
Method Article
Transkripsiyon Faktörlerinin Zorunlu İfadesi Ile İnsan Fibroblastlarının Hemojenik Yeniden Programlanması
Bu Makalede
Erratum Notice
Özet
Bu protokol, insan dermal fibroblastlarında hemojenik bir programın hematopoetik kök ve döl hücreleri üretmek için gata2, GFI1B ve FOS transkripsiyon faktörlerinin zorunlu ekspresyonu ile indüksiyonuna neden olduğunu göstermektedir.
Özet
İnsan hematopoetik kök hücrelerinin (HSC) özelliklerinin altında yatan hücresel ve moleküler mekanizmalar hala zor dur. İnsan ın HSC'nin in vitro olarak ortaya çıkışLarını özetleme stratejileri, bu karmaşık gelişimsel sürecin incelenmesindeki sınırlamaların üstesinden gelmek için gereklidir. Burada, doğrudan hücre yeniden programlama yaklaşımı kullanan insan dermal fibroblastlarından hematopoetik kök ve ata benzeri hücreler üreten bir protokol uyguluyoruz. Bu hücreler, HSC belirtiminin endotel-to-hematopoetik geçiş (EHT) özelliğini andıran hemojenik ara hücre tipinden geçerler. Fibroblastlar GATA2, GFI1B ve FOS transkripsiyon faktörleri ile transdüksiyon yoluyla hemojenik hücrelere yeniden programlandı. Morfolojik değişiklikler, hemojenik ve hematopoetik belirteçlerin ekspresyonu ve dinamik EHT transkripsiyonel programlarının bu kombinasyonu. Yeniden programlanan hücreler hematopoetik soyundan gelir ve immünofif fareleri üç ay boyunca yeniden çoğaltır. Bu protokol, yeniden programlamanın ilk aşamalarında GATA2 hedeflerini tanımlayarak insan EHT sürecinin mekanistik diseksiyonuna uyarlanabilir. Böylece, insan hemojenik yeniden programlama yeni belirteçleri ve insan HSC ortaya çıkması düzenleyicileri tanımlamak için basit ve çekici bir yaklaşım sağlar. Gelecekte, fibroblastlarda hemojenik kaderin sadık indüksiyonu transplantasyon için hastaya özgü HSC'lerin oluşmasına yol açabilir.
Giriş
Kesin hematopoetik kök ve döl hücreleri (HSPCs) aort-gonad-mesonephros (AGM) bölgesinde ortaya çıkar ve hemojen kapasiteye sahip endotel öncüllerinden plasenta, endotel-to-hematopoetik geçiş yoluyla (EHT)1, 2 . Hemojenik öncüller (HP'ler) hem endotel hem de hematopoetik belirteçleri ifade eder, ancak kesin tanımlamaları özellikle insan sisteminde zor durabilir. Memelilerde nispeten korunmuş bir süreç olmasına rağmen, hematopoetik kök hücre (HSC) gelişimi hala insanlar ve fare modelleri arasında önemli ölçüde farklılık gösterir3,4. Bu nedenle, insan HSC gelişimini özetlemek için in vitro yaklaşımlar gereklidir.
Pluripotent kök hücrelerin (PsC' lerin) HSC'lere farklılaşması, umut verici olsa da, son 20 yılda, çoğunlukla mevcut farklılaşma protokolleri nedeniyle sınırlı bir başarıya ulaşmış ve bu da kötü engreftleme ile ilkel hematopoetik atalara yol açmıştır. yetenek5,6,7. Alternatif olarak, transkripsiyon faktörleri (TFs)8,9kullanılarak, birden fazla hücre türünden HSPC benzeri hücreler oluşturmak için doğrudan hücre yeniden programlama metodolojileri uygulanmıştır. Özellikle, üç TFs, Gata2, Gfi1b ve cFos aşırı ekspresyonu, tanımlanmış bir fenotip (Prom1 +Sca-1+CD34+CD45-)10ile hp ara yoluyla HSPCs içine fare embriyonik fibroblastlar dönüştürülmüş . Bu süreç kesin hematopoezis belirtimi sırasında, embriyo ve plasenta oluşur EHT benzer. Bu fenotip, fare plasentasındaki bir popülasyonun tanımlanmasını ve izole edilmesine olanak sağladı, kısa süreli kültür ve Çentik aktivasyonundan sonra seri olarak nakledilebilir HSC'ler11.
Şimdiye kadar, hiçbir fenotip onların öncülleri insan HSC'ler ayıran kurulmuştur, ancak bazı moleküller integrin alfa 6 (ITGA6 veya CD49f) yüksek uzun vadeli yeniden hscs, en olgunlaşmamış ifade olduğu bilinmektedir HSC bölmesi hücreleri12, ve anjiyotensin dönüştürücü enzim (ACE veya CD143) embriyonik kan oluşturan dokularda CD34 negatif hematopoetik öncüleri mevcuttur13.
Son zamanlarda, biz göstermiştir ki üç TFs, GATA2, FOS ve GFI1B insan versiyonu insan dermal fibroblastlar yeniden program (HDFs) kısa vadeli engraftment kapasitesi ile HPs içine14. Yeniden programlamanın ilk aşamalarında, GATA2 açık kromatin ile uğraşır ve GFI1B ve FOS'u fibroblast genlerini bastırmak ve endotel ve hematopoetik genleri aktive etmek için işe alır. İndüklenen hücreler yüksek oranda CD49f ve ACE ifade ve HSPC marker CD34 ifade hücrelerinin küçük bir yüzdesi içeriyordu. HSC15'te ifade edilen ve HSC16için önemli olan CD9 geninin GATA2'nin doğrudan hedefi olduğu ve yeniden programlanmış hücrelerde en yüksek regüle edilmiş genler arasında olduğu gösterilmiştir14. CD9 bu nedenle insan kesin hematopoiesis HPs için ek bir belirteç teşkil edebilir.
Bu protokolde, gata2, GFI1B ve FOS'un zorunlu ekspresyonu ile insan fibroblastlarından HSPC benzeri hücrelerin oluşumunu ve kromatin immünopresidibatan (ChIP)-sektinite (seq) analizinin başlangıcında uyarlanmış bir yöntemi tanımladık. yeniden programlama. TF'ler tetrasiklin yanıt elemanı (TRE) ve minimal CMV promotörü içeren bir doksisiklin (DOX)-indüklenebilir lentiviral vektör (pFUW-tetO) kodlanmış ve ters tetrasiklin içeren bir kurucu vektör ile birlikte transe transaktivatör proteini (pFUW-M2rtTA). Transdüksiyondan sonra DOX (tetrasiklin analogu) eklendiğinde, TF transkripsiyonuna (Tet-On sistemi) izin veren TRE ile etkileşime giren rtTA proteinine bağlanır. Prosedürün tamamlanması için 25 gün gerekiyor. ChIP-seq deneyleri için HDF'ler GATA2 (pFUW-tetO-3xFLAG-GATA2) ve GFI1B (pLV-tetO-HA-GFI1B) etiketli versiyonları ile, ayrıca pFUW-tetO-FOS ve TF bağlama siteleri DOX takviyesinden iki gün sonra analiz edildi.
Sonuç olarak, insan fibroblastlarının hemojenik yeniden programlanması, insan gelişimsel hematopoyonunun altında yatan mekanizmaları incelemek için in vitro traktörsistemi ve gelecekteki klinik uygulama için hastaya özgü HSPC'lerin potansiyel kaynağını sağlar.
Protokol
Bu protokol Lund Üniversitesi'nin insan araştırma etik komitesi yönergelerine uygun olarak gerçekleştirilmiştir ve bireysel kurumsal yönergelere uygun olarak yapılmalıdır.
1. Reaktif Hazırlama
- Dulbecco'nun modifiye Eagle's medium (DMEM)/%20 fetal sığır serumu (FBS) için sodyum pirüuvat içeren yüksek glikozlu DMEM %20 FBS, %1 penisilin-streptomisin (kalem/strep), %1 L-glutamin, %1 esansiyel olmayan amino asitler ve 10-4-M 2- mercaptoethanol.
- Komple DMEM için sodyum pirüuvat içeren yüksek glikozlu DMEM 'i %10 FBS, %1 Kalem/strep ve %1 L-glutamin ile karıştırın.
- Hematopoetik ortam için hematopoetik ortamı(Malzeme Tablosu)ile 10-6 M hidrokortizon ve %1 kalem/strep ile karıştırın.
- Kalsiyum veya magnezyum olmadan fosfat tamponlu salin (PBS) kullanın.
2. İnsan Dermal Fibroblast İzolasyon
NOT: HDF'ler sertifikalı tedarikçilerden(Malzeme Tablosu)satın alınabilir. Bu durumda, fibroblastları genişletin ve doğrudan yeniden programlama deneylerinde kullanın (bölüm 4). Alternatif olarak, HDF'ler bağışçılardan izole edilebilir. Fibroblastlar farklı donörlerden izole edilmişse, protokolün her adımında numuneleri birbirinden ayrı tutun. Her donörün kimlik numarası ile etiket plakaları/kuyuları ve toplama tüpleri.
- Uzman hekimler tarafından yapılan 3 mm yuvarlak deri yumruk biyopsilerinden HDF elde edin.
- 500 μL %0,1 jelatin ve 37 °C'de 20 dakika kuluçka ya da kuluçka ya da 37 °C'de kuluçkaya yatan, doku kültürü yle tedavi edilen 6 kuyuluk üç kuyuyu kaplayın.
- Kalan jelatin çözeltisini aspire edin ve her kuyuya 750 μL DMEM/%20 FBS ekleyin. Kuyunun tüm yüzeyi orta ile kaplı olmalıdır.
- Steril 100 mm petri kabı kapağının iç yüzeyine 1,5 mL DMEM/%20 FBS ekleyin ve 5 mL serolojik pipet yardımıyla damlayı yayın.
- Sterilize forceps ile kapak üzerinde orta cilt biyopsisi yerleştirin.
- Deri biyopsisini dokuz özdeş bölüme ayırın, biyopsiyi yerinde tutmak için sterilize edilmiş neşter ve kesilmesi gereken ikinci bir neşter kullanarak.
- Sivri uçlu ponponlar kullanarak her kuyuya üç biyopsi parçası yerleştirin. Parçaların kuyunun dibine iliştirdiğinden emin olun.
- Parçaların üzerine 22 mm'lik bir kapak kaydırın ve biraz basınç uygulayın.
- Bir hafta boyunca 37 °C, %5 CO2'deplakayı kuluçkaya yatırın. Hücreleri günlük olarak kontrol edin ve buharlaşan ortamı değiştirmek için her 2 günde bir 200 μL DMEM/20% FBS ekleyin.
- Bir hafta sonra, 2 mL'ye kadar DMEM/20% FBS ekleyin ve orta ortamı 2−3 günde bir değiştirin.
-
Kuyular konca olduğunda (yaklaşık 4−8 hafta) 1:4 oranında geçiş hücreleri.
- % 0.1 jelatin kaplı doku kültürü tedavi 6-well plakalar hazırlayın.
- Kuyulardan %80 birleşme ile aspirasyon ortamı ve 1 mL PBS ile bir kez yıkayın.
- Steril çalgılarla coverslip'i çıkarın ve kapak kapağını doku tarafı yukarı olan 6 kuyuluk yeni bir kuyuya yerleştirin.
NOT: Kapak kaymasına bağlı kalan hücreler de hasat edilecektir. - Kuyu başına 500 μL ayrışma çözeltisi(Malzeme Tablosu)ekleyin (kapaklı kuyular dahil) ve 37 °C'de kuluçkaya yatırın, 5−10 dk için %5 CO2. her kuyuya 500 μL DMEM/%20 FBS ek.
- 15 mL konik tüp içine tüm kuyulardan fibroblastlar toplayın. Kalan hücreleri toplamak için kuyulara ekstra orta ekleyin. 5 dk için 350 x g tüp santrifüj.
- Bu arada, daha önce jelatin kaplı plakaların her kuyuya 500 μL DMEM/%20 FBS ekleyin.
- 6 mL DMEM/%20 FBS'de aspire orta ve resuspend fibroblastlar.
- Her kuyuya 500 μL fibroblast süspansiyon ekleyin (numune/donör başına toplam iki adet 6 kuyulu plaka). 37 °C, %5 CO2'debir gecede inkübasyon hücreleri .
- Ertesi gün, her kuyuya 1 mL DMEM/%20 FBS ekleyin. Kuyular %80 konakıcı olana kadar orta ortamı 2 mL DMEM/20% FBS ile 2−3 günde bir değiştirin.
- Üçüncü bölüme ulaşılına kadar üç confluent kuyu için bölüm 2.11'i tekrarlayın.
-
Konfluent kuyulardan fibroblastları dondurun (pasajlar 1 ve 3).
- Kuyulardan aspirasyon ortamı ve 1 mL PBS ile bir kez yıkayın.
- 2.11.4 ve 2.11.5 adımlarında açıklandığı gibi fibroblastları ayırın ve toplayın.
- Bir hemositometre ile hücreleri saymak ve 5 dakika için 350 x g tüp santrifüj.
- Santrifüj den sonra FBS'de 5 x 105 hücre/mL yoğunlukta %10 DMSO ile aspirasyon orta sı ve resuspend fibroblastlar.
- Cryovial başına hücre süspansiyon1 mL ekleyin ve dondurucu bir kap kullanarak -80 °C gecede hücreleri dondurma. Uzun süreli depolama için şişeleri -150 °C'ye (sıvı nitrojen) taşıyın.
3. Lentiviral Üretim
- HEK293T hücrelerini 100 mm doku kültürü yle işlenmiş tam DMEM 10 mL'lik birçanakta, 37 °C,%5 CO 2'de, biraraya ulaşınana kadar yetiştirin.
- Transfeksiyondan önceki gün, orta aspirasyon ve 5 mL PBS ile yemeği dikkatlice yıkayın.
- PBS'i çıkardıktan sonra, hücreleri yemekten çıkarmak için 1,5 mL ayrışma çözeltisi ekleyin ve 37 °C'de %5 CO2, 5−10 dk için %5 CO 2'ye inkübül.
NOT: Kullanmadan önce hem PBS hem de ayrışma çözeltisini ısıtmak için önerilir, böylece hücreler termal şoka uğramaz. - 3 mL tam DMEM ile dissosilasyon çözeltisini inaktive edin ve hücre süspansiyonunu 15 mL konik tüpe aktarın. Kalan bağlı hücreleri çıkarmak için 5 mL tam DMEM ile bulaşıkyıyıyı yıkayın ve bu hacmi 15 mL konik tüpe aktarın.
- Santrifüj hücre süspansiyonu 350 x g 5 dk.
- Aspire süpernatant ve çanak başına tam DMEM 10 mL son hacminde altı 100 mm doku kültürü tedavi yemekleri arasında eşit hücre pelet bölünmüş. Hücreler transfeksiyon zamanına kadar yaklaşık %60 oranında konfluent olmalıdır.
- Ertesi gün, plazmid karışımları ile transfect hücreleri aşağıdaki gibi:
NOT: Protokolün bu bölümünde plazmid karışımı başına 100 mm doku kültürü ile tedavi edilen bir çanak içinde lentivirüslerin üretimi açıklanmaktadır. Konsantrasyon için daha yüksek hacimlerde lentiviral supernatant elde etmek için karışım başına en az dört adet 100 mm HEK293T hücre kültürü yemeği kullanın.- 15 mL konik bir tüpte, üç transfer plazmidinin 10 μg'sini birlikte ekleyin: 3,33 μg pFUW-tetO-GATA2 (Addgene plazmid #125028)14, 3,33 μg pFUW-tetO-GFI1B (Addgene #125597)14 ve 3,33 μg pFUW-tetO-FOS (Addgene) (Add #125598gene14) 10 μg2 nesil psPAX2 ambalaj vektörü Gagkodlama , Pol, Tat ve Rev genleri (Addgene #12260) ve pMD2.G zarf vektör vsv-G gen kodlama 5 μg (Addgene #12259). 500 μL'ye kadar su ekleyin.
- İki yeni 15 mL konik tüpler de 10 μg FUW-M2rtTA plazmid (Addgene #20342)17, 10 g psPAX2 ambalaj vektörü ve 5 g pMD2.G zarf vektörü her tüpe ekleyin. 500 μL'ye kadar su ekleyin. Bir tüp kontrol olarak kullanılacak.
- Her tüp için 2 M CaCl262,5 μL ekleyin. Daha sonra, bir pipet denetleyicisi içine yerleştirilen bir Pasteur pipet kullanarak her karışımiçine kabarcıklar bırakın. Kabarcıklar oluşurken, pipet 500 μL N,N-bis(2-hidroksitil)-2-aminoetanesülfonik asit (BES) tamponlu salin (pH 7.1, 25 °C), P1000 pipet ile, Pasteur pipet karşı damla ve karışım üzerine.
- En az 15 dakika oda sıcaklığında tüpler inkübasyon. Karışımları bir süre sonra biraz bulutlu görünecektir.
- Bu arada, HEK293T hücre yemeklerinden aspire ortamı (bir gün önce pasajlı) ve antibiyotiksiz tam DMEM 10 mL ekleyin. Dikkatli olun ve HEK293T hücreleri yarı yapışık olduğu için orta yavaş yavaş ekleyin.
- Her bir karışımı (yaklaşık 1 mL) eşit ve damla akıllıca ayrı yemeklere dağıtın ve bir gecede 37 °C, %5 CO2'dekuluçkaya yatırın.
- Orta yı 4 mL tam DMEM, kuluçkadan 24 saat sonra değiştirin. Bir gecede 37 °C,% 5 CO2kuluçka . Varsa, 32 °C, %5 CO2'dekuluçkaya yatırın, çünkü azalan sıcaklık lentiviral partiküllerin yarı ömrünü uzatır.
- 50 mL konik tüp için üç kez lentiviral parçacıklar ile supernatant toplamak. Bu noktada farklı lentiviral parçacıkları karıştırmayın. Her çanak lentiviral supernatant 12 mL neden olacaktır. Aynı viral preparatdört tabak bir 50 mL konik tüp sığar.
DİkKAT: Lentiviral iş için özel bir laminar akış kaputunda biyogüvenlik seviyesi-2 laboratuarında lentiviral toplama gerçekleştirin ve biyolojik tehlikeli maddeler için uygun bir kapta viral kontamine atık (tüpler, ipuçları, yemekler) yerleştirin.- Son kuluçkadan sonra ilk koleksiyonu 16 saat yapın ve 4 mL tam DMEM ekleyin. 37 °C,% 5 CO2inkübat.
- İkinci koleksiyonu aynı tüpe ilk indikten 8 saat sonra yapın, 4 mL tam DMEM ekleyin ve 37 °C,%5 CO2'dekuluçkaya yatırın.
- Aynı tüp için ikinci sonra üçüncü toplama 16 saat yapın ve bulaşıkları atın.
NOT: Her koleksiyondan sonra lentiviral supernatants'ı 4 °C'de saklayın.
- Her lentiviral supernatant'ı 0,45 m düşük proteinli bağlama filtresi ile selüloz asetat membranı(Malzeme Tablosu)ile temiz bir tüpe süzün.
- Rejenere selüloz membranı(Malzeme Tablosu)bulunan bir santrifüj filtre ünitesine maksimum 15 mL filtrelenmiş süpernatant ekleyin ve 4 °C'de 25 dakika boyunca 4.000 x g'da döndürün. Akış tan atın. Lentivirüsler içeren viskoz bir sıvı filtre ünitesinde kalır.
- Daha fazla lentiviral supernatant kalana kadar, filtre ünitesinin üstüne 15 mL supernatant ekleyerek adım 3.13 tekrarlayın.
NOT: Konsantre olmak için süpernatant sadece birkaç mililitre olduğunda, 10 dakika iplik süresini azaltmak. Filtrede hala ekstra sıvı (viskoz olmayan) varsa, ek bir 10 dakika santrifüj. - Her bir konsantre lentivirüs türünün aliquots (50−200 μL) yapmak ve uzun süreli depolama için -80 °C 'de (1−2 yıl) veya 4 °C'de kısa süreli depolama için (1−2 hafta) saklayın.
NOT: Konsantre veya konsantre olmayan lentivirüsler de taze kullanılabilir. Bu azaltılmış titre ile sonuçlanır gibi yeniden dondurma ve çözülme etmeyin.
4. Hemojenik Yeniden Programlama
NOT: Yeniden programlama denemeleri gerçekleştirmek için üç (P3) veya daha yüksek (P10'a kadar) geçiş sayısına sahip HDF'ler kullanın.
- 100 mm doku kültürü yle işlenmiş bir yemeği %0,1 jelatin 5 mL ve 37 °C'de 20 dk. Kalan jelatin çözeltisini aspire edin.
- % 0.1 jelatin kaplı çanak bir fibroblast şişe ve plaka hücreleri çözülme. Bir gecede 37 °C,% 5 CO2kuluçka . Gerekirse, istenilen geçit ve kesişme ulaşılana kadar daha uzun bir süre için fibroblastlar genişletin.
- Coat 6-iyi doku kültürü tedavi plaka ile 500 μL 0.1% jelatin çözeltisi ve kuluçka 37 °C 20 dk. Ekstra jelatin çıkarın.
- Plaka HDF'leri, her kuyuda 2 mL tam DMEM'de plaka başına 150.000 hücre (kuyu başına 25.000 hücre) yoğunluğunda dır. Hücre eki sağlamak için 37 °C, %5 CO2'degece boyunca kuluçkaya yatırın.
- Orta ortamı 2 mL tam DMEM artı 8 g/mL polibrene değiştirin. Yeni bir mikrosantrifüj tüp havuz üretilen TF lentiviruses ve M2rtTA bir 1:1 oran karışımı hazırlayın.
NOT: Bu protokolde, üç TF için lentiviruses havuz üretimi gerçekleştirilir, hangi, yazarların elinde, daha yüksek yeniden programlama verimliliği ile sonuçlanır. Alternatif olarak, standart bir hücre hattı üzerinde qPCR18tarafından bireysel lentiviral parçacıkların bir titrasyon gerçekleştirmek için önerilmektedir. Bu, eş-transdüksiyon ve hemojenik yeniden programlama için en uygun enfeksiyon (MOI) çokluğunu karşılamak için gerekli olan tek tek virüslerin hacmini tanımlamak için kullanılacaktır. - HDF'leri dağıtmak için kuyu başına 10 ila 100 μL mercek karışımı dağıtın. Bu gün -2 yeniden programlama.
NOT: Hücre canlılığından ödün vermeden verimli yeniden programlama için lentiviral karışımın en uygun hacminin tanımlanması en iyi duruma getirilmesi gerektirir (daha fazla ayrıntı için Ek Şekil 1'e bakınız). 7'den fazla geçişi olan HDF'ler, daha düşük geçişli hücrelere göre daha yüksek virüs hacimleri gerektirebilir. - Kuluçka 16 saat sonra, virüsleri kaldırmak ve tam DMEM ekleyin. Hücrelerin 6−8 saat boyunca toparlanmasına izin verin.
- İyileştikten sonra orta aspire edin ve 8 g/mL polibrene sahip 2 mL tam DMEM ekleyin.
- Adım 4.6'da açıklandığı gibi ikinci bir transdüksiyon yapın ve 37 °C'de kuluçkaya yatırın, 16 saat için %5 CO2. Bu gün -1 yeniden programlama. Lentiviral karışım her iki transdüksiyon için -2 gün hazırlanabilir ve 4 °C'de tutulabilir.
- Ertesi gün, virüsleri çıkarın ve 1 μg/mL DOX ile takviye tam DMEM ekleyin. Bu yeniden programlama gün 0. 37 °C'de kuluçka, 48 saat için %5 CO2.
-
Yeniden programlamanın 2.
- 1 mL PBS ile aspire ortamı ve yıkama hücreleri.
- PBS'yi aspire edin ve 500 μL ayrışma çözeltisi ile hücreleri ayırın. Kuluçka 5−10 dk 37 °C, %5 CO2.
- 1 mL tam DMEM ile dissosilasyon çözeltisini inaktive edin ve hücreleri konik bir tüpe toplayın. Santrifüj 350 x g 5 dk.
- Peletin hematopoetik ortamda yeniden askıya alınması (bkz. adım 1.3), 1 μg/mL DOX ile desteklenmiş, ve plaka hücreleri yeni doku kültürü yle tedavi edilmiş 6-iyi plakalara % 0,1 jelatin le kaplanmış ve kuyu başına 2 mL'lik son bir hacimde.
- Yeniden programlama kültürleri (25 gün) süresince haftada iki kez orta (hematopoetik ortam artı DOX) değiştirin.
- Brightfield veya floresan mikroskobu ile farklı zaman noktalarında yeniden programlanan hücreleri analiz etmek (Bkz. Ek Şekil 2),akış sitometrisi, dökme ve tek hücreli RNA dizilimi ve transplantasyon tahlilleri hematopoetik morfolojisi, endotel ve hematopoetik belirteçlerin varlığı, endotel/hematopoetik gen ekspresyonu profili ve rejenerasyon kapasitesi14.
5. Hemojenik Reprogramming Başlangıcında ChIP-seq Analizi için Fibroblast Genişlemesinin Optimizasyonu
- Plaka 300.000 HDFs (2kuluçka .
- Ertesi gün orta ortamı 8 g/mL polibren ile tamamlanmış tam DMEM ile değiştirin.
- Hücreleri tek tek faktörlerle aktarın: pFUW-tetO-FOS14, pLV-tetO-HA-GFI1B (Addgene #125599)14 ve pFUW-tetO-3xFLAG-GATA2 (Addgene #125600)14 veya üç faktörden oluşan bir havuz, artı FUW-M2rtTA 1:1 oranında. 10−20 μL toplam virüs (bireysel TF + M2rtTA veya üç TFs + M2rtTA) kullanın. 37 °C, %5 CO2'debir gecede inkübasyon hücreleri .
NOT: Her koşulda on iki adet 6-iyi plaka kullanılması tavsiye edilir (her tf ve üç TF kombine için). - Lentivirüsleri çıkarın ve ilk transdüksiyondan sonra tam DMEM 16 saat ekleyin. Hücrelerin 6−8 saat boyunca iyileşmesine izin verin.
- Hücreleri ikinci kez aynı miktarda virüs le koşul başına ve 37 °C,% 5 CO2'dekuluçkaya yatırın.
- Ertesi gün virüsleri kaldırın ve tam DMEM ekleyin. 37 °C'de kuluçka, 24 saat için %5 CO2.
- Her kuyuyu %0,1 jelatin kaplı doku kültürü yle işlenmiş 100 mm'lik bir tabağa tam DMEM ile çanak başına 10 mL'lik son bir hacme yeniden plakalayın. Bu yaklaşık 1:6 geçişi temsil eder.
- Hücrelerin 37 °C, %5 CO2'de6 gün boyunca büyümesine izin verin.
- Gün 6 yeniden kaplama sonra, aspire orta ve 1 μg/mL DOX ile tam DMEM ekleyin. 37 °C'de, %5 CO2'de 2 gün boyunca kuluçkaya yatırın.
- Fibroblastlar toplamak ve üç TFs genomik bağlama siteleri ayrı ayrı veya birlikte transduced analiz, ChIP-seq tarafından 2 gün DOX takviyesi sonra14.
NOT: Son yetmiş iki 100 mm çanak 20−50 x 106 hücreleri arasında, ChIP-seq deneyleri ve çoğaltmayapmak için yeterli içerecektir.
Sonuçlar
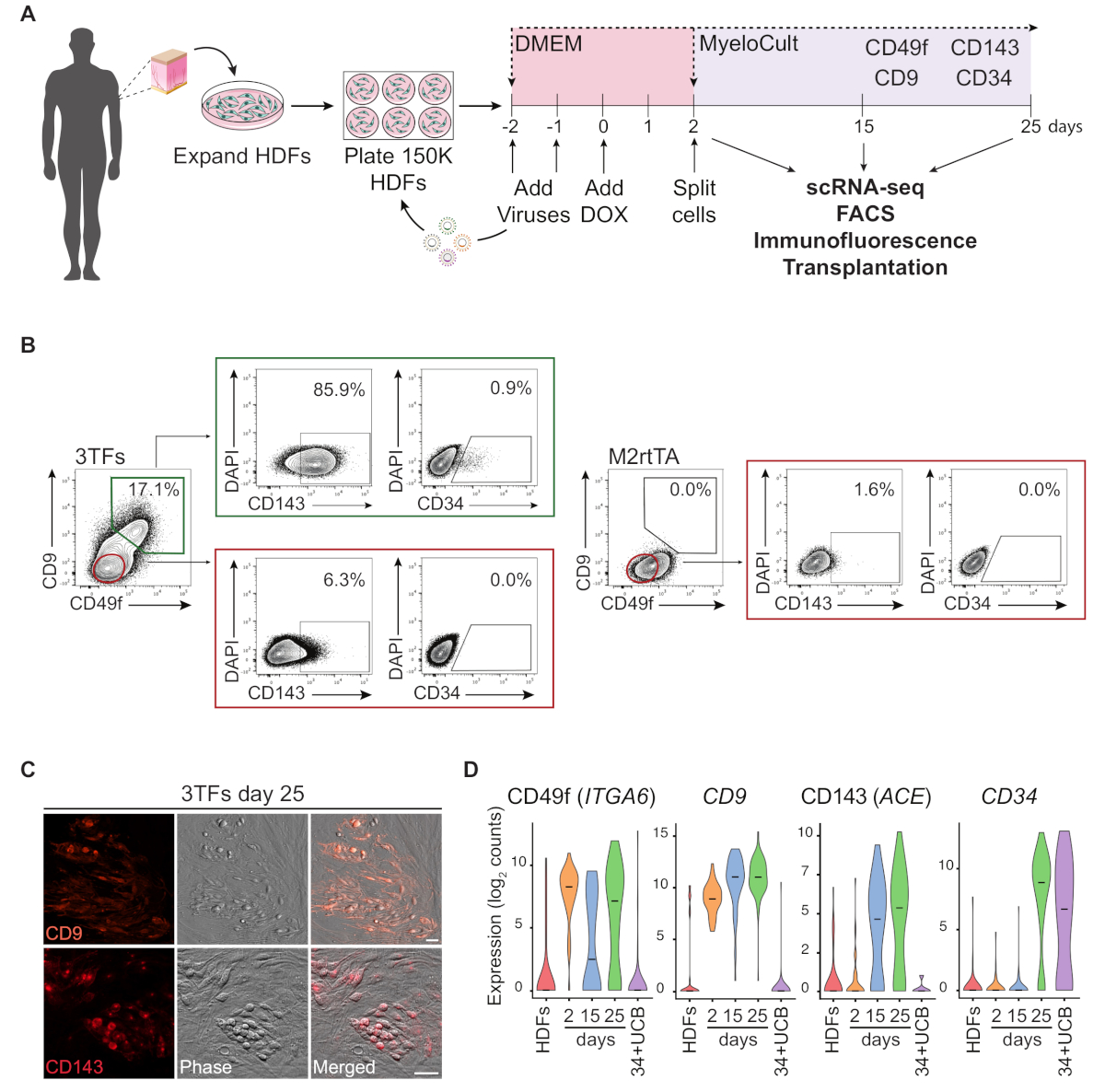
HDF'ler kullanılarak yeniden programlama yaklaşımının şematik bir gösterimi Şekil 1A'dagösterilmiştir. Fibroblastlar ticari kaynaklardan elde edilir veya insan donörlerinden toplanır ve yeniden programlamadan önce in vitro olarak genişletilir. Kaplamadan sonra hücreler GATA2, GFI1B ve FOS (ve M2rtTA) lentivirüsleri ile iki kez transkripsiyonu yapılır ve yeniden programlamanın 0. 2. gün, hücreler bölünür ve kültürün 25. Yeniden programlanmış hücreler, bağışıklık sistemi zayıf farelerde transplantasyon, saflaştırılmış hücre popülasyonlarının tek hücreli RNA-sekesi (scRNA-seq) (2. gün sıralanmamış, gün 15 CD49f+ CD34 ve gün 25 CD49f+CD34+ hücreleri), yanı sıra hücre yüzey belirteçleri CD49f, CD34, CD9 ve CD143 için mikroskopi ve akış sitometri analizi. Temsili sitometri çizimleri, 25 günlük yeniden programlamadan sonra cd49f ve CD9(Şekil 1B, sol panel)ifade eden yeniden programlanmış hücrelerin ~%17'sini gösterir. Çift pozitif hücrelerin çoğunluğu CD143 (~86%), ve küçük bir popülasyon ifade CD34 (%0.9), dinamik bir hemojenik kader indüksiyon düşündürmektedir. Bu belirteçler M2rtTA transe hdf'lerde 25 gün boyunca(Şekil 1B, sağ panel)etkinleştirilmez. İmmünofloresans görüntüleri, bu belirteçler için negatif olan fibroblastlardan morfolojik olarak farklı olan yapışık ve yuvarlak hücrelerde CD9 ve CD143 ekspresyonunu doğrular (Şekil 1C). İnsan hemojenik koloniler de IFADE CD49f ve CD3414. HDF'lerin ScRNA-seq analizi, 2. CD49f ve CD9 pozitif hücreler ilk olarak yeniden programlama işlemi sırasında, 2 ve 15. CD143 ifadesi 15. CD34+ göbek kordon kanı (UCB) hücreleri referans olarak kullanılmıştır (Şekil 1D).
Şekil 2A, hemojenik yeniden programlamanın ilk aşamalarında ChIP-seq analizi için yeterli sayıda hücre oluşturmak için değiştirilmiş bir protokolü açıklar (gün 2). İlk olarak, HDF'ler standart protokolden iki kat daha yüksek bir yoğunlukta kaplanır (300.000 hücre, plaka başına 150.000 hücre). Transdüksiyondan sonra, her kuyu 100 mm'lik bir tabağa dönüştürülerek hücrelerin DOX ile orta maddetakviyesinden önce 6 gün boyunca genişlemesini sağlar. Hücreler DOX ve buna ek olarak TF ifadesi ekledikten 2 gün sonra analiz edilir. Şekil 2B, hücreler üç faktör (3TF) veya GATA2 ile tek tek birlikte transetik olduğunda, GATA2'nin ITGA6 ve ACE genomik düzenleyici bölgelerine bağlanan genom tarayıcı profillerini göstermektedir. GATA2 ayrıca CD9 ve CD34 genlerinin kromatin bölgelerini açmaya bağlanır14.

Şekil 1: İnsan dermal fibroblastlarında hemojenik kaderin indüksiyonu. (A) İnsan dermal fibroblastlarının (HDF) hemojenik yeniden programlaması için deneysel strateji. Deri yumruk biyopsilerinden elde edilen fibroblastlar donörlerden toplanır, GATA2, GFI1B, FOS ve M2rtTA lentivirüsleri ile genişletilir ve aktarılır. Doksisiklin (DOX) yeniden programlama gün 0 kültüre eklenir ve hücreler gün 25 kadar çeşitli zaman noktalarında analiz edilir. scRNA-seq, tek hücreli RNA-sıralama. FACS, Floresan-Aktif Hücre Sıralama. (B) Transdüksiyon sonrası transdüksiyon dan sonra 25. Sitometri çizimleri CD49f ve CD9 için çift pozitif hücrelerin yüzdesini gösterir, canlı hücre popülasyonunda (DAPI-negatif) geçitli. Çift pozitif popülasyon içinde CD143 ve CD34 ifadesi gösterilir. Aynı kültür koşullarında sadece M2rtTA virüsü ile transet edilen HDF'ler kontrol olarak kullanılmaktadır. (C) Cd9 (üst panel) ve CD143 (alt panel) ekspresyonunu doğrulayan 25. Hücreler antikorlarla boyandı(Malzeme Tablosu)PBS/2% FBS'de fare serumu ile 1:100 seyreltildi, 37 °C'de 20 dk, %5 CO2,üç kez yıkandı ve PBS/2% FBS'de görüntülendi. Faz, faz-gradyan kontrast. Ölçek çubukları = 50 μm. (D) Farklı zaman noktalarında 253 hücrenin ScRNA-seq analizi. ITGA6, CD9, ACE ve CD34 ifadesi yeniden programlama sırasında etkinleştirilir. Hücreler 2. gün (sıralanmamış), 15. gün (CD49f+CD34-) ve25. HDF'ler ve CD34+ göbek kordon kanı (34+UCB) hücreleri referans olarak kullanılır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: ChIP-seq analizi için insan dermal fibroblastlarının genişlemesi. (A) ChIP-seq için yeniden programlamanın 2. 300.000 hücre 6-iyi plakalar içine kaplanır ve bireysel faktörler (pFUW-tetO-FOS, pLV-tetO-HA-GFI1B veya pFUW-tetO-3xFLAG-GATA2) veya üç faktörün (artı M2rtTA) bir kombinasyonu ile iki kez transe. Virüsler ilerlerken fibroblastlar 100 mm'lik tabaklarda altı gün boyunca genişletilir. Doksisiklin (DOX) gün 0 eklenir ve hücreler DOX eklenmesinden iki gün sonra toplanır. (B) Itga6 ve ACE loci'de GATA2 bağlayıcı bölgeleri (gri kutular) vurgulayan genom tarayıcı profilleri transdüksiyondan iki gün sonra üç transkripsiyon faktörü (3TF) veya sadece GATA2 ile. Eşlenen okumaların toplam sayısı y ekseninde temsil edilir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Ek Şekil 1: Verimli hemojenik yeniden programlama için optimize edilmiş bir lentiviral hacmin tanımlanması. İnsan dermal fibroblastlarını (HDF' ler) ve M2rtTA ile protokolün 4.5−4.12. Yeniden programlanan hücreler, canlı hücrelerde (DAPI-negatif) kaplanmış CD49f+CD9+ hücrelerin yüzdesi ile verilen hemojenik yeniden programlama için en uygun transdüksiyon hacmini tanımlamak için 25. Hücre canlılığı, 25. M2rtTA (100 μL) ile transetlenen HDF'ler negatif kontrol olarak kullanılır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Ek Şekil 2: İnsan dermal fibroblastlarının hemojenik yeniden programlaması sırasında morfoloji değişiklikleri. İnsan Dermal Fibroblast (HDF) kültürleri ilk transdüksiyon gününde (gün -2), DOX kültürlere eklendiğinde (gün 0), iki gün (gün 2) ve on beş gün (gün 15) DOX takviyesi sonra ve deneyin bitiş noktasında (gün 25). 15 ve 25. Ölçek çubukları = 100 μm. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
Bu makalede, bir yöntem doğrudan insan fibroblastlar hematopoetik progenitor hücreleri oluşturmak için açıklanmıştır, hangi bir HP hücre ara geçmesi, kesin HSCs benzer14.
GATA2, GFI1B ve FOS'u kodlayan merceksi partiküllerin havuz üretimi bireysel üretime göre tercih edilmiştir, çünkü bizim elimizde daha yüksek yeniden programlama verimliliği (yayınlanmamış veriler) ile sonuçlanır. Lentiviruses, Retroviridae ailesinin üyeleri olarak, normalde pozitif tek iplikçikli RNA19iki kopya içerir. Yeniden programlama veriminin artması, aynı lentiviral parçacıkta iki farklı transgenin paketlenmesinden kaynaklanabilir ve bu da üç transkripsiyon faktörüyle birlikte transeneden hücre sayısının artmasına neden olabilir. Bu protokolün başarısını sağlamak için, 4.6 adımda tavsiye edilen yeniden programlama verimliliği ve hücre canlılığı arasında en iyi dengeyi elde etmek için hücre geçişine bağlı olarak yeterli miktarda virüs içeren HDF'ler ile aktarılması gerekmektedir. Ayrıca, taze konsantre olmayan virüsler kullanılabilir. 0.5-3 mL 3TFs havuzu ve M2rtTA ile hücrelerin aktarılması tavsiye edilir. Ayrıca, hücre yoğunluğu uygulamaya göre ayarlanmalıdır. 6 kuyulu plaka başına 150.000 HDF (adım 4.4) facs yapmak için en uygun yoğunluğu sağladı, transplantasyon ve yeniden programlanmış hücrelerin akış sitometri analizi. ChIP-seq deneyleri için başlangıçtan itibaren daha fazla hücre ye ihtiyaç duyulur (adım 5.1). İndüklenen hematopoetik hücrelerin ortaya çıkışını desteklemek için hücreleri morfolojik değişiklikler için düzenli olarak kontrol etmek ve haftada iki kez hematopoetik ortamıdeğiştirmek önemlidir. Besleyici katmanlarda hematopoetik sitokinlerin veya ortak kültürün eklenmesi yeniden programlama verimliliğini artırabilir.
Bu yöntemle hemojenik yeniden programlama sırasında dinamik olarak ifade edilen yeni hematopoetik belirteçleri tanımlayabiliriz. Transkripsiyonel düzey14'teyeniden programlanmış hücrelerde yukarı-regüle edildiği gösterilen CD9, insan HSC öncüllerinin yeni bir belirteci olarak hizmet veren CD49f ve CD143 ile birlikte yeniden programlamanın ilk aşamalarında hücre yüzeyinde hızla ifade edilir. Ayrıca ITGA6 ve ACE'nin hemojenik yeniden programlamanın ilk aşamalarında GATA2'nin doğrudan hedefi olduğunu, CD9 ve CD3414'eek olarak insan hemojenik arasında doğrudan mekanik bir bağlantı sağladığını da gösteriyoruz. öncüfenot ve GATA2.
Bu sistemin bir avantajı nispeten homojen fibroblast kültürlerin kullanımında bulunur. PsCs kolayca genişletilmiş ve in vitrokorunur iken, farklılaşma protokolleri hematopoetik ataları içeren heterojen popülasyonlar oluşturmak, hangi kötü5engraft 5 ,6,7. Ayrıca, PSC kaynaklı HSPC'ler nakledilirken tümörigenez riski vardır, çünkü farklılaşmamış PSC'ler farklılaşma protokolleri uyguladıktan sonra bile kültürde kalmaya devam edebilirler. Fibroblastlara alternatif olarak, HSC'lere doğrudan yeniden programlama kana bağlı atalara20 ve endotel hücrelerine21uygulanmıştır. Ancak, kan kısıtlı ata hücreleri ile başlayan hasta kök / atajemihematopoetik popülasyon etkileyen mutasyonlar taşırsa ortaya çıkan HSCs terapötik uygulama engeller22. Endotel hücreleri söz konusu olduğunda, fibroblastlara göre elde edilmesi daha zordur ve organa bağımlı olan fenotip, fonksiyon ve yapı açısından çok heterojen hücre popülasyonunu oluşturmaktadır23. Diğer çalışmalar engraftable hematopoetik ataları içine fare fibroblastlar yeniden programlama başarılı olmuş24,25 henüz, şimdiye kadar, başka bir protokol insan fibroblastlar HSPC benzeri hücrelerin nesil açıklar.
Bu yaklaşım, farmakolojik inhibisyon, gen nakavt veya nakavt izinleri ile birleştiğinde doğrudan insan HSC'leri indüklemek için gerekli olan faktörlerin bireysel veya kombinasyonunu tanımlamak için izin verir. Yeniden programlamadan önce HDF'lerde CRISPR-Cas9 teknolojileri, insan kesin hematopoyonunun yeni düzenleyicilerini tanımlamak için heyecan verici bir çabayı temsil eder. Gelecekte, fibroblastlar gibi kan ait olmayan insan hücre tiplerinin yeniden programlanması, klinik uygulamalar için hastaya özel hematopoetik progenitor hücreleri oluşturmak için bir platform görevi göreceksiniz.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Knut ve Alice Wallenberg vakfı, Lund Üniversitesi ve Bölge Skåne Tıp Fakültesi cömert mali destek için kabul edilmektedir. Bu çalışma Olle Engkvists Stiftelse (194-0694 Filipe Pereira için) ve Fundação para a Ciência e Tecnologia (PTDC/BIM-MED/0075/2014 Filipe Pereira için doktora bursları) bir hibe tarafından desteklenmiştir, ve SFRH/BD/135725/2018 ve SFRH/BD/51968/2012 rita Alves için ve Andreia Gomes). Bu çalışma ayrıca NIH ve NYSTEM (1R01HL119404 ve C32597GG Kateri A. Moore) fonları tarafından desteklenmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 0.45 μm low-protein binding filter, 150 mL Bottle Top Vacuum Filter | Corning | #430625 | |
| 2-Mercaptoethanol | Sigma-Aldrich | #M6250 | |
| Alexa Fluor 488 anti-human CD34 clone 581 | BioLegend | #343518 | |
| BD Pharmingen APC Mouse Anti-Human Angiotensin Converting Enzyme (CD143) clone BB9 | BD Biosciences | #557929 | |
| BES buffered saline | Sigma-Aldrich | #14280 | |
| Calcium chloride (CaCl2) | Sigma-Aldrich | #449709 | |
| Centrifugal filter unit, Amicon Ultra-15 Centrifugal Filter Unit | Sigma-Aldrich | #UFC903096 | |
| Dissociation solution, TrypLE Express Enzyme (1x) no phenol red | Gibco | #12604-021 | |
| Doxycycline hyclate (DOX) | Sigma-Aldrich | #D9891 | |
| eBioscience CD49f (Integrin alpha 6) Monoclonal Antibody (eBioGoH3 (GoH3)), PE-Cyanine7 | Invitrogen | #25-0495-82 | |
| FUW-M2rtTA | Addgene | #20342 | |
| Gelatin from Porcine Skin Type A | Sigma-Aldrich | #G1890 | |
| Gibco L-Glutamine (200 mM) | ThermoFisher Scientific | #25030-024 | |
| Gibco MEM Non-Essential Amino Acids Solution (100x) | ThermoFisher Scientific | #11140-035 | |
| Hematopoietic medium, MyeloCult H5100 | STEMCELL Technologies | #05150 | |
| Hexadimethrine bromide (polybrene) | Sigma-Aldrich | #H9268 | |
| Human Dermal Fibroblasts (HDFs) | ScienCell | #2320 | |
| HyClone Dulbecco's Modified Eagles Medium (DMEM) | GE Healthcare | #SH30243.01 | |
| HyClone Fetal Bovine Serum (FBS) | GE Healthcare | #SV30160.03 | |
| HyClone Penicillin Streptomycin 100x Solution (Pen/Strep) | GE Healthcare | #SV30010 | |
| HyClone Phosphate Buffered Saline solution (PBS) | GE Healthcare | #SH30256.01 | |
| Hydrocortisone | STEMCELL Technologies | #7904 | |
| Mouse serum | Sigma-Aldrich | #M5905 | |
| PE anti-human CD9 Antibody clone HI9a | BioLegend | #312105 | |
| pFUW-tetO-3xFLAG-GATA2 | Addgene | #125600 | |
| pFUW-tetO-FOS | Addgene | #125598 | |
| pFUW-tetO-GATA2 | Addgene | #125028 | |
| pFUW-tetO-GFI1B | Addgene | #125597 | |
| pLV-tetO-HA-GFI1B | Addgene | #125599 | |
| pMD2.G | Addgene | #12259 | |
| psPAX2 | Addgene | #12260 |
Referanslar
- Ivanovs, A., et al. Highly potent human hematopoietic stem cells first emerge in the intraembryonic aorta-gonad-mesonephros region. Journal of Experimental Medicine. 208, 2417-2427 (2011).
- Tavian, M., Biasch, K., Sinka, L., Vallet, J., Péault, B. Embryonic origin of human hematopoiesis. International Journal of Developmental Biology. 1065, 1061-1065 (2010).
- Medvinsky, A., Rybtsov, S., Taoudi, S. Embryonic origin of the adult hematopoietic system: advances and questions. Development. 138, 1017-1031 (2011).
- Ivanovs, A., et al. Human haematopoietic stem cell development: from the embryo to the dish. Development. 144, 2323-2337 (2017).
- Daniel, M. G., Pereira, C. F., Lemischka, I. R., Moore, K. A. Making a Hematopoietic Stem Cell. Trends in Cell Biology. 26, 202-214 (2016).
- Vo, L., Daley, G. De novo generation of HSCs from somatic and pluripotent stem cell sources. Blood. 125, 2641-2648 (2015).
- Rafii, S., et al. Human ESC-derived hemogenic endothelial cells undergo distinct waves of endothelial to hematopoietic transition. Blood. 121, 770-781 (2013).
- Ebina, W., Rossi, D. J. Transcription factor-mediated reprogramming toward hematopoietic stem cells. EMBO Journal. 34, 694-709 (2015).
- Sugimura, R., et al. Haematopoietic stem and progenitor cells from human pluripotent stem cells. Nature. 545, 432-438 (2017).
- Pereira, C. F., et al. Induction of a Hemogenic Program in Mouse Fibroblasts. Cell Stem Cell. 13, 205-218 (2013).
- Pereira, C. F., et al. Hematopoietic Reprogramming In vitro Informs In Vivo Identification of Hemogenic Precursors to Definitive Hematopoietic Stem Cells. Developmental Cell. 36, 525-539 (2016).
- Notta, F., et al. Isolation of Single Human Hematopoietic Stem Cells Capable of Long-Term Multilineage Engraftment. Science. 333, 218-221 (2011).
- Sinka, L., Biasch, K., Khazaal, I., Péault, B., Tavian, M. Angiotensin-converting enzyme (CD143) specifies emerging lympho-hematopoietic progenitors in the human embryo. Blood. 119, 3712-3724 (2012).
- Gomes, A. M., et al. Cooperative Transcription Factor Induction Mediates Hemogenic Reprogramming. Cell Reports. 25, 2821-2835 (2018).
- Karlsson, G., et al. Report The Tetraspanin CD9 Affords High-Purity Capture of All Murine Hematopoietic Stem Cells. Cell Reports. 4, 642-648 (2013).
- Leung, K. T., et al. The tetraspanin CD9 regulates migration, adhesion, and homing of human cord blood CD34+ hematopoietic stem and progenitor cells. Blood. 117, 1840-1851 (2011).
- Hockemeyer, D., et al. A drug-inducible system for direct reprogramming of human somatic cells to pluripotency. Cell Stem Cell. 3, 346-353 (2008).
- Kutner, R. H., Zhang, X., Reiser, J. Production concentration and titration of pseudotyped HIV-1-based lentiviral vectors. Nature Protocols. 4, 495-505 (2009).
- Suzuki, Y. S., Suzuki, Y., Ke, X. Gene Regulatable Lentiviral Vector System. Viral Gene Therapy. , (2011).
- Riddell, J., et al. Reprogramming committed murine blood cells to induced hematopoietic stem cells with defined factors. Cell. 157, 549-564 (2014).
- Lis, R., et al. Conversion of adult endothelium to immunocompetent haematopoietic stem cells. Nature. 545, 439-445 (2017).
- Pereira, C., Lemischka, I. R., Moore, K. From blood to blood’: de-differentiation of hematopoietic progenitors to stem cells. EMBO Journal. 33, 1511-1513 (2014).
- Nolan, D. J., et al. Molecular Signatures of Tissue-Specific Microvascular Endothelial Cell Heterogeneity in Organ Maintenance and Regeneration. Developmental Cell. 26, 204-219 (2013).
- Batta, K., Florkowska, M., Kouskoff, V., Lacaud, G. Direct Reprogramming of Murine Fibroblasts to Hematopoietic Progenitor Cells. Cell Reports. 9, 1871 (2014).
- Cheng, H., et al. Reprogramming mouse fibroblasts into engraftable myeloerythroid and lymphoid progenitors. Nature Communications. 7, 1-15 (2016).
Erratum
Formal Correction: Erratum: Hemogenic Reprogramming of Human Fibroblasts by Enforced Expression of Transcription Factors
Posted by JoVE Editors on 12/03/2019. Citeable Link.
An erratum was issued for: Hemogenic Reprogramming of Human Fibroblasts by Enforced Expression of Transcription Factors. The author affiliations were updated.
The second author affiliation was updated from:
2Wallenberg Center for Molecular, Lund University
to:
2Wallenberg Center for Molecular Medicine, Lund University
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır