
Method Article
Applicazione dell'interferenza CRISPR (CRISPRi) per il silenziamento genico nelle specie patogene di Leptospira
In questo articolo
Riepilogo
Qui viene descritta l'applicazione dell'interferenza CRISPR (CRISPRi) per un silenziamento genico specifico nelle specie di Leptospira. Le cellule leptospira sono trasformate coniugando con plasmidi che esprimono dCas9 (cas9 cataliticamente "morto") e un RNA a guida singola (sgRNA), responsabile dell'accoppiamento di base al bersaglio genomico desiderato. Vengono presentati metodi per convalidare il silenziamento genico.
Abstract
La leptospirosi è una zoonosi trascurata a livello globale, responsabile di almeno 1 milione di casi all'anno e quasi 60 mila decessi. La malattia è causata da batteri patogeni e virulenti del genere Leptospira, sia per contatto diretto con i batteri che indirettamente per esposizione ad acqua o suolo contaminati. Gli animali domestici e selvatici agiscono come ospiti serbatoi di infezione, versando leptospire da tubuli renali colonizzati del rene, attraverso l'urina, nell'ambiente. La generazione di ceppi mutanti di Leptospira è fondamentale per valutare e comprendere i meccanismi patogeni dell'infezione. L'interferenza CRISPR (CRISPRi) si è dimostrata uno strumento semplice, conveniente e specifico per il silenziamento genico nel Leptospira patogeno. Pertanto, verranno descritti i dettagli metodologici per ottenere i costrutti plasmidi contenenti sia dCas9 che RNA guida, la consegna di plasmidi a Leptospira coniugazione con il ceppo E. coli β2163 e il recupero e la valutazione transcongiunti. Inoltre, i supporti Hornsby-Alt-Nally (HAN) recentemente descritti consentono l'isolamento relativamente rapido e la selezione delle colonie mutanti sulle placche di agar.
Introduzione
La leptospirosi è una zoonosi trascurata in tutto il mondo causata da specie patogene e virulenti del genere Leptospira. Nell'uomo, la malattia rappresenta più di un milione di casi e 60.000 decessi all'anno intutto il mondo 1,2. Finora non esiste un vaccino efficace e a lungo termine per la malattia. L'identificazione dei fattori di virulenza e dei meccanismi patogeni è fondamentale per lo sviluppo di migliori strategie terapeutiche e profilattiche. Pertanto, la capacità di generare mutazioni genetiche e valutare il fenotipo risultante è fondamentale per l'analisi genomica funzionale3.
La costruzione di mutanti in Leptospira patogeno era considerata, fino ad ora, intrinsecamente inefficiente, laboriosa, costosa e difficile da implementare. Questo scenario è cambiato drasticamente con l'applicazione della recente interferenza CRISPR (CRISPRi) alle leptospire saprofite4 e patogene 5.
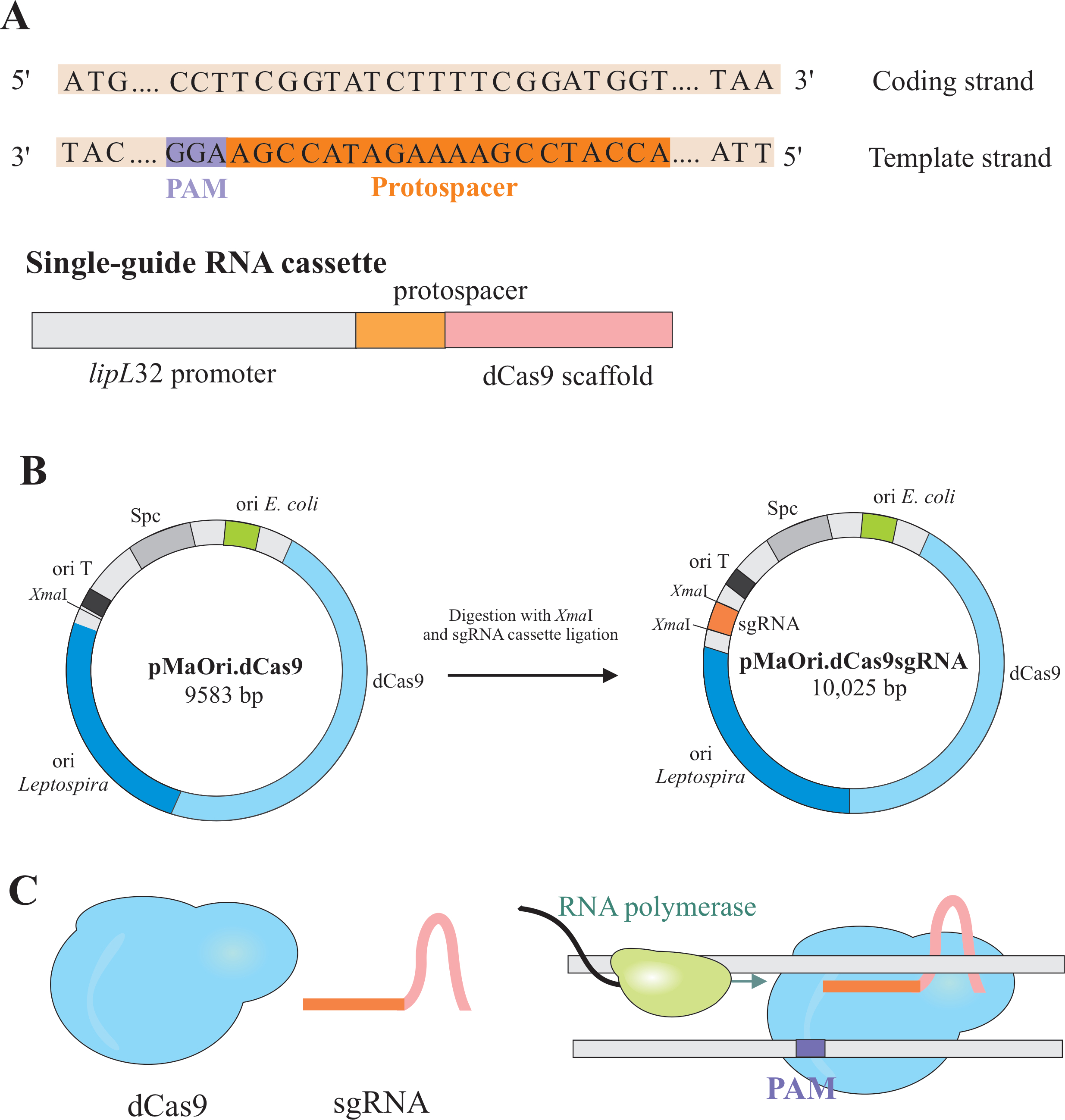
Il silenziamento genico si ottiene con l'espressione di due componenti: una variante dell'enzima CRISPR/Cas(clustro regularly interspaced short palindromic repeat/CRISPR associated) Cas9 da Streptococcus pyogenes, chiamato Catalytically dead Cas9 (dCas9) e un RNA a guida singola (sgRNA), che può essere modificato in base al target desiderato6,7,8. La proteina dCas9, quando legata allo sgRNA, è diretta a specifici bersagli di DNA dall'accoppiamento di base di Watson e Crick, causando un blocco sterico all'allungamento dell'RNA polimerasi, con conseguente silenziamento genico a causa della trascrizione genicaostruita 7 (Figura 1).
Questo manoscritto ha lo scopo di descrivere la costruzione del plasmide per esprimere sia dCas9 che sgRNA, coniugazione tra il donatore E. coli β2163 e le cellule leptospira riceventi, recupero transcongiuntino e, infine, convalida di colonie mutanti selezionate.
Protocollo
1. Definizione protospaziale e costruzione plasmide
NOTA: In questa sezione viene descritto il primo passaggio della selezione dei protospaziatori appropriati per la costruzione dello sgRNA e l'ulteriore ligazione in pMaOri.dCas9 (Figura 1). Questa sequenza protospaziale comprende una sequenza di 20 nucleotidi contro il bersaglio desiderato.
- Ottenere la sequenza nucleotidica del gene di interesse per il silenziamento a GenBank (https://www.ncbi.nlm.nih.gov/genbank). Invialo al server web CHOPCHOP (http://chopchop.cbu.uib.no/), con parametri definiti per Streptococcus pyogenes Cas9 e il motivo adiacente protospaziale NGG dopo aver selezionato il "Fasta Target". Definite i parametri "CRISPR/Cas9" e PAM (protospazio adiacente motif) NGG.
- Sulla base dei risultati ottenuti, selezionare i protospaziatori con il miglior punteggio possibile (freccia verde), che si trovano il più vicino possibile alla fine 5'della regione codificante e, soprattutto, sono contenuti nel filamento modello (meno) poiché lo sgRNA deve essere accoppiato al filamento codificante del gene per il silenziamento genico completo.
NOTA: Il motivo NGG non è incluso nella sequenza finale di sgRNA. - Utilizzare il promotore lipL32 per esprimere l'RNA a guida singola che contiene una sequenza nucleotidica variabile 20 all'estremità 5' e una sequenza di impalcature dCas9 conservata. Unire la sequenza di 20 nt, definita protospazior, al promotore lipL32 (alla sua estremità di 5') e all'impalcatura sgRNA (3'end)(Figura 1B).
NOTA: Per un promotore lipL32 ben definito, utilizzare la regione promotrice che comprende -334 per il TSS (Transcription Start Site, basato su Zhukova et al.9). Controllare il file supplementare per la cassetta sgRNA finale. - Generare la cassetta sgRNA tramite PCR5 sequenziale o sintetizzare da un provider commerciale.
- Dopo aver ottenuto la cassetta, ligatela in pMaOri.dCas9 plasmide presso il sito di restrizione XmaI ad entrambe le estremità (cccggg)4.
- Digerire sia la cassetta sgRNA che il plasmide pMaOri.dCas9 con enzima di restrizione XmaI e procedere alla legatura (Figura 1B).
- Eseguire i passaggi di clonazione nel ceppo dT auxotrophic E. coli π110, a causa dell'origine pMaOri11 (e per estensione, pMaOri.dCas9) della replicazione, R6K-gamma.
NOTA: Per un protocollo dettagliato per la legatura e la selezione dei cloni, fare riferimento alle precedenti pubblicazioni di Fernandes e Nascimento12. DCas9 guidato da sgRNA si legherà al filamento codificante del gene di interesse selezionato e, pertanto, ostacolerà l'allungamento dell'RNA polimerasi (Figura 1C), con conseguente silenziamento genico.
2. Trasformazione di Leptospira per coniugazione
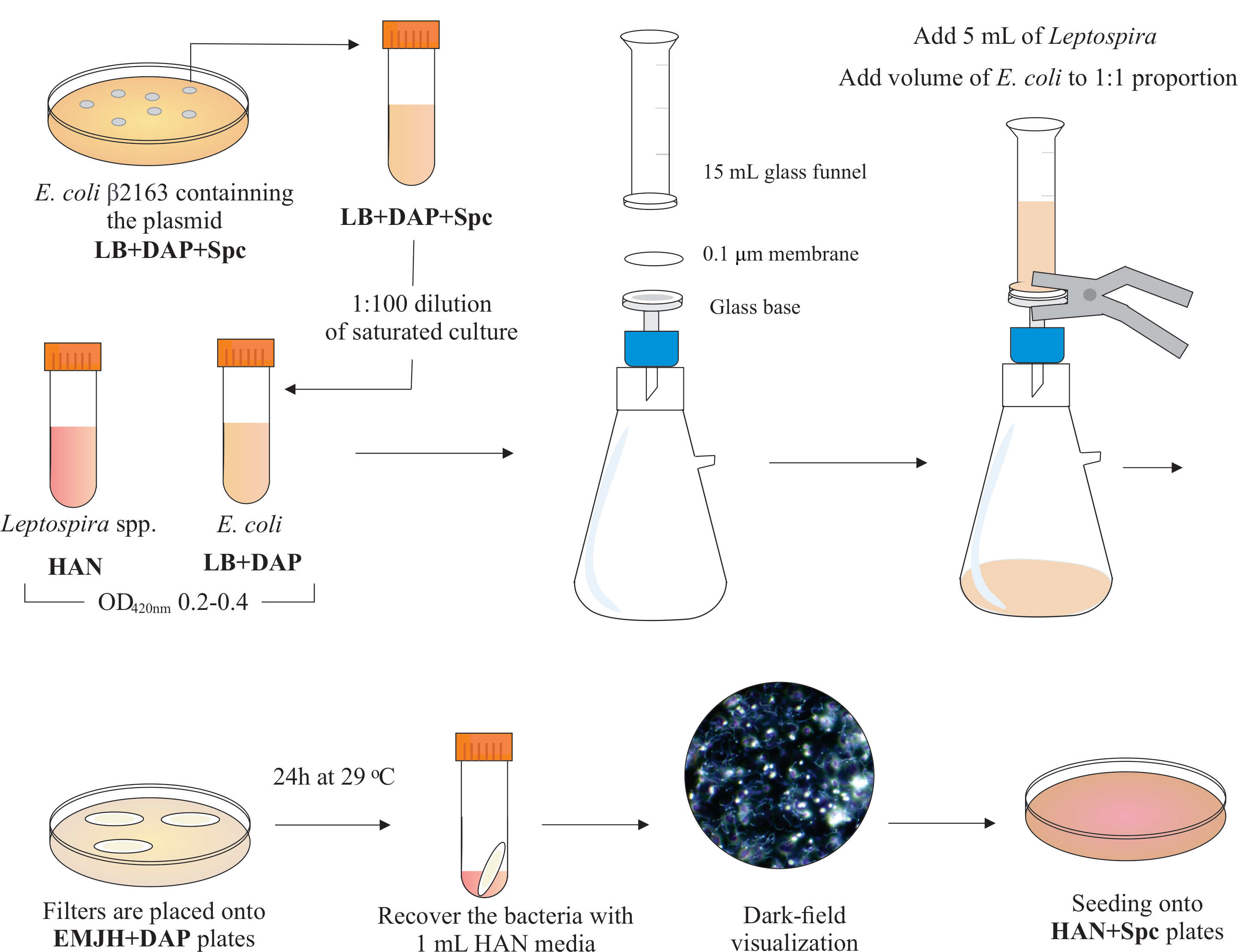
NOTA: Uno schema grafico di questo passaggio è presentato nella figura 2. Per realizzare supporti HAN e piastre HAN, fare riferimento a Hornsbyetal.
- Far crescere cellule leptospira patogene a 29 o 37 °C nei mezzi HAN13 sotto agitazione diluendo una coltura satura in HAN fresco a 1:100; tipicamente, L. interrogans serovar Copenhageni strain Fiocruz L1-130 impiega 4-6 giorni per raggiungere la densità cellulare appropriata.
- Assicurarsi che le colture raggiungano un 0,2-0,4 O.D. a 420 nm (da 2 a 5 x 108 celle/mL) prima di utilizzarle per la coniugazione.
NOTA: Poiché i mezzi HAN cambiano colore con l'aumentare della densità cellulare (a causa del rosso fenolo contenuto nei supporti DMEM), centrifuga (4.000 x g, 15 min, temperatura ambiente) 1 mL del supporto di coltura per rimuovere i leptospiri e applicare il supernatante come vuoto per misurare l'O.D.
- Assicurarsi che le colture raggiungano un 0,2-0,4 O.D. a 420 nm (da 2 a 5 x 108 celle/mL) prima di utilizzarle per la coniugazione.
- Trasformare il ceppo coniugato E. coli β2163, auxotrofico per l'acido diaminopimelico (DAP), con il pMaOri.dCas9 plasmide contenente la cassetta sgRNA. Per E. trasformazione coli, utilizzare protocolli di shock termico o elettroporazione. Includere la trasformazione con il pMaOri.dCas9 plasmide senza cassetta sgRNA come controllo.
- Per la trasformazione dello shock termico, mescolare il DNA plasmide (100 ng) con cellule E. coli chimicamente competenti e incubare sul ghiaccio per 30 minuti. Eseguire uno shock termico a 42 °C per 90 s e posizionarlo nuovamente sul ghiaccio per 5 minuti. Recuperare le cellule aggiungendo 1 mL di mezzi LB, incubare a 37 °C per 1 h e procedere alla placcatura.
- Per l'elettroporazione, utilizzare cellule elettrocompetenti mescolate con 100 ng di DNA plasmide. Utilizzare i seguenti parametri per l'impulso: 1,8 kV, 100 Ω e 25 μF. Recuperare le cellule come spiegato sopra.
- Placcare le cellule E. coli del donatore trasformato in mezzo agar LB integrato con acido diaminopimelico (DAP) (0,3 mM) e spectinomicina (40 μg/mL) da selezionare per i plasmidi.
- Per la coniugazione, selezionare una colonia da ogni piastra un giorno prima del giorno di coniugazione (che è determinato monitorando l'O.D. delle colture di leptospiri).
- Selezionate una colonia di E. coli β2163 dalla pMaOri.dCas9 vuota e una dalle placche pMaOri.dCas9sgRNA. Consentire loro di crescere durante la notte in 10 mL di LB più DAP e spectinomicina a 37 °C.
- Il giorno dopo, diluire le colture sature 1:100 in 10 mL di LB fresco più DAP (non includere l'antibiotico qui) fino a OD420nm di 0,2-0,4. Normalmente, ci vogliono 2-3 ore perché E. coli raggiunga queste densità.
- All'interno di una cappa di biosicurezza BSL2, assemblare un apparecchio di filtrazione posizionando un diametro di 25 mm, dimensioni dei pori da 0,1 μm, filtro a membrana per esteri di cellulosa mista sulla parte superiore della base di vetro. Posizionare un imbuto in vetro da 15 mL sulla parte superiore e tenere entrambi i pezzi con morsetti a molla. Collegare il vetro a una pompa per vuoto e aggiungere le colture all'imbuto per la filtrazione.
- Aggiungi 5 mL di cultura di Leptospira all'imbuto. Aggiungere un volume di E. coli per costituire la proporzione 1:1 basata sui valori OD420nm di entrambe le culture. Accendere la pompa per vuoto e concentrare le celle mediante filtrazione. Dopo la concentrazione cellulare nel filtro a membrana, recuperarlo con cura. Assicurarsi che il mezzo sia filtrato attraverso la membrana.
NOTA: La filtrazione richiede da 5 a 10 minuti. - Posizionare il filtro su una piastra EMJH disponibile in commercio (vedi Tabella dei materiali)integrata con DAP (0,3 mM). Assicurarsi che il lato batteri sia in alto. Incubare le piastre a 29 °C per 24 ore.
NOTA: Se vengono utilizzate piastre HAN o EMJH14 integrate internamente, E. coli può proliferare e superare la proporzione 1:1 prevista, che a sua volta può ridurre l'efficienza coniugazione5. - Dopo 24 ore, recuperare i filtri dalle piastre e posizionare ogni singolo filtro in un tubo conico da 50 ml.
- Utilizzare 1 mL di mezzo HAN liquido per rilasciare le celle dalla superficie del filtro mediante pipettazione e vortice estesi.
- Visualizza le soluzioni batteriche miste recuperate mediante microscopia a campo scuro per verificare la vitalità e la motilità cellulare e le proporzioni leptospira:E. coli.
NOTA: In questa fase, si possono vedere numeri equivalenti di E. coli e Leptospira. - Stendere 100-200 μL di questa coltura su piastre HAN contenenti siero di coniglio inattivato dello 0,4% e 40 μg/mL di spectinomicina. Incubare le piastre a 37 °C in un'atmosfera di CO2 del 3%.
NOTA: Normalmente, le cellule di L. interrogans serovar Copenhageni strain Fiocruz L1-130 formano colonie in 5-7 giorni su piastre di controllo e in 8-10 giorni su piastre di spectinomicina. In questa fase, E. coli non crescerà poiché sono auxotrofici per DAP. - Come controllo, diluire le colture a 104 leptospires/mL e aggiungere 100 μL su piastre senza antibiotico, per monitorare la crescita leptospirale.
3. Selezione e crescita e convalida delle colonie e transconjugant
NOTA: Le colonie dovrebbero essere evidenti entro il giorno 10. Tuttavia, non sono troppo facili da visualizzare. Normalmente in questo momento, le piastre HAN sono un po 'opache a causa delle cellule essiccate che sono state diffuse e le colonie di Leptospira possono apparire come un alone trasparente sullo sfondo biancastro. Si consiglia di visualizzare le piastre da diverse angolazioni per ottenere una diversa incidenza della luce, quindi, rendendo le colonie più evidenti. A tempi di incubazione più lunghi, le colonie possono acquisire un aspetto più denso e, in questo caso, si presentano come aloni lattici su uno sfondo scuro.
- Aggiungere 100 μL di supporti HAN liquidi a ogni microtubo da 1,5 mL per recuperare i mutanti. Prendi almeno 3 colonie da ogni piatto.
- Con l'aiuto di una punta di micropipetta, "scavare" l'agar per recuperare le colonie dalle placche poiché le colonie leptospirali possono essere sottosuperficie.
NOTA: Agar dovrebbe essere portato con sé in questa fase. Le colonie devono essere prelevate da piastre di controllo contenenti pMaOri.dCas9 plasmide vuoto e piastre con leptospiri contenenti plasmidi che esprimono sia dCas9 che RNA a guida singola, progettati per il gene bersaglio. - Erogare la colonia raccolta in 100 μL di mezzi HAN in un microtubo da 1,5 mL e omogeneizzare vigorosamente. In questa fase, garantire la massima rottura dell'integrità dell'agar per rilasciare le cellule. Vortice la sospensione per 10 s.
- Con l'aiuto di una punta di micropipetta, "scavare" l'agar per recuperare le colonie dalle placche poiché le colonie leptospirali possono essere sottosuperficie.
- Visualizzare le cellule recuperate mediante microscopia a campo scuro con un ingrandimento di 200-400x aggiungendo una goccia di 5 μL su uno scivolo di vetro e coprire immediatamente i campioni con un coverslip.
- Confermare la presenza di leptospire vivi e vitali recuperati dalle colonie.
- Dopo la visualizzazione e la conferma di leptospires vitali, trasferire 100 μL di cellule su supporti HAN liquidi contenenti 40 μg/mL di spectinomicina.
- Dopo la crescita dei mezzi HAN liquidi, valutare le colture per la presenza del plasmide con primer pMaOri2 F (ACGCAATGTATCGATACCGAC) e R (ATAGGTGAAGTAGGCCCACCC), che riconoscono la regione che fiancheggia la cassetta sgRNA.
- Raccogliere 200 μL di coltura, centrifuga (4.000 x g, 15 min), scartare il supernatante e rimescolare il pellet risultante in 20 μL di acqua.
- Utilizzare questa sospensione come modello per pcr aggiuntivi, senza la necessità di estrazione del DNA12.
NOTA: Le cellule con pMaOri.dCas9 renderanno un'amplicon di 281 bp, rispetto a quelle contenenti i plasmidi con la cassetta sgRNA che renderà un'amplicon di 723 bp.
- Per la conferma del silenziamento genico, eseguire un immunoblot utilizzando estratti cellulari da transconjuganti contenenti solo pMaOri.dCas9 (controllo negativo) e pMaOri.dCas9sgRNA.
- Applicare l'equivalente di 5 x 107 cellule per corsia del gel di poliacrilammide dodecil solfato di sodio (SDS).
- Elettrotrasferire le proteine a una membrana per l'incubazione con anticorpi appropriati. Oltre all'anticorpo contro il gene bersaglio per il silenziamento, usane un altro per il controllo del carico.
- Mantieni le colture mutanti nei media HAN più la spectinomicina per mantenere il plasmide. Se non vengono applicati antibiotici al supporto, è possibile osservare un silenziamento genico completo per almeno tre passaggi5.
Risultati
Anche se il contenuto di CG nei genomi di Leptospira spp. è probabile che praticamente ogni gene contenga il PAM 5'NGG 3'; questo motivo deve essere considerato nel filo del modello. Dopo aver inserito la sequenza di codifica di un gene (dall'inizio alla fine dei codoni), in base ai risultati di CHOPCHOP, i protospaziatori devono essere selezionati nel filamento meno (-, modello). È importante non includere il motivo NGG nel protospazio sgRNA da 20 nt.
Se la coniugazione viene eseguita con una proporzione di cellule donatrici:riceventi 1:1, per 24 ore sulla superficie delle piastre di agar EMJH più DAP, e 200 μL della sospensione batterica recuperata vengono distribuite su piastre di agar HAN più spectinomicina, le colonie di transconijuganti dovrebbero essere visibili a circa 8-10 giorni. La diffusione di questo volume si traduce normalmente in 20-40 colonie per piastra (Figura 3A). Al fine di verificare la vitalità cellulare dopo la coniugazione, le cellule possono essere diffuse su piastre HAN senza selezione di antibiotici. In questo caso, le colonie possono essere osservate non appena 7 giorni. Le piastre HAN diventano giallo pallido in un'atmosfera di CO2 del 3%.
Dopo la raccolta e la crescita della colonia nei mezzi liquidi più la spectinomicina, la PCR utilizzando cellule intere e primer pMaOri2 può essere utilizzata per un primo controllo di qualità dei transcongiunti (Figura 3B). Le cellule leptospirali contenenti il plasmide pMaOri.dCas9 di controllo dovrebbero provocare un'amplicone di 281 bp, mentre quelle cellule contenenti il plasmide per il silenziamento, cioè contenenti sia dCas9 che sgRNA, dovrebbero causare un amplicon di 723 bp. I primer pMaOri2 F e R sono stati progettati per fiancheggiare il sito di restrizione XmaI, che è il sito utilizzato durante la legatura della cassetta sgRNA.
Con la conferma della presenza di plasmide, le cellule possono essere raccolte dal supporto, lavate due volte con PBS e quindi utilizzate per preparare un estratto di cellule intere per l'immunoblotting. Se si è verificato il silenziamento, le proteine bersaglio, in questo caso LipL32 o sia LigA che LigB, devono essere osservate solo nelle cellule di tipo selvatico e in quelle contenenti pMaOri.dCas9; anche con tempi di esposizione più elevati, nessuna proteine corrispondente dovrebbe essere visibile nelle cellule contenenti pMaOri.dCas9sgRNA (Figura 3C).
Se sono previsti esperimenti per valutare la virulenza leptospirale dopo il silenziamento genico, le colture utilizzate per la coniugazione devono essere leptospiravirulenti a basso passaggio . Dopo la conferma del silenziamento genico, diverse aliquote possono essere congelate come riserva. Se il gene silenziato ha un fenotipo misurabile, ad esempio, basato su precedenti lavori con proteine ricombinanti, le colture possono essere utilizzate per la convalida e, in questo caso, le cellule contenenti solo pMaOri.dCas9, possono essere incluse come controllo negativo.

Figura 1: Sviluppo di dCas9 e sgRNA che esprimono plasmide. (A) Un protospazio lungo 20 nt, seguito dal S. pyogenes dCas9 PAM 5'-NGG-3', è selezionato all'interno del filamento modello del gene bersaglio in modo che il successivo sgRNA possa eseguire l'accoppiamento di base Watson e Crick al filamento di codifica corrispondente, con conseguente completo silenziamento genico. (B) La cassetta sgRNA è composta dal promotore lipL32, dal protospazio da 20 nt e dall'impalcatura dCas9. Il plasmide pMaOri.dCas9 è usato come spina dorsale per la legatura della cassetta sgRNA nel sito di restrizione XmaI. Il plasmide risultante, fonato pMaOri.dCas9sgRNA viene consegnato ai leptospires, e l'espressione di dCas9 e sgRNA è responsabile del silenziamento genico. (C) dCas9 diretto da sgRNA funge da barriera fisica all'allungamento dell'RNA polimerasi, quindi, ostacolando la trascrizione. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Rappresentazione schematica del protocollo di coniugazione. La specie di Leptospira desiderata viene coltivata nei media HAN, sotto agitazione, fino a 0,2-0,4 (fase di registro intermedio) a 420 nm. Un giorno prima della coniugazione, una colonia di donatore ricombinante E. coli β2163 contenente il plasmide di interesse viene raccolta dalle piastre di agar LB+DAP+Spc, poiché le cellule vengono coltivate durante la notte in LB liquido con la stessa integrazione. Il giorno dopo, saturo E. le colture di coli vengono diluite in LB più DAP e coltivate fino a 0,2-0,4 a 420 nm. Sia il donatore E. coli che il ricevente Leptospira sono mescolati a una proporzione cellulare di 1:1 sulla superficie di un filtro da 0,1 μm da un apparato di filtrazione sotto pressione negativa. Quindi, i filtri vengono posizionati sopra le piastre di agar EMJH integrate con DAP e l'incubazione procede per 24 ore a 29 °C. L'uso di EMJH limita la proliferazione di E. coli e la proporzione 1:1 prevista viene mantenuta. I batteri vengono recuperati dai filtri con pipettazione con mezzi HAN da 1 mL e le sospensioni vengono visualizzate in microscopia a campo scuro. Infine, 100-200 μL di ciascuna sospensione vengono seminati su piastre di agar HAN contenenti siero di coniglio allo 0,4% e incubati a 37 °C nel 3% di CO2. In questa fase, il DAP viene omesso e, di conseguenza, l'auxotrofico E. coli non crescerà. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Risultati rappresentativi per la valutazione dei mutanti. (A) Le colonie di piastre contenenti Leptospira trasformate con pMaOri.dCas9 vuoto (controllo negativo per ulteriori esperimenti) e plasmidi pMaOri.dCas9sgRNA (con gene mirato silenziato) vengono raccolte, vigorosamente omogeneizzate in HAN liquido e coltivate in HAN liquido contenente spectinomicina. Le cellule ricombinanti possono essere convalidate da PCR con primer che fiancheggiano il sito XmaI all'interno di pMaOri.dCas9. (B) In questo caso, le cellule contenenti pMaOri.dCas9 hanno prodotto solo un'amplicone di 281 bp, mentre quelle cellule contenenti il plasmide per il silenziamento, contenenti sia dCas9 che sgRNA, hanno mostrato un amplicone di 723 bp. Dopo la conferma della presenza dei plasmidi, il silenziamento genico è stato convalidato dall'analisi immunoblot. (C) Si raccomanda l'incubazione con anticorpi sia per le proteine bersaglio che per una proteina di controllo del carico; nell'immunoblot rappresentativo, vengono visualizzati estratti a cellule intere da transconjuganti contenenti pMaOri.dCas9 da soli o con cassette sgRNA rivolte ai geni lipL32 (pMaOri.dCas9sgRNAlipL32) e sia LigA che LigB (pMaOri.dCas9sgRNAligAB). La co-incubazione con anti-LipL32, anti-LigAB e anti-LipL41 (non bersaglio, controllo del carico) conferma che l'espressione della proteina LipL32 è abolita nelle cellule contenenti pMaOri.dCas9sgRNAlipL32 e sia LigA che LigB in cellule contenenti pMaOri.dCas9sgRNAligAB. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
File supplementare: Sequenza di cassette a RNA (sgRNA) a guida singola. La trascrizione dello sgRNA è diretta dal promotore costitutivo lipL32 (nucleotidi audaci). lo sgRNA è composto da 20 nucleotidi riferiti al protospaziore, responsabile dell'accoppiamento di base al filamento codificante del gene bersaglio, e dalla sequenza di impalcature dCas9 (nucleotidi sottolineati). Xma I siti di restrizione (cccggg) sono inclusi ad entrambe le estremità per la legatura al plasmide pMaOri.dCas9. Clicca qui per scaricare questo file.
Discussione
Dopo il sequenziamento precoce delle specie patogene15,16,17,18 e saprophytic19Leptospira, l'estrazione dei dati del genoma ha fatto luce su diversi aspetti della patogenesi leptospirale. Nella maggior parte dei casi, la funzione proteica è stata esplorata utilizzando la controparte ricombinante di proteine putative leptospirali esposte in superficie e la successiva speculazione della funzione proteica nativa20,21,22,23,24,25,26.
La generazione di mutanti, e la valutazione del loro rispettivo fenotipo, sono componenti chiave dell'analisi genomica funzionale. I primi tentativi di generare mutantiin Leptospira spp. tuttavia, dopo un'analisi approfondita e laboriosa per aver dedotto l'identità dei geni interrotti, è stato notato che solo il 15% di tutti i geni in L. interrogans serovar Manilae sono statiinterrotti 27. Il knockout genico mirato è stato ulteriormente raggiunto mediante ricombinazione omologa utilizzando plasmidi suicidi per fornire una cassetta di resistenza agli antibiotici affiancata da bracci omologhi all'interno del bersagliodesiderato 31,32.
Applicando queste tecnologie, sono stati esplorati diversi aspetti della biologia di base leptospirale e della virulenza31,33,34,35,36,37. Lo sviluppo del vettore navetta coniugale E. coli-Leptospira, pMaOri 11, ha permesso la consegna di componenti per silenziamento genico episomale.
È stato precedentemente dimostrato che la rottura a doppio filamento indotta da Cas9 è letale per Leptospira spp. Utilizzando il plasmide pMaOri.dCas9 come spina dorsale per la legatura a cassetta sgRNA, è possibile ottenere silenziamento genico specifico e stabile grazie all'espressione sia di dCas9 che di sgRNA; Lo sgRNA legato a dCas9 porterà la proteina al bersaglio desiderato dall'accoppiamento di base Watson-Crick.
Per un silenziamento genico completo, il protospaziore deve essere progettato in base al filamento modello del gene desiderato in modo che l'accoppiamento di base dello sgRNA avvenga con il filamento codificante. Sulla base di un contenuto medio di C+G del 35% in Leptospira spp., il PAM 5'-NGG-3' si verificherà almeno 3 volte ogni 100 bp. Pertanto, praticamente qualsiasi gene all'interno del genoma di Leptospira conterrà almeno un PAM. Tuttavia, se il motivo NGG non viene trovato, è possibile valutare il motivo NAG alternativo.
Le precedenti tecniche di silenziamento genico, come le dita di zinco e TALE (effettori attivatori di trascrizione), si basavano sulla costruzione di una proteina diversa per ogni bersaglio, rendendo queste tecniche laboriose ecostose 38. Nel caso di CRISPRi, il componente variabile è lo sgRNA, rendendo necessario modificare solo il 20 bp alla fine di 5 '. Il silenziamento genico completo, stabile e mirato è stato osservato non solo in Leptospira spp.4,5, ma anche in altribatteri 8,39,40,41.
Lo sviluppo dei media HAN13 favorì il recupero dei mutanti riducendo drasticamente i tempi di incubazione per la formazione della colonia e permettendo a Leptospira di crescere a 37 oC. Tuttavia, durante la fase di coniugazione, il suo uso non è raccomandato poiché E. coli può proliferare vigorosamente in questo mezzo e superare la proporzione 1:1 prevista tra le cellule donatrici e ricevente. In questa fase, EMJH più DAP è la scelta migliore, dal momento che E. coli si replica male in questo supporto. Vale la pena ricordare che alcuni laboratori protetiamo EMJH integrato internamente, che può contenere componenti aggiuntivi che potrebbero anche sostenere la crescita delle cellule E. coli.
Il protocollo di coniugazione qui presentato è stato ottimizzato per il ceppo L. interrogans serovar Copenhageni Fiocruz L1-130, ed è stato anche dimostrato essere efficace nella trasformazione di un ceppo patogeno recentemente isolato dai campioni di terreno5. I primi tentativi con diversi sierovar di specie L. borgpetersenii indicano minori efficienze coniuganti con il protocollo descritto. Pertanto, quando si lavora con diverse specie/sierovar di Leptospira, le condizioni ottimali per la coniugazione devono essere determinate empiricamente, considerando le proporzioni delle cellule donatrici:riceventi, le densità iniziali delle cellule, i mezzi di coniugazione e il tempo (24 e 48 ore). È ragionevole supporre che diverse specie di Leptospira e sierovar si comporteranno in modo diverso con diversi protocolli di coniugazione.
Anche se le colonie saprofite di Leptospira sono relativamente facili da visualizzare sulle placche, le colonie patogene possono essere più difficili da osservare. Normalmente, utilizzando mezzi HAN integrati con siero di coniglio allo 0,4% e spectinomicina, le colonie transconigiunti possono essere osservate al giorno 10. Nella nostra esperienza, le colonie inizialmente presenti come un alone trasparente sulla superficie dei media. Nel protocollo video, le colonie più dense, dopo 14 giorni di crescita, vengono mostrate poiché quelle trasparenti erano difficili da filmare. In questa fase, ruotare la piastra per ottenere una diversa incidenza della luce e spostarsi tra sfondi bianchi e scuri può aiutare a identificare le colonie.
Per la convalida mutante, l'immunoblotting offre un approccio semplice; tuttavia, poiché gli anticorpi non sono sempre disponibili contro le proteine bersaglio, è possibile perseguire strategie alternative per convalidare il silenziamento genico. La PCR quantitativa a trascrizione inversa (qRT-PCR) che utilizza primer per il gene bersaglio e un controllo costitutivo è efficace per convalidare il silenziamento genico poiché dCas9 guidato dallo sgRNA è responsabile del blocco della trascrizione genica. Se il gene bersaglio codifica una banda proteica chiaramente definita nei gel proteici, SDS-PAGE può dimostrare silenziamento e, secondo il gene lipL32 silenziamento5. Se i geni della biosintesi LPS vengono silenziati, è possibile utilizzare la colorazione LPS; nel caso della codifica dei geni silenzianti per enzimi con substrati ben definiti, i test cinetici con substrati cromogenici sono strategie valide; β silenziamento della β-galattosidasi in L. biflexa è stato convalidato dall'uso di substrati X-gal e ONPG (orto-Nitrofenil-β-galattoside)4.
Dopo la conferma del silenziamento genico, gli esperimenti possono essere progettati per valutare ulteriormente il fenotipo. I test di legame possono essere eseguiti in caso di silenziamento di adesine batteriche; i saggi siero-sfida hanno confermato il ruolo di LigA e LigB nella sopravvivenza del siero mostrato dalla patogena Leptospira5. I mutanti possono anche essere usati per inoculare gli animali per valutare l'attenuazione della virulenza; in questo caso, gli animali inoculati con il mutante devono essere confrontati con quelli infetti da cellule contenenti solo pMaOri.dCas9.
In conclusione, l'attuale protocollo descrive l'applicazione di CRISPRi per il silenziamento genico nelle specie patogene di Leptospira utilizzando i media HAN per facilitare il recupero mutante entro 10 giorni. Il silenziamento genico combinato con l'analisi genomica funzionale migliorerà la nostra comprensione dei meccanismi patogeni di Leptospirae, in ultima analisi, porterà allo sviluppo di migliori strategie profilattiche per il controllo delle malattie.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
L'USDA è un fornitore e un datore di lavoro per le pari opportunità. La menzione di nomi commerciali o prodotti commerciali in questa pubblicazione ha lo scopo di fornire informazioni specifiche e non implica una raccomandazione o un'approvazione da parte del Dipartimento dell'Agricoltura degli Stati Uniti. L'agenzia brasiliana FAPESP (sovvenzione 2014/50981-0) ha sostenuto finanziariamente questo lavoro; LGVF è finanziato con una borsa di studio della FAPESP (2017/06731-8 e 2019/20302-8). I finanziatori non hanno avuto alcun ruolo nella progettazione, raccolta e analisi dei dati, decisione di pubblicare o preparazione di manoscritti. Gli autori ringraziano anche Hannah Hill e Alexander Grimes dell'USDA Visual Services per aver filmato e modificato il protocollo video.
Materiali
| Name | Company | Catalog Number | Comments |
| 0.1 µm pore size mixed cellulose esters membrane | Millipore | VCWP02500 | Filtration for bacterial conjugation |
| 2,6-Diaminopimelic acid (DAP) | Sigma | D1377 | Growth of auxotrophic E. coli β2163 |
| Agar Noble | BD & Company | 214230 | Used for preparation of solid EMJH and HAN plates |
| Bacto Agar | BD & Company | 214010 | Used for preparation of solid LB plates |
| Clarity Western ECL substrate | Biorad | 170-5060 | Chemiluminescent substrate |
| dNTP set | Thermo Fisher | 10297-018 | dNTPs for PCR reaction |
| Glass Microanalysis Filter Holder | Millipore | XX1012530 | Filtration for bacterial conjugation |
| Imaging System | Biorad | ChemiDoc MP | Chemiluminescence detection |
| LB broth, Miller | BD & Company | 244620 | Lysogenic liquid medium for E. coli culturing |
| Leptospira Enrichment EMJH | BD & Company | 279510 | Supplementation of EMJH media |
| Leptospira Medium Base EMJH | BD & Company | 279410 | EMJH medium for Leptospira |
| Mini-PROTEAN TGX Gels 12% | Biorad | 4568043 | Used for polyacrylamide gel eletrophoresis |
| Optical density reader | Molecular Devices | SpectraMax M2 | For optical density measurements of bacterial cultures |
| Phosphate Buffered Saline 7.4 | Sigma | 806552 | Saline solution for washing bacterial pellets |
| Spectinomycin | Sigma | S0692 | Selection of pMaOri backbone plasmids |
| Taq DNA Polymerase | Thermo Fisher | EP0402 | Enyme, buffer and MgCl2 for PCR reaction |
| Thermocycler | Applied Biosystem | GeneAmp PCR System 9700 | Used for PCR reaction cycling |
| Thymidine (dT) | Sigma | T9250 | Growth of auxotrophic E. coli π1 |
| XmaI restriction enzyme | New Englan BioLabs | R0180L | Digestion of plasmids and inserts |
Riferimenti
- Bharti, A. R., et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infectious Disease. 3 (12), 757-771 (2003).
- Costa, F., et al. Global morbidity and mortality of Leptospirosis: A systematic review. PLoS Neglected Tropical Disease. 9 (9), 0003898(2015).
- Shapiro, R. S., et al. A CRISPR-Cas9-based gene drive platform for genetic interaction analysis in Candida albicans. Nature Microbiology. 3 (1), 73-82 (2018).
- Fernandes, L. G. V., et al. Gene silencing based on RNA-guided catalytically inactive Cas9 (dCas9): a new tool for genetic engineering in Leptospira. Science Reports. 9 (1), 1839(2019).
- Fernandes, L. G. V., Hornsby, R. L., Nascimento, A. L. T. O., Nally, J. E. Genetic manipulation of pathogenic Leptospira: CRISPR interference (CRISPRi)-mediated gene silencing and rapid mutant recovery at 37 C. Science Reports. 11 (1), 1768(2021).
- Jinek, M., et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 337 (6096), 816-821 (2012).
- Qi, L. S., et al. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell. 152 (5), 1173-1183 (2013).
- Choudhary, E., Thakur, P., Pareek, M., Agarwal, N. Gene silencing by CRISPR interference in mycobacteria. Nature Communication. 6, 6267(2015).
- Zhukova, A., et al. Genome-wide transcriptional start site mapping and sRNA identification in the pathogen. Frontiers in Cell and Infectious Microbiology. 7, 10(2017).
- Demarre, G., et al. A new family of mobilizable suicide plasmids based on broad host range R388 plasmid (IncW) and RP4 plasmid (IncPalpha) conjugative machineries, and their cognate Escherichia coli host strains. Research in Microbiology. 156 (2), 245-255 (2005).
- Pappas, C. J., Benaroudj, N., Picardeau, M. A replicative plasmid vector allows efficient complementation of pathogenic Leptospira strains. Applied Environmental Microbiology. 81 (9), 3176-3181 (2015).
- Fernandes, L. G. V., Nascimento, A. L. T. O. Specific gene silencing in Leptospira biflexa by RNA-guided catalytically inactive Cas9 (dCas9). Methods in Molecular Biology. 2134, 109-122 (2020).
- Hornsby, R. L., Alt, D. P., Nally, J. E. Isolation and propagation of leptospires at 37 °C directly from the mammalian host. Science Reports. 10 (1), 9620(2020).
- Haake, D. A. Hamster model of leptospirosis. Current Protocols in Microbiology. , Chapter 12, Unit 12E.12 (2006).
- Nascimento, A. L., et al. Comparative genomics of two Leptospira interrogans serovars reveals novel insights into physiology and pathogenesis. Journal of Bacteriology. 186 (7), 2164-2172 (2004).
- Nascimento, A. L., et al. Genome features of Leptospira interrogans serovar Copenhageni. Brazillian Journal of Medical Biology Research. 37 (4), 459-477 (2004).
- Ren, S. X., et al. Unique physiological and pathogenic features of Leptospira interrogans revealed by whole-genome sequencing. Nature. 422 (6934), 888-893 (2003).
- Bulach, D. M., et al. Genome reduction in Leptospira borgpetersenii reflects limited transmission potential. Proceedings of the National Academy of Science U. S. A. 103 (39), 14560-14565 (2006).
- Picardeau, M., et al. Genome sequence of the saprophyte Leptospira biflexa provides insights into the evolution of Leptospira and the pathogenesis of leptospirosis. PLoS One. 3 (2), 1607(2008).
- Fernandes, L. G., et al. OmpL1 is an extracellular matrix- and plasminogen-interacting protein of Leptospira spp. Infections and Immunity. 80 (10), 3679-3692 (2012).
- Fernandes, L. G., et al. Leptospira spp.: Novel insights into host-pathogen interactions. Veterinary Immunology and Immunopathology. 176, 50-57 (2016).
- Castiblanco-Valencia, M. M., et al. Leptospiral immunoglobulin-like proteins interact with human complement regulators factor H, FHL-1, FHR-1, and C4BP. Journal of Infectious Diseases. 205 (6), 995-1004 (2012).
- Choy, H. A., et al. The multifunctional LigB adhesin binds homeostatic proteins with potential roles in cutaneous infection by pathogenic Leptospira interrogans. PLoS One. 6 (2), 16879(2011).
- Siqueira, G. H., et al. The recombinant LIC10508 is a plasma fibronectin, plasminogen, fibrinogen and C4BP-binding protein of Leptospira interrogans. Pathogen and Diseases. 74 (2), (2016).
- Teixeira, A. F., et al. Features of two new proteins with OmpA-like domains identified in the genome sequences of Leptospira interrogans. PLoS One. 10 (4), 0122762(2015).
- Kochi, L. T., et al. The interaction of two novel putative proteins of Leptospira interrogans with E-cadherin, plasminogen and complement components with potential role in bacterial infection. Virulence. 10 (1), 734-753 (2019).
- Murray, G. L., et al. Genome-wide transposon mutagenesis in pathogenic Leptospira species. Infections and Immunity. 77 (2), 810-816 (2009).
- Bourhy, P., Louvel, H., Saint Girons, I., Picardeau, M. Random insertional mutagenesis of Leptospira interrogans, the agent of leptospirosis, using a mariner transposon. Journal of Bacteriology. 187 (9), 3255-3258 (2005).
- Pětrošová, H., Picardeau, M. Screening of a Leptospira biflexa mutant library to identify genes involved in ethidium bromide tolerance. Applied Environmental Microbiology. 80 (19), 6091-6103 (2014).
- Slamti, L., Picardeau, M. Construction of a library of random mutants in the spirochete Leptospira biflexa using a mariner transposon. Methods in Molecular Biology. 859, 169-176 (2012).
- Croda, J., et al. Targeted mutagenesis in pathogenic Leptospira species: Disruption of the LigB gene does not affect virulence in animal models of leptospirosis. Infections and Immunity. 76 (12), 5826-5833 (2008).
- Picardeau, M., Brenot, A., Saint Girons, I. First evidence for gene replacement in Leptospira spp. Inactivation of L. biflexa flaB results in non-motile mutants deficient in endoflagella. Molecular Microbiology. 40 (1), 189-199 (2001).
- King, A. M., et al. High-temperature protein G is an essential virulence factor of Leptospira interrogans. Infections and Immunity. 82 (3), 1123-1131 (2014).
- Lambert, A., et al. FlaA proteins in Leptospira interrogans are essential for motility and virulence but are not required for formation of the flagellum sheath. Infections and Immunity. 80 (6), 2019-2025 (2012).
- Murray, G. L., et al. Leptospira interrogans requires heme oxygenase for disease pathogenesis. Microbes and Infections. 11 (2), 311-314 (2009).
- Ristow, P., et al. The OmpA-like protein Loa22 is essential for leptospiral virulence. PLoS Pathogens. 3 (7), 97(2007).
- Sasaki, Y., et al. Leptospiral flagellar sheath protein FcpA interacts with FlaA2 and FlaB1 in Leptospira biflexa. PLoS One. 13 (4), 0194923(2018).
- Gaj, T., Gersbach, C. A., Barbas, C. F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends in Biotechnology. 31 (7), 397-405 (2013).
- Cress, B. F., et al. Rapid generation of CRISPR/dCas9-regulated, orthogonally repressible hybrid T7-lac promoters for modular, tuneable control of metabolic pathway fluxes in Escherichia coli. Nucleic Acids Research. 44 (9), 4472-4485 (2016).
- Zhao, C., Shu, X., Sun, B. Construction of a gene knockdown system based on catalytically inactive ("dead") Cas9 (dCas9) in Staphylococcus aureus. Applied Environmental Microbiology. 83 (12), (2017).
- Zhao, Y., et al. CRISPR/dCas9-mediated multiplex gene repression in Streptomyces. Biotechnology Journal. , 1800121(2018).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati