
Method Article
İzole Murine Ventriküler Miyositlerin Optik Görüntülemesi
Bu Makalede
Özet
Murine miyositlerin izolasyonu ve aynı anda gerilim veya kalsiyum izlerinin nasıl elde edilecegini, eşzamanlı dijital hücre geometrisi ölçümleri ile floresan fotometri kullanarak sarcomere kısaltma izleriyle birlikte saglamak için metodolojiyi savuruyoruz.
Özet
Erişkin kardiyak miyositleri izole etme yeteneği, araştırmacıların tek hücre düzeyinde çeşitli kardiyak patolojileri incelemelerine olanak samıştır. Kalsiyuma duyarlı boyalarda gelişmeler tek hücreli kalsiyum dinamiğinin sağlam optik kaydına izin verirken, sağlam transmembran optik voltaj sinyallerinin kaydedilmesi zor olmuştur. Tartışmalı, bu gürültü oranı, fototoksisite ve geleneksel potansiyometrik boyalar fotobeyazlatma düşük tek nedeniyle. Bu nedenle, tek hücregerilim ölçümleri uzun altın standart iken, teknik olarak zorlu ve düşük üretim yama kelepçe tekniği ile sınırlı olmuştur. Ancak, yeni potansiyometrik boyaların geliştirilmesi ile, voltaj değişiklikleri için büyük, hızlı optik yanıtlar hiçbir fototoksisite ve fotobeyazrlama çok az elde edilebilir. Bu protokol, hücresel kısaltma, kalsiyum ve optik voltaj ölçümleri için kullanılabilecek yetişkin minürmiositlerin nasıl izole edilebildiğini ayrıntılı olarak açıklamaktadır. Özellikle, protokol nasıl bir oranmetrik kalsiyum boya, tek uyarma kalsiyum boya ve tek bir uyarma gerilim boya sıyrık kullanılır açıklar. Bu yaklaşım çeşitli kimyasal ajanların kardiyotoksisite ve aritmiojenite sini değerlendirmek için kullanılabilir. Fototoksisite hala tek hücre düzeyinde bir sorun iken, metodoloji nasıl azaltmak için tartışılır.
Giriş
Sağlıklı ve patolojik durumlarda kalbi incelemek için, genellikle tek hücre düzeyinde fenotip incelemek yararlıdır. Bilimsel gelişmeler tek hücreli kalsiyum dinamiğinin sağlam ölçümüne izin verirken, tek hücreli optik voltaj ölçümleri1'deaz kalmıştır. Tartışmalı, bu gürültü oranı (SNR), fototoksisite ve geleneksel potansiyometrik boyalar fotobeyaztma düşük sinyal nedeniyle2,3. Yine de, izole miyosit optik eylem potansiyelleri elde edilmiştir2,3,4. Ayrıca, kimya ve voltaj duyarlı boyaların fizik gelişmeler ile, SNR5geliştirdi. Daha yeni membran potansiyeli probları(Tablo Malzemeler)membran potansiyelindeki değişikliklere milisaniyelerin altında yanıt verir ve 100 mV'de yaklaşık %25 florojenik tepki aralığına sahiptir. Ayrıca, membran potansiyel kitinin uyarılması/salınımı (örn. FluoVolt; Malzeme Tablosu) Bu protokolde kullanılan standart floresan izotiyosiyanat (FITC) veya yeşil floresan protein (GFP) ayarları6ile çalışır.
FITC ve GFP uyarma/emisyon spektrumları flor-4 kalsiyum bağlı spektrum7ile örtüşmektedir. Floresan fotometrinin dijital hücre geometrisi ölçümleri ile eşzamanlı olarak edinimi geleneksel olarak kalsiyum ve hücresel kısaltma ölçümlerinin eşzamanlı olarak elde edilmesi için kullanılmıştır8. Bu protokol, murine miyositlerin nasıl izole edilebildiğini ve standart FITC ayarlarını kullanarak kalsiyum veya voltaj sinyallerinin nasıl kaydedilenini ayrıntılı olarak açıklar. Buna ek olarak, görüntüleme iş istasyonundaki uyarma/emisyon filtrelerinde basit bir anahtarın, metrik kalsiyum boya fura-2 oranını kullanarak kalsiyum ve kısaltma ölçümleri elde etmek için nasıl kullanılabileceğini açıklar. Fluo-4 ile karşılaştırıldığında, fura-2 kalsiyum için daha yüksek bir yakınlık vardır ve fotobeyazrlama nispeten dayanıklı9. Sonuç olarak, tek bir iş istasyonu kullanarak bu protokol tek bir miyosit uyarma-daralma kaplin ayrıntılı bir inceleme sağlar.
Protokol
Bu protokolde açıklanan tüm yöntem ve prosedürler Case Western Reserve Üniversitesi Kurumsal Hayvan Bakım ve Kullanım Komitesi (IACUC) tarafından onaylanmıştır.
1. Çözelti, Alet ve Kapakların Hazırlanması
NOT: 1x çözümleri bir aya kadar kullanılabilir.
- Kalsiyum olmadan 10x Krebs-Henseleit tampon HEPES tampon olun (KHB-HB) NaCl 68.96 g ekleyerek, 3.57 g KCl, 59.58 g HEPES, 2.18 g K2HPO4,MgSO4 3.08 g ve 19.82 g glukoz 800 mL çift distile su 1.000 mL şişe. İçeriği tamamen çözüldükten sonra, 1.000 mL hacimli şişede hacim haline getirin.
NOT: Geleneksel Krebs Henseleit çözeltisi tampon olarak sodyum bikarbonat kullanır ve bu protokoldeki çözelti HEPES tamponu ile Krebs Henseleit çözeltisi kullanır. Steril filtre edilirse çözelti 6 ay stabildir. - NaCl 86.51 g ekleyerek 10x Tyrode çözeltisi olun, NaH 0.552 g2PO4,MgCl2 2.03 g , glikoz 9.03 g, KCl 4.03 g,CaCl2 2 2.65 g , ve HEPES 35.76 g 800 mL çift distile su 1000 mL flask. İçeriği tamamen çözüldükten sonra, 1000 mL hacimli şişede sese kadar getirin.
NOT: Steril filtre edilirse çözelti 6 ay stabildir. - 10x stoğun 100 mL'ini ölçerek ve 1.000 mL'lik bir şişede 875 mL çift distile suya ekleyerek 1x KHB-HB yapın. Şişeyi 37 °C su banyosuna yerleştirin. Çözelti 37 °C'ye ulaştığında, pH'ı 7,39'a çıkarmak için NaOH kullanın. pH'ı ayarladıktan sonra, 1000 mL hacimli şişede çözeltiyi hacim haline getirin. Steril filtre çözeltisi vakum filtrasyon sistemi kullanarak.
- 10x stoğun 100 mL'ini ölçerek ve 1.000 mL'lik bir şişeye 875 mL çift distile su ekleyerek 1x Tyrode'un çözeltisini yapın. Şişeyi 37 °C su banyosuna yerleştirin. Çözelti 37 °C'ye ulaştığında, pH'ı 7,39'a çıkarmak için NaOH kullanın. pH'ı ayarladıktan sonra, 1.000 mL hacimli şişede çözeltiyi hacimhaline getirin. Vakum lu filtrasyon sistemi kullanılarak steril filtre.
- 10x stoğun 100 mL'ini ölçerek ve 1.000 mL'lik bir şişeye 875 mL çift distile su ekleyerek 1x modifiye tyrode çözeltisini yapın. Flask azaltılmış L-glutatyon 3.07 g çözünür. Şişeyi 37 °C su banyosuna yerleştirin. Çözelti 37 °C'ye ulaştığında, pH'ı 7,39'a çıkarmak için NaOH kullanın. pH'ı ayarladıktan sonra, 1.000 mL hacimli şişede çözeltiyi hacimhaline getirin. Steril filtre çözeltisi vakum filtrasyon sistemi kullanarak.
- 25 mg toz 855 μL dimetil sülfoksit (DMSO) ekleyerek 100 mM blebbistatin stok çözeltisi yapın. Aliquot 20 μL artışlarla dışarı ve altı aya kadar -80 °C dondurucuda saklayın.
- 1x KHB-HB 100 mL'ye 2 g büyükbaş serum albumin (BSA) ve 1 şişe aliquoted blebbistatin stoğu ekleyerek tamponu durdurun ve vakum filtrasyon sistemi kullanarak çözeltiyi steril filtreleyin.
- M199 HEPES'in 95 mL'sine 5 mL fetal büyükbaş serum ve 1 şişe aliquoted blebbistatin stoğu ekleyerek kaplama tamponu yapın. Steril filtre çözeltisi vakum filtrasyon sistemi kullanarak.
- M199 'un (25 mM HEPES) 396 mL'sine 1 şişe aliquoted blebbistatin stoğu ve 4 mls penisilin-streptomisin ekleyerek miyosit kültür tamponu yapın. Steril filtre çözeltisi vakum filtrasyon sistemi kullanarak.
- Otoklav 2 çift Dumont cımbız, Iris kavisli makas 2 çift, 2 hemostats, plastik cerrahi forceps bir çift, 6 siyah örgülü ipek 4-0 dikiş ler cerrahi çift atmak düğüm olarak kullanılmak üzere düzenlenmiş ve dört 100 mL beaers.
- 22 x 22 mm2 cam kapaklı kapakları sterilize edin. İlk olarak, altı kuyu plaka her kuyuya tek bir coverslip yerleştirin. Daha sonra, kapak çıkarıldığında, biyogüvenlik kabininin UV lambasını açın ve kapakları 1 saat uv ışığına maruz bırakın.
- Önce buz üzerinde şişe eriterek çalışma laminin stok çözüm olun. 0.04 mg/mL nihai konsantrasyonulaşmak için yeterli soğuk steril fosfat tamponlu salin (PBS) için bir şişe içeriğini ekleyin. Aliquot dışarı 1.3 μL otoklavlı 1.5 mL santrifüj tüpler içine. -80 °C'de saklayın.
NOT: Her tüp tek bir altı kuyu plakası için yeterli laminin vardır. Birden çok donma çözülme döngülerinden kaçının. - Kat önce buz üzerinde çalışan laminin çözeltisi eriterek coverslip sterilize. P1000 pipet kullanarak, 200 μL laminin aspire edin. Pipet ucunu kapak kaymasının bir kenarı boyunca hafifçe sürükleyin ve kılcal damar hareketinin altı kuyu plakasına kapak kayması eki kolaylaştırmak için küçük miktarda laminin çıkarmasını bekleyin.
- Sonra, kapak ortasında kalan laminin dışarı. Dairesel bir hareketle, laminin damlacıkkapak boyunca yayıldı. İzolasyondan önce en az 1 saat ve 24 saate kadar 37 °C'lik bir kuluçka makinesine yerleştirin.
2. Langendorff Aparatının Hazırlanması
NOT: Bu protokolde kullanılan Langendorff cihazlarınıntek tek bileşenleri Malzeme Tablosu'nda listelenmiştir.
- Dolaşan su banyosunu açın. Sıcaklığı 37 °C'lik bir sıcaklığa sahip olacak şekilde ayarlayın.
NOT: Çözelti rezervuarları 60 cm yüksekliğe ayarlı, perfusat 37 °C olması için sirkülasyon su doğum 41 °C olarak ayarlanması gerekir. Daha önce bildirilen protokollerin aksine, rezervuarYüksekliğinin değiştirilmesi gerekmez. - Langendorff cihazını %70 etanolle durulayın ve ardından otoklavlı çift distile su ile iki durulama. Durulama dan sonra, KHB-HB ile rezervuar doldurun ve% 100 oksijen ile oksijen.
- Oksijenli KHB-HB'nin ilk olarak 100 mL'lik bir kabın içine akmasına izin vererek sistemi asallayın. 50 mL çözelti kabına aktıktan sonra, KHB-HB rezervuarından gelen akışı durdurmak için 3 yönlü stop-cock pozisyonunu değiştirin. Kabından kollajenaz haznesine 50 mL oksijenli KHB-HB dökün.
- 5 mL kollajenaz rezervuar kalır kadar sindirim rezervuar ıNDAN KHB-HB drenaj sağlar. Kollajenaz rezervuar priming sırasında, çizgiler degas izin vermek için rezervuarlar arasında tekrar tekrar 3-yol stop horoz geçiş. Sistem astarlandıktan sonra, kalan havanın sistemden çıkmasına izin vermek için ısıtma bobininin üstünde bulunan gaz giderme kapanını kullanmayı unutmayın.
- Kollajenaz çözeltisini yapın. Sıçanlar için tip II kollajenaz 100 mg birleştirmek, oksijenli KHB-HB 100 mL, ve blebbistain stok 2 şişeleri. Fareler için, tip II kollajenaz 100 mg birleştirmek, oksijenli KHB-HB 40 mL, ve blebbistain stok 2 şişeleri. Bir kez karıştırılır, çözelti 1 saat için kararlı olmalıdır.
NOT: Miyosit canlılığı tip II kollajenaz kuraları arasında değişebilir. Toplu sipariş önce çok test etmek için bir kollajenaz örnekleme programı yararlanın.
3. Miyosit İzolasyon
- Hayvana 1000 U heparin enjekte et. 5 dk bekleyin.
NOT: Her yaştaki fare ve sıçanlar kullanılabilir. Ancak, genel olarak yaşlı veya daha fazla hasta hayvan, düşük miyosit verimi. - Bir pentobarbital karışım (150 mg / kg intraperitoneal) ile hayvan ötenazi önce açık-damla yöntemi (500 cc hacim başına Isoflurane 1 cc) kullanarak isoflurane ile anestezi ile ilk hayvan kurban.
- Hızlı bir şekilde ilk xiphoid süreci üzerinde kürk kapma tarafından kalp çıkarmak. Iris makas ile, xiphoid sürecinin hemen altında küçük bir kesi yapmak ve deri açığa baş doğru yukarı doğru kürk çekin.
- Ksifoid işlemini yakalayın ve göğüs boşluğuna maruz kalan diyaframı kesin. Bir tuzak kapı kesi yapmak, bir hemostat kullanarak göğüs geri çekin, ve soğuk KHB-HB artan aort üzerinde kalp çıkarmak ve yer için kavisli forceps kullanın.
- Bir stereo mikroskop ve 5 numaralı bürlipler kullanarak kalbi kanüle. Kalbin sular altında olduğundan ve kanüle emboli önlemek için kalp eksizyonu önce astarlı olduğundan emin olun. Kanülün ucunu ventriküle aort eklemenin yaklaşık 1 mm üzerinde görselleştirerek kanülün doğru konumlandırın.
NOT: Kanülasyon süresi ne kadar hızlı sayılsa, miyosit verimi de o kadar iyi. - Langendorff üzerinde stopcock döndürerek KHB-HB akışını başlatın. Kanülleri Langendorff'a bağlayın. 5 dakika için kalp perfuse.
NOT: Perfüzyon yerçekimi ne dayalı bir sistem tarafından sağlandığı için, kalpten akan akış koroner arter uyumunun bir fonksiyonu olacaktır. - KHB-HB rezervuarından sindirim tampon haznesine perfüzyon uyguluyoruz. Sindirim tamponu kalbe ulaştığında, bir zamanlayıcı ayarlayın (fare için 5 dk veya sıçan için 15 dk). Steril 100 mL kabında perfusate toplamak için emin olun. Sindirim süresi dolana kadar perfusate ile gerektiği gibi sindirim tampon rezervuar yeniden doldurun.
- Sindirimden sonra kalbin odalarını terlikli 100 mL kabında forceps ve iris makası ile ayırın. Her odayı altı kuyu tabağından ayrı bir kuyuya yerleştirin. Her kuyuya 5 mL kollajenaz çözeltisi dökün.
- Hemen makas kullanarak kalp dokusu kıyma başlar. Doku parçaları yaklaşık 1 mm3olmalıdır. Steril transfer pipetleri kullanarak, hafifçe kıyılmış kalp dokusu triturate. Çözüm bulutlu dönecek.
- Doku parçaları beyaz ve tüylü hale geldikten sonra, ters bir mikroskop kullanarak hücreleri inceleyin. Canlı hücre sayısı %80'den fazlaysa, hücreleri 100 μm'lik bir hücre süzgeci kullanarak 50 mL konik tüpe zorlayın. Kalbin her odası için farklı bir tüp ve süzgeç kullanın.
- Canlı hücre sayısı %80'den azsa, kanültinin ne kadar sürdüğünü kontrol edin. Kanülasyon süresi 5 dk'dan fazlaise, başka bir kalp deneyin. Değilse, kollajenaz örnekleme programı ile yeni kollajenaz çok say.
- 2 dk için 215 x g santrifüj ederek hücreleri pelet. Pelet kompakt ve gevşek olmamalıdır. Pelet gevşek ise, hazırlık birçok ölü hücreleri içerir. Bir doku kültürü başlık olarak, tampon durdurma 10 mL pelet resuspend.
- 2 dk için 215 x g santrifüj ederek hücreleri pelet. Pelet kompakt ve gevşek olmamalıdır. Pelet gevşek ise, hazırlık birçok ölü hücreleri içerir.
- Kaplama tamponunun 5 mL'lik hücrelerini yeniden askıya alın. Hücre sayımı gerçekleştirin. ML başına 2 x 104 hücreli son bir miyosit konsantrasyonuna ulaşmak için kaplama tamponunun mililitrelerini ayarlayın.
- Kuvözdeki laminin kaplı kapakları çıkarın. Laminin damlacıkaspire.
- Her kapak kapağında 200 μL miyosit süspansiyonplakası. 37 °C'lik bir kuluçka makinesine (%21 O2, %5 CO2)2 saat boyunca ilave yetirin. 2 saat sonra, bekar hücreleri aspire, kültür medya 2 mL ekleyin ve 4 güne kadar kültür.
4. Fura-2 Boya Yükleme
- 2 mM fura-2 asetoksimetil ester (fura-2 AM) 50 μg fura-2 tozuna (1 şişe) 25 μL DMSO ekleyerek stok çözeltisi yapın. Aliquot 6 μL aliquots içine dışarı. Fura-2 1 aliquot alın ve kaplama orta 6 mL ekleyin. Girdap karıştırmak için.
- Kuvözden 16 tane miyosit tabağı çıkarın. Aspire medya. Her kuyuya 1 mL fura-2 ortam karışımı ekleyin. Folyo ile kapak plaka, oda sıcaklığında plaka bırakın ve 15 dk bekleyin.
- Aspire fura-2 ortam karışımı ve her kuyuya Tyrode çözeltisi 1 mL ekleyin. Folyo ile kaplayın. Görüntülemeden önce boya yıkama için izin vermek için oda sıcaklığında 20 dakika bekleyin.
5. Fluo-4 Boya Yükleme
- 50 μg floro-4 tozuna (1 şişe) 25 μL DMSO ekleyerek 1,82 mM flor-4 asetoksimetil ester (flor-4 AM) stok çözeltisi yapın. Aliquot 8.333 μL aliquots içine dışarı. Fluo-4 stok 1 aliquot alın ve kaplama orta 6 mL ekleyin. Girdap karıştırmak için.
- Kuvözden 16 tane miyosit tabağı çıkarın. Aspire medya. Her kuyuya 1 mL flor-4 ortam karışımı ekleyin. Folyo ile kapak plaka, oda sıcaklığında plaka bırakın ve 15 dk bekleyin.
- Aspire floro-4 ortam karışımı ve her kuyuya 1 mL Tyrode çözeltisi ekleyin. Folyo ile kaplayın. Görüntülemeden önce boya yıkama için izin vermek için oda sıcaklığında 20 dakika bekleyin.
6. Membran Potansiyel Boya Yükleme
- Membran potansiyel kitinden Bileşen A ve Bileşen B'yi çıkarın. 15 mL konik bir tüpte, 50 μL b bileşeni ni ve 5 μL'lik a. vortex bileşenini birleştirmek. Voltaj boyakarışımı içeren 15 mL konik boruya 10 mL kaplama ortamı ekleyin. Girdap karıştırmak için.
- Kuvözden 16 tane miyosit tabağı çıkarın. Medyayı aspire edin. Her kuyuya membran potansiyel boya karışımından 800 μL ekleyin. Folyo ile kapak plaka, oda sıcaklığında plaka bırakın ve 15 dk bekleyin.
- Aspire boya medya karışımı ve her kuyuya modifiye-Tyrode çözeltisi 1 mL ekleyin. Folyo ile kaplayın.
7. Fotometri ve Şarj Birleştirilmiş Cihaz Kayıtları
- Aşağıdaki sırayla ekipmanı açın: mikroskop, ark lambası, hyperswitch, floresan arayüz sistemi, Myocam güç kaynağı, alan uyarıcı ve bilgisayar.
- Uyarma/emisyon filtre kümelerinin görüntüleme boyası için uygun olduğundan emin olun.
NOT: Fura-2 340 nm ve 380 nm ışık ta heyecanlanır. 510 nm ışık yayır. Fluo-4 ve voltaj membran boyası 485 nm ışıkta heyecanlanır ve 520 nm ışıkta yakımır. - Vakumu açarak, hortum kelepçesini tamamen açarak ve manifoldda kullanılan her 60 mL şırıngayı hafifçe dalarak sistemi asallayın. Kalsiyum kayıtları için Tyrode'un çözeltisi kullanılır. Voltaj kayıtları için modifiye Tyrode çözeltisi kullanın.
- Perfüzyon borusundaki silindir kelepçeyi ayarlayarak ısıtıcıyı açın ve akışı ayarlayın. 36 ± 1 °C'de kayıt yapın.
- Satın alma yazılımını açın. Parametrelerin doğru görüntüleme boyası için ayarlandıkundan emin olun.
- Karanlıkta, altı kuyu plakasından folyo çıkarın ve pacing odasına bir coverslip yerleştirin. Bu adımda uyarıcının kapalı olduğundan emin olun. 10x amacını kullanarak miyositlere odaklanın.
- Bir kez odak, 1 Hz alan uyarıcı tarafından pacing başlayın, 0.2 V. Yavaş yavaş 1:1 pacing elde edilene kadar voltajı artırmak. Daha sonra 1.5x eşik ulaşılır kadar voltajı artırın.
NOT: Uyarma-daralma kaplini ısıya bağlı olduğundan, hücrelerin kayıt yapmadan önce 15 dakika boyunca perfüzyona maruz kaldığından emin olun. Bu, miyositlerin oda sıcaklığından 37 °C'ye geri dönmenin şokundan kurtulmasına ve gevşek bağlanmış hücrelerin uçup gitmelerine olanak sağlar. - 10x nesnelden 40x hedefine geçin. 1:1'lik bir tempoyu takip eden bir hücreye odaklanın. Plastik tonları, görüş alanında yalnızca bir hücre olacak şekilde ayarlayın.
- Yazılımı kullanarak, iyi tanımlanmış sarcomeres üzerinde ilgi alanı kutusu yerleştirin. Uyarma ışığını başlatmak için satın alma yazılımını başlatın. Nötr yoğunluk filtrelerini kullanarak, yeterli bir SNR elde etmek için yoğunluk ayarını buna göre ayarlayın.
Sonuçlar
Şekil 1A Langendorff cihazını gösterir. Oksijenatör KHB-HB rezervuarında. Kollajenaz çözeltisi orta 60 mL şırınga haznesindedir. Gaz giderme hattı boş 60 mL şırınga haznesine bağlıdır. Başarılı bir izolasyon dan sonra, hücrelerin çoğu çubuk şeklinde ve striated olmalıdır. 40x hedefi altında, en miyositler görünür açık striations olmalıdır. Şekil 1B,C sağlıklı sıçan miyosit örneklerini gösterir. İzole edildikten sonra, hücreler morfolojileri ve elektriksel özelliklerini korurken 4 güne kadar kültürlenebilirler.
Uyarma-daralma kaplinölçmek için, hücreler daha sonra ısıtılmış bir pacing odasına yerleştirilir. Miyositler sıcaklık değişimlerine duyarlı olduğundan, kayıt yapmadan önce kapak kaymasının 15 dakika boyunca haznede dengede durmasını sağlamak önemlidir. Floresan kayıtları için uyarma dalga boyu 75 W xenon-arc ampul tarafından oluşturulur. Ksenin-ark ampulleri doğal güneş ışığını taklit eden bir ışık spektrumu üretir. Işığın yoğunluğu ve dalga boyu nötr yoğunluk/emisyon filörleri tarafından kontrol edilir. Uyarma ışığı daha sonra hedeften miyosite geçer. Emisyon dalga boyu daha sonra bir fotoçarpan tüp tarafından toplanır. Burada açıklanan sistemi kullanarak, hem uyarma hem de emisyon filtrelerinin el ile değiştirilmesi gerekir.
Öte yandan kısaltma bir şarj birleştirilmiş cihaz sensörü ile elde edilir. Saniyede 1.000 keze kadar gerçek zamanlı olarak ölçüm yapan satın alma yazılımı, iyi çözülmüş bir striation deseni oluşturmak için ilgi alanı içindeki satırların ortalamasını gerçekleştirir. Daha sonra hızlı fourier dönüşümü (FFT) hesaplanır. Güç spektrumu içindeki tepe ortalama sarcomere aralığını temsil eder. Daha sonra pacing sırasında sarcomere aralıkdeğişiklikleri çizilir ve daha sonra ölçülür.
Şekil 2, kalsiyum boya fura-2 yüklü bir C57/B6 fare miyositinden kaydedilen kalsiyum ve kısaltma izlerini gösterir. Pacing protokolü daha önce açıklanan pacing protokolleri bir değişiklik10,11. Sağlıklı fare miyositleri istirahat kalp hızında tempolu olmalıdır 10 Hz. Şekil 3, c57/B6 farelerinden elde edilen ortalama verilerin ve potasyum kanalına bir nokta mutasyonu getirilen transgenik (TG) çöplerinin ölçülmesidir. Dikkat edin 10 Hz pacing de gevşeme süresi dışında gruplar arasında hiçbir fark yoktur.
Çift uyarma boyası olan fura-2'nin aksine, voltaj boyası ve flor-4, uyarma/emisyon çalışmaları standart FITC uyarma ve emisyon spektrumu (494/506 nm) ile çalışan tek dalga boyu uyarma boyalarıdır. Bu nedenle bu filtre seti kullanılarak kalsiyum ve sarcomere kısaltma sıyrık veya gerilim ve sarcomere kısaltma kayıtları elde edilebilir.
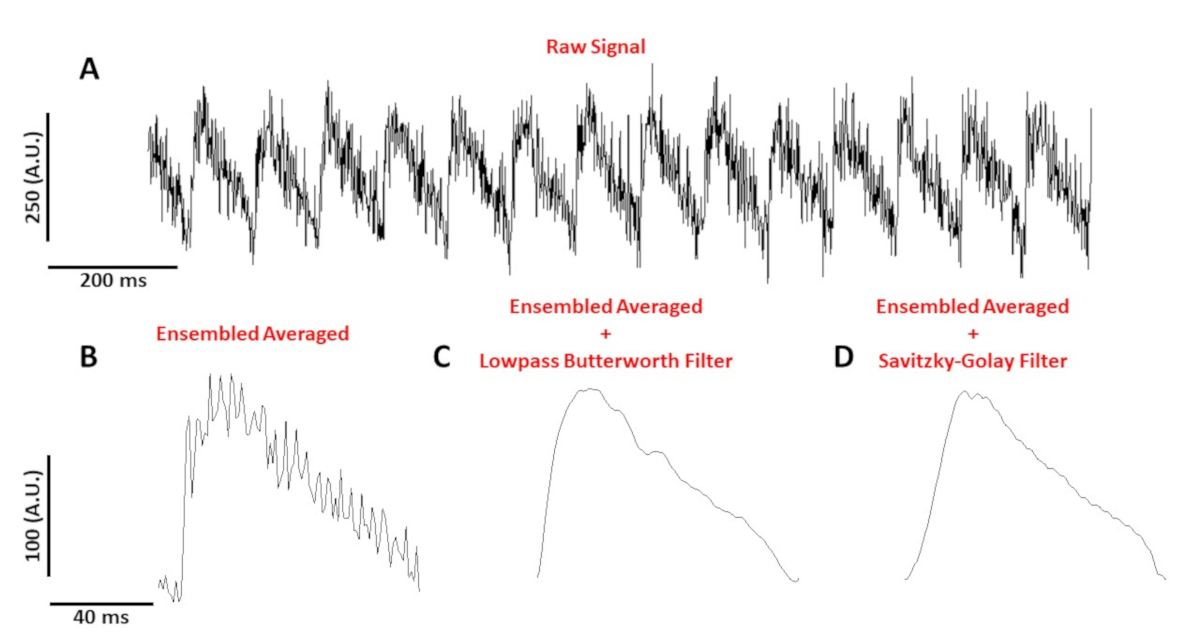
Şekil 4A, 10 Hz.'de bir C57/B6 fare miyositinden kaydedilen bir gerilim izlemesini gösterir. Şekil 4B, Şekil 4A'dakiAP'lerden yapılan ortalama bir eylem potansiyelini (AP) gösterir. Şekil 4C,D düşük geçiş Butterworth veya Savitzky-Golay dijital filtre uygulandıktan sonra bir topluluk ortalama AP gösterir. Sinyal filtreleme yaparken gerçek verileri bozmamaya özen gerekir. Şekil 4B-D'dekiAP'lerin şeklindeki ince farklılıklara dikkat edin.
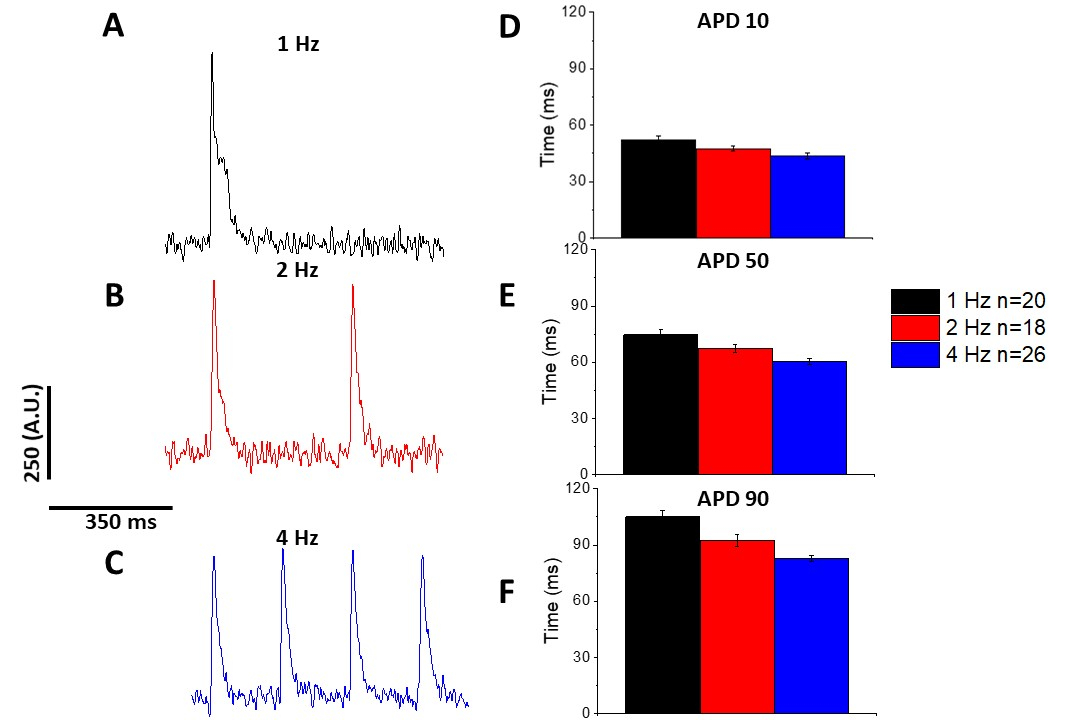
Şekil 5 1 Hz'de ilerleyen fare miyositlerinden kaydedilen izleri göstermektedir. Gerilim sinyalinin kalsiyum sinyalinden daha düşük olmasına ek olarak, kasılma kinetiği de farklıdır. Bunun nedeni kalsiyum boyaları tampon kalsiyum ise voltaj boyaları yok olmasıdır.
Kalsiyum geçicisinde olduğu gibi(Şekil 3),miyositler optik etki potansiyel sürelerinde (APD) da pacing bağımlı değişiklikler göstermiştir (Şekil 6). Fura-2 izleri sayısallaştırılmadan önce ortalama olarak gruplandırılırken, gerilim izleri ortalama ve ölçülmeden önce Savitzky-Golay polinom yumuşatma filtresi (genişlik 5, sıra 2) ile filtrelenmiştir.
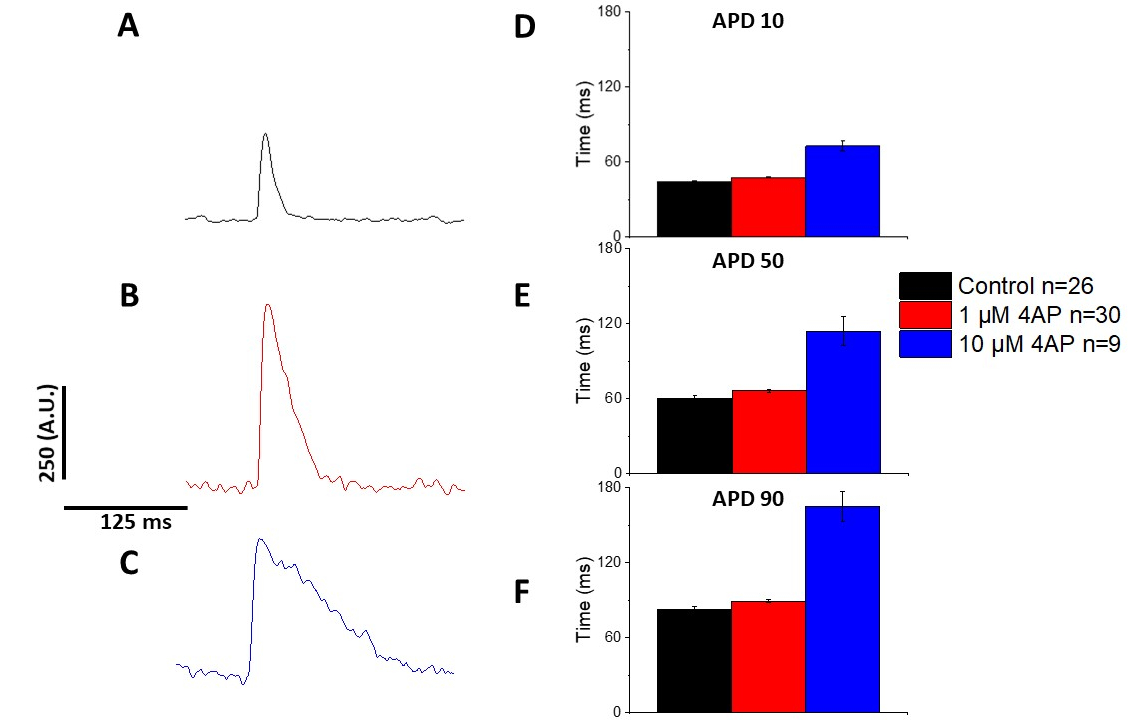
Şekil 6 ve Şekil 7'dede belirtildiği gibi, APD'de pacing kaynaklı değişiklikleri göstermenin yanı sıra, APD'nin uyuşturucuya bağlı uzamasını da göstermiştir. 4 Hz pacing, geçici dış akım konsantrasyon bağımlı abluka(I)ile 4-aminopiridin APD uzaması ile sonuçlandı.
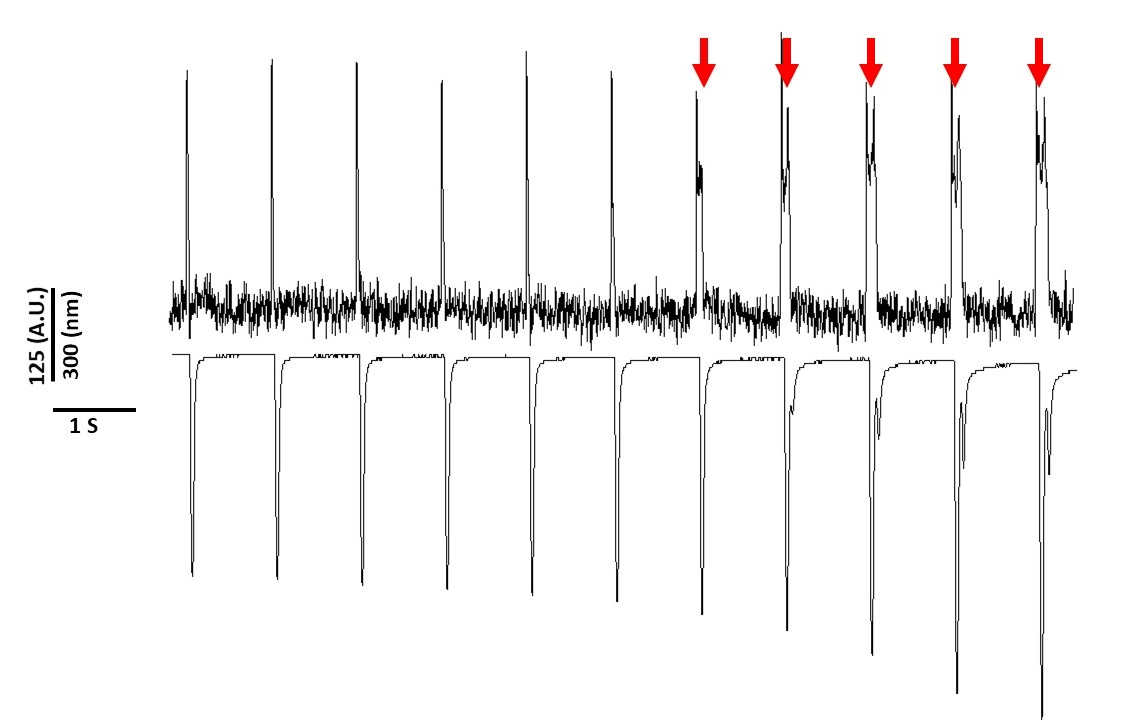
Son olarak, sitotoksisite önlemek için dikkatli olunmalıdır. Şekil 8, 20'lik bir kaydın son 11'i. Şekil 8'dekikırmızı oklarla gösterilen miyositlerin mavi ışığa uzun süre maruz kalmaları tetiklenen aktiviteye yol açar.

Şekil 1: Sabit basınç Langendorff aparatı. (A) Langendorff Aparatı ve her bileşen beyaz harflerle etiketlenmiştir. (B) İzole Sprague-Dawley sıçan miyositleri 10x nesnel olarak izlenir. (C) İzole sıçan miyositleri 40x nesnel olarak izlenir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: Fura-2 kullanılarak C57/B6 miyonlarından kaydedilen temsili kalsiyum ve sarcomere kısaltma izleri. 1, 2, 4, 10, 0.5 ve 0.75 Hz.
{kind=link}

Şekil 3: C57/B6 yabani tip (WT) ve transgenik (TG) farelerden kaydedilen sarcomere kısalması, pik kalsiyum, gevşeme süresi ve geri alım süresinin ölçülmesi. (A) Sarcomere kısaltma. (B) Pik kalsiyum. (C) Gevşeme süresi olarak tanımlanan 90% kısaltma iz taban çizgisine dönüş. (D) Kalsiyum izinin taban çizgisine %90 geri dönüş olarak tanımlanan geri alım süresi. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: C57/B6 fare miyositinden kaydedilen optik etki potansiyeli 10 Hz.'de hareket etti. (A) 1 saniye filtresiz izleme. (B) Ortalama optik aksiyon potansiyeli. (C) Lowpass Butterworth filtresi uygulandıktan sonra ortalama optik aksiyon potansiyeli elde edilebilmektedir. (D) Savitzky-Golay polinom yumuşatma filtresi uygulandıktan sonra ortalama optik aksiyon potansiyeli elde edilebilmektedir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 5: Sprague-Dawley sıçan miyositlerinden kaydedilen temsili kalsiyum, gerilim ve sarcomere kısaltma izleri 1 Hz'de ilerler. (A) Flor-4 kullanılarak 1 Hz pacing'de kaydedilen kalsiyum ve sarcomere kısaltma izleri. (B) Gerilim boyası kullanılarak 1 Hz pacing'de kaydedilen voltaj ve sarcomere kısaltma izleri. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 6: Sprague-Dawley sıçan miyositlerinden kaydedilen optik aksiyon potansiyelleri 1, 2 ve 4 Hz tempoludur. (A) 1 Hz pacing'de kaydedilen filtrelenmiş iz. (B) Filtrelenmiş iz 2 Hz pacing kaydedilir. (C) 4 Hz pacing'de kaydedilen filtrelenmiş iz. (D) Eylem potansiyel süresi 10, %10 olarak ölçülen taban çizgisine geri dönüş. (E) Eylem potansiyel süresi 50, %50 olarak ölçülen taban çizgisine geri dönüş. (F) Eylem potansiyel süresi 90, %90 olarak ölçülen taban çizgisine geri dönüş. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 7: 4 Hz pacing kaydedilen Sprague-Dawley sıçan optik aksiyon potansiyelleri üzerinde 4-aminopyridine Etkileri. (A) 4 Hz'de kaydedilen ortalama iz çözeltide 4-Aminopiridin yoktur. (B) Çözeltide 1 μM 4-Aminopyridine ile 4 Hz pacing'de kaydedilen ortalama iz. (C) Çözeltide 10 μM 4-Aminopyridine ile 4 Hz pacing'de kaydedilen ortalama iz. (D) Eylem potansiyel süresi 10, %10 olarak ölçülen taban çizgisine geri dönüş. (E) Eylem potansiyel süresi 50, %50 olarak ölçülen taban çizgisine geri dönüş. (F) Eylem potansiyel süresi 90, %90 olarak ölçülen taban çizgisine geri dönüş. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 8: Gerilim boyası Sürekli ışığa maruz kaldıktan sonra Sprague-Dawley sıçan miyositlerinde fototoksisiteye neden oldu. Kırmızı oklar sitotoksik olayları gösterir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
Kardiyak miyositleri izole edebilmek, kardiyak fizyoloji, patoloji ve toksikolojiyi anlamak için kullanılabilecek güçlü bir yöntemdir. Yukarıdaki protokolde, tek kardiyak miyosit elde etmek için sabit bir yerçekimi basıncı Langendorff cihazı kullanan bir yöntem açıklanmıştır. Daha sonra, floresan fotometri sistemini kullanarak, aynı anda ya kalsiyum ve kısaltma veya voltaj ve kısaltma izleri elde etmek için nasıl açıklayın.
Kalsiyum boyalar arasındaki farklı kinetik nedeniyle, hangi boyanın seçilene dikkat edilmelidir. Bu protokol için, hem fura-2 hem de fluo-4, hücre içi esterlerin grubunu ayırması ve boyayı hücreye hapsetmesi için bir yıkama adımı gerektiren esterleri ile tasarlandı. Hem fura-2 hem de fluo-4 yüksek afinite kalsiyum boyaları olarak kabul edilirken, fura-2 için Kd 145 nM fluo-49için 345 nM ile karşılaştırıldığında. Ayrıca, fura-2 oranmetriktir. Bu nedenle, hücre içi kalsiyum düzeyleri ölçmek için kullanılabilir9,12. Fluo-4 ise tek dalga kalsiyum sondadır. Fluo-4 kullanmanın avantajı daha parlak bir floresan sinyali üretmesidir. Hangi kalsiyum boya kullanılırsa kullanılsın, kalsiyum boyasına göre membran voltaj probları daha düşük SNR'ye sahiptir.
Şekil 4 ve Şekil 5'tegösterildiği gibi, kalsiyum izlerine göre gerilim izleri genlik olarak daha küçüktür. Yazılımın dijital izleme filtreleme sini kullanarak SNR'yi artırmak ve verileri ölçmek mümkündür(Şekil 4 ve Şekil 7). Bir kez sayısallaştırılmış, hem kalsiyum geçici hem de optik APD'ler geri alma gösterirler ve daha hızlı tempo frekanslarında sürelerini kısaltırlar (Şekil 2, Şekil 3, Şekil 6, ve Şekil 7). Daha hızlı pacing döngüleri sırasında daha kısa APDs diastole sırasında ventriküler dolgu için yeterli zaman sağlamak için gereklidir. Bu fenomendeğişiklikler arrythmias riski bir artış göstergesi olduğu düşünülmektedir13,14,15,16. APD değişiklikler hastalık neden olabilir iken, onlar da kimyasallar neden olabilir. Şekil 7'degösterildiği gibi, baskın murine repolarize potasyum akımı, I,bloke olduğunda, optik APD daha uzun olur.
Yine de, voltaj duyarlı boyalar ile daha önce bildirildiği gibi, ışık yoğunluğu ve süresiAPD2,5,17değiştirebilirsiniz. Bu reaktif oksidatif türlerin nesil sonucu olduğuna inanılmaktadır (ROS)5. Daha önce, bu kayıt çözeltisine antioksidan eklenmesi voltaj duyarlı boya sitotoksisite önleyebilir gösterilmiştir5. Sonuç olarak, Tyrode'un çözümüne antioksidan L-glutatyon (10 mM) ekledik. Shown in Figure 8 is the last 11 s of a 20 s recording obtained at 1 Hz pacing. Kırmızı oklarda belirtildiği gibi, APD'de 15 s'ye kadar değişiklikler meydana gelmedi; bu nedenle, modifiye Tyrode's çözüm fototoksisite önlemek olmasa da önemli ölçüde geciktirdi. Modifiye Tyrode'un çözeltisini kullanarak, düşük ışık yoğunluğu ayarını kullanarak ve kayıt süresini 5'in altında tutmak, APD'de boya kaynaklı değişikliklerden kaçınmak mümkündür. Bu önemlidir, çünkü fototoksisiteden kaçınmaya özen yapmadan, veriler depolarizasyondan sonra erken veya gecikmeye neden olduğu için yanlış yorumlanabilir. Mavi ışığa maruz kalma sınırlama ek olarak, verilerin yanlış yorumlanmasını önlemek için alınabilir ek önlemler vardır.
Bunlardan ilki, sadece bire bir tempoyu takip eden ve 1,75 μm'den daha büyük veya eşit bir istirahat sarcomere uzunluğuna sahip hücrelerden kaydetmektir. 1.75 μm kesme Gordon ve ark.18 tarafından sarcomere uzunluğu bu miktarın altında olduğunda gerilim hızla azalır gözlem alınır. Bununla birlikte, bazı patolojiler sarcomere uzunluğu istirahat önemli değişikliklere neden olabilir. Fenotipin izolasyon un bir artifakı değil de gerçek olduğundan emin olmak için aşağıdaki sorun giderme yaklaşımları alınmalıdır.
Miyositler sürekli olarak 1:1 pacing'i takip etmiyorsa, sarcomere uzunlukları 1,75 μm'nin altındaysa, ağır membran blebbing'i varsa veya izolasyondan sağ çıkamazsa, kontrol edilmesi gereken ilk şey kalbi kanüle etmek için gereken süredir. Kanülasyon süresi ne kadar uzun sayılsa, verim o kadar düşük olur. Uzun bir kanülasyon süresi gerekiyorsa, kardiyoplegic çözelti19kalp yerleştirerek canlılık geliştirilebilir. Yine de, kollajenaz bir enzim olduğu için, aktivite ve belirli bir çok özgüllük zaman içinde değişir. Genel verimleri giderek iyi kanülasyon kez rağmen kötüleşirse, yeni çok titretilmelidir. Protokolümüz 5 s kayıtları için optimize edilmiş olsa da, daha uzun voltaj izlemeleri gerekiyorsa, ek nötr yoğunluk filtreleri satın alınması gerekecektir. Protokolde tanımlanan sistem, iletilen ışığı %37, %50, %75, %90 ve %95 azaltan nötr yoğunluk filtreleriyle birlikte gelir.
Özetle, kalsiyum, voltaj ve sarcomere kısaltma ölçümleri için kullanılan erişkin mintriküler miyositlerin izolasyonuna izin veren bir metodoloji tanımladık.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Dana Morgenstern'e müsveddeleri dikkatlice okuduğu için teşekkür ederiz.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 0.25 Liter Water Jacketed Reservoir | Radnoti, LLC | 120142-025 | |
| 1 liter volumetric flask | Fisher Scientific | 10-205F | |
| 100 ml beaker | Fisher Scientific | FB-100-100 | |
| 100 ml graduated cylinder | Fisher Scientific | 08 562 5C | |
| 1000 ml flask | Fisher Scientific | FB-500-1000 | |
| 2-Bar Lab Stand with Stabilizer Bar and 24" Stainless Steel Rods | Radnoti, LLC | 159951-2 | |
| 4-Aminopyridine | Sigma-Aldrich | 275875 | |
| 40X Oly UApo/340 Non-Immersion Objective (NA 0.9, WD 0.2mm) | IonOptix | MSCP1-40 (b) | |
| 60-mL syringe, BD Luer-Lok tip | BD | 309650 | |
| Aortic Metal Cannulae | Harvard Apparatus | 73-0112 | |
| Bovine Serum Albumin | Fisher Scientific | BP9703-100 | |
| C-6 Standard Heating Circulator | Chemyx | A30006 | |
| CaCl2 | Fisher Scientific | BP510500 | |
| Cell framing adapter | IonOptix | CFA300 | |
| CellPr0 Vacuum Filtration System, 1 liter, 0.22µm,Cs/12 | Labratory Product Sales, Inc | V100022 | |
| CellPro Vacuum Filtration System, 250mL, 0.22µm,Cs/12 | Labratory Product Sales, Inc | V25022 | |
| CellPro Vacuum Filtration System, 500mL, 0.22µm,12/CS | Labratory Product Sales, Inc | V50022 | |
| CMC (mTCII) Temp Control w/ inline flow heater | IonOptix | TEMPC2 | |
| Cole-Parmer Large-bore 3-way, male-lock, stopcocks | Cole-Parmer | EW-30600-23 | |
| Cole-Parmer Luer fittings, Large-bore stopcocks, male lock, 4-way | Cole-Parmer | EW-30600-12 | |
| Cole-Parmer Stopcocks with Luer Connections; 1-way; male slip | Cole-Parmer | EW-30600-01 | |
| Collagenase Type II | Worthington | LS004177 | |
| Corning Sterile Cell Strainers | Fisher Scientific | 07-201-432 | |
| Dell Optiplex 790 mini-tower, 4G RAM, 250G HD, Windows 7 Pro | IonOptix | CPUD7M | |
| DMSO | Fisher Scientific | 50980367 | |
| Dumont Tweezers Style 5 | Amazon | B00F70ZDEQ | |
| FHD Rapid Change Stimulation Chamber | IonOptix | FHDRCC1 | |
| Fluo-3/4 Optics Package | IonOptix | IonOP-Fluo | |
| Fluorescence system interface – (w PCI-I/O card) | IonOptix | FSI700 | |
| Gibco Penicillin-Streptomycin (10,000 U/mL) | Fisher Scientific | 15-140-122 | |
| Glucose | Fisher Scientific | D16-1 | |
| Hemostat, Curved 5-1/2" | Amazon | B00GGAAPD0 | |
| HEPES | Fisher Scientific | BP310500 | |
| HyperSwitch dual excitation light source | IonOptix | HSW400 | |
| Inverted Motic Fluorescence Microscope | IonOptix | MSCP1-40 (a) | |
| IonWizard Core + Analysis | IonOptix | IONWIZ | |
| Iris Scissors, curved | Amazon | B018KRRMY6 | |
| K2HPO4 | Fisher Scientific | P288-100 | |
| KCl | Fisher Scientific | BP3661 | |
| L-Glutathione reduced | Sigma-Aldrich | G4251 | |
| LOOK Silk Spool, Black Braided, 4-0, 100yds | SouthernAnesthesiaSurgical Inc. | SP116-EA | |
| M199 Media | Fisher Scientific | 12 340 030 | |
| MgCl2 | Fisher Scientific | MP021914215 | |
| MgSO4 | Fisher Scientific | BP2131 | |
| MyoCam-S Digital CCD video system | IonOptix | MCS100 | |
| MyoPacer Field Stimulator | IonOptix | MYP100 | |
| NaCl | Fisher Scientific | BP358212 | |
| NaH2PO4 | Fisher Scientific | 56-754-9250GM | |
| Oxygenator Bubbler with Fluid Inlet for 0.25 Liter | Radnoti, LLC | 140143-025 | |
| Photomultiplier sub-system | IonOptix | PMT400 | |
| PMT Acquisition add-on | IonOptix | PMTACQ | |
| Radnoti Heating Coil 5 mL with Degasing Trap | Radnoti, LLC | 158830 | |
| Ring Clamp 60 – 80mm Dia. for 250ml Reservoir | Radnoti, LLC | 120141-025 | |
| Ring Clamp for Bubble Trap Compliance Chamber | Radnoti, LLC | 120149RC | |
| Saint-Gobain ACF000010 5/32 in.9/32 in. | Fisher Scientific | 14-171-214 | |
| Saint-Gobain ACF000013 3/16 in.3/8 in. | Fisher Scientific | 14-171-217 | |
| Saint-Gobain ACF000016 1/4 in.5/16 in. | Fisher Scientific | 14-171-219 | |
| Saint-Gobain ACF000025 5/16 in.5/8 in. | Fisher Scientific | 14-171-226 | |
| Saint-Gobain ACF00003 1/16 in.3/16 in. | Fisher Scientific | 14-171-209 | |
| Saint-Gobain ACF00005 1/16 in.3/32 in. | Fisher Scientific | 14-171-210 | |
| Saint-Gobain ACF00009 5/32 in.7/32 in. | Fisher Scientific | 14-171-213 | |
| Sarcomere Length Recording add-on | IonOptix | SARACQ | |
| T/C Adson Tissue Platic Surgery Forceps 4.75" | Amazon | B00JDWRBGC | |
| VETUS Anti-Static Curved Tip Tweezers | Amazon | B07QMZC94J | |
| Vistek 3200 Motic Vibration Isolation Platform | IonOptix | ISO100 |
Referanslar
- Hagen, B. M., Boyman, L., Kao, J. P., Lederer, W. J. A comparative assessment of fluo Ca2+ indicators in rat ventricular myocytes. Cell Calcium. 52 (2), 170-181 (2012).
- Schaffer, P., Ahammer, H., Muller, W., Koidl, B., Windisch, H. Di-4-ANEPPS causes photodynamic damage to isolated cardiomyocytes. Pflügers Archiv: European Journal of Physiology. 426 (6), 548-551 (1994).
- Hardy, M. E., Lawrence, C. L., Standen, N. B., Rodrigo, G. C. Can optical recordings of membrane potential be used to screen for drug-induced action potential prolongation in single cardiac myocytes. Journal of Pharmacological and Toxicological Methods. 54 (2), 173-182 (2006).
- Tian, Q., et al. Optical action potential screening on adult ventricular myocytes as an alternative QT-screen. Cellular Physiology and Biochemistry. 27 (3-4), 281-290 (2011).
- Warren, M., et al. High-precision recording of the action potential in isolated cardiomyocytes using the near-infrared fluorescent dye di-4-ANBDQBS. The American Journal of Physiology-Heart and Circulatory Physiology. 299 (4), 1271-1281 (2010).
- FluoVolt™ Membrane Potential Kit. ThermoFisher Scientific Available from: https://assets.thermofisher.com/TFS-Assets/LSG/manuals/MAN0009668_FluoVolt_Membrane_Potential_Kit_UG.pdf (2018)
- Fluorescence SpectraViewer. ThermoFisher Scientific Available from: https://www.thermofisher.com/us/en/home/life-science/cell-analysis/labeling-chemistry/fluorescence-spectraviewer.html (2019)
- Paredes, R. M., Etzler, J. C., Watts, L. T., Zheng, W., Lechleiter, J. D. Chemical calcium indicators. Methods. 46 (3), 143-151 (2008).
- Davis, J., et al. Diastolic dysfunction and thin filament dysregulation resulting from excitation-contraction uncoupling in a mouse model of restrictive cardiomyopathy. The Journal of Molecular and Cellular Cardiology. 53 (3), 446-457 (2012).
- Ren, J., Davidoff, A. J. Diabetes rapidly induces contractile dysfunctions in isolated ventricular myocytes. The American Journal of Physiology. 272, 148-158 (1997).
- Roe, M. W., Lemasters, J. J., Herman, B. Assessment of fura-2 for measurements of cytosolic free calcium. Cell Calcium. 11 (2-3), 63-73 (1990).
- Goldhaber, J. I., et al. Action potential duration restitution and alternans in rabbit ventricular myocytes: the key role of intracellular calcium cycling. Circulation Research. 96 (4), 459-466 (2005).
- Weiss, J. N., et al. The dynamics of cardiac fibrillation. Circulation. 112 (8), 1232-1240 (2005).
- Baher, A., et al. Short-term cardiac memory and mother rotor fibrillation. The American Journal of Physiology-Heart and Circulatory Physiology. 292 (1), 180-189 (2007).
- Kleber, A. G., Rudy, Y. Basic mechanisms of cardiac impulse propagation and associated arrhythmias. Physiological Reviews. 84 (2), 431-488 (2004).
- McPheeters, M. T., Wang, Y. T., Werdich, A. A., Jenkins, M. W., Laurita, K. R. An infrared optical pacing system for screening cardiac electrophysiology in human cardiomyocytes. PLoS ONE. 12 (8), 0183761 (2017).
- Gordon, A. M., Huxley, A. F., Julian, F. J. The variation in isometric tension with sarcomere length in vertebrate muscle fibres. The Journal of Physiology. 184 (1), 170-192 (1966).
- Li, Y., et al. Three Preservation Solutions for Cold Storage of Heart Allografts: A Systematic Review and Meta-Analysis. The Journal of Artificial Organs. 40 (5), 489-496 (2016).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır