
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
التحليل المورفولوجي والتركيبي لمصائد العدلات خارج الخلية التي تسببها المحفزات الميكروبية والكيميائية
In This Article
Summary
يظهر هنا بروتوكول لتحريض وتحليل مصائد العدلات خارج الخلية في المختبر (NETs). أسفر القياس الكمي للحمض النووي والكاثيليسيدين (LL37) ونشاط الإنزيم عن بيانات تظهر التباين في تكوين ومورفولوجيا الشبكات التي تسببها المحفزات الميكروبية والكيميائية في ظل ظروف مضبوطة مماثلة.
Abstract
تعمل العدلات كخط أول للدفاع الخلوي في الاستجابة المناعية الفطرية من خلال استخدام آليات متنوعة ، مثل تكوين مصائد العدلات خارج الخلية (NETs). تحلل هذه الدراسة التغيرات المورفولوجية والتركيبية في الشبكات التي تسببها المحفزات الميكروبية والكيميائية باستخدام منهجيات موحدة في المختبر لتحريض NET وتوصيف الخلايا البشرية. تسمح الإجراءات الموضحة هنا بتحليل مورفولوجيا NET (lytic أو non-lytic) وتكوينها (هياكل بروتين الحمض النووي والنشاط الأنزيمي) ، وتأثير العوامل القابلة للذوبان أو الاتصال الخلوي على هذه الخصائص. بالإضافة إلى ذلك ، يمكن تعديل التقنيات الموضحة هنا لتقييم تأثير العوامل الخارجية القابلة للذوبان أو الاتصال الخلوي على تكوين NET.
تشمل التقنيات المطبقة تنقية الخلايا متعددة الأشكال النووية من الدم المحيطي البشري باستخدام تدرج مزدوج الكثافة (1.079-1.098 جم / مل) ، مما يضمن النقاء الأمثل والجدوى (≥ 95٪) كما يتضح من تلطيخ رايت ، واستبعاد تريبان الأزرق ، وقياس التدفق الخلوي ، بما في ذلك تحليل FSC مقابل SSC وتلطيخ 7AAD. يتم تحفيز تكوين NET مع الميكروبات (Pseudomonas aeruginosa ، المكورات العنقودية الذهبية ، والمبيضات البيض) والمحفزات الكيميائية (أسيتات phorbol myristate ، HOCl) ، وتتميز NETs بتلطيخ DNA-DAPI ، والتلوين المناعي لكاثيليسيدين الببتيد المضاد للميكروبات (LL37) ، وقياس النشاط الأنزيمي (إيلاستاز العدلات ، والكاثيبسين G ، والميلوبيروكسيداز). يتم الحصول على الصور من خلال الفحص المجهري الفلوري وتحليلها باستخدام ImageJ.
Introduction
العدلات هي الكريات البيض الأكثر وفرة في مجرى الدم ، وتلعب دورا أساسيا أثناء إزالة العوامل المسببة للأمراض من خلال عدة آليات ، بما في ذلك إطلاق هياكل الكروماتين الكبيرة المكونة من الحمض النووي والعديد من البروتينات المضادة للبكتيريا النووية والسيتوبلازمية والحبيبية 1,2. تم إجراء السابقة المباشرة التي تصف هذا الدور المضاد للميكروبات للعدلات بواسطة Takei et al.3 في عام 1996. أبلغ هؤلاء المؤلفون عن شكل جديد من الموت يختلف عن موت الخلايا المبرمج والتنخر في العدلات ، وأظهروا تغيرات مورفولوجية تظهر تمزقا نوويا ، يليه انسكاب من بلازما النواة إلى السيتوبلازم ، وزيادة في نفاذية الغشاء من 3 ساعات من الحضانة مع أسيتات ميريستات فوربول (PMA) 2,3. ومع ذلك ، لم يتم استخدام مصطلح "مصائد العدلات خارج الخلية (NETs)"حتى عام 2004 4.
لوحظ تكوين NET في ظروف مختلفة ، مثل الالتهابات البكتيرية والفطرية5 والفيروسية6 والطفيلية ، لتحييد وقتل ومنع انتشار الميكروبات7. تظهر دراسات أخرى أنه يمكن أن يحدث أيضا في الحالات غير المسببة للأمراض عن طريق المنبهات المعقمة ، مثل السيتوكينات وحمض اليوريك أحادي الصوديوم أو بلورات الكوليسترول والأجسام المضادة الذاتية والمجمعات المناعية والصفائح الدموية المنشطة7. كانت عديد السكاريد الشحمي (LPS) وإنترلوكين -8 (IL-8) و PMA من بين المحفزات الأولى في المختبر الموصوفة بأنها محرضات NET ، وتم إثبات مشاركة NET في الجسم الحي في العمليات المسببة للأمراض في نموذجين من الالتهاب الحاد: الزحار التجريبي والتهاب الزائدة الدودية البشري التلقائي4. الحمض النووي هو مكون NET أساسي. هيكلها وتكوينها المناسبان ضروريان لعزل الكائنات الحية الدقيقة وقتلها عن طريق توصيل تركيز محلي عال من جزيئات مضادات الميكروبات نحو الميكروبات التي تم اصطيادها ، كما يتضح من علاج ديوكسي ريبونوكلياز (DNase) الموجز الذي يفكك الشبكات وخصائصها القاتلةللميكروبات 4. إلى جانب الحمض النووي ، تشتمل الشبكات على بروتينات متصلة مثل الهستونات ، وإيلاستاز العدلات (NE) ، والكاثيبسين G (CG) ، والبروتيناز 3 ، واللاكتوفيرين ، والجيلاتيناز ، والمايلوبيروكسيديز (MPO) ، والببتيدات المضادة للميكروبات (AMPs) مثل الببتيد الكاتيوني المؤيد للالتهابات cathelicidin LL-37 من بين أمور أخرى 8,9. قد تشكل هذه المجاميع خيوطا أكبر بأقطار تصل إلى 50 نانومتر. هذه العوامل يمكن أن تعطل عوامل الفوعة الميكروبية أو سلامة غشاء الخلية الممرضة. بالإضافة إلى ذلك ، يمكن ل AMPs تثبيت الحمض النووي المشتق من NET ضد التدهور بواسطة النيوكليازات البكتيرية10.
لم يتم بعد توضيح الآليات المحددة التي تنظم تكوين NET بشكل كامل. أفضل مسار مميز يؤدي إلى إطلاق NET هو من خلال إشارات ERK ، مما يؤدي إلى تنشيط NADPH oxidase وإنتاج أنواع الأكسجين التفاعلية (ROS) ، بالإضافة إلى زيادة الكالسيوم داخل الخلايا الذي يؤدي إلى تنشيط مسار MPO. وهذا بدوره يحول بيروكسيد الهيدروجين إلى حمض هيبوكلوروس ، وينشط NE عن طريق الأكسدة11,12. NE مسؤول عن تحلل خيوط الأكتين في الهيكل الخلوي لمنع البلعمة ونقلها إلى النواة للمعالجة عن طريق الانقسام المحلل للبروتين وإزالة الأمين بواسطة PAD4 التي تدفع إزالة حساسية ألياف الكروماتين ، والتي ترتبط بالبروتينات الحبيبية والسيتوبلازمية ، ثم يتم إطلاقها خارج الخلية7. تشمل هذه البروتياز تلك المنبعثة من مجمع الأزوروسوم لحبيبات الأزوروفيل والبروتياز الأخرى مثل cathepsin G13.
اعتمادا على التغيرات المورفولوجية في العدلات ، يتم تصنيف الشبكات إلى نوعين: تكوين NET انتحاري أو lytic يؤدي إلى موت الخلية4 ، وتشكيل NET حيوي أو غير lytic ينتج عن خلايا قابلة للحياة بوساطة إطلاق حويصلي من الحمض النووي أو الميتوكوندريا ، مع بقايا من cytoplast anucleated مع القدرة البلعمية14,15. بشكل عام ، تقدم الشبكات المكونة من الحمض النووي للميتوكوندريا مورفولوجيا ممدودة من الألياف14 ، في حين أن تلك المكونة من الحمض النووي النووي لها مظهر يشبه السحابة3. ومع ذلك ، من غير المعروف كيف يختار العدلة أصل الحمض النووي. على عكس الدراسات السابقة التي وصفت المسارات الكنسية للشبكات بأنها تتطلب عدة ساعات ، يتم تنشيط المسار الحيوي بسرعة في 5-60 دقيقة15 دقيقة فقط.
على الرغم من هذه التطورات ، يختلف التكوين الصافي اعتمادا على الحافز. على سبيل المثال ، تحفز سلالات مخاطية وغير مخاطية مختلفة من P. aeruginosa تكوين شبكات تحتوي على 33 بروتينا شائعا وما يصل إلى 50 بروتينا متغيرا7. وبالتالي ، من الضروري تجانس التقنيات التي تسمح بتوليد استنتاجات موضوعية في مجموعات البحث. تصف هذه الورقة بروتوكولا بتقنيات مختلفة تسمح بمقارنة وتقييم تكوين وهيكل ومورفولوجيا الشبكات المستحثة مع الكائنات الحية الدقيقة المختلفة: المكورات العنقودية الذهبية (بكتيريا إيجابية الجرام) ، Pseudomonas aeruginosa (بكتيريا سالبة الجرام) ، والمبيضات البيض (الفطريات) ، وكذلك المحفزات الكيميائية (PMA ، HOCl) في العدلات البشرية من الأفراد الأصحاء. توضح النتائج التمثيلية عدم تجانس الشبكات اعتمادا على محفزاتها المحفزة في ظل ظروف مماثلة في المختبر ، والتي تتميز بتلطيخ DNA-DAPI ، والتلطيخ المناعي ل LL37 ، والقياس الكمي للنشاط الأنزيمي (NE و CG و MPO).
Access restricted. Please log in or start a trial to view this content.
Protocol
تم الحصول على عينات الدم كتبرعات من المشاركين الأصحاء سريريا بعد الموافقة المستنيرة. تم إجراء جميع التجارب بإذن من لجنة أخلاقيات البحوث البشرية التابعة لكلية العلوم الكيميائية الحيوية ، جامعة أوتونوما "بينيتو خواريز" في أواكساكا.
ملاحظة: كانت معايير التضمين في الدراسة غير واضحة الجنس والعمر ، وصحية سريريا وفقا لردود المشاركين على استبيان قبل أخذ عينة دم. تم إجراء تحليل دموي لتحديد عدد الخلايا واستبعاد العدوى أو فقر الدم ، بالإضافة إلى اختبار البروتين التفاعلي C لاستبعاد الالتهاب في المتبرع.
1. جمع الدم المحيطي والحصول على حزمة كرات الدم الحمراء والكريات البيض
- اجمع 10 مل من الدم المحيطي عن طريق بزل الوريد في أنابيب تحتوي على 1.8 مجم / مل من K2 · EDTA كمضاد للتخثر (انظر جدول المواد) من الأفراد الأصحاء سريريا بعد الحصول على الموافقة المستنيرة. بعد ذلك ، قم بإجراء القياس الحيوي القياسي للدم واختبار البروتين التفاعلي C لاستبعاد العدوى أو الالتهاب ، مما يضمن جودة العينة.
- جهاز طرد مركزي لعينة الدم المحيطية عند 82 × جم لمدة 15 دقيقة لإزالة البلازما الغنية بالصفائح الدموية ، تليها عملية طرد مركزي ثانية عند 630 × جم لمدة 5 دقائق. تجاهل البلازما المتبقية للحصول على حزمة كريات الدم الحمراء والكريات البيض.
- قم بتخفيفه بنسبة 1: 1 (v / v) باستخدام محلول ملحي مخزن بالفوسفات من Dulbecco (DPBS).
2. تنقية العدلات متعددة الأشكال النووية (PMN) باستخدام تدرج مزدوج الكثافة
ملاحظة: قم بإجراء تنقية العدلات مباشرة بعد جمع الدم ، لأن عمرها المختبري محدود يبلغ حوالي 8 ساعات.
- قم بإيداع ما يلي في أنبوب زجاجي معقم سعة 10 مل (انظر جدول المواد) بالترتيب: 1 مل من محلول كثافة 1.098 جم / مل ، 1 مل من محلول كثافة 1.079 جم / مل (انظر جدول المواد) ، ثم 4 مل من حزمة كريات الدم الحمراء المخففة والكريات البيض. صب على الجدران دون كسر التوتر السطحي بين الطبقات لمنعها من الاختلاط.
- جهاز طرد مركزي عند 320 × جم لمدة 20 دقيقة عند 4 درجات مئوية ، وتجنب التسارع / التباطؤ حتى لا تزعج القوى العالية لجهاز الطرد المركزي الانحدار.
- استنشاق المرحلة التي تتوافق مع الخلايا المحببة (الشكل 1 أ) عن طريق السحب ، ونقلها إلى أنبوب زجاجي معقم آخر سعة 10 مل. يغسل ب 4 مل من 1x DPBS عند 300 × جم لمدة 10 دقائق عند 4 درجات مئوية.
- تخلص من المادة الطافية وعالج الخلايا بالصدمة التناضحية لإزالة كريات الدم الحمراء المتبقية. أضف 4 مل من محلول ملحي 0.2٪ لمدة دقيقتين عند 4 درجات مئوية ، وأجهزة طرد مركزي عند 300 × جم لمدة 10 دقائق عند 4 درجات مئوية. تخلص من المادة الطافية. ثم أضف 4 مل من المحلول متساوي التوتر (0.65٪ محلول ملحي) لمدة 5 دقائق عند 4 درجات مئوية لاستعادة سلامة الغشاء ، وأجهزة الطرد المركزي عند 300 × جم لمدة 10 دقائق عند 4 درجات مئوية.
ملاحظة: المحلول الملحي 0.2٪ هو وسط منخفض التوتر ، مع تركيز مذاب أقل بالنسبة إلى تركيز الوسط داخل الخلايا RBC. يسمح التلامس مع hypotonicmedium بانتشار الماء في كرات الدم الحمراء ، مما يؤدي إلى تورمها وانحلال الدم. تم تأكيد إزالة كرات الدم الحمراء من الطافي من خلال الملاحظة المجهرية. - إزالة طاف . أعد تعليق الخلايا في 4 مل من 1x DPBS لإزالة الحطام الخلوي ، ثم أجهزة الطرد المركزي عند 300 × جم لمدة 10 دقائق عند 4 درجات مئوية. أخيرا ، أعد تعليق حبيبات الخلية في 2 مل من محلول الملح المتوازن البارد من هانك (HBSS).
3. مورفولوجيا العدلات وصلاحيتها (الشكل 1 ب)
- اختبار استبعاد التريبان الأزرق
- تمييع 5 ميكرولتر من تعليق الخلية في 20 ميكرولتر من 0.4 ٪ التربيق الأزرق (نسبة 1: 5). عد الخلايا في حجرة نيوباور وحدد صلاحية الخلية باستخدام اختبار الاستبعاد. ضع في اعتبارك الخلايا التي تحافظ على سلامة غشائها دون اختراق الصبغة على أنها قابلة للحياة.
- جبل 5 ميكرولتر من تعليق الخلية على شريحة ؛ تجف وصمة عار مع وصمة عار رايت لمدة 15 ثانية. قم بإصلاح العينة على الفور باستخدام درجة الحموضة 6.4 العازلة للفوسفات لمدة 30 ثانية. تغسل مع كمية كافية من الماء المقطر ومراقبة مورفولوجيا تحت المجهر الضوئي (100x).
- 7AAD- تلطيخ وتحليل التدفق الخلوي
- أضف 1 × 105 خلايا لتدفق أنابيب القياس الخلوي ، وقم بتلطيخها ب 1 ميكرولتر من 7AAD في 100 ميكرولتر من المخزن المؤقت FACS (1x DPBS ، 0.1٪ أزيد الصوديوم ، و 10٪ بلازما ذاتية المكتمل) لمدة 15 دقيقة عند 4 درجات مئوية في الظلام.
- يغسل مع 500 ميكرولتر من المخزن المؤقت FACS في 300 × غرام لمدة 10 دقائق. ثبت الخلايا ب 500 ميكرولتر من 2٪ بارافورمالدهيد ، وقم بتخزينها عند 4 درجات مئوية حتى تحليلها في مقياس التدفق الخلوي.
- للتحكم في الخلايا الميتة ، قم بإصلاح 1 × 105 خلايا مع 200 ميكرولتر من 4٪ بارافورمالدهيد لمدة 30 دقيقة ، واغسلها ب 500 ميكرولتر من 1x PBS عند 300 × جم لمدة 10 دقائق عند 4 درجات مئوية. اسحب الطافية وتخلص منها. ثم أضف 200 ميكرولتر من 0.1٪ Triton X-100 لمدة 1 ساعة عند 4 درجات مئوية. يغسل مع 500 ميكرولتر من 1x PBS وصمة عار مع 7AAD كما في الخطوة 3.2.1.
- باستخدام مقياس التدفق الخلوي (انظر جدول المواد) ، قم بإجراء تحليل FSC مقابل SSC لتحليل نقاء الخلية و SSC مقابل تلطيخ 7AAD لتحليل صلاحية الخلية. اقرأ 3 × 104 أحداث في 100 ميكرولتر من حجم الامتصاص عند التدفق المتوسط (1000 خلية / ثانية) في الإعدادات متعددة الأشكال النووية (FSC ، 400-490 و SSC ، 300-320).
- تحليل البيانات التي تم التقاطها في برنامج مقياس التدفق الخلوي (انظر جدول المواد) ، وتحديد النسبة المئوية للنقاء والخلايا الإيجابية ل 7AAD في السكان متعددي الأشكال النووية ، المقدمة من خلال المخططات النقطية والرسوم البيانية.
4. تلطيخ CFSE من الكائنات الحية الدقيقة
- أضف 1 × 108 بكتيريا أو 1 × 106 خيوط كاذبة فطرية في أنابيب دقيقة سعة 1.5 مل ، وصمة عار مع 200 ميكرولتر من 5 ميكرومتر كربوكسي فلوريسئين سكسينيميديل إستر (CFSE) المذاب في 1x PBS. تخلط لبضع ثوان ، وتحتضن عند 37 درجة مئوية لمدة 10 دقائق في الظلام.
- أوقف التفاعل بإضافة 500 ميكرولتر من البلازما غير المكتملة ، وأجهزة الطرد المركزي عند 620 × جم لمدة 10 دقائق للخيوط الكاذبة أو عند 1800 × جم لمدة 10 دقائق للبكتيريا.
- تخلص من المواد الطافية واغسل الكريات ب 1 مل من 1x PBS مع الطرد المركزي كما في الخطوة 4.2. أخيرا ، أعد تعليق الكائنات الحية الدقيقة في 250 ميكرولتر من 1x PBS.
- تحضير 50 ميكرولتر من القسمة في أنابيب دقيقة سعة 1.5 مل مع 2 × 107 بكتيريا (MOI: 100) أو 2 × 105 pseudohyphae (MOI: 1) للحث الصافي.
5. صافي الحث
- ضع أغطية زجاجية معقمة مقاس 10 مم × 10 مم في لوحة من 24 بئرا وقم بتغطيتها ب 10 ميكرولتر من 0.001٪ poly-L-lysine لمدة ساعة واحدة في درجة حرارة الغرفة. اغسل مرتين ب 100 ميكرولتر من 1x PBS ، وجفف في الهواء ، وقم بالإشعاع بضوء الأشعة فوق البنفسجية لمدة 15 دقيقة.
- استبدل محلول HBSS لتعليق العدلات في الخطوة 2.5 بوسط RPMI 1640 المكمل ببلازما ذاتية المنشأ بنسبة 10٪. إلى لوحة 24 بئر (الخطوة 5.1) ، أضف 350 ميكرولتر من معلق الخلية هذا ، للحصول على تركيز نهائي قدره 2 × 105 عدلات / بئر.
- اسمح للخلايا بالالتصاق بقاع الآبار عن طريق الحضانة لمدة 20 دقيقة عند 37 درجة مئوية مع 5٪ CO2.
- أضف المحفزات للحث على تكوين NET في 50 ميكرولتر: المحفزات الميكروبية - بكتيريا إيجابية الجرام S. aureus (ATCC 25923) ، والبكتيريا سالبة الجرام P. aeruginosa (ATCC 10145) في MOI 100 ، و pseudohyphae من C. albicans (ATCC 10231) في MOI: 1 ؛ المحفزات الكيميائية الحيوية-PMA (200 نانومتر) و HOCl (4.5 mM) ، والتحكم مع غياب التحفيز (50 ميكرولتر من HBSS).
- الحصول على حجم نهائي قدره 400 ميكرولتر لكل بئر. تخلط على لوحة شاكر عند 140 دورة في الدقيقة لمدة 30 ثانية ، وتحتضن لمدة 4 ساعات عند 37 درجة مئوية و 5٪ CO2.
6. تصور الشبكات بواسطة الفحص المجهري الفلوري
- الحمض النووي و LL37 المناعي
- بعد الحث الصافي ، قم بإزالة المواد الطافية من الآبار عن طريق السحب بعناية ، وقم بإصلاح الخلايا ب 300 ميكرولتر من 4٪ بارافورمالدهيد لمدة 30 دقيقة.
- اغسل الخلايا ب 200 ميكرولتر من 1x PBS بدون طرد مركزي ، وأضف 200 ميكرولتر من المخزن المؤقت للحجب (10٪ بلازما غير مكتملة في 1x PBS) لمدة 30 دقيقة.
- بالنسبة لصبغة LL-37 ، قم باختراق الخلايا ب 200 ميكرولتر من 0.2٪ Triton X-100 في 1x PBS لمدة 10 دقائق للسماح للجسم المضاد بدخول الخلايا. اغسل 2x بعناية باستخدام 1x PBS لإزالة المنظف الزائد.
- قم بتركيب أغطية الأغطية على شرائح زجاجية (أربعة أغطية على كل شريحة). يقوم الحمض النووي بتلطيخ الخلايا ب 2 ميكرولتر من DAPI (انظر جدول المواد) ، وختم أغطية الغطاء ، وتخزينها عند -20 درجة مئوية حتى تحليلها بواسطة الفحص المجهري الفلوري متحد البؤر.
- اقتناء وتحليل الصور الفلورية
- التقط صور NET لتحديد مكوناتها ، واستخدم المرشحات المقابلة في مجهر التألق متحد البؤر (انظر جدول المواد) للحصول على الصور باستخدام برنامج الكمبيوتر.
ملاحظة: ضع في اعتبارك أن الحمض النووي ملطخ ب DAPI (اللون الأزرق) ، مما يدل على الإثارة عند 360 نانومتر والانبعاث عند 460 نانومتر. الكائنات الحية الدقيقة ملطخة ب CFSE (اللون الأخضر) ، الذي يبلغ إثارته 492 نانومتر وانبعاث 521 نانومتر. يتم تمييز الببتيد LL37 بجسم مضاد ل LL37 Alexa Fluor 594 (اللون الأحمر) ، والذي يبلغ إثارته 594 نانومتر وانبعاث 614 نانومتر. - معايرة المجهر. ضع الشريحة والتركيز البؤري باستخدام تباين التداخل التفاضلي (DIC) مع تشغيل الضوء العادي. اختر مباشر لعرض الصورة على الشاشة.
- أطفئ الضوء وحدد القناة المقابلة للفلوروكروم. على سبيل المثال ، حدد عامل التصفية 365 نانومتر / أزرق ل DAPI ، أو 43 HE DsRed ل Alexa 594 ، أو 38 HE GFP ل CFSE.
- اضبط الإعدادات باستخدام الجسم المضاد للتحكم في النمط المتماثل ل LL37 والخلايا غير الملوثة ل DAPI و CFSE. اضبط نفس وقت التعريض الضوئي والجهد والتباين وإعدادات العدسة لالتقاط جميع الصور في نفس الظروف.
ملاحظة: في هذه الدراسة ، تم ضبط وقت التعرض والجهد والتباين على 1.0 مللي ثانية و 4.0 فولت و 0.0 على التوالي ، بهدف 40x. يمكن ضبط هذه القيم لتسهيل أفضل التقاط للصور للعينات. - حدد Snap لالتقاط الصورة. احفظ خمس صور (أربعة أطراف والمركز) لكل بئر ، والتوطين المشترك (دمج) الحمض النووي / LL37 / CFSE.
- حدد الفئات الثلاث من وحدات البكسل كخلفية مع الصور المستقلة لكل لون وقم بتحليل قيمة متوسط الإشارة الرمادية لكل منطقة باستخدام برنامج Image J.
- التقط صور NET لتحديد مكوناتها ، واستخدم المرشحات المقابلة في مجهر التألق متحد البؤر (انظر جدول المواد) للحصول على الصور باستخدام برنامج الكمبيوتر.
7. قياس النشاط الأنزيمي
- في لوحة 96 بئر ، أضف 90 ميكرولتر من معلق الخلية في HBSS التي تحتوي على 1 × 10 5 عدلات للتحريض الصافي ، واحتضانها لمدة 20 دقيقة عند 37 درجة مئوية و5 ٪ CO2.
- على الفور ، أضف 10 ميكرولتر من المحفزات المقابلة (التركيز كما في الخطوة 5.4) واحتضان لمدة 4 ساعات عند 37 درجة مئوية مع 5٪ CO2.
- تخلص من المواد الطافية واغسل الخلايا ب 100 ميكرولتر من HBSS. عالج ب 1 وحدة / مل من DNase لمدة 10 دقائق عند 37 درجة مئوية لصالح إطلاق هياكل بروتين الحمض النووي ، وأجهزة الطرد المركزي عند 1800 × جم لمدة 10 دقائق.
- استعادة المواد الطافية وتقييم نشاط الإنزيم في الطافي باستخدام التفاعلات اللونية كما وصفها سابقا White et al.17.
- حدد أقصى نشاط إنزيمي ل NE و CG و MPO في العدلات في ظل نفس الظروف التجريبية دون إضافة أي محفزات للحث الصافي. بعد ذلك ، قم بتجميد عينة الخلية عند -70 درجة مئوية وذوبان الجليد عند 37 درجة مئوية في حمام مائي ، مما يولد صدمة في درجة الحرارة لصالح إطلاق البروتينات داخل الخلية عن طريق تحلل الخلايا. جهاز طرد مركزي بسرعة 1800 × جم لمدة 10 دقائق واستعادة المواد الطافية.
- أضف 50 ميكرولتر من المادة الطافية إلى كل بئر في 96 لوحة بئر ، ثم أضف 50 ميكرولتر من كل ركيزة كما هو موضح في الخطوة 7.7.
- أضف 0.5 M من N-ميثوكسي سكسينيل-Ala-Ala-Pro-Val-p-nitro aniline كركيزة ل NE ، و 1 mM من N-succinyl-Ala-Ala-Pro-Phe-p-nitroanilide ل CG. احتضان لمدة 3 ساعات في درجة حرارة الغرفة. بالنسبة ل MPO ، أضف 1.6 مللي متر من 3,3 '، 5,5'-رباعي ميثيل بنزيدين (TMB) واحتضانه لمدة 30 دقيقة في درجة حرارة الغرفة.
- بعد الحضانة ، أضف 50 ميكرولتر من محلول التوقف (0.5 M H2SO4) ل MPO وقم بقياس الامتصاص عند 405 نانومتر ل NE و CG و 450 نانومتر ل MPO ، باستخدام مقياس الطيف الضوئي.
- قارن القيم التي تم الحصول عليها مع منحنيات المعايرة المقابلة وأظهر نتائج كل حالة بالنسبة إلى الحد الأقصى لنشاط الإنزيم (100٪).
8. التحليل الإحصائي
- تحليل بيانات القياس في ثلاث نسخ لكل تجربة مستقلة (ن = 10) وإجراء تحليل ANOVA للتحليل الإحصائي من خلال مقارنة المجموعات بمستوى ثقة 95٪.

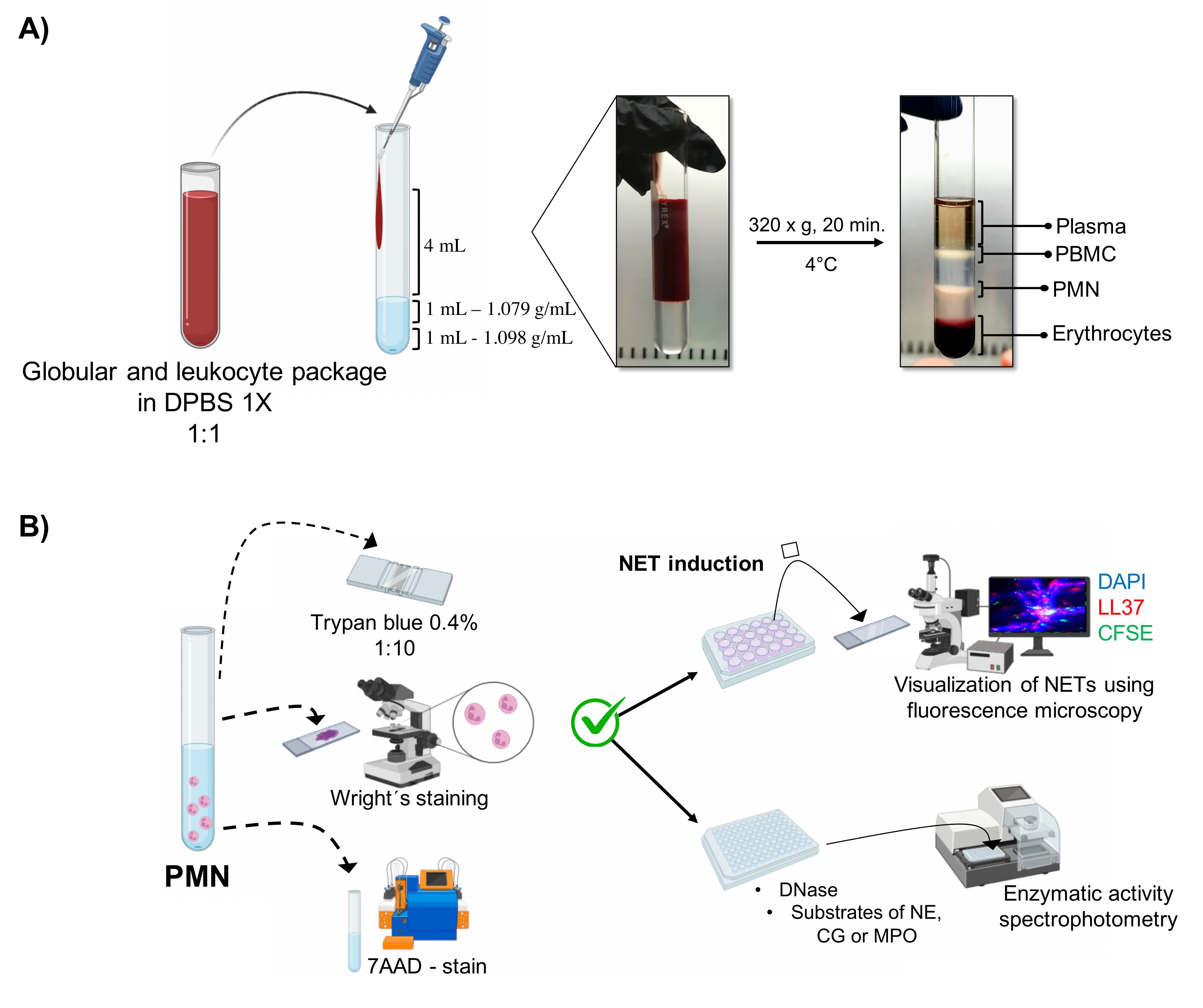
الشكل 1: تنقية PMN وبروتوكول الحث الصافي. (أ) تمت إزالة البلازما من الدم المحيطي للحصول على حزمة كريات الدم الحمراء والكريات البيض وتخفيفها 1: 1 (v / v) مع 1x DPBS. بعد ذلك ، تمت إضافة 4 مل من التخفيف على طول الجدار إلى أنبوب التدرج مزدوج الكثافة ، وتم طرده بالطرد المركزي عند 320 × جم لمدة 20 دقيقة عند 4 درجات مئوية ، والحصول على فصل طبقات الخلية المختلفة واستعادة الطبقة المقابلة ل PMN. (ب) تم عد الخلايا النقية، وحللت مورفولوجيتها بواسطة تلطيخ رايت. تم تحديد الجدوى من خلال استبعاد تريبان الأزرق وتلطيخ 7AAD باستخدام قياس التدفق الخلوي. بمجرد التحقق من نقاء العدلات المثلى وقابليتها للحياة ، تم تحفيز تكوين NET عن طريق إضافة الميكروبات (S. aureus و P. aeruginosa و C. albicans) أو المواد الكيميائية (PMA و HOCl) في 24 لوحة بئر للتحليل بواسطة الفحص المجهري الفلوري باستخدام DAPI-DNA و anti-LL37 Alexa Fluor 594 وتلطيخ الكائنات الحية الدقيقة CFSE. من أجل تحديد كمية الإنزيم ، تم تحفيز NETs في 96 صفيحة بئر لمدة 3 ساعات وعولجت ب DNase ، تليها إضافة ركائز لكل إنزيم: NE و CG و MPO. تم قياس تغيرات اللون بواسطة القياس الطيفي. DPBS = محلول ملحي مخزن بالفوسفات في دولبيكو ؛ PBMC = خلايا الدم أحادية النواة المحيطية ؛ PMN = العدلات متعددة الأشكال النووية ؛ NE = إيلاستاز العدلات ؛ CG = كاثيبسين G ؛ MPO = الميلوبيروكسيداز. PMA = خلات فيربول ميريستات ؛ HOCl = حمض هيبوكلوروس. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
النتائج
نقاء وصلاحية العدلات
يتم تصور المراحل الخلوية الديناميكية في الأنبوب من تنقية التدرج مزدوج الكثافة. داخل هذه الطبقات ، تكون الطبقة المقابلة للخلايا المحببة أعلى من طبقة الكثافة 1.079 جم / مل ، والتي تتميز عن مراحل الخلايا وحيدة الدم المحيطية (PBMCs) وكريات الدم الحمراء (ا?...
Access restricted. Please log in or start a trial to view this content.
Discussion
يجب الحصول على مجموعة عالية النقاء من العدلات القابلة للحياة للحث على إطلاق الشبكات لأن هذه الخلايا لها عمر محدود خارج الجسم الحي يبلغ 8 ساعات في المتوسط ، وهي فترة يجب خلالها إجراء جميع التجارب. تحقيقا لهذه الغاية ، فإن المنهجية المثالية هي التدرج مزدوج الكثافة لتحسين وقت التنقية عن ...
Access restricted. Please log in or start a trial to view this content.
Disclosures
يعلن المؤلفون أنه ليس لديهم تضارب في المصالح.
Acknowledgements
تم دعم هذا العمل من خلال منحة العلوم الأساسية (# 285480) من CONACyT وقسم أبحاث المناعة السريرية في كلية العلوم الكيميائية الحيوية ، جامعة أوتونوما "بينيتو خواريز" دي أواكساكا. A.A.A، S.A.S.L، و W.J.R.R. لديها زمالات الدكتوراه من أرقام CONACyT # 799779 و # 660793 و # 827788 ، على التوالي.
Access restricted. Please log in or start a trial to view this content.
Materials
| Name | Company | Catalog Number | Comments |
| 24 Well plate for cell culture | Corning | 3526 | |
| 7-aminoactinomycin D (7-AAD) | BD Pharmingen | 51-668981E | |
| 96 Well plate for cell culture | Costar | 3596 | Flat bottom |
| Agitator | CRM Globe | CRM-OS1 | |
| Antibody LL37 | Santa Cruz Biotechnology | sc-166770 | |
| Blood collection tubes | BD VACUTAINER | 368171 | K2 EDTA 7.2 mg |
| Carboxyfluorescein succinimidyl ester (CFSE) | Sigma-Aldrich | 21878 | |
| Centrifuge | Hettich | 1406-01 | |
| Coverslip | Madesa | M03-CUB-22X22 | 22 mm x 22 mm |
| Dulbecco´s phosphate-buffered saline (DPBS) | Caisson | 1201022 | |
| Falcon tubes 50 mL | CORNING | 430829 | |
| Flow Cytometry Tubes | Miltenyi Biotec | 5 mL - Without caps | |
| FlowJo Software | BD Biosciences | Analyze flow cytometry data | |
| Fluorescence microscope | DM 2000 | LEICA | |
| Fluoroshield with DAPI | Sigma-Aldrich | F6057 | |
| Incubator | NUAIRE | UN-4750 | |
| MACSQuant Analyzer | Miltenyi Biotec | Flow cytometer | |
| Microplate reader photometer | Clarkson Laboratory - CL | ||
| Microtubes 1.5 mL | Zhejiang Runlab Tech | 35200N | wire snap |
| Minitab Software | Minitab | Statistical analysis | |
| Needles | BD VACUTAINER | 301746 | Diameter 1.34 mm |
| Optical microscope | VELAB | VE-B50 | |
| Percoll | GE Healthcare | 17-0891-01 | Solution for density gradient |
| Phosphate Buffered Saline (10x) | Caisson | PBL07-500ML | |
| Pyrex culture tubes | CORNING | CLS982025 | N°9820 |
| RPMI 1640 1x | Corning | 10-104-CV | contains Glutagro |
| Slides | Madesa | PDI257550 | 22 mm x 75 mm |
| Trypan Blue solution 0.4% | SIGMA | T8154-100ML |
References
- De Buhr, N., Maren, K. B. How neutrophil extracellular traps become visible. Journal of Immunology Research. 2016, 4604713(2016).
- Karlsson, A., Nixon, J. B., McPhail, L. C. Phorbol myristate acetate induces neutrophil NADPH-oxidase activity by two separate signal transduction pathways: dependent or independent of phosphatidylinositol 3-kinase. Journal of Leukocyte Biology. 67 (3), 396-404 (2000).
- Takei, H., Araki, A., Watanabe, H., Ichinose, A., Sendo, F. Rapid killing of human neutrophils by the potent activator phorbol 12-myristate 13-acetate (PMA) accompanied by changes different from typical apoptosis or necrosis. Journal of Leukocyte Biology. 59 (2), 229-240 (1996).
- Brinkmann, V., et al. Neutrophil extracellular traps kill bacteria. Science. 303 (5663), 1532-1535 (2004).
- Kenny, E. F., et al. Diverse stimuli engage different neutrophil extracellular trap pathways. eLife. 6, 24437(2017).
- Schultz, B. M., Acevedo, O. A., Kalergis, A. M., Bueno, S. M. Role of extracellular trap release during bacterial and viral infection. Frontiers in Microbiology. 13, 798853(2022).
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nature Reviews Immunology. 18 (2), 134-147 (2018).
- Delgado-Rizo, V., et al. Neutrophil extracellular traps and its implications in inflammation: An overview. Frontiers in Immunology. 8, 81(2017).
- Petretto, A., et al. Neutrophil extracellular traps (NET) induced by different stimuli: A comparative proteomic analysis. PLoS One. 14 (7), 0218946(2019).
- Neumann, A., et al. Novel role of the antimicrobial peptide LL-37 in the protection of neutrophil extracellular traps against degradation by bacterial nucleases. Journal of Innate Immunity. 6 (6), 860-868 (2014).
- Hakkim, A., et al. Activation of the Raf-MEK-ERK pathway is required for neutrophil extracellular trap formation. Nature Chemical Biology. 7 (2), 75-77 (2011).
- Sabbatini, M., Magnelli, V., Renò, F. NETosis in wound healing: When enough Is enough. Cells. 10 (3), 494(2021).
- Metzler, K. D., Goosmann, C., Lubojemska, A., Zychlinsky, A., Papayannopoulos, V. A myeloperoxidase-containing complex regulates neutrophil elastase release and actin dynamics during NETosis. Cell Reports. 8 (3), 883-896 (2014).
- Clark, S. R., et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nature Medicine. 13 (4), 463-469 (2007).
- Pilsczek, F. H., et al. A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to Staphylococcus aureus. Journal of Immunology. 185 (12), 7413-7425 (2010).
- Sosa, S. A., et al. Structural differences of neutrophil extracellular traps induced by biochemical and microbiologic stimuli under healthy and autoimmune milieus. Immunologic Research. 69 (3), 264-274 (2021).
- White, P. C., et al. Characterization, quantification, and visualization of neutrophil extracellular traps. Methods in Molecular Biology. 1537, 481-497 (2017).
- Boeltz, S., et al. To NET or not to NET: current opinions and state of the science regarding the formation of neutrophil extracellular traps. Cell Death and Differentiation. 26 (3), 395-408 (2019).
- Yousefi, S., Mihalache, C., Kozlowski, E., Schmid, I., Simon, H. U. Viable neutrophils release mitochondrial DNA to form neutrophil extracellular traps. Cell Death and Differentiation. 16 (11), 1438-1444 (2009).
- Brinkmann, V., Zychlinsky, A. Neutrophil extracellular traps: is immunity the second function of chromatin. The Journal of Cell Biology. 198 (5), 773-783 (2012).
Access restricted. Please log in or start a trial to view this content.
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved