
Method Article
Application of CRISPR Interference (CRISPRi) for Gene Silencing in Pathogenic Species of Leptospira
In This Article
Summary
Here, the application of CRISPR interference (CRISPRi) for specific gene silencing in Leptospira species is described. Leptospira cells are transformed by conjugation with plasmids expressing dCas9 (catalytically "dead" Cas9) and a single-guide RNA (sgRNA), responsible for base pairing to the desired genomic target. Methods to validate gene silencing are presented.
Abstract
Leptospirosis is a global neglected zoonosis, responsible for at least 1 million cases per year and almost 60 thousand deaths. The disease is caused by pathogenic and virulent bacteria of the genus Leptospira, either by direct contact with the bacteria or indirectly by exposure to contaminated water or soil. Domestic and wild animals act as reservoir hosts of infection, shedding leptospires from colonized renal tubules of the kidney, via urine, into the environment. The generation of mutant strains of Leptospira is critical to evaluate and understand pathogenic mechanisms of infection. CRISPR interference (CRISPRi) has proven to be a straightforward, affordable, and specific tool for gene silencing in pathogenic Leptospira. Therefore, the methodological details of obtaining the plasmid constructs containing both dCas9 and guide RNA, delivery of plasmids to Leptospira by conjugation with the E. coli strain β2163, and transconjugant recovery and evaluation, will be described. In addition, the recently described Hornsby-Alt-Nally (HAN) media allows for the relatively rapid isolation and selection of mutant colonies on agar plates.
Introduction
Leptospirosis is a neglected worldwide zoonosis caused by pathogenic and virulent species of the genus Leptospira. In humans, the disease accounts for more than one million cases and 60,000 deaths per year worldwide1,2. So far, there is no long-term and effective vaccine for the disease. The identification of virulence factors and pathogenic mechanisms is pivotal to the development of better therapeutic and prophylactic strategies. Therefore, the ability to generate genetic mutations and assess the resulting phenotype is critical to functional genomic analysis3.
The construction of mutants in pathogenic Leptospira was considered, until now, inherently inefficient, laborious, costly, and difficult to implement. This scenario drastically changed with the application of the recent CRISPR interference (CRISPRi) to saprophytic4 and pathogenic5 leptospires.
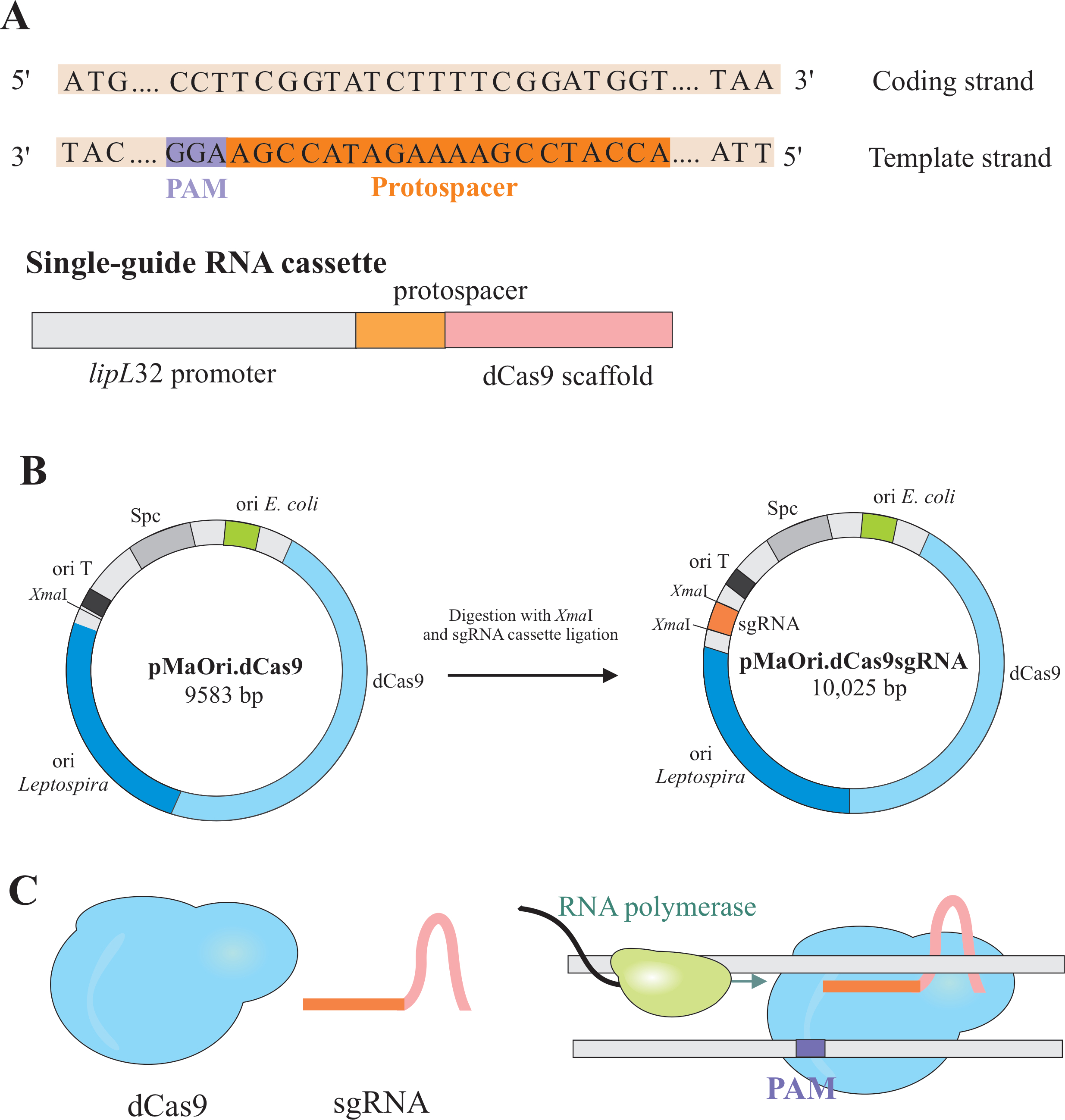
Gene silencing is achieved by the expression of two components: a variant of the CRISPR/Cas (clustered regularly interspaced short palindromic repeat/CRISPR associated) enzyme Cas9 from Streptococcus pyogenes, called catalytically dead Cas9 (dCas9) and a single-guide RNA (sgRNA), that can be edited according to the desired target6,7,8. dCas9 protein, when bound to the sgRNA, is directed to specific DNA targets by Watson and Crick base pairing, causing a steric blockage to RNA polymerase elongation, resulting in gene silencing due to the obstructed gene transcription7 (Figure 1).
This manuscript aims to describe the construction of the plasmid for expressing both dCas9 and sgRNA, conjugation between donor E. coli β2163 and recipient Leptospira cells, transconjugant recovery, and finally, validation of selected mutant colonies.
Protocol
1. Protospacer definition and plasmid construction
NOTE: In this section, the first step of selecting appropriate protospacers for constructing the sgRNA and further ligation into pMaOri.dCas9, is described (Figure 1). This protospacer sequence comprises of a 20 nucleotides sequence against the desired target.
- Obtain the nucleotide sequence of the gene of interest for silencing at GenBank (https://www.ncbi.nlm.nih.gov/genbank). Submit it to the CHOPCHOP webserver (http://chopchop.cbu.uib.no/), with parameters defined for Streptococcus pyogenes Cas9 and protospacer adjacent motif NGG after selecting the "Fasta Target". Define the parameters to "CRISPR/Cas9" and PAM (protospacer adjacent motif) NGG.
- Based on the results obtained, select protospacers with the best score possible (green arrow), that are located as close as possible to the 5´end of the coding region and, most importantly, are contained in the template (minus) strand since the sgRNA must pair to the coding strand of the gene for complete gene silencing.
NOTE: The NGG motif is not included in the final sgRNA sequence. - Use the lipL32 promoter to express the single guide RNA that contains a variable 20 nucleotide sequence at the 5' end and a conserved dCas9 scaffold sequence. Merge the 20-nt sequence, termed protospacer, to the lipL32 promoter (at its 5´ end) and sgRNA scaffold (3'end) (Figure 1B).
NOTE: For a well-defined lipL32 promoter, utilize the promoter region comprising -334 to the TSS (Transcription Start Site, based on Zhukova et al.9). Check the Supplementary File for the final sgRNA cassette. - Generate the sgRNA cassette by sequential PCR5 or have it synthesized by a commercial provider.
- After obtaining the cassette, ligate it into pMaOri.dCas9 plasmid at the XmaI restriction site at both ends (cccggg)4.
- Digest both the sgRNA cassette and pMaOri.dCas9 plasmid with XmaI restriction enzyme and proceed to ligation (Figure 1B).
- Perform the cloning steps in the dT auxotrophic E. coli strain π110, due to the pMaOri11 (and by extension, pMaOri.dCas9) origin of replication, R6K-gamma.
NOTE: For a detailed protocol for ligation and clone selection, refer to previous publications by Fernandes and Nascimento12. sgRNA-guided dCas9 will bind to the coding strand of the selected gene of interest and, therefore, will obstruct RNA polymerase elongation (Figure 1C), resulting in gene silencing.
2. Leptospira transformation by conjugation
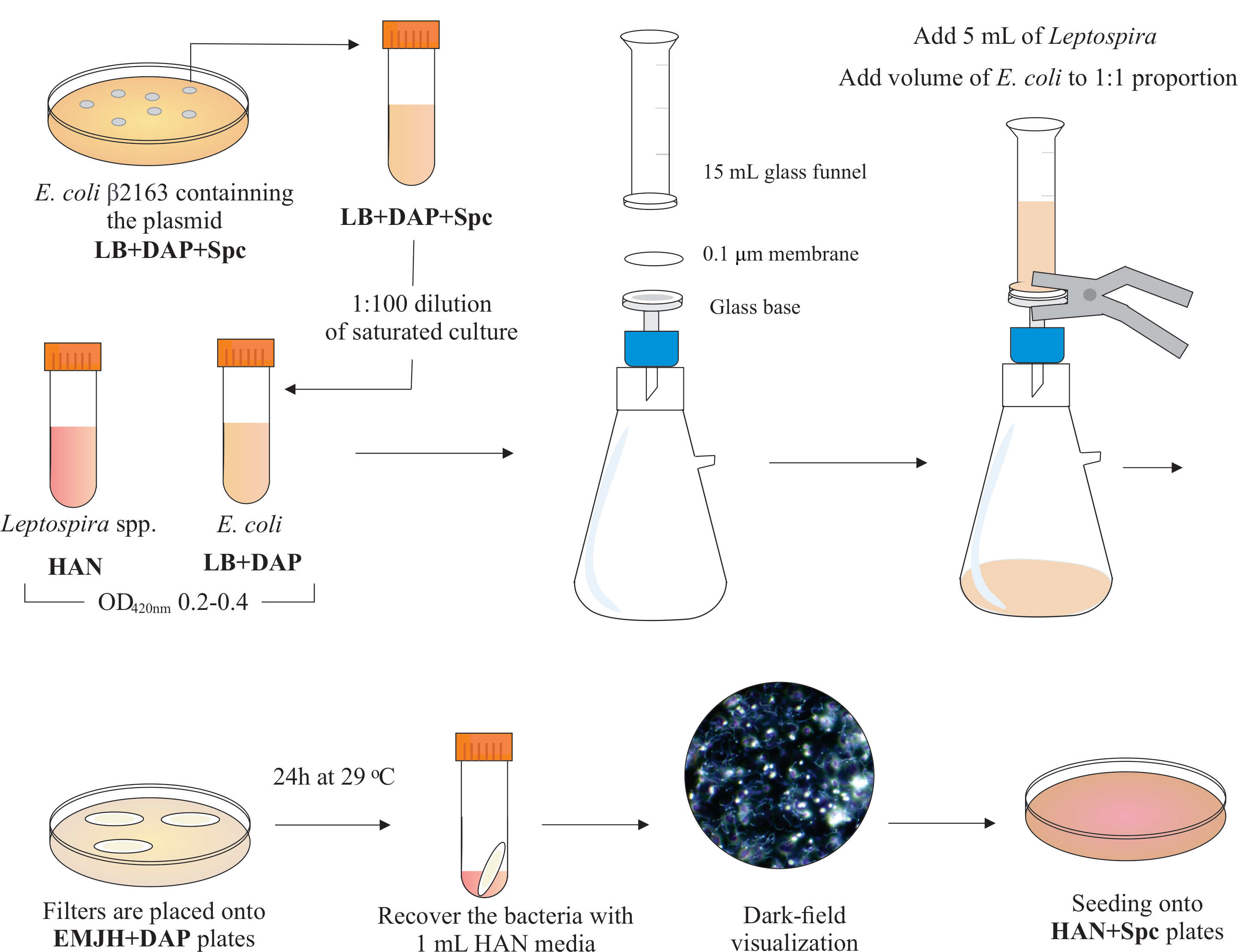
NOTE: A graphical scheme of this step is presented in Figure 2. To make HAN media and HAN plates, refer to Hornsby et al.13 and Fernandes et al.5.
- Grow pathogenic Leptospira cells at 29 or 37 °C in HAN media13 under agitation by diluting a saturated culture in fresh HAN at 1:100; typically, L. interrogans serovar Copenhageni strain Fiocruz L1-130 takes 4-6 days to reach the appropriate cell density.
- Ensure cultures reach an O.D. of 0.2-0.4 at 420 nm (2 to 5 x 108 cells/mL) before using for conjugation.
NOTE: Since HAN media changes color as cell densities increase (due to phenol red contained in DMEM media), centrifuge (4,000 x g, 15 min, room temperature) 1 mL of the culture media to remove leptospires and apply supernatant as a blank for measuring the O.D.
- Ensure cultures reach an O.D. of 0.2-0.4 at 420 nm (2 to 5 x 108 cells/mL) before using for conjugation.
- Transform the conjugative E. coli strain β2163, auxotrophic for Diaminopimelic acid (DAP), with the plasmid pMaOri.dCas9 containing the sgRNA cassette. For E. coli transformation, use either heat-shock protocols or electroporation. Include transformation with the plasmid pMaOri.dCas9 with no sgRNA cassette as a control.
- For heat-shock transformation, mix the plasmid DNA (100 ng) with chemically competent E. coli cells and incubate on ice for 30 min. Perform heat shock at 42 °C for 90 s and place it again on ice for 5 min. Recover the cells by adding 1 mL of LB media, incubate at 37 °C for 1 h and proceed to the plating.
- For electroporation, use electrocompetent cells mixed with 100 ng of plasmid DNA. Use the following parameters for pulse: 1.8 kV, 100 Ω, and 25 µF. Recover the cells as explained above.
- Plate the transformed donor E. coli cells in LB agar medium supplemented with Diaminopimelic acid (DAP) (0.3 mM) and spectinomycin (40 μg/mL) to select for plasmids.
- For conjugation, select one colony from each plate one day before the day of conjugation (which is determined by monitoring the O.D. of cultures of leptospires).
- Select one colony of E. coli β2163 from the empty pMaOri.dCas9, and one from pMaOri.dCas9sgRNA plates. Allow them to grow overnight in 10 mL of LB plus DAP and spectinomycin at 37 °C.
- The next day, dilute the saturated cultures 1:100 in 10 mL of fresh LB plus DAP (do not include the antibiotic here) until OD420nm of 0.2-0.4. Normally, it takes 2-3 h for E. coli to reach these densities.
- Inside a BSL2 biosafety hood, assemble a filtration apparatus by placing a 25 mm diameter, 0.1 µm pore size, mixed cellulose esters membrane filter on the top of the glass base. Place a 15 mL glass funnel on the top and hold both pieces with spring clamps. Connect the glass to a vacuum pump and add the cultures to the funnel for filtration.
- Add 5 mL of Leptospira culture to the funnel. Add a volume of E. coli to constitute the 1:1 proportion based on the OD420nm values of both cultures. Turn on the vacuum pump and concentrate cells by filtration. After cell concentration in the membrane filter, carefully retrieve it. Ensure medium is filtered through the membrane.
NOTE: Filtration takes 5 to 10 min. - Place the filter on a commercially available EMJH plate (see Table of Materials) supplemented with DAP (0.3 mM). Ensure that bacteria side is up. Incubate the plates at 29 °C for 24 h.
NOTE: If HAN or supplemented in-house EMJH14 plates are used, E. coli can proliferate and overcome the intended 1:1 proportion, which in turn can decrease conjugation efficiency5. - After 24 h, recover the filters from the plates and place each individual filter in a 50 mL conical tube.
- Use 1 mL of liquid HAN medium to release the cells from the filter surface by extensive pipetting and vortexing.
- Visualize the recovered mixed bacterial solutions by dark field microscopy to check for cell viability and motility, and Leptospira:E. coli proportions.
NOTE: At this stage, equivalent numbers of E. coli and Leptospira can be seen. - Spread 100-200 μL of this culture onto HAN plates containing 0.4% inactivated rabbit serum and 40 μg/mL spectinomycin. Incubate plates at 37 °C in a 3% CO2 atmosphere.
NOTE: Normally, L. interrogans serovar Copenhageni strain Fiocruz L1-130 cells form colonies in 5-7 days on control plates and in 8-10 days on spectinomycin plates. At this stage, E. coli will not grow since they are auxotrophic for DAP. - As a control, dilute cultures at 104 leptospires/mL and add 100 μL onto plates without antibiotic, for monitoring leptospiral growth.
3. Colony selection and transconjugant growth and validation
NOTE: Colonies should be apparent by day 10. However, they are not too easy to visualize. Normally at this time point, HAN plates are a bit opaque because of the dried cells that were spread and Leptospira colonies can appear as a transparent halo against the whitish background. It is recommended to view the plates at different angles to achieve different light incidence, therefore, making the colonies more apparent. At longer incubation times, colonies can acquire a denser appearance, and in this case, they present as milky halos against a dark background.
- Add 100 μL of liquid HAN media to each 1.5 mL microtube to recover mutants. Take at least 3 colonies from each plate.
- With the aid of a micropipette tip, "dig" the agar to retrieve the colonies from the plates since leptospiral colonies can be subsurface.
NOTE: Agar is expected to be taken along at this stage. Colonies should be taken from control plates containing empty pMaOri.dCas9 plasmid, and plates with leptospires containing plasmids expressing both dCas9 and single guide RNA, designed for the target gene. - Dispense the collected colony in 100 μL of HAN media in a 1.5 mL microtube and vigorously homogenize. At this stage, ensure maximal break of the agar integrity to release cells. Vortex the suspension for 10 s.
- With the aid of a micropipette tip, "dig" the agar to retrieve the colonies from the plates since leptospiral colonies can be subsurface.
- Visualize the recovered cells by dark field microscopy at a 200-400x magnification by adding a 5 µL drop onto a glass slide and cover the samples immediately with a coverslip.
- Confirm the presence of live and viable leptospires recovered from the colonies.
- After visualization and confirmation of viable leptospires, transfer 100 μL of cells to liquid HAN media containing 40 μg/mL spectinomycin.
- After growth in liquid HAN media, evaluate the cultures for the presence of the plasmid with primer pMaOri2 F (ACGCAATGTATCGATACCGAC) and R (ATAGGTGAAGTAGGCCCACCC), which recognize the region that flanks the sgRNA cassette.
- Collect 200 μL of culture, centrifuge (4,000 x g, 15 min), discard the supernatant, and resuspend the resulting pellet in 20 μL of water.
- Use this suspension as a template for additional PCR, without the need for DNA extraction12.
NOTE: Cells with pMaOri.dCas9 will render an amplicon of 281 bp, compared to those containing the plasmids with the sgRNA cassette which will render an amplicon of 723 bp.
- For confirmation of gene silencing, perform an immunoblot utilizing cell extracts from transconjugants containing only pMaOri.dCas9 (negative control) and pMaOri.dCas9sgRNA.
- Apply the equivalent of 5 x 107 cells per lane of the sodium dodecyl sulfate (SDS) polyacrylamide gel.
- Electrotransfer proteins to a membrane for incubation with appropriate antibodies. Besides the antibody against the target gene for silencing, use another one for loading control.
- Keep the mutant cultures in HAN media plus spectinomycin for maintaining the plasmid. If no antibiotics are applied to the media, complete gene silencing can be observed for at least three passages5.
Results
Even though the CG content in Leptospira spp. genomes is typically around 35%; virtually every gene is likely to contain the PAM 5'NGG 3'; this motif needs to be considered in the template strand. After inputting the coding sequence of a gene (from start to stop codons), based on the CHOPCHOP results, protospacers must be selected at the minus (-, template) strand. It is important to not include the NGG motif in the 20-nt sgRNA protospacer.
If conjugation is performed with a 1:1 donor:recipient cell proportion, for 24 h on the surface of EMJH agar plates plus DAP, and 200 μL of the recovered bacterial suspension are spread onto HAN plus spectinomycin agar plates, transconjugants colonies should be visible at approximately 8-10 days. Spreading of this volume normally results in 20-40 colonies per plate (Figure 3A). In order to check for cell viability after conjugation, cells can be spread onto HAN plates with no antibiotic selection. In this case, colonies can be observed as soon as 7 days. HAN plates turn pale yellow in a 3% CO2 atmosphere.
After colony picking and growth in liquid media plus spectinomycin, PCR using whole cells and pMaOri2 primers can be used for an initial quality check of the transconjugants (Figure 3B). Leptospiral cells containing the control pMaOri.dCas9 plasmid should result in an amplicon of 281 bp, while those cells containing the plasmid for silencing, that is, containing both dCas9 and sgRNA, should result in a 723 bp amplicon. pMaOri2 F and R primers were designed to flank the XmaI restriction site, which is the site used during sgRNA cassette ligation.
With the confirmation of plasmid presence, cells can be harvested from the media, washed twice with PBS, and then used to prepare a whole-cell extract for immunoblotting. If the silencing occurred, the target proteins, in this case, either LipL32 or both LigA and LigB, should be observed only in the wild type cells and in those containing pMaOri.dCas9; even with higher exposure times, no corresponding proteins should be visible in the cells containing pMaOri.dCas9sgRNA (Figure 3C).
If experiments to assess leptospiral virulence after gene silencing are planned, cultures used for conjugation should be low passage virulent Leptospira. After gene silencing is confirmed, several aliquots can be frozen as a backup. If the silenced gene has a measurable phenotype, e.g., based on previous work with recombinant proteins, cultures can be used for validation and, in this case, cells containing pMaOri.dCas9 only, can be included as a negative control.

Figure 1: Development of dCas9 and sgRNA expressing plasmid. (A) A 20-nt long protospacer, followed by the S. pyogenes dCas9 PAM 5'-NGG-3', is selected within the template strand of the target gene so the subsequent sgRNA can perform Watson and Crick base pairing to the corresponding coding strand, resulting in complete gene silencing. (B) The sgRNA cassette is composed of the lipL32 promoter, 20-nt protospacer, and dCas9 scaffold. The pMaOri.dCas9 plasmid is used as a backbone for sgRNA cassette ligation at the XmaI restriction site. The resulting plasmid, termed pMaOri.dCas9sgRNA is delivered to leptospires, and the expression of both dCas9 and sgRNA is responsible for the gene silencing. (C) sgRNA-directed dCas9 acts as a physical barrier to RNA polymerase elongation, therefore, hampering transcription. Please click here to view a larger version of this figure.
{kind=link}

Figure 2: Schematic representation of conjugation protocol. The desired Leptospira species is grown in HAN media, under agitation, until O.D. of 0.2-0.4 (mid-log phase) at 420 nm. One day before conjugation, a colony of recombinant donor E. coli β2163 containing the plasmid of interest is picked from LB+DAP+Spc agar plates, as cells are grown overnight in liquid LB with the same supplementation. The next day, saturated E. coli cultures are diluted in LB plus DAP and grown until O.D. of 0.2-0.4 at 420 nm. Both donor E. coli and recipient Leptospira are mixed at 1:1 cell proportion onto the surface of a 0.1 μm filter by a filtration apparatus under negative pressure. Then, filters are placed on top of the EMJH agar plates supplemented with DAP, and incubation proceeds for 24 h at 29 °C. The use of EMJH limits E. coli proliferation, and the intended 1:1 proportion is maintained. Bacteria are recovered from filters by pipetting with 1 mL HAN media, and suspensions are visualized under darkfield microscopy. Finally, 100-200 μL of each suspension are seeded onto HAN agar plates containing 0.4% rabbit serum and incubated at 37 °C in 3% CO2. At this stage, DAP is omitted, and as a result, auxotrophic E. coli will not grow. Please click here to view a larger version of this figure.
{kind=link}

Figure 3: Representative results for mutants' evaluation. (A) Colonies from plates containing Leptospira transformed with empty pMaOri.dCas9 (negative control for further experiments) and plasmids pMaOri.dCas9sgRNA (with targeted gene silenced) are picked, vigorously homogenized in liquid HAN and grown in liquid HAN containing spectinomycin. Recombinant cells can be validated by PCR with primers flanking the XmaI site within pMaOri.dCas9. (B) In this case, cells containing pMaOri.dCas9 only resulted in an amplicon of 281 bp, while those cells containing the plasmid for silencing, containing both dCas9 and sgRNA, showed a 723 bp amplicon. After confirmation of the presence of the plasmids, gene silencing was validated by immunoblot analysis. (C) Incubation with antibodies to both target protein and a loading control protein is recommended; in the representative immunoblot, whole-cell extracts from transconjugants containing pMaOri.dCas9 alone or with sgRNA cassettes targeting lipL32 (pMaOri.dCas9sgRNAlipL32) and both LigA and LigB (pMaOri.dCas9sgRNAligAB) genes are displayed. Co-incubation with anti-LipL32, anti-LigAB and anti-LipL41 (non-target, loading control) confirms that the expression of LipL32 protein is abolished in cells containing pMaOri.dCas9sgRNAlipL32 and both LigA and LigB in cells containing pMaOri.dCas9sgRNAligAB. Please click here to view a larger version of this figure.
{kind=link}
Supplementary File: Single guide RNA (sgRNA) cassette sequence. The sgRNA transcription is directed by the constitutive lipL32 promoter (bold nucleotides). sgRNA is composed of 20 nucleotides referring to the protospacer, responsible for base pairing to the coding strand of the target gene, and dCas9 scaffold sequence (underlined nucleotides). XmaI restriction sites (cccggg) are included at both ends for ligation at pMaOri.dCas9 plasmid. Please click here to download this File.
Discussion
After the early sequencing of pathogenic15,16,17,18 and saprophytic19 Leptospira species, data mining of the genome shed light on several aspects of leptospiral pathogenesis. In most cases, protein function was explored by using the recombinant counterpart of putative leptospiral surface-exposed proteins and subsequent speculation of the native protein function20,21,22,23,24,25,26.
The generation of mutants, and evaluation of their respective phenotype, are key components of functional genomic analysis. Initial attempts to generate mutants in Leptospira spp. were achieved by random transposon mutagenesis27,28,29,30; however, after extensive and laborious analysis for inferring the identity of disrupted genes, it was noted that only 15% of all genes in L. interrogans serovar Manilae were disrupted27. Targeted gene knockout was further achieved by homologous recombination utilizing suicide plasmids to deliver an antibiotic resistance cassette flanked by homologous arms within the desired target31,32.
By applying these technologies, several aspects of leptospiral basic biology and virulence were explored31,33,34,35,36,37. The development of the E. coli-Leptospira conjugative shuttle vector, pMaOri 11, allowed the delivery of components for episomal gene silencing.
It was previously shown that the Cas9-induced double-strand break is lethal to Leptospira spp. and, as an alternative, the catalytically inactive variant of the enzyme, dCas9, can be used to achieve gene silencing in both saprophytic and pathogenic species4, 5. By using the plasmid pMaOri.dCas9 as a backbone for sgRNA cassette ligation, specific and stable gene silencing can be obtained due to the expression of both dCas9 and sgRNA; dCas9-bound sgRNA will lead the protein to the desired target by Watson-Crick base pairing.
For complete gene silencing, the protospacer should be designed based on the template strand of the desired gene so that base pairing of the sgRNA occurs with the coding strand. Based on an average C+G content of 35% in Leptospira spp., the PAM 5'-NGG-3' will occur at least 3 times every 100 bp. Therefore, virtually any gene within the genome of Leptospira will contain at least one PAM. However, if the motif NGG is not found, the alternative NAG motif can be evaluated.
Previous gene silencing techniques, such as zinc fingers and TALE (transcription activator-like effectors), relied on the construction of one different protein to each target, making these techniques laborious and costly38. In the case of CRISPRi, the variable component is the sgRNA, making it necessary to only change the 20 bp at the 5' end. Complete, stable, and targeted gene silencing has been observed not only in Leptospira spp.4,5, but also in other bacteria8,39,40,41.
The development of HAN media13 favored the recovery of mutants by drastically reducing the incubation time for colony formation and allowing Leptospira to grow at 37 oC. However, during the conjugation step, its use is not recommended since E. coli can vigorously proliferate in this media and overcome the intended 1:1 proportion between donor and recipient cells. At this stage, EMJH plus DAP is the better choice, since E. coli replicate poorly in this media. It is worth mentioning that some laboratories make in-house supplemented EMJH, which can contain additional components that might also support the growth of E. coli cells.
The conjugation protocol presented here was optimized for L. interrogans serovar Copenhageni strain Fiocruz L1-130, and it was also proven to be effective in the transformation of a recently isolated pathogenic strain from soil samples5. Initial attempts with different serovars of L. borgpetersenii species indicate lower conjugation efficiencies with the described protocol. Thus, when working with different species/serovars of Leptospira, optimal conditions for conjugation should be determined empirically, considering donor:recipient cell proportions, initial cell densities, conjugation media and time (24 and 48 h). It is reasonable to assume that different Leptospira species and serovars will behave differently with different conjugation protocols.
Even though saprophytic Leptospira colonies are relatively easy to visualize on plates, pathogenic colonies can be more difficult to observe. Normally, by using HAN media supplemented with 0.4% rabbit serum and spectinomycin, transconjugant colonies can be observed at day 10. In our experience, colonies initially present as a transparent halo at the media surface. In the video protocol, denser colonies, after 14 days of growth, are shown since the transparent ones were difficult to film. At this stage, rotating the plate to achieve different light incidence and shifting between white and dark backgrounds can help identify colonies.
For mutant validation, immunoblotting offers a straightforward approach; however, since antibodies are not always available against target proteins, alternative strategies to validate gene silencing can be pursued. Quantitative reverse-transcriptase PCR (qRT-PCR) using primers for the target gene and a constitutive control is effective to validate gene silencing since sgRNA-guided dCas9 is responsible for blockage of gene transcription. If the target gene encodes a clearly defined protein band in protein gels, SDS-PAGE can demonstrate silencing, and as per the lipL32 gene silencing5. If LPS biosynthesis genes are silenced, LPS staining can be employed; in the case of silencing genes encoding for enzymes with well-defined substrates, kinetic assays with chromogenic substrates are valid strategies; β-galactosidase silencing in L. biflexa was validated by the use of X-gal and ONPG (ortho-Nitrophenyl-β-galactoside) substrates4.
After confirmation of gene silencing, experiments can be designed to further evaluate phenotype. Binding assays can be performed in the case of silencing bacterial adhesins; serum-challenge assays confirmed the role of LigA and LigB in serum survival displayed by pathogenic Leptospira5. Mutants can also be used to inoculate animals to assess attenuation of virulence; in this case, animals inoculated with the mutant should be compared to those infected with cells containing pMaOri.dCas9 only.
In conclusion, the current protocol describes the application of CRISPRi for gene silencing in pathogenic Leptospira species using HAN media to facilitate mutant recovery within 10 days. Gene silencing combined with functional genomic analysis will improve our understanding of pathogenic mechanisms of Leptospira, and ultimately leading to the development of better prophylactic strategies for disease control.
Disclosures
The authors have nothing to disclose.
Acknowledgements
USDA is an equal opportunity provider and employer. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information, and does not imply recommendation or endorsement by the U.S. Department of Agriculture. The Brazilian agency FAPESP (grant 2014/50981-0) financially supported this work; LGVF is funded with a fellowship from FAPESP (2017/06731-8 and 2019/20302-8). The funders had no role in study design, data collection and analysis, decision to publish, or manuscript preparation. The authors also thank Hannah Hill and Alexander Grimes from the USDA Visual Services for filming and editing the video protocol.
Materials
| Name | Company | Catalog Number | Comments |
| 0.1 µm pore size mixed cellulose esters membrane | Millipore | VCWP02500 | Filtration for bacterial conjugation |
| 2,6-Diaminopimelic acid (DAP) | Sigma | D1377 | Growth of auxotrophic E. coli β2163 |
| Agar Noble | BD & Company | 214230 | Used for preparation of solid EMJH and HAN plates |
| Bacto Agar | BD & Company | 214010 | Used for preparation of solid LB plates |
| Clarity Western ECL substrate | Biorad | 170-5060 | Chemiluminescent substrate |
| dNTP set | Thermo Fisher | 10297-018 | dNTPs for PCR reaction |
| Glass Microanalysis Filter Holder | Millipore | XX1012530 | Filtration for bacterial conjugation |
| Imaging System | Biorad | ChemiDoc MP | Chemiluminescence detection |
| LB broth, Miller | BD & Company | 244620 | Lysogenic liquid medium for E. coli culturing |
| Leptospira Enrichment EMJH | BD & Company | 279510 | Supplementation of EMJH media |
| Leptospira Medium Base EMJH | BD & Company | 279410 | EMJH medium for Leptospira |
| Mini-PROTEAN TGX Gels 12% | Biorad | 4568043 | Used for polyacrylamide gel eletrophoresis |
| Optical density reader | Molecular Devices | SpectraMax M2 | For optical density measurements of bacterial cultures |
| Phosphate Buffered Saline 7.4 | Sigma | 806552 | Saline solution for washing bacterial pellets |
| Spectinomycin | Sigma | S0692 | Selection of pMaOri backbone plasmids |
| Taq DNA Polymerase | Thermo Fisher | EP0402 | Enyme, buffer and MgCl2 for PCR reaction |
| Thermocycler | Applied Biosystem | GeneAmp PCR System 9700 | Used for PCR reaction cycling |
| Thymidine (dT) | Sigma | T9250 | Growth of auxotrophic E. coli π1 |
| XmaI restriction enzyme | New Englan BioLabs | R0180L | Digestion of plasmids and inserts |
References
- Bharti, A. R., et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infectious Disease. 3 (12), 757-771 (2003).
- Costa, F., et al. Global morbidity and mortality of Leptospirosis: A systematic review. PLoS Neglected Tropical Disease. 9 (9), 0003898(2015).
- Shapiro, R. S., et al. A CRISPR-Cas9-based gene drive platform for genetic interaction analysis in Candida albicans. Nature Microbiology. 3 (1), 73-82 (2018).
- Fernandes, L. G. V., et al. Gene silencing based on RNA-guided catalytically inactive Cas9 (dCas9): a new tool for genetic engineering in Leptospira. Science Reports. 9 (1), 1839(2019).
- Fernandes, L. G. V., Hornsby, R. L., Nascimento, A. L. T. O., Nally, J. E. Genetic manipulation of pathogenic Leptospira: CRISPR interference (CRISPRi)-mediated gene silencing and rapid mutant recovery at 37 C. Science Reports. 11 (1), 1768(2021).
- Jinek, M., et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 337 (6096), 816-821 (2012).
- Qi, L. S., et al. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell. 152 (5), 1173-1183 (2013).
- Choudhary, E., Thakur, P., Pareek, M., Agarwal, N. Gene silencing by CRISPR interference in mycobacteria. Nature Communication. 6, 6267(2015).
- Zhukova, A., et al. Genome-wide transcriptional start site mapping and sRNA identification in the pathogen. Frontiers in Cell and Infectious Microbiology. 7, 10(2017).
- Demarre, G., et al. A new family of mobilizable suicide plasmids based on broad host range R388 plasmid (IncW) and RP4 plasmid (IncPalpha) conjugative machineries, and their cognate Escherichia coli host strains. Research in Microbiology. 156 (2), 245-255 (2005).
- Pappas, C. J., Benaroudj, N., Picardeau, M. A replicative plasmid vector allows efficient complementation of pathogenic Leptospira strains. Applied Environmental Microbiology. 81 (9), 3176-3181 (2015).
- Fernandes, L. G. V., Nascimento, A. L. T. O. Specific gene silencing in Leptospira biflexa by RNA-guided catalytically inactive Cas9 (dCas9). Methods in Molecular Biology. 2134, 109-122 (2020).
- Hornsby, R. L., Alt, D. P., Nally, J. E. Isolation and propagation of leptospires at 37 °C directly from the mammalian host. Science Reports. 10 (1), 9620(2020).
- Haake, D. A. Hamster model of leptospirosis. Current Protocols in Microbiology. , Chapter 12, Unit 12E.12 (2006).
- Nascimento, A. L., et al. Comparative genomics of two Leptospira interrogans serovars reveals novel insights into physiology and pathogenesis. Journal of Bacteriology. 186 (7), 2164-2172 (2004).
- Nascimento, A. L., et al. Genome features of Leptospira interrogans serovar Copenhageni. Brazillian Journal of Medical Biology Research. 37 (4), 459-477 (2004).
- Ren, S. X., et al. Unique physiological and pathogenic features of Leptospira interrogans revealed by whole-genome sequencing. Nature. 422 (6934), 888-893 (2003).
- Bulach, D. M., et al. Genome reduction in Leptospira borgpetersenii reflects limited transmission potential. Proceedings of the National Academy of Science U. S. A. 103 (39), 14560-14565 (2006).
- Picardeau, M., et al. Genome sequence of the saprophyte Leptospira biflexa provides insights into the evolution of Leptospira and the pathogenesis of leptospirosis. PLoS One. 3 (2), 1607(2008).
- Fernandes, L. G., et al. OmpL1 is an extracellular matrix- and plasminogen-interacting protein of Leptospira spp. Infections and Immunity. 80 (10), 3679-3692 (2012).
- Fernandes, L. G., et al. Leptospira spp.: Novel insights into host-pathogen interactions. Veterinary Immunology and Immunopathology. 176, 50-57 (2016).
- Castiblanco-Valencia, M. M., et al. Leptospiral immunoglobulin-like proteins interact with human complement regulators factor H, FHL-1, FHR-1, and C4BP. Journal of Infectious Diseases. 205 (6), 995-1004 (2012).
- Choy, H. A., et al. The multifunctional LigB adhesin binds homeostatic proteins with potential roles in cutaneous infection by pathogenic Leptospira interrogans. PLoS One. 6 (2), 16879(2011).
- Siqueira, G. H., et al. The recombinant LIC10508 is a plasma fibronectin, plasminogen, fibrinogen and C4BP-binding protein of Leptospira interrogans. Pathogen and Diseases. 74 (2), (2016).
- Teixeira, A. F., et al. Features of two new proteins with OmpA-like domains identified in the genome sequences of Leptospira interrogans. PLoS One. 10 (4), 0122762(2015).
- Kochi, L. T., et al. The interaction of two novel putative proteins of Leptospira interrogans with E-cadherin, plasminogen and complement components with potential role in bacterial infection. Virulence. 10 (1), 734-753 (2019).
- Murray, G. L., et al. Genome-wide transposon mutagenesis in pathogenic Leptospira species. Infections and Immunity. 77 (2), 810-816 (2009).
- Bourhy, P., Louvel, H., Saint Girons, I., Picardeau, M. Random insertional mutagenesis of Leptospira interrogans, the agent of leptospirosis, using a mariner transposon. Journal of Bacteriology. 187 (9), 3255-3258 (2005).
- Pětrošová, H., Picardeau, M. Screening of a Leptospira biflexa mutant library to identify genes involved in ethidium bromide tolerance. Applied Environmental Microbiology. 80 (19), 6091-6103 (2014).
- Slamti, L., Picardeau, M. Construction of a library of random mutants in the spirochete Leptospira biflexa using a mariner transposon. Methods in Molecular Biology. 859, 169-176 (2012).
- Croda, J., et al. Targeted mutagenesis in pathogenic Leptospira species: Disruption of the LigB gene does not affect virulence in animal models of leptospirosis. Infections and Immunity. 76 (12), 5826-5833 (2008).
- Picardeau, M., Brenot, A., Saint Girons, I. First evidence for gene replacement in Leptospira spp. Inactivation of L. biflexa flaB results in non-motile mutants deficient in endoflagella. Molecular Microbiology. 40 (1), 189-199 (2001).
- King, A. M., et al. High-temperature protein G is an essential virulence factor of Leptospira interrogans. Infections and Immunity. 82 (3), 1123-1131 (2014).
- Lambert, A., et al. FlaA proteins in Leptospira interrogans are essential for motility and virulence but are not required for formation of the flagellum sheath. Infections and Immunity. 80 (6), 2019-2025 (2012).
- Murray, G. L., et al. Leptospira interrogans requires heme oxygenase for disease pathogenesis. Microbes and Infections. 11 (2), 311-314 (2009).
- Ristow, P., et al. The OmpA-like protein Loa22 is essential for leptospiral virulence. PLoS Pathogens. 3 (7), 97(2007).
- Sasaki, Y., et al. Leptospiral flagellar sheath protein FcpA interacts with FlaA2 and FlaB1 in Leptospira biflexa. PLoS One. 13 (4), 0194923(2018).
- Gaj, T., Gersbach, C. A., Barbas, C. F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends in Biotechnology. 31 (7), 397-405 (2013).
- Cress, B. F., et al. Rapid generation of CRISPR/dCas9-regulated, orthogonally repressible hybrid T7-lac promoters for modular, tuneable control of metabolic pathway fluxes in Escherichia coli. Nucleic Acids Research. 44 (9), 4472-4485 (2016).
- Zhao, C., Shu, X., Sun, B. Construction of a gene knockdown system based on catalytically inactive ("dead") Cas9 (dCas9) in Staphylococcus aureus. Applied Environmental Microbiology. 83 (12), (2017).
- Zhao, Y., et al. CRISPR/dCas9-mediated multiplex gene repression in Streptomyces. Biotechnology Journal. , 1800121(2018).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved