
Method Article
Применение Membrane и клеточной стенки селективные флуоресцентные красители для Live-Cell Imaging filamentous Грибы
В этой статье
Резюме
Жизненно важные флуоресцентные красители являются важными инструментами для анализа живой клетки изображения в современной биологии грибковых клеток. В этой статье подробно описано применение установленных и менее известных флуоресцентных красителей для отслеживания динамики плазменной мембраны, эндо-/экзоцитоза и морфогенеза клеточной стенки в нитевидных грибах.
Аннотация
Применение мембраны и клеточной стенки селективных флуоресцентных красителей для анализа живой клетки изображения органеллы динамики в грибковых клетках началось два десятилетия назад и с тех пор продолжает вносить большой вклад в наше понимание нитевидных грибковых Образ жизни. В этой статье содержится практическое руководство по использованию двух мембранных красителей FM 1-43 и FM 4-64 и четырехклеточных стенок пятна Calcofluor White M2R, Solophenyl Flavine 7GFE 500, Pontamine Fast Scarlet 48 и Congo Red. Основное внимание уделяется их низкой дозы применения для выяснения артефактов, свободной окрашивания, их со-изображения свойства, и их количественной оценки. Представленные методы применимы ко всем нитевидным грибковым образцам, которые могут быть подготовлены описанными способами. Фундаментальные подходы к окрашиванию могут служить отправной точкой для адаптации к видам, которые могут потребовать различных условий культивирования. Во-первых, биофизические и биохимические свойства рассматриваются как их понимание имеет важное значение для использования этих красителей, как действительно жизненно важные флуоресцентные пятна. Во-вторых, представлены пошаговые протоколы, в которых подробно описывается подготовка различных грибковых типов образцов для флуоресцентной визуализации живых клеток. Наконец, эксперименты иллюстрируют различные подходы к: (1) выявление дефектов в пространственно-временной организации эндоцитоза у генетических мутантов, (2) сравнительно характеризуют общую и четкую солокализацию целевых белков с маркировкой GFP в эндоцитарном пути, (3) определить морфогенетические дефекты стенки клеток в генетическом мутант, и (4) контролировать биогенеза клеточной стенки в режиме реального времени.
Введение
Двадцать лет назад, способ, в котором гифальный морфогенез и основной молекулярной клеточной биологии может быть визуализирована в нитевидных грибов был революционизирован путем применения мембраны селективного флуоресцентного Fei Mao краситель FM 4-641. Позже, благо хитина-связывающего красителя Calcofluor Белый как жизненно флуоресцентный маркер грибковой динамики стенки клеток была реализована2. С тех пор, как красители и варианты из них стали неотъемлемой частью живой клетки анализа изображений органеллы динамики в грибах, и продолжают предоставлять беспрецедентные идеи в нитевидный грибковый образ жизни. В этой статье подробно описано применение установленных и менее известных флуоресцентных красителей для отслеживания динамики плазменной мембраны, эндо- и экзоцитоза и морфогенеза клеточной стенки в нитевидных грибах. Эндоцитоз отслеживания анализы позволяют различные клетки биологических вопросов, связанных с общим изучением эндоцитоза, которые будут рассмотрены3. Для этого локализация, скорость и последовательность окрашенных отсеков на FM красителя дополнение регистрируется промежуток времени микроскопии и количественно по сравнению между испытаниями грибковых штаммов4. Красители клеточной стенки разграничения внешней границы клетки и позволяют отслеживать морфогенетические события, в том числе поляризованный рост гифального наконечника2, гифаль ветвления5, гифальный синтез6,7 и перегородка формирования8. Кроме того, они облегчают количественную оценку локализованного осаждения стенок клеток и выявление дефектов во время биогенеза клеточной стенки9. Поскольку детальное знание биохимических и биофизических свойств любого флуоресцентного маркера является фундаментальным условием для его успешного применения in vivo, эти характеристики сначала обобщены для шести красителей, описанных в этой статье.
Мембрана селективных красителей
FM (Fei Mao) стирил красители небольшие молекулы амфифилов, которые не могут пройти, но обратимо ассоциировать с внешней листовки липидного двухслойного биологических мембран10. Они практически не флуоресцентные в вквоковом растворе, но становятся интенсивно флуоресцентными при интеграции плазменной мембраны, создавая отличные коэффициенты сигнала к шуму (S/N)-коэффициенты11. Эти свойства делают их идеально подходящими для визуализации плазменной мембраны и внутриклеточной динамики органелл, включая отслеживание эндо- и экзоцитоза12. Зелено-флуоресцентные FM 1-43 и красно-флуоресцентные FM 4-64 являются двумя наиболее широко используемыми флуоресцентными маркерами мембран для этих целей. SynaptoGreen C4 и SynaptoRed C2 являются общими молекулами от альтернативных поставщиков, которые могут быть взаимозаменяемы использованы вместо FM 1-43 и FM 4-64, соответственно.
Стирил красители состоят из трех ключевых структурных областей: (1) липофильный хвост, который облегчает вставку красителя в липидный двухслойный, (2) ядро фторофора, которое определяет спектральные свойства красителя и состоит из двух ароматических колец, соединенных от одного до трех двойных связей, и (3) положительно заряженная гидрофифийная головка, которая предотвращает полную вставку и проницаемость красителя через мембрану (Рисунок 1A).
Чем длиннее липофильный хвост, тем выше гидрофобность красителя и, таким образом, связывающая близость к мембране, но чем ниже его растворимость воды и скорость де-окрашивания мембран. Следовательно, различные варианты FM красителя производят различные динамики окрашивания и моделей. Более высокая гидрофобность C4-хвостой FM 1-43 обеспечивает более сильный и стабильный сигнал флуоресценции на плазменных мембранах и внутренних органеллах быстрее, чем более короткий C2-хвост FM 4-64, при применении при эквимолярных концентрациях(рисунок 2).
Важно отметить, что постоянная и высокая скорость ассоциации/диссоциации обоих FM красителей11 со средним временем удержания 1-6 с на индивидуальную молекулу красителя13 уменьшает шансы на локализованное нарушение мембранной функции, например, за счет изменения текучести мембраны или принудительного постоянного взаимодействия мембранных белков. Это, вероятно, ключевая причина, почему эти молекулы могут быть использованы в качестве жизненно важных красителей. Тем не менее, FM-красителя концентрации выше 50 ММ являются токсичными для грибковых и растительных клеток2,14, и данные из ББ-2 табачных протопластов указывает на то, что более 20 ММ FM краситель привести к насыщению плазменной мембраны14. Таким образом, желательно не превышать этот предел, особенно учитывая тот факт, что отличная визуализация была достигнута всего лишь 2-5 мкм15,16.
Примечательно, что спектральные свойства FM красителей сильно различаются в зависимости от конкретной мембранной микросреды(рассмотрено 14). Как правило, возбуждение и эмиссионные спектры FM красителей в чистых растворителях (как это обычно приводится в информации о продукте) значительно отличаются от того, что в клеточной среде, и в большинстве случаев не могут быть непосредственно консультироваться для выбора настроек изображения живых клеток. Возбуждение / выброс максима FM 1-43 и FM 4-64, например, становятся сине-сдвинуты на 37/46 нм и 43/64 нм, соответственно, при привязке к грибковых мембран по сравнению с их решениями в метаноле (Таблица 1).
Новаторские основы для использования FM 4-64 и FM 1-43 для отслеживания плазменной мембраны, эндо-/экзоцитоза и органеллы динамики, в том числе Спитценкёрпер и митохондрии, уже были всесторонне документированы для широкого спектра нитевидных грибковых видов ранее2,4,17,18,19. Рекомендуемые настройки изображения для обоих FM красителей, которые работают в различных нитевидных грибковых видов изображены на рисунке 1B. Однако технические ограничения имеющегося оборудования или конкретных клеточных и экспериментальных условий, таких, как культурная среда, рН или температура, могут потребовать некоторой адаптации. К счастью, FM красители работают в широком спектральном диапазоне, и очень хорошие результаты изображения достигаются захватывающим FM 1-43 с 514 нм или FM 4-64 с 488 нм. Следовательно, оптимальные настройки визуализации должны определяться индивидуально для каждого типа образца и предполагаемого применения.
Значительный сдвиг Сток более чем 135 нм FM 4-64 позволяет отличный, одновременный совместное изображение с флюорофорами, испускающих зеленый свет; это часто используется для оценки внутриклеточной динамики локализации зеленого флуоресцентного белка (GFP) помечены синтеза белков по отношению к плазменной мембране и эндоцитарный путь9,20.
Сотовые стены селективных красителей
Calcofluor White M2R (CFW), также продаваемый как флуоресцентный светлатель 28, вероятно, самый известный флуоресцентный краситель, используемый для окрашивания клеточных стенок бактерий, грибков, водорослей, высших растений и насекомых. Первоначально использовался в качестве оптического отбеливания агента в бумаге, текстильной и моточной промышленности, его преимущества для клинической диагностики грибковых инфекций была реализована на ранней стадии21,22. Потому что CFW интеркалаты необратимо в зарождающейся цепи хитина он нарушает нормальную хитиновый микрофибрил сборки во время биогенеза клеточной стенки тем самым генерации клеточной стенки стресс23. Это, в свою очередь, вызывает механизм восстановления повреждения клеточной стенки, что приводит к локально повышенному осаждению клеточной стенки в результате глюкан и хитина активации синтаза24,25. Это явление может произойти с любым красителем, который работает при устойчивой связывания с полимерами клеточной стенки, зависит от концентрации и наиболее заметно на hyphal советы, которые представляют собой наиболее плодовитый растет и, следовательно, наиболее чувствительных частей мицелия (Рисунок 3). Всеобъемлющее резюме молекулярного механизма, который реагирует на повреждение клеточной стенки недавно было предоставлено26.
Передозировка красителя в сочетании с фототоксичностью может привести к быстрому клеточному лисису гифальных отсеков(фильм 1). Тем не менее, повышенная чувствительность к концентрациям красителей, которые являются "жизненно важными" в диком типе могут быть использованы для выявления дефектов в биосинтезе клеточной стенки генов потери функции мутантов9. Для CFW и Конго Красный (CR), другой текстильный краситель также известен как Прямой Красный 28 и используется в качестве з-и-хитин-специфических клеточных стенок пятно для грибов и насекомых27,28, пороговые концентрации, которые сильно вызывают хитин синтазов были определены с 60 мкм CFW и Зgt; 70 КМ CR, соответственно, в то время как концентрации злт;15 мкм либо красителя не изменили или препятствуют грибкового роста29,30,31. Хики и др. поместили эту пороговую концентрацию для CFW на уровне 25 мкм2. Таким образом, желательно использовать концентрации красителей No 5 мКм, чтобы исключить связанные со стрессом артефакты и обеспечить использование этих молекул в качестве действительно "жизненно флуоресцентных красителей"2,32. Это в равной степени относится к Solophenyl Flavine 7GFE 500 (SPF) и Pontamine Fast Scarlet 4B (PFS), синонимом Direct Yellow 86 и Direct Red 23, соответственно, двух других полезных красителей клеточной стенки, применение которых для грибов было зарегистрировано впервые более десяти лет назад33. Но, несмотря на их замечательные спектральные свойства34,35, использование обоих красителей с тех пор было очень ограничено36,37. Как было показано ранее для 1,5 мкм CFW2, 2 ММ SPF достаточно для решения динамики стенки ячейки в родных условиях с очень высоким временным разрешением(фильм 2). Такие же результаты можно получить с 2 cr или PFS.
Вместе эти четыре красителя, CFW, SPF, PFS и CR, состоят из набора клеточных стенок селективных флуоресцентных маркеров, которые охватывают почти полный видимый спектр света эмиссии (400-700 нм), используемых на современных флуоресцентных микроскопах(рисунок 4). Значительное увеличение интенсивности флуоресценции при привязке к полимерам клеточной стенки присуще всем четырем и генерирует отличные S/N-коэффициенты. Это, в свою очередь, позволяет сохранить концентрации красителей и возбуждение интенсивности света очень низко йен и позволяет выполнять клеточной стенки окрашивания, как "низкая доза" живой клетки визуализации техники2. Потому что эти красители стенки клетки плазменная мембрана непроницаемая, они одновременно действуют как живые/мертвые пятна. Примечательно, что из-за их чрезвычайно широкого излучения световых спектров, некоторые ограничения в отношении совместной визуализации свойства CFW и SPF с другими флюорофорами должны быть тщательно рассмотрены.
протокол
1. Подготовка грибковых образцов
- Грибковые предкультуры
- Прививать желаемую нагрузку на соответствующую твердую агар-среду, такую как картофельный агар декстроза (PDA) для трикодерма атровирид или минимальный средний фогель (VMM) для Neurospora crassa. Добавьте соответствующий маркер выбора при работе с трансформаторными штаммами.
- Инкубировать предкультуру при оптимальной температуре организма. Например, T. atroviride при 25 градусах Цельсия и N. crassa при 30 градусах Цельсия и 12 h/12 h светлых/темных циклах до тех пор, пока не развился спорный мицелий, но еще не достиг края пластины. На стандартном размере чашка Петри (9,2 см) занимает дикий тип T. atroviride на 4-6 дней в среднем, в то время как N. crassa дикий тип достигает этой стадии после 3-4 дней в среднем.
- Культивирование грибковых колоний
- Используя стерильный скальпель, вырежьте небольшой блок агара 3 мм х 3 мм, несущий неспорующий мицелий, от края колонии докультуры.
- Поместите агар-блок в центре свежей твердой средней пластины, чтобы привить экспериментальную культуру.
- Инкубировать экспериментальную культуру в соответствии с стадией развития, предназначенной для исследовании. Например, дикий тип T. atroviride требует 20-22 часов при 25 градусах Цельсия в темноте для развития колоний диаметром около 2 см на КПК, в то время как дикий тип N. crassa достигает диаметра колонии около 4 см после 14-16 ч инкубации при 30 градусах по Цельсию в темноте на VMM.
ПРИМЕЧАНИЕ: Инкубация в темноте предотвращает образование пигментов, которые могут ввести автофлюоресценцию. Для того, чтобы исключить среднюю фоновому флуоресценцию из экспериментальной культуры, замените агар на 1,5% ж/ч прозрачного затвердевающего агента (см. Таблицу Материалов),и любую сложную среду с определенной минимальной средой.
- Культивирование твердых зародышевых культур
- Используйте 5 мл стерильного физиологического солевого раствора (0,9% w/v NaCl) для сбора кониальных спор из докультурной пластины и собирайте полученную суспензию в 15 мл винтовой крышки.
- Смешайте суспензию хорошо энергичным вихрем и затем отфильтруйте ее на полосе 1 см х 5 см стерильной фильтровой ткани (см. таблицу материалов)слегка фаршированной в 1 мл наконечник пипетки (как собранные, так и автоматически заранее) в свежую стерильную трубку.
- Определите плотность спорок с камерой подсчета клеток и подготовить 1 х 107 клеток / мл суспензии с физиологическим раствором соли.
ПРИМЕЧАНИЕ: Подвеска для разношершие может храниться при 4 градусах Цельсия в течение двух недель. - Приготовьте стандартное блюдо Петри (9,2 см) с 20 мл твердой среды и добавьте 15-20 стерильных стеклянных бусин (3 мм) сверху.
- Пипетка 200 л из суспензии на средней пластине и равномерно распределить клетки по всей пластине нежной тряской. Соберите стеклянные бусы в стакан с 70% этанола для повторного использования.
- Инкубировать экспериментальную культуру в соответствии с стадией развития, предназначенной для исследовании. Например, T. atroviride дикий тип требует 5-6 ч при 25 градусов по Цельсию в темноте, чтобы развивать кониальные зародыши на КПК, в то время как N. crassa дикий тип развивается conidial зародышей после 3-4 ч инкубации при 30 градусов по Цельсию в темноте на VMM.
ПРИМЕЧАНИЕ: Для того, чтобы исключить любой средний фон флуоресценции из экспериментальной культуры, заменить агар с 1,5% ж / v прозрачного затвердевного агента, и любой сложной среде с определенной минимальной среде.
- Культивирование жидких зародышевых культур
- Заполните 190 л жидкой культуры среды в каждом колодце 8-наилучшим камерным микро-слайдом.
- Добавьте 10 кл. 1 х 107 клеток/мл раствора (подготовленного шагами 1.4.1-1.4.3) и перемешайте, аккуратно трубя вверх-вниз несколько раз. В результате общее количество ячеек составляет 1 х 105 на скважину.
- Инкубировать экспериментальную культуру в соответствии с стадией развития, предназначенной для исследовании. Например, дикий тип T. atroviride требует 5-6 ч при 25 градусах Цельсия в темноте для развития кониальных зародышей в картофельном бульоне декстрозы (PDB), в то время как дикий тип N. crassa развивает сяровые зародыши после 3-4 ч инкубации при 30 градусах по Цельсию в темноте в жидком ВММ.
2. Подготовка рабочих растворов красителя
- Чтобы гарантировать полную растворимость каждого красителя, приготовьте 2 мМ штучные растворы в диметилсулькидной сулькости (DMSO), добавив соответствующее количество (см. точный вес в таблице 1) до 1 мл 100% DMSO и хорошо перемешайте вихрем.
ВНИМАНИЕ: Убедитесь в том, чтобы взять DMSO из перегородки запечатанной бутылки; она должна быть прозрачной жидкостью. При контакте с воздухом, DMSO становится коричневым, вероятно, из-за окисления следов примесей, и может негативно повлиять на рост клеток или окрашивания красителя. - Фильтр стерилизовать бульонный раствор через мембранный фильтр шприца 0,2 мкм в свежую стерильную реакционную трубку объемом 1,5 мл. Чтобы свести к минимуму отбеливание красителя, заверните трубку в алюминиевую фольгу.
ПРИМЕЧАНИЕ: Раствор склада красителя можно алицитировать в более малые тома для того чтобы во избежание оттаивать/замораживать циклы, и держать на 4 c на несколько месяцев. - Подготовьте 20 мкм aqueous красителя рабочий раствор путем растворения 2 л раствора красителя запасов в 198 Л стерильной дистиллированной воды в свежей стерильной 1,5 мл реакции трубки. Чтобы свести к минимуму отбеливание красителя, заверните трубку в алюминиевую фольгу.
ПРИМЕЧАНИЕ: Рабочий раствор красителя должен быть приготовлен заново в день эксперимента. - Во время монтажа образца (см. раздел 3) рабочий раствор красителя будет стандартно разбавлен 1:10, что приведет к окончательной концентрации красителя в размере 2 мкм и 0,1% w/v окончательной концентрации DMSO.
ПРИМЕЧАНИЕ: Выбор различных факторов разбавления, просто изменив соотношение громкости между раствором для работы красителя и монтажной жидкостью, позволяет легко адаптировать нужную конечную концентрацию красителя.
ВНИМАНИЕ: Для предотвращения нежелательных эффектов из-за красителя или токсичности DMSO, коэффициент разбавления не должен подпадать под 1:4, чтобы привести к максимальной конечной концентрации 5 ММ красителя и 0,4% w/v DMSO. Более высокие концентрации красителей быстро насыщают систему и предотвращают надежную количественную оценку сигнала, в то время как более 0,5% w/v (62,5 мМ) DMSO может нарушить развитие клеток38.
3. Подготовка образца для микроскопии
- Маунт образцы из грибковых колоний (шаг 1.2) или твердых зародышевых культур (шаг 1.3) по перевернутой агар блока метода.
- Держите чистый 24 мм х 60 мм стеклянной крышкой скольжения (#1 0,13-0,16 мм толщиной) готовы и добавить 18 л жидкой минимальной среде (VMM или M9) или физиологический солевой раствор на центр.
- Добавьте 2 злицы из 20 мм красителя рабочего раствора к 18 л жидкости и хорошо перемешайте, несколько раз промешивая вверх и вниз, избегая при этом производства пузырьков воздуха.
ПРИМЕЧАНИЕ: При работе с несколькими образцами, желательно подготовить мастер смесь раствора жидкого красителя для всех, чтобы обеспечить равную концентрацию красителя на протяжении всего эксперимента. - С помощью чистого скальпеля вырежьте образец 15 мм х 15 мм с периферии колонии или твердой зародышевой культуры и поместите его вертикально рядом со средним падением на крышку скольжения.
- Использование скальпеля для поддержки верхнего края блока и палец, чтобы держать заднюю сторону блока на месте, медленно опустите сторону проведения мицелия или зародышей на жидкость. Образец готов к передаче на стадию микроскопа.
ВНИМАНИЕ: Важно делать это медленно и очень осторожно, чтобы свести к минимуму механическую нагрузку на клетки и избежать пузырьков воздуха, оказавшихся между образцом и крышкой скольжения.
- Смонтировать жидкие зародышевые культуры со ступени 1.4.
ПРИМЕЧАНИЕ: Наиболее удобно, жидкие зародышевые культуры в камерных микроколодцы слайды могут быть непосредственно переданы и дальнейшего манипулировать на стадии микроскопа.- Добавьте 22 л рабочего раствора красителя к 200 л жидкой среды, чтобы привести к стандартным конечным концентрациям 2 мм красителя и 0,1% w/v DMSO.
ПРИМЕЧАНИЕ: Жидкие зародышевые культуры имеют большое преимущество, что флуоресцентные красители (или другие химические вещества, такие как ингибиторы) могут быть добавлены в любой нужный момент времени эксперимента, а также во время записи. В этом случае, специальная осторожность должна быть принята для введения жидких капель очень медленно, чтобы не беспокоить клетки. Вибрации системы и броуновское движение могут уже ввести некоторые движения клеток.
- Добавьте 22 л рабочего раствора красителя к 200 л жидкой среды, чтобы привести к стандартным конечным концентрациям 2 мм красителя и 0,1% w/v DMSO.
4. Микроскопия живых клеток
- Отрегулируйте основные настройки приобретения изображения. Следующие настройки приобретения изображения позволяют запечатлеть динамику окрашивания в отдельных гифах и применимы к обоим следующим анализам
- Нанесите 5-10% лазерной мощности 20% от полной мощности устройства.
- Используйте план Apo 60x-63x глицерол или цель погружения воды с высокой численной диафрагмой No 1.2.
- Ограничьте область приобретения изображения контуром гифа, установив размер изображения 1024 x 256 пикселей и используя оптический коэффициент масштабирования 2-3.
- Используйте двунаправленное сканирование с 400 Гц. Отрегулируйте размер пинхола до 1 кондиционера.
- Установите усиление наиболее чувствительного детектора до 100%.
- Для записи временных кругов, начать приобретение изображения с одного кадра каждые 15 с, чтобы разумное временное разрешение без производства отбеливания красителя или фото стресса.
- Для 3D-записи установите верхний и нижний пространственный предел границы гифаев и космических оптических секций 1 мкм друг от друга, чтобы обеспечить разумное пространственное разрешение.
ПРИМЕЧАНИЕ: Из-за быстрого роста гиф, высокое пространственное разрешение в оси часто приносится в жертву для высокого временного разрешения в оси X/Y или наоборот. Только очень современные конфокальные лазерные сканирующие микроскопы достаточно быстры, чтобы удовлетворить оба требования.
- Эндоцитоз поглощения анализы
- Проконсультируйтесь по рисунку 1 и таблице 1, чтобы определить наилучшие настройки возбуждения/выбросов для FM 1-43 и/или FM 4-64, доступных в системе микроскопии, и соответствующим образом отрегулировать их.
ПРИМЕЧАНИЕ: С рекомендуемой концентрацией 2 ММ, включение FM-красителя в плазменной мембране мгновенно в нормальных здоровых клеток. Весь процесс от первоначального окрашивания плазменной мембраны до появления красителя в трубчатых вакуолах обычно завершается в течение 30-45 минут при комнатной температуре. Увеличение концентрации FM красителя увеличивает S/ N-коэффициент и, таким образом, производит более высокие контрастные изображения быстрее. Тем не менее, это также ускоряет процесс маркировки, что затрудняет дифференцировать хронологическую последовательность окрашивания органелл. - Начните запись изображения, используя рекомендуемые выше настройки приобретения изображений, и оцените результаты.
- Оптимизируйте настройки приобретения изображения с пространственным и временным разрешением, необходимым для захвата аспекта плазменной мембраны или динамики эндоцитоза, на котором сосредоточен эксперимент.
- Например, для того, чтобы захватить очень быструю динамику в X/Y, уменьшить общий размер изображения, изображение только одного фокусного плоскости и увеличить скорость сканирования до 1 fps. Для более высокого разрешения в оси, уменьшить разрешение в X/ Y, уменьшить размер изображения и уменьшить расстояние между оптическими секциями до 0,5 мкм.
- Проконсультируйтесь по рисунку 1 и таблице 1, чтобы определить наилучшие настройки возбуждения/выбросов для FM 1-43 и/или FM 4-64, доступных в системе микроскопии, и соответствующим образом отрегулировать их.
- Динамика клеточной стены
- Проконсультируйтесь с рисунком 4 и таблицей 1, чтобы определить наилучшие настройки возбуждения/выбросов для прикладного клеточного красила, доступного в системе микроскопии, и соответствующим образом отрегулировать их.
ПРИМЕЧАНИЕ: Из-за их широкой спектры выбросов, CFW и SPF не очень хорошо подходят для одновременного совместного изображения с другими флюорофорами, преимущественно GFP. Некоторые ограничения даже применяются для последовательных подходов изображения с этими красителями, и, таким образом, должны быть оптимизированы индивидуально. - Начните запись изображения, используя рекомендуемые выше настройки приобретения изображений, и оцените результаты.
ПРИМЕЧАНИЕ: С рекомендуемой концентрацией 2 ММ, включение красителя в клеточной стенке не обязательно мгновенно, но достаточно быстро. Весь процесс образования перегородки, например, занимает в среднем около 5-7 мин при комнатной температуре20. Увеличение концентрации красителя стенки клетки увеличивает S/N-коэффициент и таким образом производит более высокие изображения контраста более быстро. Тем не менее, он также быстро вводит артефакты из-за индуцированного ремонта повреждения стенки клетки. - Оптимизация параметров приобретения изображения для пространственного и временного разрешения, необходимого для захвата аспект амортогенеза клеточной стенки, на котором эксперимент сосредоточен, как указано в разделе 4.2.3.
- Проконсультируйтесь с рисунком 4 и таблицей 1, чтобы определить наилучшие настройки возбуждения/выбросов для прикладного клеточного красила, доступного в системе микроскопии, и соответствующим образом отрегулировать их.
Результаты
Количественный анализ изображений
Помимо «простой» визуализации клеточных процессов, живая клеточная визуализация позволяет извлечь количественную информацию из записанных данных. Как правило, количественный анализ изображений является сложной темой, надлежащее обсуждение которой выходит далеко за рамки этой статьи, следовательно, читатель относится к выделенным учебникам и статьям39,40,41. Тем не менее приводятся некоторые основные руководящие принципы, связанные со следующими примерами данных. Необходимо выполнить ряд важнейших предпосылок для количественной оценки изображений, в том числе: (1) определенные моляриты флуоресцентных красителей должны применяться ко всем образцам, с тем чтобы обеспечить точное относительное сравнение; (2) настройки приобретения изображения должны быть скорректированы таким образом, чтобы детекторы света излучения никогда не насыщались, в противном случае максимальная интенсивность отрезана; (3) настройки приобретения изображений должны оставаться фиксированными в ходе согласованного экспериментального набора, в противном случае вводятся искусственные изменения интенсивности; (4) данные изображений должны быть сохранены в формате безинформационно-необлагаемых файлов вместе с мета-информацией, содержащей все настройки инструмента; и (5) анализ изображений должен ограничиваться минимальным числом шагов по постобработке, необходимых для извлечения желаемой количественной информации.
Как правило, определенные стандарты, которые позволили бы абсолютную количественную оценку зарегистрированных сигналов, не доступны в живой клетке. Таким образом, в своей простейшой форме количественный анализ изображений основывается на относительном сравнении интенсивности пикселей в одном изображении или между различными изображениями, записанными с одинаковыми настройками. Программное обеспечение для управления микроскопом производителя обычно включает в себя основные инструменты для изображения после обработки и количественного анализа, или может быть обновлено с дополнительными функциями для сегментации изображений, порогового значения, изображения соотношения и т.д. Доступно несколько платформ обработки изображений с открытым исходным кодом, которые по-разному подходят для различных типов данных изображений, включая ImageJ (https://imagej.net; https://imagej.nih.gov/ij/), ледяную (http://icy.bioimageanalysis.org/), CMEIAS Bioimage Informatics (http://cme.msu.edu/cmeias/) и Wimasis (https://www.wimasis.com/en/).
Представленные примеры данных были обработаны и проанализированы с помощью платформы ImageJ. Короче говоря, конкретные области в ячейке, такие как вершина гифалия наконечника или септа, отмечены значительными инструментами выбора области, и интенсивность всех содержащихся пикселей считывается с программным обеспечением реализован "инструмент измерения". Данные интенсивности элементов управления и экспериментальных образцов передаются в файл электронной таблицы, математически анализируются и готовятся в виде графика. Более подробную информацию можно найти в цитируемых оригинальных публикациях.
Пример данных 1: FM 4-64 поглощения анализы
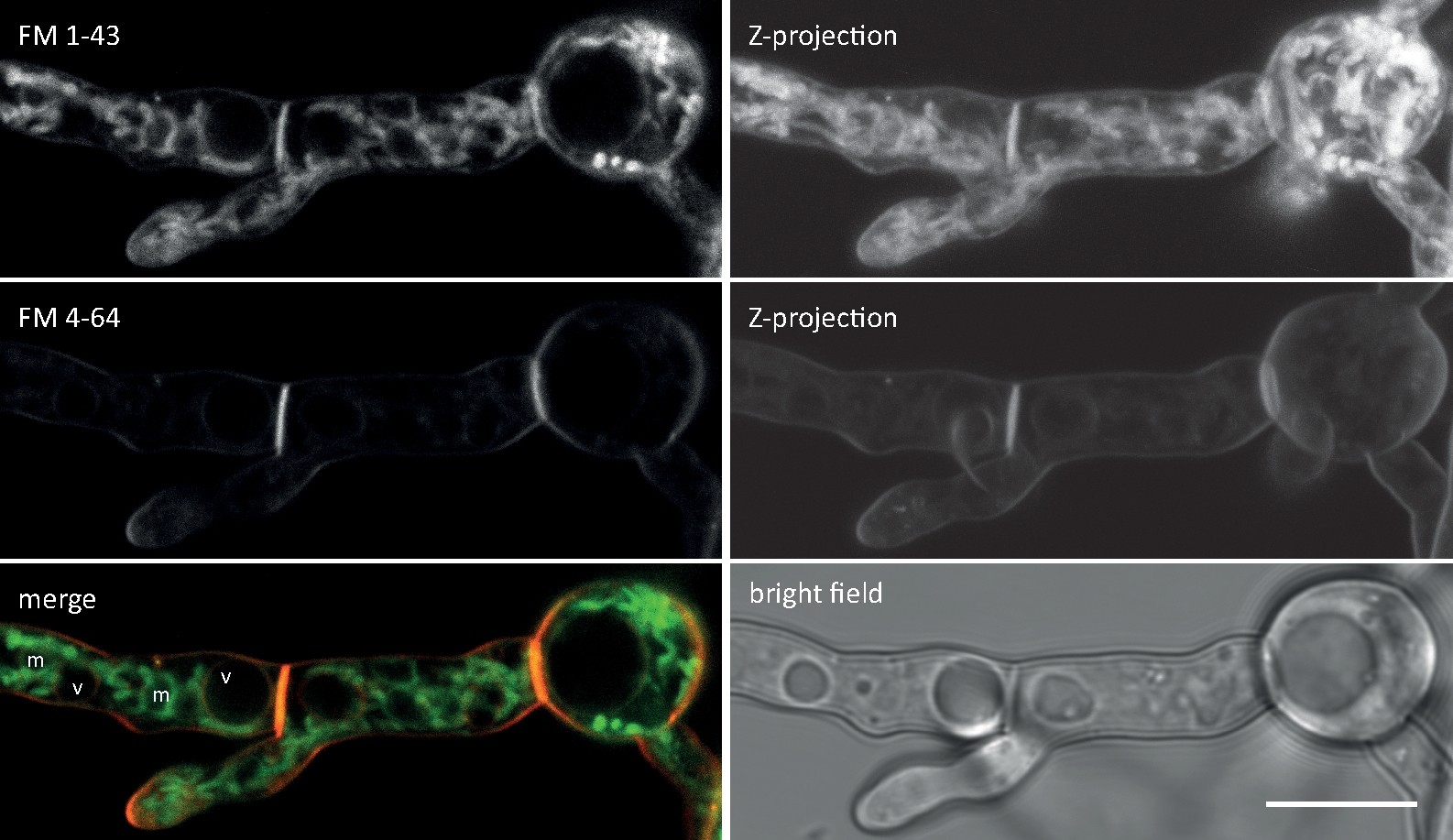
Грибковые образцы культивировались как колонии (шаг 1.2) и смонтированы инвертированным методом агарного блока (шаг 3.1). Окончательная концентрация FM 4-64 составила 1,67 мкм. Настройки визуализации: HCX PL APO 63x/1.3 NA glycerol цель погружения на перевернутый конфокальный лазерный сканирующий микроскоп (см. Таблицу Материалов); FM 4-64 возбуждение на 488 нм и выбросы на 600-700 нм; один кадр каждую минуту на срок до 150 мин. FM 4-64 поглощения анализы определены дефекты в пространственно-временной организации эндоцитоза в делегации генов и гена overexpressing мутантов грибковых конкретных Sur7-семейный белок 2 (Sfp2) T. атровирид9 (Рисунок 5).
Пример данных 2: FM 4-64 совместное окрашивание флуоресцентных белков синтеза, ориентированных на эндоцитарные отсеки
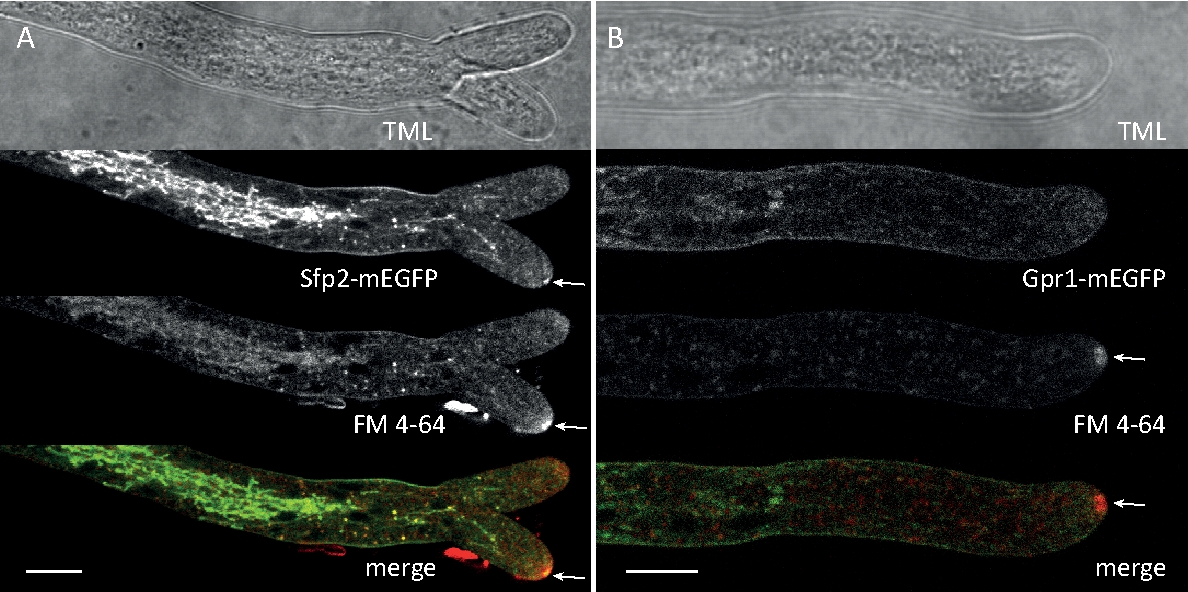
Грибковые образцы культивировались как колонии (шаг 1.2) и смонтированы инвертированным методом агарного блока (шаг 3.1). Окончательная концентрация FM 4-64 составила 2 мКМ. Настройки визуализации: CFI Plan Apo VC 60x/1.2 NA XC цель погружения воды на перевернутом конфокальном лазерном сканирующем микроскопе (см. Таблицу Материалов); Возбуждение GFP при 488 нм и выбросна при 500-530 нм, FM 4-64 возбуждение на 488 нм и выброс на 600-700 нм, и яркое поле с передаваемым светом детектор, все одновременно; один кадр каждые 15 с на срок до 15 мин. FM4-64 совместное окрашивание было использовано для связи субклеточного распределения двух расширенных зеленых флуоресцентных белков (EGFP) помечены трансмембранных белков Sfp2 и Gpr1 к эндоцитарной пути в T. atroviride (Рисунок 6, Фильм 3, Фильм 4).
Пример данных 3: FM 4-64 совместное окрашивание для выявления морфогенетических различий
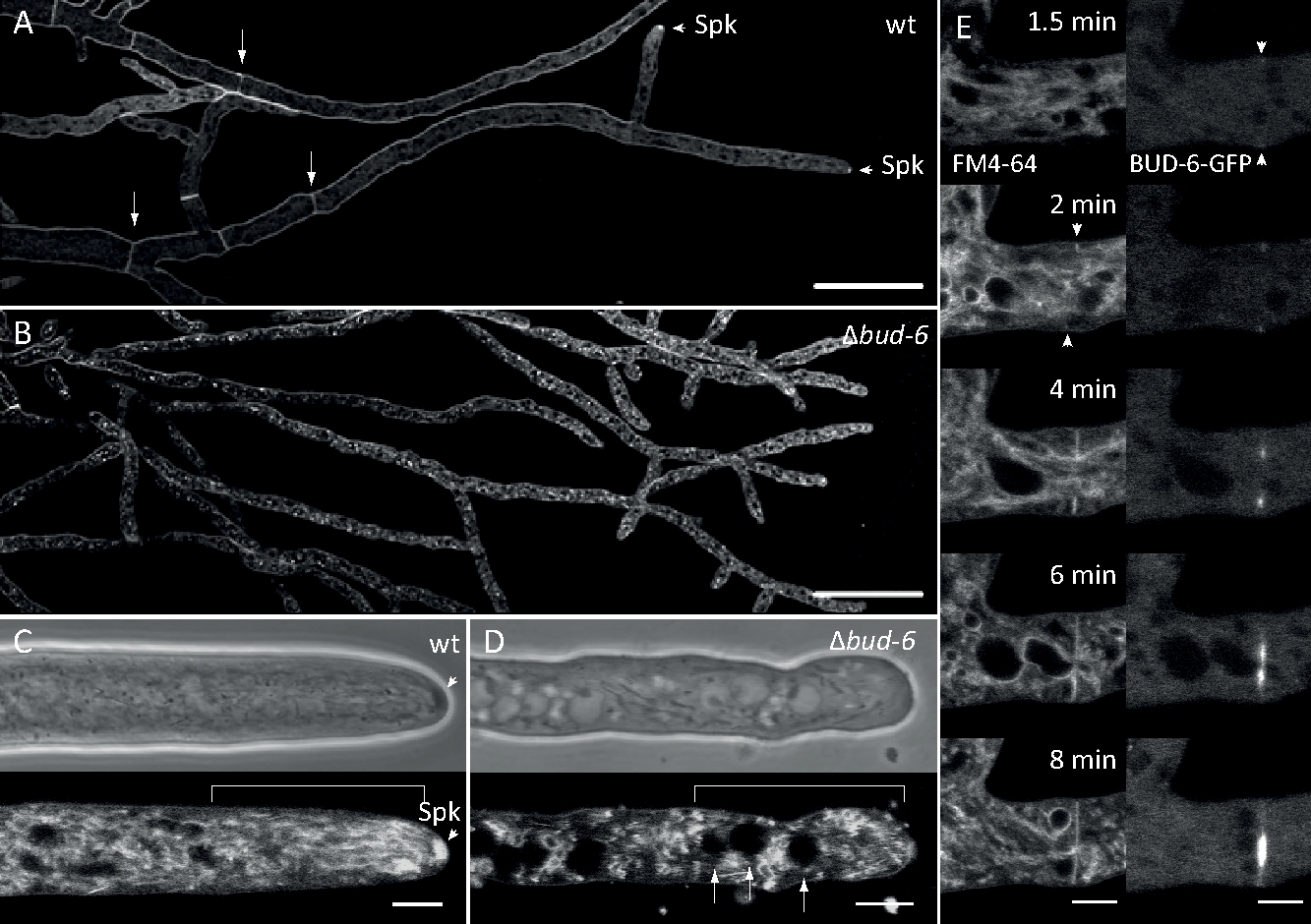
Грибковые образцы культивировались как колонии (шаг 1.2) и смонтированы инвертированным методом агарного блока (шаг 3.1). Окончательная концентрация FM 4-64 составила 2 мКМ. Настройки визуализации: План Апохромат 63x/1.4 NA цель погружения масла на перевернутый конфокальный лазерный сканирующий микроскоп (см. Таблица материалов); Возбуждение GFP при 488 нм и выбросна при 505-550 нм, FM 4-62 возбуждение на 488 нм и выброс на 574-691 нм, и яркое поле с передаваемым светом детектор, все одновременно; один кадр каждые 8,5 с на срок до 15 минут. FM4-64 совместное окрашивание позволило связать динамику субклеточной локализации флуоресцентно помечены BUD-6 polarisome сложный белок эндосомы торговли зависимых процессов, таких как формирование септы и поляризованных гифальный рост наконечника, и характеризуется различия в субклеточной организации и гифальной архитектуры между диким типом и мутант штаммов N. crassa (Рисунок 7, Фильм 5).
Пример данных 4: Окрашивание клеточной стенки выявляет морфогенетические различия
Грибковые образцы культивировались как колонии (шаг 1.2) и смонтированы инвертированным методом агарного блока (шаг 3.1). Использовались окончательные концентрации 2 МКм CFW, 20 МКм SPF и 100 мкм CR. Настройки изображения: CFI Plan Apo VC 60x/1.2 NA XC цель погружения воды на перевернутом конфокальном лазерном сканирующем микроскопе (см. Таблицу Материалов); CFW и SPF возбуждение на 405 нм и выбросы на 430-470 нм, CR возбуждение на 543 нм и выбросы на 580-620 нм. Различные свойства взаимодействия CFW, SPF и CR с полимерами клеточной стенки подчеркивают морфогенетические различия между мутантомsfp2 и штаммом дикого типа T. atroviride9. Увеличение стресса клеточной стенки, вызванного повышенной концентрацией красителей происходит быстрее и более выраженным у мутанта по сравнению с диким типом. Кроме того, одни и те же изображения позволяют количественно очислить морфогенетические различия в диаметре гифаля и расстоянии перегородки между обоими штаммами(рисунок 8).
Пример данных 5: Мониторинг биосинтеза клеточной стенки в режиме реального времени
Зародышицы культивировались как жидкая культура (шаг 1.4) в 8-колодцах камерных микро-слайдов (шаг 3.2). Окончательная концентрация CFW составила 0,12 мкм. Настройки визуализации: CFI Plan Apo VC 60x/1.2 NA XC цель погружения воды на перевернутом конфокальном лазерном сканирующем микроскопе; CFW возбуждение на 405 нм и выбросы на 420-470 нм; один кадр каждые 20 с на срок до 35 минут. Очень низкая концентрация CFW предотвращает насыщение клеточной стенки молекулами красителя и позволяет осуществлять количественный мониторинг биосинтеза клеточной стенки в режиме реального времени. Это показывает, что осаждение нового материала стенки клетки не является однородным, но очень быстро реагирует на локализованные физические нагрузки в результате относительного смещения одной клетки на клеточной привязанности до зародывания слияния в N. crassa (Рисунок 9, Фильм 6).

Рисунок 1: Биохимические и биофизические свойства FM красителей. ()Химические структуры FM 1-43/SynaptoGreen C4 и FM 4-64/SynaptoRed C2. (B) Абсорбция и спектры выбросов обоих FM красителей, накладные с оптимальными настройками изображения для мембранных красителей в нитевидных грибов: 445-495 нм синий свет будет возбуждать FM 1-43 с 100-80% эффективности, в то время как 488 нм лазера Аргона будет возбуждать краситель с 91% эффективности. Из-за синей сдвига на мембранной связывании (яп. ) оптимальный диапазон обнаружения выбросов FM 1-43 составляет 520-590 нм. Аналогичным образом, оптимальные настройки изображения для FM 4-64 в грибах 471-541 нм (100-80% эффективности) при использовании полихромного источника света возбуждения или 514 нм (99% эффективности) при использовании лазера Аргон, и 650-750 нм для обнаружения света выбросов. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: Одновременное совместное изображение FM 1-43 и FM 4-64. Эквимолярная смесь обоих красителей была добавлена в культуру жидких зародышевых N. crassa, приносящую окончательную концентрацию 10 мкм. На 25 мин после красителя того, FM 1-43 запятнал плазменную мембрану и уже накапливается во внутренних мембранах, в том числе сильно окрашенных митохондрий (м), но в значительной степени за исключением вакуолярных мембран (v), и более чем в восемь раз сильнее по сравнению с FM 4-64 (средняя интенсивность флуоресценции от 176 до 21, соответственно), чья более низкая гидрофобность / более высокая гидрофилийность замедляет его внутреннюю скорость, ведущую к длительному времени. Шкала бар 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

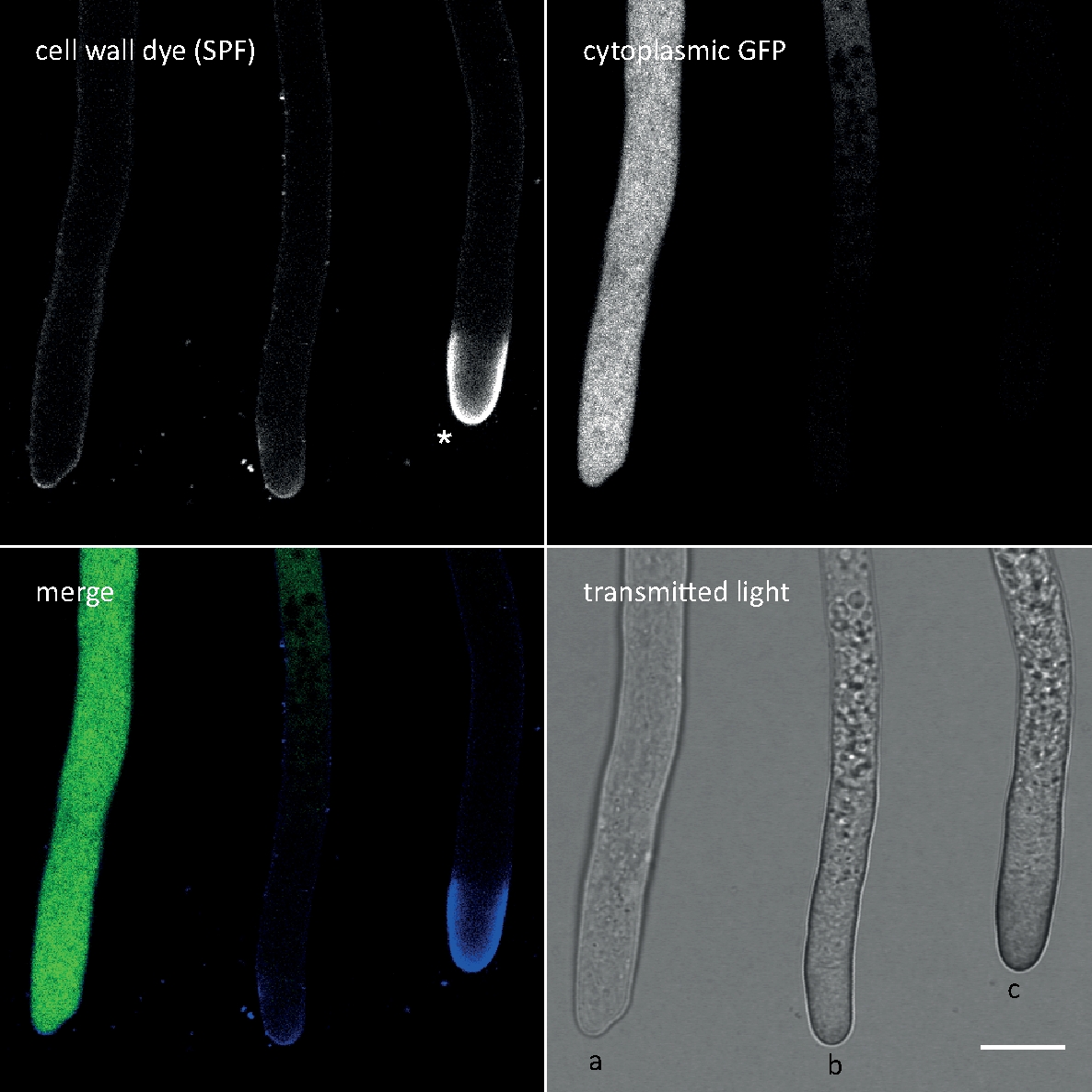
Рисунок 3: Клеточная стенка стресс-индуцированного осаждения полимеров клеточной стены на вершине кончика часто влечет за собой автолиза клеток. Hyphae T. atroviride, выражающий цитоплазмический GFP, были окрашены 10 мкм Solophenyl Flavine 7GFE 500 (SPF) и изображены сразу после монтажа. Не-stressed гифы (a), подчеркнул гифы со слегка увеличенным осаждением глюкана/хитина на вершине и прогрессирующей автолизом (b), и сильно подчеркнутыми гифами с выраженным атическим глюканом/хитиновой крышкой (звездочка) и терминальной автолизом (с), очевидной полной потерей фуфесценции GFP и обширной вакуолизации. Шкала бар 10 мкм. Смотрите фильм 1 для полной последовательности курса. Обратите внимание, что три гифа действительно были расположены рядом друг с другом. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 4: Биохимические и биофизические свойства селективных красителей клеточной стенки. Химические особенности даны, что из натрия соли каждого красителя. Абсорбционные и эмиссионные спектры соответствуют спектрам клеток. Показанные монохромные лазерные линии возбуждения (написанные в цвете), полихромные диапазоны возбуждения, применимые для эпифлюоресцентных микроскопов, и диапазоны обнаружения света на выбросах рекомендуются для визуализации в нитевидных грибах. Две линии лазерного возбуждения указаны, когда обе работают одинаково хорошо. (*) Спектр выбросов клеточных стенных ПФС значительно более красно-сдвинуты, чем ранее отмечалось33, однако, в результате чего очень хорошие S / N-коэффициенты с более низкой концентрации красителя, чем раньше. Полный спектр CR в настоящее время недоступен, следовательно, что Нила Красного (CAS No: 7385-67-3) показано, как ближайший матч. Подробную информацию о спектральных свойствах ЧР можно найти в другом месте42. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 5: Влияние Sfp2 на эндоцитарное поглощение FM4-64. Три последовательных ключевых этапа поглощения FM красителя легко различимы в диком типе (WT). Этап I: эксклюзивное окрашивание плазменной мембраны, II этап: первое появление FM-красителя в эндоцитарных пузырьках, а iii этап: эксклюзивное окрашивание эндоцитарных пузырьков и эндомембран. Эквивалентные узоры окрашивания показаны в самый ранний момент их появления. По сравнению с T. atroviride дикого типа, эндоцитоз немного ускоряется в sfp2 чрезмерно-выражения мутант (OEsfp2), в то время как поглощение красителя резко задерживается в sfp2 удаления мутанта (sfp2 ). Например, поглощение красителя в плазменной мембране происходит мгновенно в OEsfp2, но занимает 2 минуты в диком типе; и полная интернализация FM-красителя из плазменной мембраны происходит в 10 раз быстрее в OEsfp2 по сравнению сsfp2. Шкала баров 5 мкм. Рисунок воспроизводится с Atanasova et al.9 в согласии с лицензией Creative Commons (https://creativecommons.org/licenses/by/4.0/). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 6: Совместное окрашивание мембранных белков с маркировкой EGFP с FM 4-64 способствует дифференциации различных динамики субклеточной локализации в T. atroviride. (A) Четырех-трансмембранный домен белка Sfp2 совместно локализуется с FM4-64 помечены органеллы, в том числе плазменной мембраны и септы, Спитценкёрпер (Spk; стрелка) и предполагаемых трубчатых vacuoles. (B) GPCR-как семь-трансмембранный белок Gpr1 совместно локализуется с FM4-64 в те же органеллы, как Sfp2, за исключением Spk. Шкала баров, 10 мкм. Смотрите фильм 3 и фильм 4 для полного времени курс последовательности. Цифра была изменена с Atanasova et al.9 в согласии с лицензией Creative Commons. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 7: FM 4-64 окрашивание отличаетбут-6 мутанта от дикого типа, и локализует BUD-6 на переплетном кольце. (A) FM 4-64 окрашивание гифы N. crassa дикого типа (стрелки указывают септа; наконечники стрел указывают на Spk). (B) Септа и Spk отсутствуют вБут-6. Шкала баров, 50 мкм. (C и D) Крупным планом гифальной вершины и subapex дикого типа (C) иБут-6 (D). Spk (стрелка) дифференцируется в диком типе, но небутон-6. Кронштейны указывают на ядерную зону отчуждения, не установленную в«Бут-6». Шкала баров 5 мкм. (E) BUD-6-GFP набора в зарождающейся септации сайте, предшествующем плазменной мембраны инвагинации (стрелы) и сопровождающих сужения перегородки. Шкала баров 5 мкм. Смотрите фильм 5a и movie 5b для полного последовательности курса времени. Рисунок был воспроизведен с изменениями из Lichius et al.16 в согласии с лицензией Creative Commons. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 8: Удаление sfp2 изменяет структуру осаждения материала клеточной стенки и влияет на гифальный морфогенез Т. атровирида. (A) CFW и SPF окрашивания показывает увеличение осаждения стенки клетки вsfp2 (стрелки) по сравнению с диким типом (WT). CR окрашивания вызывает обширный отек кончика только вsfp2 (стрелы). Шкала баров No 10 мкм. (B) Морфогенетические дефекты вsfp2 включают в себя значительно сниженные расстояния перегородки(sfp2 - 26,0 мкм, дикий тип No 85 мкм; n й 60; ANOVA Pr slt; 2-16) и меньшие диаметры гифального(sfp2 - 5,6 м/с, дикий тип - 12,6 м/с; n - 100; ANOVA Pr злт; 2/16). (C) Увеличение красителя флуоресценции в вершине кончика по сравнению с subapex. Шкала бар No 5 мкм. (D) Интенсивность кодированных 3D поверхности (C). (E) Количественная оценка интенсивности относительной флуоресценции вsfp2 и диком типе (n No 55). Рисунок был воспроизведен из Атанасовой и др.9 в согласии с Лицензией Creative Commons. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 9: Мониторинг биосинтеза клеточной стенки в режиме реального времени. (A) Конидиальный анастомоз трубки (CAT) слияние между зародышей N. crassa. Физический контакт проявляется в результате реакции крутящего момента (21–28 мин). Интенсивность цветной флуоресценции CFW указывает на области с небольшим (темно-синим) и интенсивным (желтым) осаждением клеточной стенки. Первоначально неокрашенные самонаводящиеся наконечник (стрелка), откладывает новый материал стенки клетки на контакте кончика и в области испытывают наибольшее физическое напряжение (стрелка). Шкала бар 5 мкм. Смотрите фильм 6 для полной последовательности курса. Рисунок,воспроизведенный из 38 с разрешения. (B) Проекция (A) с указанием четырех круговых областей, в которых интенсивность флуоресценции были измерены. Шкала бар 5 мкм. (C) Участок указанных регионов, показывающий быстрое увеличение локализованного биосинтеза клеточной стенки в ответ на физический стресс (CAT 1, стрелка). В зародышевой трубе (GT) и тело споры (конидий), биосинтез стенки клетки увеличивает устойчиво. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Фильм 1: Краситель-индуцированной клеточной стенки стресса. 10 ММ SPF (циан) были добавлены в T. atroviride гифы, выражающие цитоплазмический GFP (magenta). Обширное окрашивание кончиков происходит немедленно, с последующим быстрым лисисом гифальных отсеков в течение 2 мин; очевидно исчезновение GFP флуоресценции. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.

Фильм 2: Vital SPF окрашивания. 2 ЗМ SPF (циан) позволяют отслеживать рост кончика T. atroviride hyphae с высоким пространственным и временным разрешением, не вызывая клеточной стенки стресс артефактов на вершине кончика. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.

Фильм 3: FM 4-64 совместное окрашивание Sfp2-GFP. Совместное окрашивание T. atroviride, выражающее Sfp2-mEGFP (зеленый) с 1,67 мкм FM 4-64 (красный) выявить перекрывающийся и отчетливый локализации мембранного белка с эндоцитарным путем. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.

Фильм 4: FM 4-64 совместное окрашивание Gpr1-GFP. Совместное окрашивание T. atroviride, выражающего Gpr1-mEGFP (зеленый) с 1,67 мкм FM 4-64 (красный) выявить перекрывающиеся и различные локализации мембранного белка с эндоцитарным путем. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.

Фильм 5: FM 4-64 совместное окрашивание BUD-6-GFP. (5a) Совместное окрашивание N. crassa, выражающего BUD-6-GFP (зеленый) с 2 мкм FM 4-64 (красный) позволяют отслеживать динамику BUD-6 во время формирования перегородки относительно сопутствующей плазменной мембраны инвагинации. (5b) Обрезанные и слияния изображения (5a). Пожалуйста, нажмите здесь, чтобы скачать фильм 5a
Пожалуйста, нажмите здесь, чтобы скачать фильм 5b.

Фильм 6: Мониторинг биосинтеза клеточной стенки в режиме реального времени. N. crassa зародыши, выражающие MAK-1-GFP (зеленый) были совместно окрашены с 0,12 мкм CFW (синий), чтобы выявить локализованный биогенеза клеточной стенки во время CAT-опосредованного зародыша слияния. Обратите внимание, что есть некоторое кровотечение через сигнал CFW в GFP каналов, иллюстрирующие, что SPF или CR являются лучшим выбором, как последовательные и одновременные ко-изображения красителей для GFP, соответственно. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.
Таблица 1: Свойства мембраны и клеточной стенки селективных флуоресцентных красителей. - значения мг/мл, скорректированные для снижения содержания чистоты/красителя, что приведет к эквимолярности во всех растворах; n.i.a. - Нет информации. Пожалуйста, нажмите здесь, чтобы загрузить эту таблицу.
Обсуждение
Эта статья продолжает новаторскую работу, которая установила использование различных флуоресцентных красителей в качестве жизненно важных маркеров органеллы для нитевидных грибов в начале 2000-х2,4,43, и попытки обсудить фотофизические и клеточные биологические свойства FM красителей и отдельных красителей клеточной стенки более подробно, чем раньше. Особенно в отношении нежелательных клеточных эффектов, таких как насыщение мембраны или повреждение клеточной стенки, которые происходят выше определенных концентраций красителя. То, что раньше считалось нетоксичным на клеточном уровне, в настоящее время считается токсичным на молекулярном уровне. Даже если эти эффекты могут быть очень тонкими и непосредственно не очевидны очевидные изменения в органеллы или клеточного поведения, любое возможное вмешательство применения красителя, кроме визуализации, должно быть сведено к минимуму для исследования родной молекулярной функции. К счастью, улучшенная чувствительность и квантовая эффективность современных детекторов, таких как кремниевые лавинные фотодиодные детекторы (Si-APDs)44 или детектор зоны Airyscan45,облегчают использование еще более низких количеств красителей, чем раньше. Другой ключевой целью статьи является пример со-изображения свойства этих красителей с другими фторофорами, самое главное, те из GFP как наиболее часто используемый флуоресцентный белок в биологии. Это должно помочь в разработке изображений экспериментов, которые направлены на корреляцию субклеточной динамики локализации флуоресцентных белков синтеза к тем из грибковых клеточных стенок, плазменной мембраны или эндо- и экзоцитоза путь и т.д.
Изображение в естественных и свободных от стресса условиях является ключом к получению достоверных данных. Некоторые практические соображения, касающиеся культуры среды и подготовки образцов целью обеспечения отправной точки для поиска оптимальных условий, которые позволяют без артефактов, долгое время наблюдения здоровых, неподчеркнутых клеток с самым высоким S / N-коэффициент возможно для любого данного образца. Универсального способа получения надежных и значимых результатов визуализации не существует. Подходу присущи, что биологические различия в выборке, субъективность и ожидания микроскопа, а также обработка изображений оказывают значительное влияние соответственно на сбор и интерпретацию данных. Таким образом, практический опыт микроскопа, ее / его интимные знания о клеточной биологии гриба под следствием, а также умелый образец подготовки для создания условий, как "естественный" и ненарушенный, как это возможно в лабораторных условиях, имеют первостепенное значение для приобретения и оценки изображений данных, которые правдиво отражает изученных клеточных явлений. Как правило, возникновение нежелательных побочных эффектов флуоресцентных красителей, начиная от тонких и, следовательно, не очевидно видимых активации плазменной мембраны или клеточной стенки ремоделирования путей реакции на стресс к простой цитотоксичной индукции клеточного автолизиса, может быть только надежно предотвращено путем применения низкой концентрации красителей No 2 мкм.
Применение флуоресцентных красителей просто, но их специфика плохо характеризуется. Ключевой силой использования флуоресцентных красителей является подготовитечная простота экспериментальных протоколов. Культивирование и отбор проб гриба, добавление красителя (ы), и монтаж на стадии микроскопа (с практикой) просто. Корректировка основных настроек визуализации, включая возбуждение и длину волн выбросов, время экспозиции, настройки временных курсов и т.д., следовать простым биофизическим правилам микроскопа и биологическим правилам используемых флуоресцентных красителей внутри клеток. Таблица 1 предназначена для поддержки выявления наиболее подходящей комбинации красителей или красителей для экспериментов. Кроме того, флуоресцентные красители по разумной цене, легко доступны с надежным высоким качеством и, таким образом, обеспечить высоко воспроизводимое применение.
Двумя основными ограничениями использования мембранной или клеточной стенки селективных флуоресцентных красителей являются (часто) ограниченные знания их точных окрашивающих свойств, которые в большинстве случаев являются неспецифическими на органелле и молекулярном уровне, и их концентрацией зависимых нежелательных побочных эффектов. FM красители специфичны для липидных двухслойных, участвующих в эндо- и экзоцитоза. Однако, точно какие субклеточные органеллы становятся последовательно помечены в тестируемых условиях не сразу очевидно и требует сравнения различных вариантов FM красителя, и совместное маркировка с дополнительными органель-специфических маркеров. Предпочтение FM 1-43 для митохондриальных мембран, по сравнению с FM 4-64, является одним из примеров. Селективные красители клеточной стенки отображают различную специфичность для трех основных полимеров грибковой клеточной стенки. CFW считается неспецифическим пятном для глюканов и хитина, SPF считается наиболее избирательным для глюканов, а CR считается весьма избирательным для к- и хитинов. Информация о привязке специфики ПФС к грибковым клеточным полисахаридов стенки в настоящее время отсутствует. На какое соотношение, какое соотношение грибковых клеточных полимеров наиболее эффективно помечены при заданной концентрации красителя в исследуемых грибковых видов не легко ответить, и применение подробных измерений, приобретенных in vitro или in vivo в других организмах или других грибковых видов должны быть рассмотрены очень тщательно. К сожалению, эта информация является редким и сильно рассеяны в литературе35,42,46. Более поздние записи, которые будут следовать на предыдущих исследованиях33, чтобы обеспечить новое понимание точных свойств окрашивания признакам красителей специально в грибах в настоящее время не доступны.
Контроль изображений имеет важное значение для точной оценки шаблонов окрашивания и клеточных реакций. Вероятно, наиболее сложной частью, однако, является знать клеточной биологии гриба так хорошо, что зарегистрированные изменения в субклеточной локализации мембраны и клеточной стенки селективных флуоресцентных красителей, изменения в клеточной архитектуры или гифальный рисунок роста может исключительно и уверенно быть связаны с предполагаемыми последствиями экспериментального лечения. Для этого, очень важно иметь хорошие элементы управления наряду с любым новым экспериментом изображения живых клеток. К ним относятся необработанный дикий тип в качестве отрицательного контроля изображений, чтобы исключить фоновую аутофлуоресценцию и шум детектора из приобретенного изображения, и иметь морфологический компаратор при работе с мутантами. Кроме того, положительный контроль изображения, например, штамм, который выражает GFP или RFP в цитоплазме или другой известный флуоресцентный маркер белка, имеет важное значение для установки интенсивности возбуждения света до необходимого минимума и контроля жизнеспособности клетки. Как только эти элементы управления установлены, использование флуоресцентных красителей не ограничивается только задачами визуализации, но их концентрационно-зависимая динамика окрашивания, а также зависящие от концентрации побочные эффекты могут быть аналитически использованы; например, для количественного мониторинга биосинтеза клеточной стенки в режиме реального времени или для выявления мутантов конкретных фенотипов в восприимчивости анализы47.
Улучшенные будущие приложения зависят от детального функционального анализа красительных окрашивающих свойств. Основная текущая задача заключается в дальнейшем совершенствовании и автоматизации количественного анализа изображений в целях продвижения функциональной оценки субклеточной динамики мембраны и селективных флуоресцентных красителей клеточной стенки в нитевидных грибах. Для этого сначала требуются обширные количественные исследования совместной локализации этих красителей с известными маркерами полимера органеллы и клеточной стены в сочетании с штаммами мутантов, которых не хватает в конкретных транспортных путях или в которых отсутствуют конкретные структурные компоненты. Несколько маркеров эндоцитоза для сравнительного анализа с FM красителей доступны48,49, и в отношении еще плохо характеризуется связывания особенности клеточной стенки красителей в грибах, применение флуоресцентно помечены клекан-специфических антител50 может обеспечить одну возможность для решения этой проблемы.
Раскрытие информации
Авторы заявляют, что у них нет конкурирующих финансовых интересов и ничего раскрывать не приходится.
Благодарности
Благодарим Тирольский научный фонд (TWF) за предоставление гранта #256524 AL, Венскому научно-техническому фонду (WWTF) за предоставление гранта #LS13-086 СЗ, а также Фонду публикаций Университета Инсбрука за поддержку публикации открытого доступа. Авторы также благодарят кафедру зоологии Университета Инсбрука за предоставление конфокального лазерного сканирующего микроскопа Leica TCS SP5 II.
Материалы
| Name | Company | Catalog Number | Comments |
| BRAND cell counting chamber | Merck | BR718005 | Thoma format |
| Calcofluor White M2R | Merck/Sigma-Aldrich | F3543 | cell wall dye |
| CFI Plan Apo VC 60x/1.2 NA XC WI | Nikon | MRD07602 | water immersion objective |
| CFI Plan Apo VC 60x/1.2 NA XC WI | Nikon | MRD07602 | water immersion objective |
| Congo Red | Merck/Sigma-Aldrich | C6277 | cell wall dye |
| Dimethyl sulfoxide | VWR | 8,36,73,230 | organic solvent |
| Eclipse TE2000-E with C1 scanning unit | Nikon | custom configuration | inverted laser scanning confocal microscope used to acquire example data 2 and 4 |
| Eclipse TE2000-U with Bio-Rad Radiance 2100 scannig unit | Nikon | custom configuration | inverted laser scanning confocal microscope used to acquire example data 5 |
| FM 1-43 | Merck/Sigma-Aldrich | S6814 | membrane dye |
| FM 4-64 | Merck/Sigma-Aldrich | S6689 | membrane dye |
| Glass beads | Rettberg | 1340691030 | 3 mm glass beads |
| Glass cover slips | Thermo Fisher Scientific | BB02400600A113MNT0 | 24 x 60 # 1 glass cover slips |
| HCX PL APO 63x/1.3 NA Glyc | Leica | 15506353 | glycerol immersion objective |
| LSM 510 Meta | Zeiss | custom configuration | inverted laser scanning confocal microscope used to acquire example data 3 |
| M9 Minimal Medium | Merck/Sigma-Aldrich | M6030 | generic fungal growth medium |
| Micro-slide 8-well | ibidi | 80826 | ibiTreat #1.5 polymer coverslip |
| Miracloth | Merck/Millipore | 475855-1R | polyester filtration material |
| Petri dish | Sarstedt | 8,21,472 | 92 x 16 mm culture dish w/o cams |
| Phytagel | Merck/Sigma-Aldrich | P8169 | transparent gelling agent |
| Plan Apochromat 63x/1.4 NA Oil DIC | Zeiss | 440762-9904-000 | oil immersion objective |
| Pontamine Fast Scarlet 4B | Merck/Sigma-Aldrich | 212490 | cell wall dye |
| Potato Dextrose Agar (PDA) | BD Difco | 213400 | fungal growth medium for T. atroviride |
| Potato Dextrose Broth (PDB) | BD Difco | 254920 | fungal growth medium for T. atroviride |
| Reaction tube | Sarstedt | 72,706 | 1.5 mL SafeSeal tube |
| Scalpel | B.Braun | 5518016 | Cutfix sterile scalpel #23 |
| Screw cap tube | Sarstedt | 6,25,54,502 | 15 mL polypropylene tube |
| Solophenyl Flavine 7GFE 500 | CIBA | 1485385V6 | cell wall dye |
| SynaptoGreen C4 | Biotum | 70020 | membrane dye |
| SynaptoRed C2 | Biotum | 70021 | membrane dye |
| Syringe membrane filter | Thermo Fisher Scientific | 723-9945 | 0.45 µm SFCA syringe filter |
| TCS SP5 II | Leica | custom configuration | inverted laser scanning confocal microscope used to acquire example data 1 |
| Vogel's Minimal Medium (VMM) | FGSC | Fungal Genetics Stock Centre | fungal growth medium for N. crassa |
Ссылки
- Read, N. D., Fischer, S., Parton, R. M. Imaging Spitzenkörper, pH and calcium dynamics in growing fungal hyphae. Pesticide Science. 54 (2), 179-181 (1998).
- Hickey, P. C., Swift, S. R., Roca, M. G., Read, N. D. Live-cell imaging of filamentous fungi using vital fluorescent dyes and confocal microscopy. Microbial Imaging. , 63-87 (2004).
- Jelínková, A., et al. Probing plant membranes with FM dyes: tracking, dragging or blocking. The Plant Journal. 61 (5), 883-892 (2010).
- Fischer-Parton, S., et al. Confocal microscopy of FM4-64 as a tool for analysing endocytosis and vesicle trafficking in living fungal hyphae. Journal of Microscopy. 198 (3), 246-259 (2000).
- Harris, S. D. Branching of fungal hyphae: regulation, mechanisms and comparison with other branching systems. Mycologia. 100 (6), 823-832 (2008).
- Roca, M. G., Arlt, J., Jeffree, C. E., Read, N. D. Cell biology of conidial anastomosis tubes in Neurospora crassa. Eukaryotic Cell. 4 (5), 911-919 (2005).
- Becker, Y., et al. The fungal cell-wall integrity MAPK cascade is crucial for hyphal network formation and maintenance of restrictive growth of Epichloë festucae in symbiosis with Lolium perenne. Molecular Plant-Microbe Interactions. 28 (1), 69-85 (2015).
- Justa-Schuch, D., Heilig, Y., Richthammer, C., Seiler, S. Septum formation is regulated by the RHO4-specific exchange factors BUD3 and RGF3 and by the landmark protein BUD4 in Neurospora crassa. Molecular Microbiology. 76 (1), 220-235 (2010).
- Atanasova, L., et al. The Gpr1-regulated Sur7 family protein Sfp2 is required for hyphal growth and cell wall stability in the mycoparasite Trichoderma atroviride. Scientific Reports. 8 (1), 12064 (2018).
- Betz, W. J., Mao, F., Bewick, G. S. Activity-dependent fluorescent staining and destaining of living vertebrate motor nerve terminals. Journal of Neuroscience. 12 (2), 363-375 (1992).
- Wu, Y., Yeh, F. L., Mao, F., Chapman, E. R. Biophysical characterization of styryl dye-membrane interactions. Biophysical Journal. 97 (1), 101-109 (2009).
- Betz, W. J., Mao, F., B, S. C. Imaging exocytosis and endocytosis. Current Opinion in Neurobiology. 6, 365-371 (1996).
- Hoopmann, P., Rizzoli, S. O., Betz, W. J. Imaging synaptic vesicle recycling by staining and destaining vesicles with FM dyes. Cold Spring Harbor Protocols. 2012 (1), 77-83 (2012).
- Bolte, S., et al. FM-dyes as experimental probes for dissecting vesicle trafficking in living plant cells. Journal of Microscopy. 214, 159-173 (2004).
- Riquelme, M., et al. Spitzenkorper localization and intracellular traffic of green fluorescent protein-labeled CHS-3 and CHS-6 chitin synthases in living hyphae of Neurospora crassa. Eukayotic Cell. 6 (10), 1853-1864 (2007).
- Lichius, A., Yáñez-Gutiérrez, M. E., Read, N. D., Castro-Longoria, E. Comparative live-cell imaging analyses of SPA-2, BUD-6 and BNI-1 in Neurospora crassa reveal novel features of the filamentous fungal polarisome. PloS one. 7 (1), 30372 (2012).
- Peñalva, M. A. Tracing the endocytic pathway of Aspergillus nidulans with FM4-64. Fungal Genetics and Biology. 42 (12), 963-975 (2005).
- Dijksterhuis, J., Molenaar, D. Vesicle trafficking via the Spitzenkörper during hyphal tip growth in Rhizoctonia solani. Antonie van Leeuwenhoek. 103 (4), 921-931 (2013).
- Hickey, P. C., Read, N. D. Imaging living cells of Aspergillus in vitro. Medical Mycology. 47, 110-119 (2009).
- Delgado-Álvarez, D. L., Bartnicki-García, S., Seiler, S., Mouriño-Pérez, R. R. Septum development in Neurospora crassa: the septal actomyosin tangle. PLoS One. 9 (5), 96744 (2014).
- Hageage, G. J., Harrington, B. J. Use of Calcofluor White in clinical mycology. Laboratory Medicine. 15 (2), 109-112 (1984).
- Monheit, J. E., Cowan, D. F., Moore, D. G. Rapid detection of fungi in tissues using Calcofluor White and fluorescence microscopy. Archives of Pathology and Laboratory. 108 (8), 616-618 (1984).
- Herth, W., Schnepf, E. The fluorochrome Calcofluor White binds oriented to structural polysaccharide fibrils. Protoplasma. 105 (1-2), 129-133 (1980).
- Elorza, M. V., Rico, H., Sentandreu, R. Calcofluor White alters the assembly of chitin fibrils in Saccharomyces cerevisiae and Candida albicans cells. Journal of General Microbiology. 129 (5), 1577-1582 (1983).
- Lagorce, A., et al. Genome-wide analysis of the response to cell wall mutations in the yeast Saccharomyces cerevisiae. Journal of Biological Chemistry. 278 (22), 20345-20357 (2003).
- Sanz, A. B., García, R., Rodríguez-Peña, J. M., Arroyo, J. The CWI Pathway: regulation of the transcriptional adaptive response to cell wall stress in yeast. Journal of Fungi. 4 (1), (2017).
- Slifkin, M., Cumbie, R. Congo Red as a fluorochrome for the rapid detection of fungi. Journal of Clinical Microbiology. 26 (5), 827-830 (1988).
- Michels, J., Büntzow, M. Assessment of Congo Red as a fluorescence marker for the exoskeleton of small crustaceans and the cuticle of polychaetes. Journal of Microscopy. 238 (2), 95-101 (2010).
- Pancaldi, S., Poli, F., Dall'Olio, G., Vannini, G. L. Morphological anomalies induced by Congo Red in Aspergillus niger. Archives of Microbiology. 137 (3), 185-187 (1984).
- Roncero, C., Durán, A. Effect of Calcofluor White and Congo Red on fungal cell wall morphogenesis: in vivo activation of chitin polymerization. Journal of Bacteriology. 163 (3), 1180-1185 (1985).
- Kopeck, M., Gabriel, M. The influence of Congo Red on the cell wall and (1,3)- β-d-glucan microfibril biogenesis in Saccharomyces cerevisiae. Archives of Microbiology. 158 (2), 115-126 (1992).
- Heilmann, C. J., et al. Surface stress induces a conserved cell wall stress response in the pathogenic fungus Candida albicans. Eukayotic Cell. 12 (2), 254-264 (2013).
- Hoch, H. C., Galvani, C. D., Szarowski, D. H., Turner, J. N. Two new fluorescent dyes applicable for visualization of fungal cell walls. Mycologia. 97 (3), 580-588 (2005).
- Liesche, J., Ziomkiewicz, I., Schulz, A. Super-resolution imaging with Pontamine Fast Scarlet 4BS enables direct visualization of cellulose orientation and cell connection architecture in onion epidermis cells. BMC Plant Biology. 13, 226 (2013).
- Ursache, R., Andersen, T. G., Marhavý, P., Geldner, N. A protocol for combining fluorescent proteins with histological stains for diverse cell wall components. The Plant Journal. 93 (2), 399-412 (2018).
- Knight, N. L., Sutherland, M. W. A rapid differential staining technique for Fusarium pseudograminearum in cereal tissues during crown rot infections. Plant Pathology. 60 (6), 1140-1143 (2011).
- Fajardo-Somera, R. A., et al. Dissecting the function of the different chitin synthases in vegetative growth and sexual development in Neurospora crassa. Fungal Genetics and Biology. 75, 30-45 (2015).
- Lichius, A. . Cell Fusion in Neurospora crassa. , (2010).
- Chen, W., Li, W., Dong, X., Pei, J. A Review of Biological Image Analysis. Current Bioinformatisc. 13 (4), 337-343 (2018).
- Goldman, R. D., Swedlow, J., Spector, D. L. . Live cell imaging: A laboratory manual. , (2010).
- Eliceiri, K. W., et al. Biological imaging software tools. Nature methods. 9 (7), 697-710 (2012).
- Zemanek, G., Jagusiak, A., Chłopaś, K., Piekarska, B., Stopa, B. Congo Red fluorescence upon binding to macromolecules - a possible explanation for the enhanced intensity. Bio-Algorithms and Med-Systems. 13 (2), 1187 (2017).
- Hickey, P. C., Jacobson, D. J., Read, N. D., Louise Glass, N. Live-cell imaging of vegetative hyphal fusion in Neurospora crassa. Fungal Genetics and Biology. 37 (1), 109-119 (2002).
- . Hamamatsu Si APD - high sensitivity photodiodes having an internal gain mechanism: Avalanche photodiode selection guide 2019 Available from: https://www.hamamatsu.com/resources/pdf/ssd/si_apd_kapd0001e.pdf (2019)
- Huff, J. The Airyscan detector from ZEISS: confocal imaging with improved signal-to-noise ratio and super-resolution. Nature Methods. 12, 1205 (2015).
- Thomas, J., Idris, N. A., Collings, D. A. Pontamine Fast Scarlet 4B bifluorescence and measurements of cellulose microfibril angles. Journal of Microscopy. 268 (1), 13-27 (2017).
- Ram, A. F. J., Klis, F. M. Identification of fungal cell wall mutants using susceptibility assays based on Calcofluor White and Congo Red. Nature Protocols. 1 (5), 2253-2256 (2006).
- Toshima, J. Y., et al. Spatial dynamics of receptor-mediated endocytic trafficking in budding yeast revealed by using fluorescent alpha-factor derivatives. Proceedings of the National Academy of Science of the USA. 103 (15), 5793-5798 (2006).
- Kilaru, S., Schuster, M., Latz, M., Guo, M., Steinberg, G. Fluorescent markers of the endocytic pathway in Zymoseptoria tritici. Fungal Genetics and Biology. 79, 150-157 (2015).
- Fu, C., Tanaka, A., Free, S. J. Neurospora crassa 1,3-α-glucan synthase, AGS-1, is required for cell wall biosynthesis during macroconidia development. Microbiology. 160, 1618-1627 (2014).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены