Method Article
Caracterização de Junções Neuromusculares em Camundongos por Microscopia Confocal Combinada e Super-Resolução
* Estes autores contribuíram igualmente
Neste Artigo
Resumo
Este protocolo descreve um método para análise morfométrica de junções neuromusculares por microscopia confocal combinada e STED que é usado para quantificar alterações patológicas em modelos de camundongos de SMA e CMS relacionada ao ColQ.
Resumo
As junções neuromusculares (NMJs) são sinapses altamente especializadas entre neurônios motores inferiores e fibras musculares esqueléticas que desempenham um papel essencial na transmissão de moléculas do sistema nervoso para os músculos voluntários, levando à contração. Eles são afetados em muitas doenças humanas, incluindo distúrbios neuromusculares hereditários, como distrofia muscular de Duchenne (DMD), síndromes miastênicas congênitas (CMS), atrofia muscular espinhal (AME) e esclerose lateral amiotrófica (ELA). Portanto, o monitoramento da morfologia das junções neuromusculares e suas alterações em modelos de camundongos com doença representa uma ferramenta valiosa para estudos patológicos e avaliação pré-clínica de abordagens terapêuticas. Aqui, métodos para marcar e analisar a morfologia tridimensional (3D) das partes pré e pós-sinápticas de placas terminais motoras de fibras musculares murinas provocadas são descritos. Os procedimentos para preparar amostras e medir o volume, a área, a tortuosidade e a morfologia/ocupação terminal axônica por imagem confocal, e a distância entre as pregas juncionais pós-sinápticas e a largura da faixa do receptor de acetilcolina (AChR) por microscopia de depleção de emissão estimulada por super-resolução (STED) são detalhados. Alterações nesses parâmetros NMJ são ilustradas em camundongos mutantes afetados por SMA e CMS.
Introdução
A junção neuromuscular (NMJ) é uma estrutura complexa composta por um terminal axônico motor, uma célula de Schwann perissináptica e uma porção de miofibra esquelética envolvida na transmissão de informações químicas e no acoplamento da atividade do neurônio motor inferior à contração muscular. Em mamíferos, a morfologia da junção neuromuscular muda durante o desenvolvimento, adotando uma forma típica de pretzel após a maturação, com diferenças de forma e complexidade entre as espécies, e mostra algum grau de plasticidade em resposta a processos fisiológicos como exercício ou envelhecimento 1,2,3,4 . A placa terminal motora pós-sináptica forma invaginações de membrana denominadas pregas juncionais, onde a parte superior contendo receptores de acetilcolina (AChR) está em contato próximo com o ramo axônico terminal pré-sináptico5.
Alterações morfológicas e funcionais nas junções neuromusculares contribuem para a fisiopatologia de diversos distúrbios neurodegenerativos, como atrofia muscular espinhal (AME) e esclerose lateral amiotrófica (ELA), miopatias como distrofia muscular de Duchenne (DMD), síndromes miastênicas congênitas (CMS), miastenia gravis (MG) e miopatias centronucleares (CNM) e sarcopenia associada ao envelhecimento 3,6,7,8,9, 10,11,12. Nessas doenças, observam-se alterações estruturais do NMJ, como fragmentação da placa terminal, redução do tamanho da prega juncional pós-sináptica e/ou desnervação. A patologia dos NMJs pode ser um evento primário ou precoce durante a progressão da doença ou aparecer mais recentemente como um evento secundário que contribui para as manifestações clínicas. De qualquer forma, o monitoramento da morfologia dos NMJs em modelos animais dessas doenças representa um parâmetro valioso para estudar alterações patológicas e avaliar a eficácia de possíveis tratamentos.
A morfologia das junções neuromusculares geralmente é analisada por técnicas que utilizam microscopia confocal2,13,14,15 ou microscopia eletrônica 5,16, com suas limitações inerentes como resolução ou dificuldades técnicas, respectivamente. Mais recentemente, a microscopia de super-resolução também foi utilizada para visualizar regiões específicas do NMJ, como zonas ativas pré-sinápticas ou distribuição de AChR na membrana pós-sináptica16,17,18, como uma abordagem alternativa ou complementar à análise ultraestrutural por microscopia eletrônica.
Este protocolo tem como objetivo fornecer um método detalhado e reprodutível para avaliar os parâmetros morfológicos do NMJ combinando microscopia de fluorescência confocal e depleção de emissão estimulada (STED). Características importantes das placas finais pré-sinápticas e pós-sinápticas, como volume, área, tortuosidade relativa, largura da faixa AChR e distribuição terminal axonal em fibras musculares provocadas inervadas de gastrocnêmio e tibial anterior de camundongos foram quantificadas no contexto de condições normais e doentes. Em particular, os defeitos do NMJ foram exemplificado no modelo de atrofia muscular espinhal em camundongo Smn2B/-camundongo, uma doença neuromuscular com degeneração do neurônio motor causada por mutações no gene SMN1 11,19, e em uma subunidade de cauda semelhante ao colágeno de camundongos knockout assimétrico da acetilcolinesterase (ColQDex2/Dex2 ou ColQ-KO), como um modelo da síndrome miastênica congênita20, 21,22.
Protocolo
Os cuidados e a manipulação de camundongos foram realizados de acordo com a legislação nacional e europeia sobre experimentação animal e aprovados pelo comitê de ética institucional. Machos e fêmeas de camundongos Smn2B/- (fundo C57Bl/6J) e ColQDex2/Dex2 (fundo B6D2F1/J) com 3 e 6 semanas de idade, respectivamente, foram utilizados no estudo.
1. Eutanásia de camundongos e dissecção de músculos: tibial anterior e gastrocnêmio
- Proceder à anestesia em ratinhos por injeção intraperitoneal de uma solução mista de cetamina (87,5 mg/kg)/xilazina (12,5 mg/kg) (0,1 ml/20 g de peso corporal) antes da eutanásia por luxação cervical.
NOTA: Como a AME e a ColQ-CMS acometem indivíduos independentemente de seu sexo, camundongos machos e fêmeas foram utilizados no presente protocolo. - Remova os pelos dos membros posteriores usando um pequeno barbeador elétrico e lave as pernas com etanol a 70%.
NOTA: O procedimento de dissecção será diferente para cada músculo. Para dissecção do tibial anterior (AT), siga os passos 1.2.1-1.2.3 e, para gastrocnêmio (GA) (partes medial e lateral), siga os passos 1.2.4-1.2.6. Manuseie os músculos suavemente para evitar danos nos tecidos e esmagá-los ou esticá-los durante a dissecção.- Coloque o rato na posição supina.
- Faça uma incisão na pele de 5 mm com uma tesoura afiada e contundente ao longo da parte anteroexterna do membro posterior distal, paralela à tíbia, para expor o músculo. Use uma tesoura extrafina para remover a fáscia.
- Corte o tendão distal primeiro (perto da pata) e depois o tendão proximal (perto do joelho) usando uma tesoura extrafina e uma pinça fina curva. Manuseie o músculo com cuidado para evitar danos às miofibras e nervos.
NOTA: O tendão proximal deve ser seccionado o mais próximo possível do osso para colher todo o músculo. - Coloque o rato na posição prona, use uma tesoura afiada para fazer uma incisão na pele da parte superior do compartimento posterior distal do membro posterior até a pata e remova a pele.
- Segure o tendão de Aquiles com pinça serrilhada média, corte-o com uma tesoura extrafina e separe suavemente o GA do tecido circundante de volta à sua inserção proximal.
- No lado proximal, insira a pinça serrilhada média na bolsa formada entre o bíceps femoral (GC) e o GA. Separe os dois músculos para cortar o tendão do GA o mais próximo possível da inserção óssea com uma tesoura extrafina.
- Para fixação tecidual, colocar cada músculo em um tubo de microcentrífuga de 2 mL contendo 1 mL de solução de paraformaldeído (PFA) a 4% p/v diluída em solução salina tampão fosfato (PBS sem Ca 2+Mg 2+) e manter a 4 °C por18-24 h.
CUIDADO: O paraformaldeído e o formaldeído são tóxicos e devem ser manuseados em um exaustor químico com equipamento de proteção adequado. - No dia seguinte, lave os músculos fixos 3x por 5 min com PBS em placas de 12 poços, agitando suavemente à temperatura ambiente (RT) dentro de um exaustor químico.
Observação : O protocolo pode ser interrompido nesta etapa e continuado dentro de um mês. Neste caso, adicionar PBS suplementado com azida de sódio a 0,01% para armazenar amostras a 4 °C. - Provoque cada músculo em pequenos feixes de fibras de cerca de 1 mm de largura usando duas pinças serrilhadas finas.
NOTA: É crucial manipular os músculos muito suavemente com a pinça, sem força excessiva, para evitar danos nos tecidos durante a provocação.- Dissociar o músculo AT em 3 ou 4 feixes, dependendo do seu tamanho.
- Para GA, separe as partes medial e lateral do músculo e, em seguida, dissociar cada parte em 4-5 feixes, dependendo do seu tamanho.
2. Imunocoloração
- Proceder com a permeabilização das fibras musculares: Transfira os feixes musculares para placas de 24 poços contendo 1% (v/v) de Tritão X-100 em PBS e mantenha-os sob agitação suave (50 rpm) por 1 h a RT ou 5 h a 4 °C.
NOTA: Divida os feixes musculares entre duas placas para proceder a imunocorações separadas e minimizar o risco de confusão de anticorpos. Não os divida em mais de dois poços (1 poço/placa); caso contrário, o número (N) de NMJs que são representativos de seu status geral no músculo analisado pode ser insuficiente. - Lavar as amostras 3x por 5 min com PBS at RT e incubá-las com uma solução bloqueadora composta por albumina sérica bovina (BSA) a 4% em PBS/Triton X-100 1% por 4 h a 4 °C, sob agitação suave (50 rpm).
NOTA: Não utilize uma bomba de aspiração durante as etapas de lavagem, mas aspirar a solução manualmente com uma pipeta de 200 μL e pontas de tamanho pequeno (a referência é indicada na Tabela de Materiais). - Incubar as amostras durante a noite (O/N) a 4 °C sob agitação suave (50 rpm) com a solução de bloqueio indicada no passo 2.2 contendo anticorpos monoclonais primários contra o neurofilamento M (NF-M, 2H3, diluição 1/200) ou a glicoproteína 2 da vesícula sináptica (SV2, diluição 1/200) para rotular os terminais axónicos pré-sinápticos ou as zonas activas, respectivamente.
- No dia seguinte, lave os feixes musculares 3x por 5 min em PBS sob agitação (50 rpm).
- Para imagens confocais: Incubar os feixes musculares com anticorpos anti-camundongos secundários conjugados com um fluoróforo emissor de vermelho (F594) (diluição 1/500) e α-bungarotoxina conjugada com um fluoróforo emissor verde (α-BTX-F488) (diluição 1/1000) em PBS por 2 h a RT sob agitação (50 rpm).
- Para imagens STED: Incubar os feixes musculares com anticorpos anti-camundongos secundários conjugados com um fluoróforo emissor de verde (F488) (diluição 1/500) e α-bungarotoxina conjugada com um fluoróforo emissor de vermelho distante caracterizado por alta fotoestabilidade (α-BTX-F633) (diluição 1/1000) em PBS por 2 h em RT sob agitação (50 rpm).
NOTA: Não exponha as amostras à luz durante a incubação para evitar o fotobranqueamento.
- Lave os feixes musculares rotulados 3x por 5 min com PBS sob agitação (50 rpm) e coloque-os em uma lâmina com um meio de montagem.
NOTA: Coloque um máximo de 4 a 5 feixes musculares por lâmina para permitir a selagem. - Adicione uma tampa de vidro de grau #1,5 (ou #1,5H) (espessura de 0,17 mm) na parte superior e coloque ímãs cilíndricos em ambos os lados da lâmina para aplicar pressão e achatar os músculos.
- Mantenha as lâminas protegidas contra a luz O/N a 4°C. Sele as lâminas permanentemente com esmalte.
3. Aquisição de imagens
- Aquisições por microscópio confocal
NOTA: As imagens foram coletadas com microscópio confocal de varredura a laser invertido, utilizando uma objetiva de imersão em óleo de magnitude 63x (HCX Plan Apo CS, 1,4 abertura numérica (NA)).- Para análise cega, deixe uma pessoa não envolvida na análise codificar cada slide com um determinado número. Permanecer cego para os grupos experimentais até que a quantificação dos parâmetros NMJ esteja completa para todas as amostras.
- Inicie o software do microscópio no Modo de configuração > machine.xlhw (Figura suplementar 1).
- Coloque a lâmina no estágio do microscópio e encontre o plano de observação dentro da amostra, observando a iluminação de fluorescência de campo largo DAPI com o conjunto de filtros DAPI.
- Clique em Abrir projeto > novos projetos e crie uma pasta para armazenar aquisições de imagens (Figura 1 Suplementar).
Observação : crie um novo projeto para cada NMJ para limitar o tamanho da pasta e evitar problemas de memória do computador. - Para gerenciar os parâmetros de aquisição, clique na janela da guia Aquisição e defina o orifício confocal como unidade 1.0 Airy e potência do laser para otimizar os níveis de ganho e deslocamento para a fluorescência verde/F488 (α-BTX) usando um laser de 488 nm na placa final que deve ser fotografado (modo Live ON).
- Em seguida, otimize a aquisição de fluorescência vermelho/F594 (NF-M ou SV2) usando um laser adaptado à observação F594. Neste estudo, foi utilizado um laser de 552 nm (modo Live ON). Defina o espectro de emissão de corante com as seguintes faixas para cada laser: laser 405 (DAPI) de 414 a 483 nm, laser 488 (F488-α-BTX) de 506-531 nm e laser 552 (NF-M/SV2) de 622-650 nm.
- Colete pilhas de imagens de junções neuromusculares em cada grupo experimental com as mesmas configurações: tamanho da imagem 1024 x 1024 pixels (73,7 x 73,7 μm) a uma taxa de amostragem de 400 Hz, Bidirecional X ON, Fator de zoom 2,5, Tamanho do passo Z 0,5 μm no modo Z-Wide.
NOTA: Para cada NMJ, o número de fatias é definido para adquirir a junção inteira. As configurações de aquisição descritas acima preenchem o teorema de amostragem de Nyquist-Shanon. No entanto, o usuário pode clicar no botão Otimizar formato , presente em todos os softwares operacionais confocais recentes, para garantir que o tamanho do pixel e o passo Z atendam à taxa de amostragem ideal do Nyquist. Essa ação evitará imagens com excesso ou subamostragem, o que causará uma perda de precisão nas medições de volume. - Salve o arquivo original (.lif) ou as imagens da pilha Z (.tif) em uma pasta com um nome que inclua o codinome do slide, o tipo de coloração e o número da placa final.
NOTA: Colete sequencialmente (não simultaneamente) as varreduras usando os lasers de 488 nm e 552 nm (F488 e F594) para evitar o crosstalk da fluorescência F488 no canal F594 e vice-versa (sangria). NB: o caminho do feixe pode ser configurado com o Dye Assistant no software do microscópio. - Altere para o próximo slide codificado e repita as etapas 3.1.3-3.1.8 para cada NMJ.
- No final da sessão, clique em Abrir no Visualizador 3D e escolha um NMJ representativo de um grupo experimental para visualizar a rotulagem 3D.
Observação : esse modo de exibição ajudará a verificar se os parâmetros de aquisição estavam corretos. - Feche o software do microscópio, limpe as objetivas com os tecidos da lente e, em seguida, desligue o sistema.
- Aquisições por microscopia STED
NOTA: As imagens foram coletadas com microscópio confocal de varredura a laser invertido, equipado com Gated STED a 775 nm, utilizando uma objetiva de imersão em óleo de 100x (HC PL APO CS2 1.4 NA).- Para análise cega, deixe uma pessoa não envolvida na análise codificar cada slide com um determinado número. Permanecer cego para os grupos experimentais até que a quantificação dos parâmetros NMJ esteja completa para todas as amostras.
- Inicie o software do microscópio no Modo de configuração > machine.xlhw e STED ON (Figura 2 suplementar).
- Clique em Abrir projeto > Novos Projetos para criar uma pasta para armazenar aquisições de imagens.
Observação : gere uma nova pasta para cada slide para limitar o tamanho da pasta e evitar problemas de memória do computador. - Coloque a lâmina no palco do microscópio e visualize-a sob iluminação de fluorescência de campo largo usando o laser de 488 nm para encontrar o plano de observação dentro da amostra.
- Procure um NMJ marcado com coloração de neurofilamento M (NF-M) ou SV2 usando o laser de 488 nm com uma detecção espectral de 506-531 nm.
- Quando um NMJ tiver sido identificado, clique em Ativar STED e comece a adquirir imagens em uma região que contenha várias dobras juncionais (Figura suplementar 3) usando o laser de 635 nm com uma detecção espectral de 640-750 nm.
NOTA: Tenha cuidado com a tabela de pesquisa de saturação durante a aquisição da imagem e clique no botão LUT rápida para evitar a superexposição (valores de cinza >255; para 8 bits). - Colete as imagens de cada grupo experimental com as mesmas configurações: tamanho da imagem 2048 x 2048 pixels (38,75 x 38,75 μm) a uma taxa de amostragem de 400 Hz.
NOTA: A potência do laser de esgotamento (STED) é definida como 65%. - Salve as imagens com um nome de arquivo que inclua o código do slide.
NOTA: É possível clicar em Formato XY Otimizado: Definir Formato para obter a melhor configuração de aquisição com imagens STED. - Altere para o próximo slide codificado e repita as etapas 3.2.3-3.2.8. Repita este procedimento para todos os slides.
- No final da sessão de microscopia STED, transfira os arquivos de imagem para outro computador e salve os arquivos originais (. lif) em uma unidade ou servidor externo.
- Desligue o software do microscópio, limpe as objetivas com os tecidos da lente e, em seguida, desligue o sistema.
4. Análise de imagem - microscopia confocal
NOTA: Todas as imagens foram processadas com computadores que utilizam o sistema operativo Microsoft Windows 10 professional.
- Inicie o ImageJ e a macro personalizada para calcular o volume da placa final NMJ pós-sináptica, a área de Projeção de Intensidade Máxima (MIP) e a tortuosidade relativa.
- Processe pilhas de imagens NMJ usando o freeware23 do NIH ImageJ, o plugin iGeodesic e a macro personalizada para obter medições de parâmetros NMJ. Inicie o software ImageJ.
NOTA: A última versão do ImageJ está disponível gratuitamente e pode ser baixada24. A fim de abrir formatos de arquivo proprietários, Bio-Formats Package25 plugin tem que ser baixado26 . Esta etapa não é necessária caso o operador use Fiji porque o plugin já está instalado no software. O plugin iGeodesic27 para calcular tortuosidade também está disponível online28; verificar a disponibilidade deste plugin na versão ImageJ/Fiji que será usada. As macros personalizadas também estão disponíveis online29. - Arraste e solte o Macro_NMJ_VOL_Marinelloetal.ijm (personalizado, Arquivo de Codificação Suplementar 1) na janela ImageJ; a macro será aberta em uma segunda janela. Nesta nova janela, clique em Macros > Executar macro.
Observação : A macro pode processar arquivos proprietários e TIFF. Os arquivos devem atender aos seguintes critérios: para formatos de arquivo proprietários, salve apenas uma junção (ou seja, a pilha de imagens) por arquivo, ordenada em uma pasta; para imagens TIFF, os arquivos devem ser salvos em uma pasta contendo subpastas, cada uma chamada JunctionX (X corresponde a um número NMJ) com as pilhas de imagens de uma determinada junção (RGB TIFF) (Figura 4 Suplementar). - Selecione a pasta nativa que contém as subpastas Junction que devem ser analisadas e clique em Selecionar.
- No novo menu pop-up chamado Saving Folder, selecione a pasta de armazenamento e clique em Select.
- No novo menu pop-up chamado Tipo de imagem, selecione o formato das aquisições da pilha Z.
- Selecione o canal RGB correspondente à coloração de interesse e indique o tamanho do pixel XY e o passo Z (z). A macro executará automaticamente a análise.
Observação : caso os formatos de arquivo proprietários sejam selecionados, a macro lê diretamente o tamanho do pixel e Z-step (z). No entanto, o usuário ainda precisa indicar o canal de interesse (C1, C2 ou C3). A macro fornecerá uma DataSheet (.csv) para cada parâmetro de junção (volume da placa final, área MIP e tortuosidade) na pasta de salvamento. A macro também gera três . Os arquivos TIF, que correspondem ao perímetro dos α de coloração Drawing_MaxprojX.tif-BTX, DrawingJunctionX.tif e MIP MaxprojX.tif. Esses arquivos TIFF são gerados para verificar a qualidade das aquisições e para garantir que o processamento de imagem tenha sido realizado corretamente.
Volume NMJ pós-sináptico (V): A macro separará as imagens de um único NMJ e manterá o canal α-bungarotoxina F488 correspondente à placa terminal pós-sináptica. A pilha é segmentada usando o limiteOtsu 30 na fatia intermediária da pilha. A imagem binária resultante é dilatada em 1 pixel e o recurso Analisar partículas é usado para medir a área da placa final de cada objeto detectado. Para obter o volume NMJ pós-sináptico, a macro soma todas as áreas de placa final medidas da pilha e multiplica-o pelo valor do passo Z (z) em μm.
Área da placa final de projeção de intensidade máxima (MIP): Depois que a pilha é limiarizada, a projeção de intensidade máxima (MIP) é obtida usando o recurso ImageJ do projeto Z . O recurso Analisar partículas é então usado para quantificar a área da placa final MIP.
Tortuosidade do NMJ MIP (T): A tortuosidade NMJ, que reflete o grau de complexidade da placa terminal motora pós-sináptica, incluindo dobras e perfurações31, é calculada com base em cada MIP usando a seguinte fórmula, onde d Obj(AB) é a distância entre A e B ao longo do perímetro do objeto, e dEuc(AB) é a distância euclidiana entre A e B (linha reta).
- Defina o maior valor de tortuosidade no grupo de tipo selvagem de cada condição experimental para 100% e normalize todos os outros valores da condição experimental para este valor, a fim de obter a tortuosidade NMJ relativa.
- Processe pilhas de imagens NMJ usando o freeware23 do NIH ImageJ, o plugin iGeodesic e a macro personalizada para obter medições de parâmetros NMJ. Inicie o software ImageJ.
- Inicie o ImageJ e a macro personalizada para quantificar o acúmulo de neurofilamentos pré-sinápticos e a coloração da glicoproteína 2 da vesícula sináptica.
NOTA: O acúmulo de neurofilamentos (aqui, NF-M) e/ou a distribuição alterada das vesículas sinápticas (aqui, SV2) são marcadores de transporte axonal anormal e/ou tráfico de vesículas prejudicado e foram previamente observados em NMJs de vários modelos de camundongos com AME32,33,34.- Arraste e solte o Macro_NMJ_ACCU_Marinelloetal.ijm (personalizado, Arquivo de Codificação Suplementar 2) na janela ImageJ; a macro será aberta em uma segunda janela. Nesta nova janela, clique em Macros > Executar macro.
Observação : A macro pode processar formatos de arquivo proprietários e arquivos TIFF. Os arquivos devem atender aos critérios indicados na NOTA abaixo da etapa 4.1.2. - Selecione a pasta nativa que contém as subpastas Junction que devem ser analisadas e clique em Selecionar.
- No novo menu pop-up chamado Saving Folder, selecione a pasta de armazenamento e clique em Select.
- No novo menu pop-up chamado Tipo de imagem, selecione o formato das aquisições da pilha Z.
- No pop-up Informações de Mancha, indique o rótulo e a cor pré-sinápticos e pós-sinápticos e clique em OK. Por exemplo, rótulo pré-sináptico: SV2 ou NF, cor pré-sináptica: R, rótulo pós-sináptico: BTX, cor pós-sináptica: G.
NOTA: Para formatos de arquivo proprietários, rótulos e canais correspondentes (C1, C2 ou C3) devem ser indicados. - No pop-up Tamanho do pixel, indique o tamanho do pixel XY 0,072 μm e a etapa Z 0,5 μm (z) e clique em OK. A macro executará automaticamente a análise.
NOTA: Este parâmetro corresponde ao tamanho da imagem 1024 x 1024 pixels (73,7 x 73,7 μm) selecionado antes das aquisições do microscópio confocal e está correlacionado com as configurações de objetivo e zoom. Se os formatos de arquivo proprietários forem selecionados, a macro lerá diretamente o tamanho do pixel e o Z-step (z). A macro armazenará, na pasta de salvamento, uma Folha de Dados (.csv) de volumes pré-sinápticos e pós-sinápticos, uma imagem TIFF de várias páginas da detecção atual para cada rotulagem (pré e pós-sináptica). Como indicado acima, esses arquivos TIFF são gerados para verificar a qualidade das aquisições e para garantir que o processamento de imagem tenha sido realizado corretamente.
A macro calcula o volume de coloração do neurofilamento axonal M (volume NF) do canal NF-M-F594 que colocaliza com a marcação α-bungarotoxina-F488 e o volume da coloração da glicoproteína 2 da vesícula sináptica NMJ (volume SV2) do canal SV2-F594 que colocaliza com a marcação α-BTX-F488. A acumulação de NF-M é quantificada calculando-se a razão entre o volume de NF e o volume da placa terminal pós-sináptica (α-BTX) e a ocupação do terminal do axônio NMJ pela razão entre os volumes SV2 e α-BTX, conforme mostrado abaixo.

- Arraste e solte o Macro_NMJ_ACCU_Marinelloetal.ijm (personalizado, Arquivo de Codificação Suplementar 2) na janela ImageJ; a macro será aberta em uma segunda janela. Nesta nova janela, clique em Macros > Executar macro.
5. Análise de imagem - microscopia STED
NOTA: O processamento de imagem foi realizado com o software off-line do fabricante do microscópio STED.
- Inicie o software do microscópio.
- Abra o projeto clicando no botão Abrir Projeto . Selecione o arquivo de projeto (.lif) e abra-o. As imagens são exibidas na tela junto com seus nomes.
- Na janela Processo : Clique em Mediana de > de Redução de Ruído. Na parte inferior da janela do meio, defina Raio como 5,00 e Iteração como 1,00 e desmarque Filtragem 3D.
- Em seguida, selecione a guia Abrir projetos no canto superior esquerdo da janela e escolha uma imagem.
- Clique em Aplicar para validar parâmetros. Uma nova imagem chamada "nameofimage_median001" é criada.
NOTA: É possível clicar em Visualizar antes de Aplicar para monitorar o efeito do filtro mediano, o que melhorará o contraste da imagem e suavizará os perfis de linha usados para quantificação. - Aplique o filtro a todas as imagens, conforme indicado nas etapas 5.4-5.5.
- Nas guias Abrir Projetos , clique no ícone da unidade de disquete para salvar todos os projetos, incluindo as imagens filtradas recém-criadas.
NOTA: A próxima etapa será feita usando a imagem filtrada chamada "nameofimage_median001". - Calcular a distância entre as listras AChR
NOTA: Alterações na morfologia das pregas pós-juncionais são frequentemente observadas em distúrbios neuromusculares como um sinal de patologia NMJ (imaturidade ou degeneração). A distância (d) entre as listras AChR, que são detectadas pela coloração α-bungarotoxina, é calculada gerando perfis de intensidade e quantificando a distância entre cada pico de intensidade máxima, desenhando um perfil de linha (Figura Suplementar 5).- Usando o software do microscópio, selecione o menu Quantificar na parte superior da janela central.
- Clique na guia Ferramentas no canto superior esquerdo. Selecione Intensidade no painel superior esquerdo e clique no ícone Perfil da linha . Defina Oversampling como 1 e marque Classificar canais.
- Clique nas abas Abrir Projetos e selecione a imagem filtrada a ser analisada.
NOTA: É possível ampliar a imagem rolando com o mouse do computador. O intervalo dinâmico da imagem pode ser modificado usando a barra do lado esquerdo ao lado da imagem exibida, o que facilita a visualização das listras. - Em seguida, clique no ícone Desenhar Linha no menu superior da janela direita e trace uma linha cruzando perpendicularmente várias listras / dobras juncionais.
Observação : O perfil de intensidade é exibido na janela central. - Clique na parte superior do primeiro pico e mova o ponteiro do mouse enquanto mantém o botão esquerdo do mouse pressionado até que o próximo pico máximo seja atingido.
NOTA: A informação é mostrada no perfil de intensidade, enquanto a distância entre os dois picos é exibida sob o gráfico com denominação "dx". - Clique com o botão direito do mouse enquanto estiver na imagem da janela direita e selecione Salvar ROIs. Abra os ROIs (Regiões de Interesse) salvos clicando em Carregar ROIs.
- Clique no ícone de seta no canto superior esquerdo da janela direita, clique no ROI e exclua-o clicando no ícone de lixeira.
- Repita esta operação quantas vezes forem necessárias a partir de diferentes perfis de intensidade para obter o número previsto de distâncias de faixa AChR que representarão o valor global no músculo analisado.
NOTA: O valor ideal de N pode ser calculado com antecedência com base na diferença estimada entre grupos, α risco, potência e teste unicaudal ou bicaudal. No delineamento experimental atual, aplicou-se o teste unicaudal de Mann-Whitney (risco α = 10%; potência = 80%), e o valor de N foi estimado em pelo menos cinco distâncias de faixa AChR por NMJ, a fim de comparar os dois grupos de animais.
- Largura da faixa AChR
NOTA: A largura da faixa (w) corresponde à largura total do meio-máximo (FWHM) do perfil de intensidade, que é a distância entre os pontos em que o valor de fluorescência do sinal α-BTX é metade de sua intensidade máxima (Figura Suplementar 5).- Usando o software do microscópio, selecione o menu Quantificar na janela central.
- Clique na guia Ferramentas no canto superior esquerdo. Selecione Intensidade no painel superior esquerdo e clique no ícone Determinar FWHM . Marque Classificar canais.
NOTA: Para otimizar a detecção de pico pelo software, Definir Limite e Largura foram definidos como 50 e 3, respectivamente. Adapte esses valores para cada experimento e peça conselhos a um cientista de imagem experiente. - Clique nas abas Abrir projetos e selecione a imagem filtrada a ser analisada.
NOTA: É possível ampliar a imagem exibida na janela direita rolando com o mouse do computador. Conforme indicado acima (NOTA a seguir à etapa 5.8.3), o intervalo dinâmico da imagem pode ser modificado para uma visualização de faixa ideal. - Em seguida, clique no ícone Desenhar retângulo no menu superior da janela direita. Selecione uma faixa horizontal ou vertical e desenhe um retângulo perpendicularmente à listra. Um perfil aparece na janela central.
- Clique em Vertical ou Horizontal do menu Projeção Média localizado no painel esquerdo, dependendo se a orientação da faixa é vertical ou horizontal.
- Clique em Estatísticas na janela central e leia o valor FWHM.
- Clique com o botão direito do mouse com o mouse do computador na imagem exibida na janela direita e selecione Salvar ROIs.
NOTA: Abra os ROIs salvos clicando em Carregar ROIs. - Clique no ícone de seta no canto superior esquerdo da janela direita, clique no ROI e exclua-o clicando no ícone de lixeira.
- Repita esta operação quantas vezes forem necessárias a partir de diferentes ROIs retangulares até obter o número previsto de larguras de faixa AChR, que será representativo do valor global no músculo analisado.
6. Delineamento experimental e testes estatísticos
- Realizar análises estatísticas utilizando softwares específicos.
NOTA: Os dados foram coletados de N ≥ 3 replicações biológicas e pelo menos 20 NMJs por genótipo para imagem em microscópio confocal, e N ≥ 5 repetições biológicas e N = 5 NMJs por genótipo para imagem STED, em cada grupo experimental. A significância foi avaliada pelo teste de Mann-Whitney não pareado (não paramétrico), e os valores de p estão indicados nas legendas das figuras correspondentes.
Resultados
A fim de facilitar a análise morfológica das junções neuromusculares em nível pré e pós-sináptico de forma reprodutível, foi desenvolvido um fluxo de trabalho desde a coleta muscular até a obtenção de imagens e quantificação utilizando o software microscópio e macros personalizadas ImageJ (Figura 1). Para exemplificar a utilidade deste protocolo, a morfologia dos NMJs em dois modelos de camundongos de distúrbios genéticos, camundongos Smn2B/- e ColQ Dex2/Dex2 afetados por atrofia muscular espinhal (AME) e uma forma de síndrome miastênica congênita (CMS), respectivamente, foram avaliados e os dados foram comparados com companheiros de ninhada de controle pareados por idade.
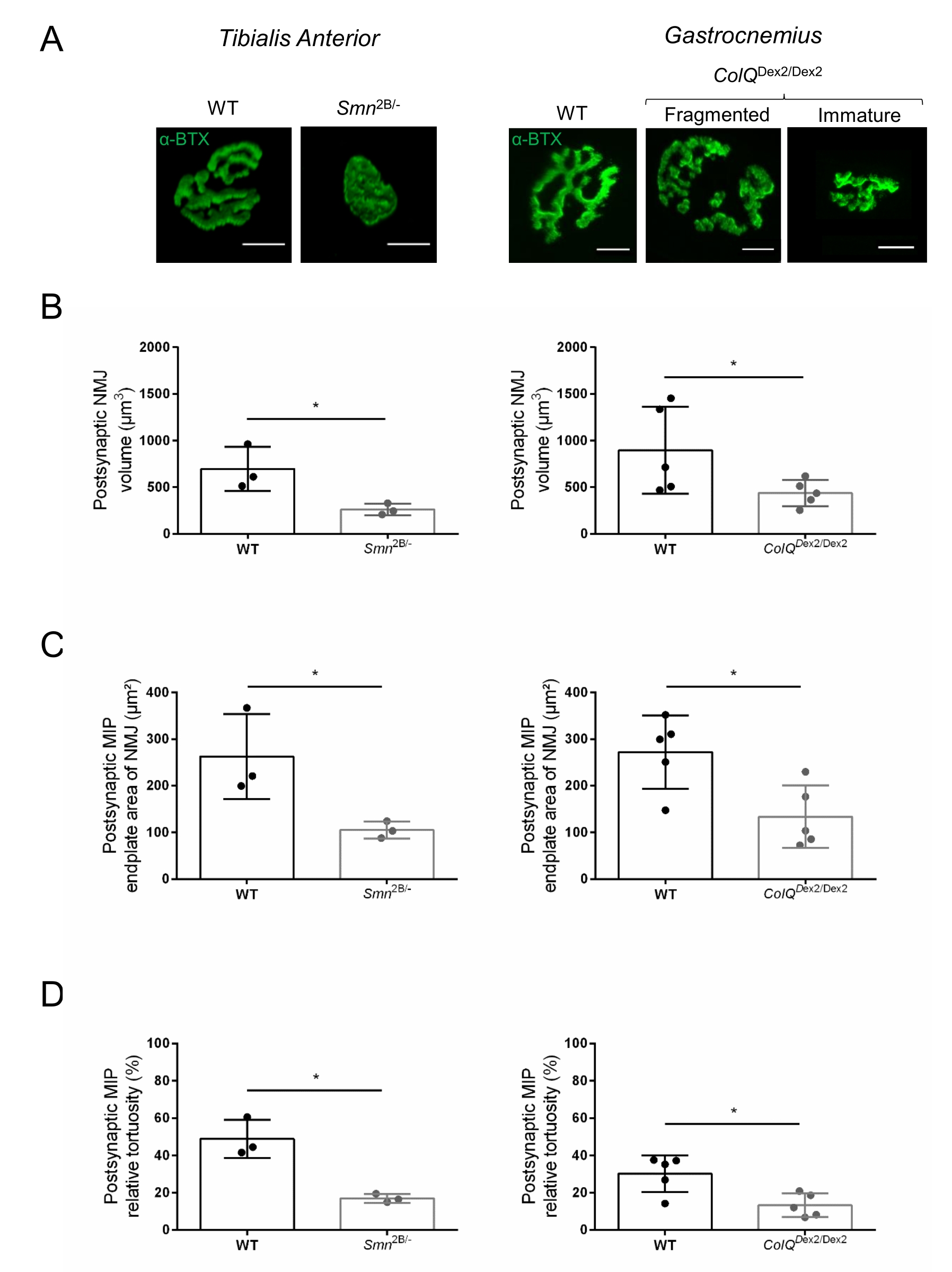
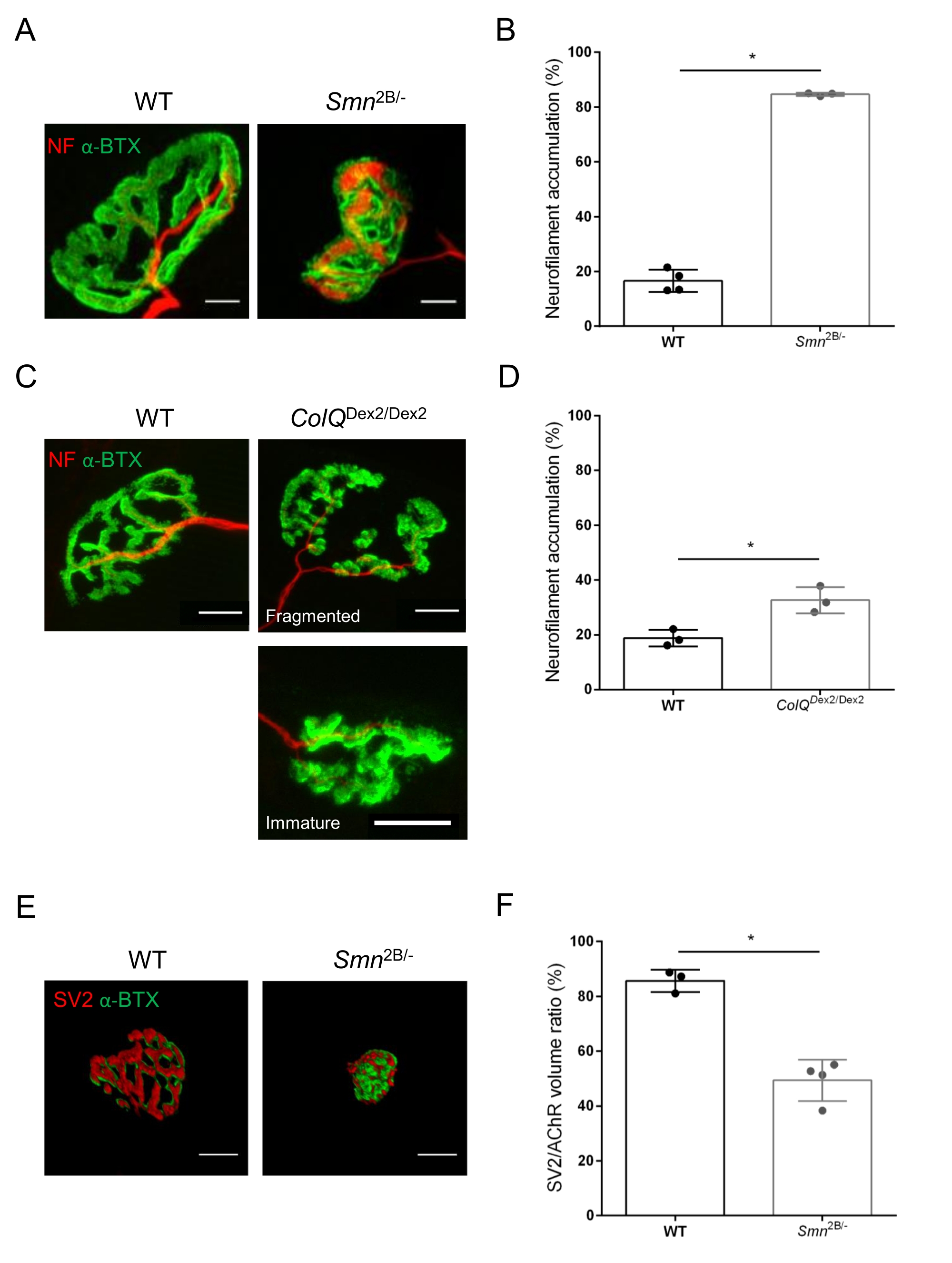
A estrutura do NMJ foi avaliada a partir dos músculos tibial anterior e gastrocnêmio de camundongos Smn2B/- (fundo C57Bl/6) e ColQDex2/Dex2 (fundo B6D2F1/J) de 3 e 6 semanas de idade, respectivamente, quando os sinais da doença já estão presentes nesses animais. Com 3 semanas de idade, camundongos Smn2B/- apresentam sinais de atraso no desenvolvimento muscular esquelético e desnervação, como atrofia e perda de NMJ35,36. Os camundongos CMS têm uma patologia primária em NMJs e manifestam uma redução no peso corporal a partir da primeira semana de vida e fraqueza muscular acentuada20 (dados não mostrados). Como mostrado na Figura 2A, a placa terminal motora pós-sináptica marcada com α-bungarotoxina fluorescente mostrou-se menor e/ou fragmentada em mutantes das duas linhas de camundongo por microscopia confocal. A quantificação de pilhas Z NMJ usando essas macros ImageJ personalizadas revelou reduções acentuadas no volume da placa final, projeção de intensidade máxima (MIP) e tortuosidade relativa em camundongos SMA e CMS em comparação com controles, como sinais de defeitos de maturação NMJ32 (Figura 2B-D). O volume da placa terminal pós-sináptica e a PImáx foram diminuídos em animais doentes (troca de dobras de 2,7 e 2,0 para o volume e 2,5 e 2,0 para PImáx, em camundongos Smn2B/- e ColQDex2/Dex2, respectivamente). A tortuosidade relativa também foi menor nos músculos deficientes em SMN e ColQ do que no WT (16,97% ± 1,33% no SMA versus 48,84% ± camundongos WT 5,90% e 13,29% ± 2,79% no CMS versus 30,20% ± 4,44% nos camundongos controle). Além disso, a quantificação da distribuição dos ramos terminais axônicos pré-sinápticos utilizando a macro personalizada ImageJ revelou um padrão alterado na distribuição do neurofilamento M nos dois modelos animais, com aumento da imunomarcação (84,65% ± 0,32% versus 16,57% ± 2,03% e 23,64% ± 2,78% versus 18,77% ± 1,73% nos camundongos Smn2B/- e ColQ Dex2/Dex2 em relação aos controles, respectivamente) (Figura 3A-D ). Pela coloração SV2, uma redução de 43% na razão de ocupação, ou seja, porcentagem de regiões contendo AChR com zonas ativas terminais nervosas adjacentes, também foi observada em camundongos Smn2B/- (49,36% ± 3,76% em SMA versus 85,69% ± 2,34% WT camundongos) (Figura 3E,F). Este parâmetro NMJ também foi calculado no GA dos mutantes ColQ Dex2/Dex2, mas não foi encontrada diferença estatisticamente significativa em comparação com os companheiros de ninhada de controle (dados não mostrados).
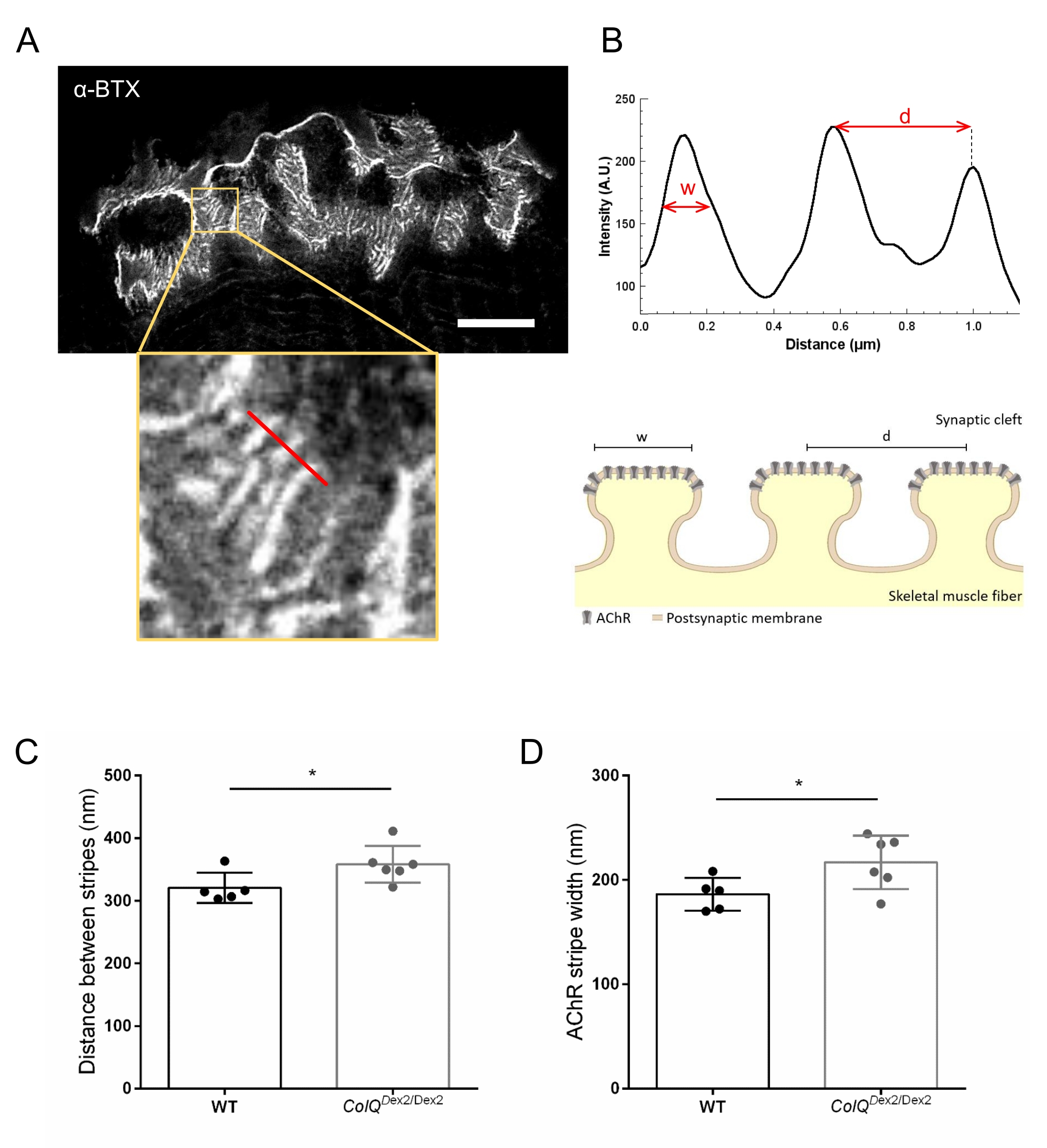
Além disso, analisamos as características da membrana pós-sináptica quantificando a distância entre as pregas juncionais e a largura das listras AChR, que estão localizadas na crista dessas dobras, no músculo deficiente em ColQ, usando microscopia de depleção de emissão estimulada por super-resolução (STED). Como mostrado na Figura 4, o aspecto dessas estruturas pode ser claramente visualizado pela marcação fluorescente de α-bungarotoxina e análise do perfil de intensidade. Avaliamos esses parâmetros do NMJ e encontramos um aumento na distância da prega juncional (d) e largura (w) das listras AChR no músculo gastrocnêmio dos mutantes (358,3 nm ± 11,97 nm e 320,8 nm ± 10,90 nm para a distância, e 216,9 nm ± 10,51 nm e 186,3 nm ± 7,015 nm para a largura, em ColQ Dex2/Dex2 em comparação com camundongos do tipo selvagem, respectivamente, p < 0,05) (Figura 4C,D).

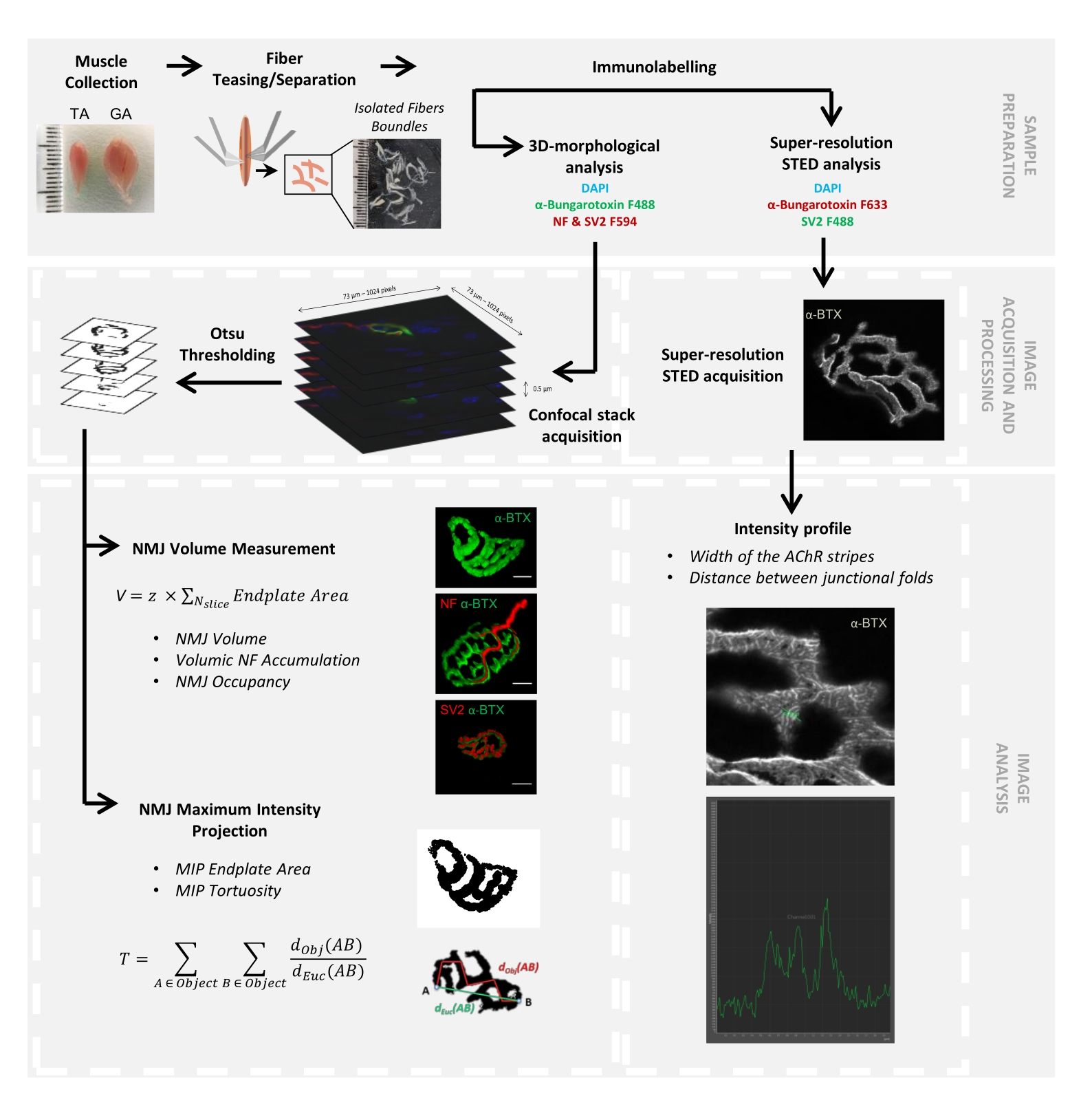
Figura 1: Fluxograma do protocolo de vídeo para caracterização de NMJ multiescala 3D por microscopia confocal e STED. Os músculos tibial anterior (AT) e gastrocnêmio (GA) foram coletados de camundongos, e as fibras musculares foram provocadas antes da marcação com α-bungarotoxina-F488 ou α-bungarotoxina-F633, DAPI, anticorpos primários direcionados contra o neurofilamento M (NF-M) e glicoproteína 2 da vesícula sináptica (SV2) e anticorpos secundários conjugados com fluoróforo (F488 ou F594). As pilhas de imagens foram adquiridas por microscopia confocal e processadas para medir o volume de NMJ pós-sináptica, o acúmulo pré-sináptico de NF-M, a ocupação terminal do axônio NMJ, a área da placa terminal da projeção de intensidade máxima pós-sináptica (MIP) e a tortuosidade (d Obj(AB) é a distância entre A e B ao longo do perímetro do objeto (linha vermelha), enquanto dEuc(AB) é a distância euclidiana entre A e B (linha verde)). Para a análise da microscopia STED, a largura das listras do receptor de acetilcolina (AChR) e a distância entre as pregas juncionais foram quantificadas a partir de perfis de intensidade da coloração α-BTX-F633. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Caracterização do NMJ pós-sináptico multiparâmetro em modelos de camundongos de atrofia muscular espinhal (AME) e síndrome miastênica congênita (CMS) relacionada ao ColQ. (A) Imagens representativas de placas terminais motoras pós-sinápticas dos músculos AT e GA marcadas com α-bungarotoxina-F488 (α-BTX). (B ) Quantificação do volume da placa terminal pós-sináptica NMJ, (C) área de projeção de intensidade máxima (PImáx) e (D) tortuosidade relativa em AT de camundongos tipo selvagem (WT) e Smn2B/- de 3 semanas de idade (gráficos à esquerda, N = 3 animais por genótipo, n = 37 e n = 56 NMJs, respectivamente) e WT e ColQ Dex2/Dex2 de 6 semanas de idade camundongos (gráficos à direita, N = 5 camundongos por genótipo, n = 89 e n = 97 NMJs, respectivamente). Os dados são expressos como a média por rato (ponto) ± EPM. As diferenças entre os grupos foram analisadas pelo teste de Mann-Whitney (* p < 0,05). A barra de escala é de 10 μm. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Análise morfométrica da distribuição terminal axônica pré-sináptica em músculos de camundongos WT e mutantes. Padrão de inervação NMJ nos músculos tibial anterior (TA) e gastrocnêmio (GA) de camundongos CMS do tipo selvagem, SMA e relacionados ao ColQ. (A, B) Junções neuromusculares representativas de AT de camundongos WT e Smn2B/- aos 21 dias de idade marcadas com anticorpos contra o neurofilamento M (NF-M, vermelho) e α-bungarotoxina-F488 (α-BTX, verde) (A), e resultados de análise quantitativa do acúmulo de neurofilamentos (B); (C, D) Junções neuromusculares representativas do GA de camundongos WT e ColQ Dex2/Dex2 de 6 semanas de idade marcadas com anticorpos contra o neurofilamento M (NF-M, vermelho) e α-bungarotoxina-F488 (α-BTX, verde), mostrando placas finais pós-sinápticas (C) fragmentadas e imaturas e resultados de acúmulo de neurofilamentos nos dois grupos de animais (D). N= 4 (n = 34 NMJs) (B) e N = 3 (n = 54 NMJs) (D) animais WT, e N=3 (n = 36 NMJs) Smn2B/- e N = 3 (n = 55 NMJs) foram analisados nos experimentos ColQ Dex2/Dex2 camundongos (B, D). (E, F) Imagens representativas da ocupação terminal axônica em NMJs de AT de camundongos WT e Smn2B/- de 3 semanas de idade, marcados com anticorpos contra glicoproteína 2 da vesícula sináptica (SV2, vermelho) e α-bungarotoxina-F488 (α-BTX, verde) (E), e resultados da ocupação do NMJ (relação volume SV2/AChR) (F). Foram analisados os músculos de N = 3 (n = 50 NMJs) do tipo selvagem e N = 4 (n = 62 NMJs) Smn2B/- camundongos. Os dados são expressos como o valor médio por rato (ponto) ± MEV. As diferenças entre os grupos foram analisadas pelo teste de Mann-Whitney (* p < 0,05). As barras de escala são de 20 μm. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Imagem STED de placas finais pós-sinápticas NMJ. (A) Imagem STED representativa de um NMJ marcado com α-bungarotoxina-F633 (α-BTX) de gastrocnêmio de um camundongo selvagem de 6 semanas de idade mostrando listras AChR pós-juncionais (a barra de escala é de 5 μm). (B) Maior ampliação de uma região com listras AChR (painel inferior) que foi utilizada para gerar o perfil de intensidade. A largura (w) das listras AChR e a distância entre duas listras adjacentes (d) dessa região foram quantificadas e apresentadas no gráfico de barras. Representação esquemática da placa final pós-sináptica para ilustrar a largura (w) e a distância da faixa AChR (d). Esses parâmetros, (C) distância da faixa AChR e (D) largura, foram medidos em camundongos ColQDex2/Dex2 e companheiros de ninhada controle às 6 semanas de idade. NMJs de 5 WT (n total = 29 NMJs) e 6 ColQ Dex2/Dex2 (n total = 43 NMJs) foram analisados cegamente. Os dados são expressos como a média por rato (ponto) ± EPM. As diferenças estatísticas entre os grupos foram analisadas pelo teste de Mann-Whitney (* p < 0,05). Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}
Figura suplementar 1: Lançamento do software LAS X e parâmetros para aquisições confocais. As várias etapas para a aquisição de imagens confocais são descritas nas seções 3.1.2 a 3.1.7 do protocolo. Para cada aquisição de pilha NMJ, um projeto é aberto (etapa 3.1.4) e os parâmetros de tamanho da imagem, velocidade de aquisição, eixos X, Y e Z são selecionados (etapa 3.1.7), com cada varredura sequencial indicada (Seq.1, laser 405 para DAPI; Seq.2, laser 488 para α-BTX-F488; e Seq.3, laser 552 para anticorpos secundários conjugados F594). Clique aqui para baixar este arquivo.
Figura suplementar 2: Lançamento do software e parâmetros LAS X para aquisições STED. As etapas para adquirir imagens STED estão descritas nas seções 3.2.2 a 3.2.8 do protocolo. O microscópio é iniciado no modo de configuração STED ON (etapa 3.2.2) e um projeto é aberto (etapa 3.2.3). Os parâmetros para aquisição de imagem (etapa 3.2.7) (tamanho da imagem, velocidade de aquisição, fator de zoom, eixo X), com cada varredura sequencial são indicados (Seq.1 para α-BTX-F633; Seq.2 para anticorpos secundários conjugados F488). Clique aqui para baixar este arquivo.
Figura suplementar 3: Imagens de pregas juncionais coradas com α-BTX obtidas por microscopia STED. Exemplos de imagem de uma placa final pós-sináptica rotulada com α-BTX-F633 de um rato selvagem de 6 semanas de idade que foram adquiridos com um foco correto (à esquerda) ou incorreto (à direita). Clique aqui para baixar este arquivo.
Figura 4 suplementar: Pop-ups do Windows para descrever os dados de entrada e saída obtidos pelas macros ImageJ personalizadas. Exemplos de dados de entrada (arquivos .tif e .lif) de imagens NMJ são mostrados na coluna da esquerda. Os dados de saída das macros (coluna da direita) são salvos em pastas (Save_Volume, Save_Accu) que contêm imagens da junção (.tif) e folhas de dados contendo os resultados (arquivos .csv). Clique aqui para baixar este arquivo.
Figura 5 Suplementar: Análise de distância e largura da faixa AChR de uma aquisição STED usando o software LAS X. As etapas para analisar as imagens de NMJ STED estão descritas na seção 5 do protocolo. A) Imagem de uma região da placa terminal pós-sináptica marcada contendo listras AChR. A região de interesse para a análise de listras é selecionada desenhando uma linha perpendicular (linha verde, para distância da faixa) ou um retângulo perpendicular (retângulo roxo, para largura da faixa). (B, C) Os perfis de intensidade das regiões selecionadas e as medições para calcular a distância entre as listras AChR (B) e a largura da faixa AChR (C) são mostradas. Clique aqui para baixar este arquivo.
Arquivo de codificação suplementar 1: Macro_NMJ_VOL_Marinelloetal. Macro personalizada ImageJ para extrair medições de parâmetros NMJ (volume NMJ, área da placa final MIP e tortuosidade NMJ). Clique aqui para baixar este arquivo.
Arquivo de codificação suplementar 2: Macro_NMJ_ACCU_Marinelloetal. Macro personalizada ImageJ para extrair acúmulo de NF-M e coloração SV2. Clique aqui para baixar este arquivo.
Discussão
O protocolo de vídeo descrito fornece um método detalhado para quantificar a estrutura 3D das junções neuromusculares, combinando microscopia confocal e STED que pode ser usada para caracterizar alterações patológicas nos níveis pré e pós-sináptico. A alta resolução da microscopia STED permite a visualização e análise morfométrica de nanoestruturas que não são identificáveis por imagens confocais convencionais. Esse procedimento permitiu mensurar alterações estruturais de NMJs em dois músculos apendiculares, tibial anterior e gastrocnêmio, de camundongos CMS relacionados à SMA e ColQ.
Para obter resultados confiáveis com esta técnica, é fundamental dissecar e provocar os músculos adequadamente, prestando especial atenção à fáscia ao redor do músculo e à força aplicada para separar os feixes musculares; caso contrário, o padrão de inervação pode ser interrompido impedindo a avaliação pré-sináptica adequada do NMJ. Embora informações detalhadas sejam fornecidas para analisar os NMJs de AT e GA, em princípio, esse protocolo poderia ser adaptado a outros músculos, incluindo músculos planos, como o diafragma ou o abdome transversal37, que não requerem a etapa de provocação. A fixação tecidual também é crucial para garantir uma coloração de boa qualidade; portanto, recomenda-se o uso de PFA de alta qualidade em um volume apropriado (15-20 vezes o do músculo). Além disso, o tempo de exposição ao fixador é um passo importante porque artefatos, como encolhimento e aglomeração, podem aparecer devido à fixação excessiva e influenciar as características do NMJ. Dado o tamanho das amostras e a taxa de penetração da solução de paraformaldeído nos tecidos38, recomenda-se um tempo de fixação de 18-24 h para este tipo de músculo. Caso a etapa de coloração esteja planejada mais de uma semana após a colheita do tecido, sugere-se manter os músculos fixos em PFA em PBS suplementados com azida de sódio a 4 °C para evitar a proliferação bacteriana.
Este protocolo apresenta uma abordagem utilizando α-BTX-F488 para imagens confocais e α-BTX-F633 para STED. Esses fluoróforos foram escolhidos para se adequarem ao delineamento experimental descrito, mas podem ser modificados de acordo com os equipamentos e materiais disponíveis. Por exemplo, a rotulagem α-BTX F488 pode ser selecionada ao usar um laser STED CW 592 nm para aquisição e quantificação de imagens. No entanto, verifica-se que a configuração que foi aplicada no presente estudo (excitação pulsada fechada STED, depleção de 775 nm) apresenta maior desempenho e melhor resolução do que outras abordagens, como a onda contínua STED39, tornando-a mais adequada para a aplicação atual. Também é importante selecionar cuidadosamente as configurações de potência do laser, especialmente para STED (excitação e esgotamento), uma vez que as características de um perfil de intensidade não podem ser medidas em caso de saturação e, portanto, qualquer sinal saturado em uma imagem NMJ pode comprometer toda a análise.
Este fluxo de trabalho detalhado, incluindo aquisições e análises de imagens usando software de microscópio e macros ImageJ, foi desenvolvido para facilitar a análise morfométrica autônoma de NMJ por microscopia confocal e STED de um único músculo. Fluxos de trabalho descritos anteriormente para análise confocal de NMJ, como NMJ-morph2 ou NMJ-Analyser14, abriram caminho para o projeto de métodos semiautomáticos que facilitam a análise morfológica de NMJs e estudos comparativos. NMJ-morph (e sua versão atualizada aNMJ-morph15) é uma plataforma livre baseada em ImageJ que usa a projeção de intensidade máxima para medir 21 características morfológicas, e NMJ-Analyser usa um script desenvolvido em Python que gera 29 parâmetros relevantes de toda a estrutura NMJ 3D. O limite manual é a única etapa durante o processamento de imagem nesses dois métodos que exigem análise do usuário. Este protocolo integrado detalha as etapas de preparação de tecidos, aquisições de imagens confocais 3D e processamento baseado em ImageJ de NMJs de músculos esqueléticos inteiros e fornece uma visão geral simplificada de cinco parâmetros importantes das placas finais pós-sinápticas (volume, área de projeção máxima e tortuosidade) e pré-sinápticas (ocupação terminal axônica e acúmulo de neurofilamentos). Um parâmetro adicional de relevância biológica, o padrão de organização AChR das pregas juncionais pós-sinápticas, foi incorporado para análise morfométrica em nível nanométrico por microscopia STED de super-resolução (resolução 20-30 nm)40. Curiosamente, o preparo tecidual para imagens STED é mais simples do que outros métodos utilizados para estudos ultraestruturais NMJ, como a microscopia eletrônica de transmissão convencional (MET)9, que é um procedimento bastante complexo e demorado que requer um manipulador qualificado para obter seções ultrafinas da região muscular apropriada. Além disso, dados quantitativos de múltiplas dobras juncionais podem ser obtidos automaticamente usando o software associado ao STED.
Este protocolo foi aplicado para ilustrar defeitos de NMJs previamente conhecidos em músculos deficientes em SMN e ColQ 20,36,41,42. Alterações comuns foram encontradas nos dois modelos de camundongos por microscopia confocal, como diminuição do volume da placa terminal pós-sináptica, área de PImáx e tortuosidade relativa e aumento do acúmulo de neurofilamentos, enquanto alguns achados mais específicos (diminuição da ocupação do NMJ), foram observados apenas em camundongos SMA, como indicador de comprometimento do tráfico de vesículas36. Finalmente, um aumento na distância e largura da faixa AChR foi detectado no ColQ-KO pela análise STED, que são sinais de defeitos ultraestruturais nas pregas juncionais pós-sinápticas, como observado anteriormente pelo MET20. É importante ressaltar que esse protocolo pode ajudar em uma caracterização morfológica mais aprofundada das junções neuromusculares durante o desenvolvimento, manutenção e sob várias condições patológicas.
Divulgações
Os autores declaram não haver conflitos de interesse relacionados a este trabalho.
Agradecimentos
Agradecemos ao "Imaging and Cytometry Core Facility" da Genethon, bem como ao serviço de histologia, que são apoiados em parte por fundos de equipamentos da Região Ile-de-France, do Conseil General de l'Essonne, do Genopole Recherche de Evry, da Universidade de Evry Val d'Essonne e do INSERM, França. Também somos gratos ao Dr. Rashmi Kothary por fornecer a linha de mouse Smn2B / 2B (Universidade de Ottawa, Canadá) e ao Dr. Eric Krejci pela linha de mouse ColQDex2 / + (inédita, Universidade de Paris, França). Agradecemos a Guillaume Corre por seu apoio na análise estatística. Os anticorpos monoclonais 2H3 (desenvolvidos por Jessel, T.M. e Dodd, J.) e SV2 (desenvolvidos por Buckley, K.M.) foram obtidos do Developmental Studies Hybridoma Bank (DSHB), criado pelo NICHD do NIH e mantido na Universidade de Iowa, Departamento de Biologia, Iowa City, IA 52242. Este trabalho foi apoiado pela Association Française contre les Myopathies (AFM-Telethon), pelo INSERM e pela Universidade de Evry Val d'Essonne.
Materiais
| Name | Company | Catalog Number | Comments |
| Buffers and Reagents | |||
| Alexa Fluor 488 goat anti-mouse IgG (F488) | Life Technologies, Thermofisher | A-11001 | |
| Alexa Fluor 488 α-bungarotoxin (F488-a-BTX) | Life Technologies, Thermofisher | B13422 | |
| Alexa Fluor 594 goat anti-mouse IgG (F594) | Life Technologies, Thermofisher | A-11032 | |
| ATTO-633 α-bungarotoxin (F633-a-BTX) | Alomone Labs | B-100-FR | |
| Bovine serum albumin (BSA) | Sigma | A2153 | |
| DAPI Fluoromount-G | Southern Biotech | 00-4959-52 | |
| DPBS | Gibco, Invitrogen | 14190-169 | |
| Ethanol Absolute | VWR | 20821.296 | |
| Immersion Oil, n = 1.518 | THORLABS | MOIL-10LF | Low autofluorescence |
| Neurofilament (NF-M) antibody | DSHB | AB_531793 | |
| Paraformaldehyde (PFA) | MERCK | 1.04005 | |
| Synaptic vesicle glycoprotein 2 (SV2) antibody | DSHB | AB_2315387 | |
| Triton X-100 | Sigma | T8787 | |
| Materials | |||
| Alnico Button cylindrical magnets | Farnell France | E822 | diameter of 19.1 mm with maximal pull of 1.9 Kg |
| 63x 1.4 NA magnitude oil immersion HCX Plan Apo CS objective | Leica Microsystems | ||
| 100x 1.4 NA HC PL APPO CS2 Objective | Zeiss | ||
| Curved thin forceps-Moria iris forceps | Fine Science Tools | 11370-31 | |
| Extra thin scissors - Vannas-Tübingen Spring Scissors | Fine Science Tools | 15-003-08 | |
| Fine serrated forceps | Euronexia | P-95-AA | |
| Gel loading tip round 1-200 µL | COSTAR | 4853 | |
| Leica laser-scanning confocal microscope TCS SP8 | Leica Microsystems | ||
| Leica Laser-scanning confocal microscope TCS SP8 Gated STED 775 nm | Leica Microsystems | ||
| Lens Cleaning Tissue | Whatman (GE Healthcare) | 2105-841 | |
| Medium serrated forceps | Euronexia | P-95-AB | |
| Microscope cover glasses 24x50 nm No 1.5H 170±5 µm | Marienfield | 107222 | High precision |
| Nunclon delta surface (12-well plates) | Thermo Scientific | 150628 | |
| Nunclon delta surface (24-well plates) | Thermo Scientific | 142475 | |
| Safeshield scalpel | Feather | 02.001.40.023 | |
| Sharp-blunt scissors - fine Scissors - Martensitic Stainless Steel | Fine Science Tools | 14094-11 | |
| Superfrost plus slides | Thermo Scientific | J1800AMNZ | |
| Software | |||
| GraphPad | Prism, San Diego (US) | Release N°6.07 | Statistical software |
| ImageJ software | National Institutes of Health | Release N° 1.53f | |
| Leica Application Suite X software | Leica Microsystems | Release N°3.7.2.2283 | Free microscope software available at https://www.leica-microsystems.com/products/microscope-software/p/leica-las-x-ls/downloads/ |
Referências
- Slater, C. R. Postnatal maturation of nerve-muscle junctions in hindlimb muscles of the mouse. Developmental Biology. 94 (1), 11-22 (1982).
- Jones, R. A., et al. NMJ-morph reveals principal components of synaptic morphology influencing structure-function relationships at the neuromuscular junction. Open Biology. 6 (12), (2016).
- Willadt, S., Nash, M., Slater, C. Age-related changes in the structure and function of mammalian neuromuscular junctions. Annals of the New York Academy of Sciences. 1412, 41-53 (2018).
- Boehm, I., et al. Comparative anatomy of the mammalian neuromuscular junction. Journal of Anatomy. 237 (5), 827-836 (2020).
- Nishimune, H., Shigemoto, K. Practical anatomy of the neuromuscular junction in health and disease. Neurologic Clinics. 36 (2), 231-240 (2018).
- Moloney, E. B., de Winter, F., Verhaagen, J. ALS as a distal axonopathy: molecular mechanisms affecting neuromuscular junction stability in the presymptomatic stages of the disease. Frontiers in Neuroscience. 8, (2014).
- Lovering, R. M., Iyer, S. R., Edwards, B., Davies, K. E. Alterations of neuromuscular junctions in Duchenne muscular dystrophy. Neuroscience Letters. 737, 135304 (2020).
- Koneczny, I., Herbst, R. Myasthenia Gravis: Pathogenic effects of autoantibodies on neuromuscular architecture. Cells. 8 (7), 671 (2019).
- Dowling, J. J., et al. Myotubular myopathy and the neuromuscular junction: a novel therapeutic approach from mouse models. Disease Models & Mechanisms. 5 (6), 852-859 (2012).
- Gibbs, E. M., et al. Neuromuscular junction abnormalities in DNM2-related centronuclear myopathy. Journal of Molecular Medicine. 91 (6), 727-737 (2013).
- Swoboda, K. J., et al. Natural history of denervation in SMA: Relation to age, SMN2 copy number, and function. Annals of Neurology. 57 (5), 704-712 (2005).
- Rodríguez Cruz, P. M., Palace, J., Beeson, D. The neuromuscular junction and wide heterogeneity of congenital myasthenic syndromes. International Journal of Molecular Sciences. 19 (6), 1677 (2018).
- Tse, N., et al. The neuromuscular junction: Measuring synapse size, fragmentation and changes in synaptic protein density using confocal fluorescence microscopy. Journal of Visualized Experiments: JoVE. (94), e52220 (2014).
- Mejia Maza, A., et al. NMJ-Analyser identifies subtle early changes in mouse models of neuromuscular disease. Scientific Reports. 11 (1), 12251 (2021).
- Minty, G., et al. aNMJ-morph: a simple macro for rapid analysis of neuromuscular junction morphology. Royal Society Open Science. 7 (4), 200128 (2020).
- Modla, S., Mendonca, J., Czymmek, K. J., Akins, R. E. Identification of neuromuscular junctions by correlative confocal and transmission electron microscopy. Journal of Neuroscience Methods. 191 (2), 158-165 (2010).
- Kittel, R. J., et al. Bruchpilot promotes active zone assembly, Ca2+ channel clustering, and vesicle release. Science. 312 (5776), 1051-1054 (2006).
- York, A. L., Zheng, J. Q. Super-resolution microscopy reveals a nanoscale organization of acetylcholine receptors for trans-synaptic alignment at neuromuscular synapses. eNeuro. 4 (4), (2017).
- Bowerman, M., Murray, L. M., Beauvais, A., Pinheiro, B., Kothary, R. A critical smn threshold in mice dictates onset of an intermediate spinal muscular atrophy phenotype associated with a distinct neuromuscular junction pathology. Neuromuscular Disorders. 22 (3), 263-276 (2012).
- Feng, G., Krejci, E., Molgo, J., Cunningham, J. M., Massoulié, J., Sanes, J. R. Genetic analysis of collagen Q: Roles in acetylcholinesterase and butyrylcholinesterase assembly and in synaptic structure and function. Journal of Cell Biology. 144 (6), 1349-1360 (1999).
- Sigoillot, S. M., et al. Neuromuscular junction immaturity and muscle atrophy are hallmarks of the ColQ-deficient mouse, a model of congenital myasthenic syndrome with acetylcholinesterase deficiency. The FASEB Journal. 30 (6), 2382-2399 (2016).
- Vanhaesebrouck, A. E., Beeson, D. The congenital myasthenic syndromes: expanding genetic and phenotypic spectrums and refining treatment strategies. Current Opinion in Neurology. 32 (5), 696-703 (2019).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Linkert, M., et al. Metadata matters: access to image data in the real world. Journal of Cell Biology. 189 (5), 777-782 (2010).
- Legland, D., Beaugrand, J. Automated clustering of lignocellulosic fibres based on morphometric features and using clustering of variables. Industrial Crops and Products. 45, 253-261 (2013).
- . GitHUb Available from: https://github.com/Genethon/ImCy (2021)
- Otsu, N. A Threshold selection method from gray-level histograms. IEEE Transactions on Systems, Man, and Cybernetics. 9 (1), 62-66 (1979).
- Sanes, J. R., Lichtman, J. W. Induction, assembly, maturation and maintenance of a postsynaptic apparatus. Nature Reviews Neuroscience. 2 (11), 791-805 (2001).
- Kong, L., et al. Impaired synaptic vesicle release and immaturity of neuromuscular junctions in spinal muscular atrophy mice. The Journal of Neuroscience. 29 (3), 842-851 (2009).
- Cifuentes-Diaz, C., et al. Neurofilament accumulation at the motor endplate and lack of axonal sprouting in a spinal muscular atrophy mouse model. Human Molecular Genetics. 11 (12), 1439-1447 (2002).
- Murray, L. M., Comley, L. H., Thomson, D., Parkinson, N., Talbot, K., Gillingwater, T. H. Selective vulnerability of motor neurons and dissociation of pre- and post-synaptic pathology at the neuromuscular junction in mouse models of spinal muscular atrophy. Human Molecular Genetics. 17 (7), 949-962 (2008).
- Boyer, J. G., et al. Myogenic program dysregulation is contributory to disease pathogenesis in spinal muscular atrophy. Human Molecular Genetics. 23 (16), 4249-4259 (2014).
- Ling, K. K. Y., Gibbs, R. M., Feng, Z., Ko, C. -. P. Severe neuromuscular denervation of clinically relevant muscles in a mouse model of spinal muscular atrophy. Human Molecular Genetics. 21 (1), 185-195 (2012).
- Murray, L., Gillingwater, T. H., Kothary, R. Dissection of the transversus abdominis muscle for whole-mount neuromuscular junction analysis. Journal of Visualized Experiments: JoVE. (83), e51162 (2014).
- Baker, J. R. . Principles of biological microtechnique; a study of fixation and dyeing. , (1958).
- Vicidomini, G., et al. STED Nanoscopy with time-gated detection: Theoretical and experimental aspects. PLoS ONE. 8 (1), 054421 (2013).
- Badawi, Y., Nishimune, H. Super-resolution microscopy for analyzing neuromuscular junctions and synapses. Neuroscience Letters. 715, 134644 (2020).
- Thomson, S. R., et al. Morphological characteristics of motor neurons do not determine their relative susceptibility to degeneration in a mouse model of severe spinal muscular atrophy. PLoS ONE. 7 (12), 052605 (2012).
- McMacken, G. M., et al. Salbutamol modifies the neuromuscular junction in a mouse model of ColQ myasthenic syndrome. Human Molecular Genetics. 28 (14), 2339-2351 (2019).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados