichlorodimethylsilane ≥99.5% | Sigma-Aldrich | 440272 | Used for silanization of glass petri dish in which PDMS sheet is prepared |
| Vacuum pump | Cole-Parmer | EW-79202-05 | Connected to desiccator |
| Sylgard 184 silicone elastomer curing agent | Dow corning | 24236-10 | Kit to make PDMS solution |
| Sylgard 184 Silicone elastomer base | |||
| Disposable scalpel | Swann-Morton | 11798343 | Used to cut the PDMS |
| Cover slips | Menzel -Gläser | MEZ102460 | 24x60 mm. Used to deposit thin film of Al2O3 |
| Atomic layer deposition system | Beneq | TFS200 (model number) | Atomic Layer deposition system used to deposit thin film of Al2O3 in microscope cover glass |
| Ellipsometer | J.A. Woollan Co. | Alpha-SE (model name) | System used to charcaterize the thickness of the film deposited on glass surface |
| Laser scanning confocal microscope | Leica Microsystems | Leica TCS SP8 X | Microscope used for visualization of the experiment |
| Objective 40x, 1.3 NA | Leica Microsystems | 1550635 | Used for visualization of the experiment |
| White light laser source | Leica Microsystems | Leica TCS SP8 X | For excitation of the membrane fluorophore |
Referências
- Chen, S., Novick, P., Ferro-Novick, S. ER structure and function. Current Opinion in Cell Biology. 25 (4), 428-433 (2013).
- Powers, R. E., Wang, S., Liu, T. Y., Rapoport, T. A. Reconstitution of the tubular endoplasmic reticulum network with purified components. Nature. 543 (7644), 257-260 (2017).
- Pendin, D., McNew, J. A., Daga, A. Balancing ER dynamics: Shaping, bending, severing, and mending membranes. Current Opinion in Cell Biology. 23 (4), 435-442 (2011).
- Bilal, T., Gözen, I. Formation and dynamics of endoplasmic reticulum-like lipid nanotube networks. Biomaterials Science. 5 (7), 1256-1264 (2017).
- Shibata, Y., et al. Mechanisms determining the morphology of the peripheral ER. Cell. 143 (5), 774-788 (2010).
- Ozcan, L., Tabas, I. Role of endoplasmic reticulum stress in metabolic disease and other disorders. Annual Review of Medicine. 63, 317-328 (2012).
- Yamanaka, T., Nukina, N. ER dynamics and derangement in neurological diseases. Frontiers in Neuroscience. 12, (2018).
- Taalab, Y. M., et al. Mechanisms of disordered neurodegenerative function: Concepts and facts about the different roles of the protein kinase RNA-like endoplasmic reticulum kinase (PERK). Reviews in the Neurosciences. , (2018).
- Shemesh, T., et al. A model for the generation and interconversion of ER morphologies. Proceedings of the National Academy of Sciences of the United States of America. 111 (49), 5243-5251 (2014).
- Bouxsein, N. F., Carroll-Portillo, A., Bachand, M., Sasaki, D. Y., Bachand, G. D. A continuous network of lipid nanotubes fabricated from the gliding motility of kinesin powered microtubule filaments. Langmuir. 29 (9), 2992-2999 (2013).
- Sackmann, E. Endoplasmatic reticulum shaping by generic mechanisms and protein-induced spontaneous curvature. Advances in Colloid and Interface Science. 208, 153-160 (2014).
- Jesorka, A., et al. Generation of phospholipid vesicle-nanotube networks and transport of molecules therein. Nature Protocols. 6, 791 (2011).
- Hook, D. A., Olhausen, J. A., Krim, J., Dugger, M. T. Evaluation of Oxygen Plasma and UV Ozone Methods for Cleaning of Occluded Areas in MEMS Devices. Journal of Microelectromechanical Systems. 19 (6), 1292-1298 (2010).
- Gözen, I., et al. Fractal avalanche ruptures in biological membranes. Nature Materials. 9 (11), 908-912 (2010).
- Lobovkina, T., Dommersnes, P., Joanny, J. -. F., Hurtig, J., Orwar, O. Zipper Dynamics of Surfactant Nanotube Y Junctions. Phys Rev Lett. 97, (2006).
- van Meer, G., Voelker, D. R., Feigenson, G. W. Membrane lipids: where they are and how they behave. Nature Reviews Molecular Cell Biology. 9 (2), 112-124 (2008).
- Gözen, I., et al. Repair of large area pores in supported double bilayers. Soft Matter. 9 (10), 2787-2792 (2013).
- Gözen, I., et al. Thermal migration of molecular lipid films as a contactless fabrication strategy for lipid nanotube networks. Lab on a Chip. 13 (19), 3822-3826 (2013).
- Sides, P. J., Hoggard, J. D. Measurement of the Zeta Potential of Planar Solid Surfaces by Means of a Rotating Disk. Langmuir. 20 (26), 11493-11498 (2004).
- Nissen, J., Jacobs, K., Rädler, J. O. Interface Dynamics of Lipid Membrane Spreading on Solid Surfaces. Physical Review Letters. 86 (9), 1904-1907 (2001).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados