
Method Article
Polarización de células y análisis de células y análisis derivados de monocitos humanos M1 y M2 con citometría de flujo sobre la infección por mycobacterium tuberculosis
En este artículo
Erratum Notice
Resumen
Este protocolo proporciona un método para estudiar la infección por Mycobacterium tuberculosis en macrófagos polarizados M1 o M2 humanos basados en la diferenciación de monocitos de sangre periférica a células similares al macrófago que están infectadas con la cepa virulenta H37Rv etiquetada por GFP, y analizada con citometría de flujo utilizando un panel de 10 colores que incluye la expresión de marcadores M1/M2 seleccionados.
Resumen
Los macrófagos humanos son células huésped primarias de la infección intracelular de Mycobacterium tuberculosis (Mtb) y, por lo tanto, tienen un papel central en el control inmunológico de la tuberculosis (TB). Hemos establecido un protocolo experimental para seguir la polarización inmune de las células derivadas del mieloide en células similares al macrófago M1 (activadas clásicamente) o M2 (alternativamente activadas) a través de la evaluación con un panel de citometría de flujo de 10 colores que permite visualizar y caracterizar profundamente la Mtb etiquetada con proteína verde fluorescente (GFP) en diversos subconjuntos de macrófagos. Los monocitos obtenidos de donantes sanguíneos sanos se polarizaron en células M1 o M2 utilizando la diferenciación con el factor estimulador de colonias de macrófago de granulocito (GM-CSF) o factor estimulante de colonias de macrófago (M-CSF) seguido de polarización con IFN-γ y lipopolisacárido (LPS) o IL-4, respectivamente. Las células M1 y M2 totalmente polarizadas fueron infectadas con Mtb-GFP durante 4 horas antes de que los macrófagos separados infectados por Mtb se mancharan con citometría de flujo a las 4 o 24 horas posteriores a la infección. La adquisición de muestras se realizó con citometría de flujo y los datos se analizaron mediante un software de análisis de citometría de flujo. Se realizó un cálculo manual, así como una reducción de la dimensionalidad con aproximación y proyección uniforme del colector (UMAP) y análisis de fenógrafos. Este protocolo dio lugar a una polarización efectiva M1/M2 caracterizada por niveles elevados de células CD64, CD86, TLR2, HLA-DR y CCR7 en células M1 no infectadas, mientras que las células M2 no infectadas mostraron una fuerte regulación de los marcadores de fenotipo M2 CD163, CD200R, CD206 y CD80. Las células polarizadas M1 normalmente contenían menos bacterias en comparación con las células polarizadas por M2. Varios marcadores M1/M2 fueron regulados después de la infección por Mtb, lo que sugiere que Mtb puede modular la polarización del macrófago. Además, se encontraron 24 grupos celulares diferentes de diferentes tamaños que se distribuyeron de forma única entre las células infectadas por M1 y M2 y Mtb a las 24 horas posteriores a la infección. Este protocolo de citometría de flujo M1/M2 podría utilizarse como columna vertebral en la investigación de Mtb-macrofáfago y adoptarse para necesidades especiales en diferentes áreas de investigación.
Introducción
Los macrófagos son células inmunes que contribuyen significativamente a la regulación de la homeostasis tisular, inflamación y patologías de la enfermedad. Al ser un componente esencial de la inmunidad innata, el linaje monocito-macrófago de las células expresa fenotipos heterogéneos en respuesta a señales ambientales alteradas, que reflejan su plasticidad y adaptación a diferentes lugares anatómicos e inmunológicos1. Dependiendo de los factores de crecimiento, citoquinas y otros mediadores presentes en el microambiente, los macrófagos se han categorizado en dos grandes poblaciones reversibles, cada una con un papel diferente en el control bacteriano y el aclaramiento2:los macrófagos polarizados M1 proinflamatorios y activados clásicamente M1 y los macrófagos antiinflamatorios y alternativamente activados con M2 polarizados que fueron nombrados originalmente para imitar la nomenclaturacelular T helper (Th). Esta agrupación de macrófagos polarizados inmunes a menudo se considera simplista, ya que la activación y diferenciación del macrófago no es lineal, sino que se ilustra con mayor precisión como un continuo donde cada población tiene diferentes características y roles funcionales en el resultado del desarrollo y progresión de la enfermedad4,5,6,7. Sin embargo, hay numerosas ventajas experimentales con el modelo de macrófago M1/M2 que se puede utilizar en varios campos diferentes de la investigación.
Mycobacterium tuberculosis (Mtb) es el agente causal de la tuberculosis (TB) y se ha estimado que infecta a una persona cada segundo y se considera el agente infeccioso único más letal del mundo (Informe mundial sobre la tuberculosis 2019). Dado que las vías respiratorias son la vía principal de la infección por Mtb, los macrófagos alveolares son las células huésped preferidas para infectarse con Mtb y representan tanto las barreras primarias como el reservorio infeccioso de Mtb en los pulmones. La polarización del macrófago en respuesta a diferentes estímulos se ha estudiado ampliamente a lo largo de los años7 y en la mayor parte del trabajo publicado, La polarización M1 de cultivos de monocitos in vitro es inducida por factor estimulante de colonias de granulocito-macrófago (GM-CSF) junto con IFN-γ y LPS8,9,mientras que la polarización M2 es inducida con factor estimulante de colonia de macrófago (M-CSF) e IL-410,11. Los macrófagos M1 son potentes células efectoras que median las respuestas antimicrobianas contra patógenos intracelulares y tienen un papel esencial en la inmunidad antitumoral12. Los macrófagos M2, por otro lado, tienen una función antiinflamatoria, alta capacidad fagocítica y están involucrados principalmente en la cicatrización de heridas y reparación de tejidos, así como en las infecciones del parásito12. En consecuencia, los macrófagos M1 se consideran más eficaces en el control intracelular de Mtb en comparación con los macrófagos M213. Sin embargo, las bacterias Mtb también tienen el potencial de modular la polarización del macrófago para subvertir la inmunidad innata14,15,16,17.
Si bien es común generar macrófagos a partir de la diferenciación de monocitos obtenidos de sangre periférica18,también se podrían generar macrófagos a partir de células madre pluripotentes inducidas (iPSC)19 o de macrófagos derivados de médula ósea de ratones20,21. Estas son técnicas factibles para estudiar células macrofágenas primarias obtenidas de progenitores monocitos/macrófago que proliferan y diferenciarán en una población homogénea de células maduras similares al macrófago. Sin embargo, estos protocolos rara vez proporcionan un conocimiento más profundo sobre el fenotipo y la función de las células obtenidas ni explican la heterogeneidad natural observada entre los macrófagos obtenidos in vivo. Como Mtb es un patógeno humano estricto, también hay una ventaja para estudiar Mtb en sistemas de modelos humanizados. La citometría de flujo es una potente tecnología que ofrece la posibilidad de evaluar múltiples características fenotípicas y funcionales de células individuales en la suspensión22,algo que podría ser bastante desafiante con células adherentes como macrófagos que también se sabe que son autofluorescentes23,24. Además del desprendimiento químico de macrófagos firmemente adherentes, la infección por Mtb puede representar un factor de estrés significativo para las células que añade otro nivel de complejidad en los análisis citométricos de flujo de macrófagos infectados con Mtb.
En este protocolo experimental, hemos utilizado un modelo de infección por macrófago humano previamente establecido basado en la polarización inmune de las células primarias derivadas de periféricos y monocitos sanguíneos que están infectadas con la virulenta cepa Mtb de laboratorio H37Rv, y analizadas con citometría de flujo utilizando un panel de 10 colores que incluye la expresión de marcadores M1 y M2 seleccionados25. Este protocolo proporciona un método eficiente y reproducible para estudiar las respuestas a la infección por Mtb en macrófagos polarizados de monocitoS M1 o M2. Además, el uso de citometría de flujo en macrófagos infectados por Mtb adherentes nos permite estudiar una variedad de marcadores de superficie asociados con macrófagos M1 y M2 convencionales y su respuesta longitudinal a la infección por Mtb. Es importante destacar que este protocolo puede adoptarse fácilmente para la investigación de infecciones con otros patógenos, en estudios antitumorales o en estudios de enfermedades inflamatorias, para la detección de fármacos, etc. y también podría ser explotado para la evaluación de la polarización del macrófago M1/M2 en muestras clínicas humanas.
Protocolo
La sangre periférica humana de donantes de sangre anónimos sanos se obtuvo del banco de sangre del Hospital Universitario Karolinska, Huddinge, Suecia (aprobación ética Dnr 2010/603-31/4). Todos los pasos experimentales que involucran mtb virulento vivo se realizaron en el laboratorio de Bioseguridad Nivel-3 (BSL-3) de la Agencia de Salud Pública de Suecia (FOHM), Solna, Suecia.
1. Preparación de medios, tampones y cultivos bacterianos
NOTA: Los detalles sobre todos los reactivos y consumibles se proporcionan en la Tabla de Materiales.
- Medio completo RPMI: Suplemento RPMI 1640 con piruvato de sodio de 1 mM, 2 mM L-glutamina, 10 mM HEPES, y 10% suero bovino fetal (FBS) inactivado por calor. Evite los antibióticos en el medio de cultivo celular cuando trabaje con la infección por Mtb.
- Medio RPMI libre de suero: Suplemento RPMI 1640 con piruvato sódico de 1 mM, 2 mM L-glutamina y 10 mM HEPES.
- Tampone de lavado: Prepare la solución salina del tó buffer de fosfato (PBS) que contenga 0,05% (v/v) Tween-80.
- Búfer FACS: Prepare PBS que contenga 2.5% (v/v) FBS y 0.5 mM EDTA.
- Búfer de fijación: Prepare PBS que contenga 4% de formaldehído a PBS. Asegúrese de que esté recién preparado antes de su uso, por ejemplo, mezclado a partir de una solución de stock de formaldehído del 37%.
- Amortiguador de permeabilización: Añadir 0.1% citrato de sodio y 0.1% Tritón X-100 a agua desionizada.
- Tampone de lavado (para inmunofluorescencia): Prepare PBS que contenga 0.1% BSA y 0.1% Tween-20.
- Búfer de bloqueo: Prepare PBS que contenga 0.1% BSA y 10% suero de cabra normal (NGS) a PBS.
- Tampone de tinción (para inmunofluorescencia): Prepare PBS que contenga 0.1% de BSA a PBS.
- Medio completo de TB: Suplemento de caldo Middle Brook 7H9 con 0.05% (v/v) Tween-80, 0.5% (v/v) glicerol, kanamycina (20 μg/mL), 10% (v/v) ácido oleico Middlebrook, albúmina, dextrosa y enriquecimiento de catalasa (Middlebrook OADC Enrichment).
- Cultivos bacterianos: Utilice la cepa estándar de laboratorio de Mtb virulenta, H37Rv, que expresa constitutivamente proteína fluorescente verde (GFP), para la infección de células derivadas de monocitos. Esta cepa de Mtb lleva un plásmido pFPV2 que contiene un gen que codifica GFP, así como un gen para la resistencia a la kanamicina. La resistencia a los antibióticos permite la selección continua de bacterias que expresan plásmidos en cultivos que contienen kanamicina. Almacene las bacterias en tb media completa y 70% glicerol (dilución 1:1) a -80 °C.
2. Aislamiento celular mononuclear de sangre periférica de abrigos buffy
NOTA: Realizar todo el trabajo con sangre humana (potencialmente contagiosa) dentro de un gabinete de bioseguridad de clase II. Inactivar los productos sanguíneos residuales con desinfectantes durante 15 minutos antes de desecharlos. La sangre se obtuvo de voluntarios sanos en este caso. Este protocolo de diferenciación de macrófago in vitro se creó para incluir 10 x 106 PBMCs aislados/donante/pozo. De cada donante, una capa buffy contiene alrededor de 50 ml de una suspensión concentrada de leucocitos originada de sangre entera, que normalmente proporciona 500-800 x 106 PBMCs de los cuales se pueden recuperar aproximadamente 10% o 50-80 x10 6 monocitos.
- Carga 15 ml de sangre de capa buffy encima de 15 ml de medio degradado de densidad preparado en tubo de 50 ml. Superponga lentamente la sangre en la parte superior de la capa de gradiente de densidad inclinando la punta de la pipeta a la pared del tubo.
- Gire los tubos a 600 x g durante 25 minutos a temperatura ambiente (RT) con 0 de aceleración y 0 de desaceleración.
NOTA: Cierre las tapas cuidadosamente antes de la centrifugación y compruebe siempre los soportes del tubo para un posible derrame después de la centrifugación. - Retire la capa superior de plasma con una pipeta pasteur estéril y, a partir de entonces, recoja cuidadosamente la capa celular mononuclear en un nuevo tubo de 50 ml utilizando una pipeta pasteur estéril. Añada el medio RPMI libre de suero al pellet PBMC para obtener un volumen final de 50 ml. Mezcle cuidadosamente invirtiendo el tubo unas cuantas veces antes de la centrifugación a 500 x g durante 5 minutos en RT.
- Deseche el sobrenadante cuidadosamente y vuelva a gastar el pellet de celda volteando la parte inferior del tubo dentro de los dedos.
- Para eliminar la contaminación media del gradiente de densidad de los PBMCs, lave las células 2-3 veces con RPMI sin suero para obtener un volumen final de 50 ml. Centrífuga a 500 x g durante 5 minutos en RT. Lave hasta que el sobrenadante celular se vuelva transparente.
- Deseche el sobrenadante y resuspend las células en 20 ml de medio RPMI libre de suero.
- Cuente las células por tinción azul trypan, manualmente usando un hemocítómetro o usando un contador de células automatizado. Diluir la suspensión celular en azul trypan en dilución 1:2 o 1:10 mezclando la muestra azul de cell-trypan en una placa de 96 pozo, por ejemplo, 50 μL + 50 μL (para el conteo de hemocitos) o 10 μL + 10 μL (para el recuento automatizado de células) y contar las células para obtener el número de células vivas / ml.
PRECAUCIÓN: El azul trypan es tóxico y debe desecharse en un residuo químico separado.
3. Diferenciación y polarización de células derivadas de monocitos
NOTA: Para la diferenciación y polarización de las células derivadas de monocitos, se siguió un protocolo que establecimos previamente para células similares a M0, M1 y M2, así como células polarizadas M1 y M2 completamente25. Para simplificar, aquí solo se describen macrófagos M1 y M2 totalmente polarizados.
- Utilice la adherencia plástica para el aislamiento de monocitos. Brevemente, semillas recién aisladas PBMCs en una placa de cultivo de 6 pozos a una concentración adecuada, por ejemplo, 10 x 106 PBMCs/pozo en 2 mL sin séricos medios RPMI e incubar a 37 °C y 5% CO2.
- Después de 2-3 h, retire las células no adherentes con una pipeta y lave los pozos 3 veces con 1 mL medio libre de suero. Las células unidas son monocitos y comprenden alrededor del 10% del total de PBMCs añadidos al pozo, es decir, 106 monocitos recuperados de 10 x 106 PBMCs añadidos por pozo.
- Para la diferenciación de macrófago, prepare una solución de trabajo que contenga 50 ng/mL GM-CSF o M-CSF para polarización de macrófago M1 y M2 respectivamente, añadida en 2 ml de medio completo RPMI por pozo. Cultiva las células en una incubadora de CO2 al 5% en 37 °C durante 3 días.
- El día 3, retire 1 ml del medio de cultivo celular cuidadosamente de la capa superior de cada pozo y complemente los cultivos celulares con 1 ml de medio completo RPMI fresco que contiene la doble concentración de M-CSF o GM-CSF para obtener 50 ng/mL de concentración final en los pozos. Añadir los factores de crecimiento en una solución de trabajo prefaseada de 100 ng/mL/well.
- En el día 6, añadir diferentes estímulos para los últimos 18-20 h de diferenciación celular para obtener M1 totalmente polarizado y maduro (interferón-γ; IFN-γ y lipopolisacárido; LPS (E. coli O55:B5)) o M2 (interleucina 4; IL-4) macrófagos. Para la polarización M1, prepare IFN-γ y LPS en el medio completo RPMI y agregue 50 μL por pozo para obtener una concentración final de 50 ng/mL IFN-γ y 10 ng/mL LPS en los cultivos celulares. Para la polarización M2, prepare IL-4 en el medio completo RPMI y agregue 50 μL por pozo para obtener una concentración final de 20 ng/mL en los cultivos celulares.
- Para la diferenciación de macrófagos polarizados M0, estimule las células únicamente con M-CSF, sin citoquinas adicionales (proporcionando un fenotipo similar a M2)25.
- Compruebe la morfología de cultivos celulares derivados de monocitos regularmente con microscopía ligera para asegurarse de que los monocitos más pequeños se diferencien en células más grandes similares al macrófago. Además, monitoree las posibles diferencias morfológicas entre la polarización M1 y M2, es decir, las células M1 alargadas y estiradas en comparación con las células M2 con una forma más redondeada25.
- El día 7, transfiera las placas con células derivadas de monocitos a un laboratorio BSL-3 para la infección con Mtb virulenta.
4. Preparación de cultivos Mtb
NOTA: Los siguientes pasos deben realizarse en una instalación de BSL-3. Para todos los trabajos con Mtb virulento, use ropa protectora, protección respiratoria y guantes resistentes al etanol.
- Descongelar un vial con 1 ml de alícuota bacteriana y mezclar con 9 ml de tb medio completo (dilución de 1:10) en un tubo de tapa filtrada de 50 ml. Cultivo de la suspensión en una incubadora a 37 °C y 5% CO2.
- Después de 24 h, gire la suspensión bacteriana a 2.300 x g durante 10 minutos y vierta cuidadosamente el medio. Resuspend el pellet bacteriano con 15-20 ml de tuberculosis fresca medio completo en un nuevo tubo de cultivo de tapa filtrada de 50 ml e incubar a 37 °C y 5% CO2. Mezcle las bacterias asentadas en el tubo cada 2-3 días para mantener un suministro homogéneo de nutrientes para todas las células bacterianas.
- Después de 7-10 días, mezcle la suspensión bacteriana correctamente pipeteando hacia arriba y hacia abajo antes de transferirse a un tubo de tapa de tornillo de 50 ml.
- Añadir 35-40 ml de tampón de lavado estéril al tubo de 50 ml y girar la suspensión bacteriana a 2.300 x g durante 10 minutos. Repita los pasos de lavado una vez. Resuspend el pellet bacteriano en 1 ml de medio RPMI libre de suero mediante pipeteo con un micropipette.
- Agregue otros 9 ml de medio RPMI libre de suero y sonice la suspensión bacteriana dentro de un armario de bioseguridad de clase II durante 5 minutos a 37 °C, para interrumpir los grumos bacterianos. Sumerja el tubo repetidamente (3-4 veces) en el sonicator del baño de agua para asegurar la máxima interrupción de los grupos bacterianos. Mida la densidad óptica (OD) de 1 ml de suspensión bacteriana a 600 nm de longitud de onda utilizando un espectrofotómetro colocado dentro del armario de bioseguridad. Utilice el medio RPMI sin suero para establecer la referencia.
- Calcule el número de unidades de formación de colonias (CFU) utilizando la fórmula: (OD+0.155)/0.161 = Y, y Y x 107= Y x 106 CFU/mL, por ejemplo, un valor OD 0.32 proporciona una concentración bacteriana de (0.32 + 0.155)/0.161 = 2.95, 2.95 x 107= 29.5 x 106 CFU/mL.
5. Infección por Mtb de células derivadas de monocitos
NOTA: Los siguientes pasos deben realizarse en una instalación de BSL-3.
- Resuspend el pellet bacteriano en medio RPMI libre de suero en un nuevo tubo estéril de 50 ml y ajustar la concentración bacteriana final a aproximadamente 5 x 106 CFU/ml.
- Retire el medio de cultivo celular de las 6 placas de pozo que contienen células derivadas de monocitos. Agregue 1 ml de medio RPMI libre de suero a cada pozo. Añadir 1 ml de suspensión bacteriana por pozo para obtener una multiplicidad de infección (MOI) 5:1, es decir, 5 x 106 CFU por 106 macrófagos en 2 ml/pozo e incubar las placas durante 4 h en 37 °C y 5% CO2.
- Después de la infección, lave las células 3 veces con 1 ml de tampón de lavado estéril para eliminar las bacterias extracelulares. Incline la placa y retire cuidadosamente todo el búfer de lavado de las esquinas. Resuspend las células derivadas de monocitos infectadas por Mtb en 2 ml de medio completo RPMI sin antibióticos y proceder a la tinción de la citometría de flujo o incubar las células durante otras 24 h (u otros puntos de tiempo) antes de la citometría de flujo.
6. Mancha de citometría de flujo de células derivadas de monocitos infectadas por Mtb
NOTA: Los siguientes pasos deben realizarse en una instalación de BSL-3. La tinción de citometría de flujo podría realizarse en una placa de 96 pozos en lugar de tubos.
- Separar las células infectadas por Mtb (y los controles no infectados) de los pozos de las 6 placas de pozo mediante incubación con 1 ml de tampón FACS por pozo durante al menos 30 min a 37 °C y 5% CO2.
- Pipetear suavemente hacia arriba y hacia abajo unas cuantas veces para asegurarse de que las células están separadas. Si es posible, confirme el desprendimiento de células con microscopía. Transfiera la suspensión celular de cada pozo a un tubo de microcentrífuga tapado con tornillo y gire los tubos a 200 x g durante 5 minutos. Deseche el sobrenadante cuidadosamente pipeteando.
- Lave el pellet celular en cada tubo dos veces con el búfer FACS y gire las celdas a 200 x g durante 5 minutos.
- Manchar las células (aproximadamente 0,5 x 106 a 1 x 106 células/tubo) con aproximadamente 50 μL cóctel de anticuerpos anticohumanos fluorocromáticos conjugados incluyendo TLR2 (AF647), CD206 (APC-Cy7), CD163 (BV605), CD80 (BV650), CCR7 (BV711), CD86 (BV786), CD200R (PE), CD64 (PE-Dazzle 594), HLA-DR (PE-Cy5)(Tabla 1)en combinación con el tinte de viabilidad Zombie-UV durante 30 minutos a 4 °C (refrigerador) en la oscuridad.
- Lave las células manchadas dos veces con 400 μL de tampón FACS y gire las células a 200 x g durante 5 minutos.
- Fijar las células manchadas con 200 μL de búfer de fijación (recién preparado) durante 30 minutos en RT en la oscuridad para asegurar la inactivación completa de las micobacterias.
- Lave las celdas dos veces con 400 μL de búfer FACS y gire a 200 x g durante 5 minutos para eliminar el exceso de búfer de fijación.
- Resuspend las células fijas en 400 μL de tampón FACS y transferir las muestras a nuevos tubos de microcentrífuga de 1 ml antes de sacarlas del laboratorio BSL-3 para la citometría de flujo en BSL-2. Almacene las celdas manchadas en +4 °C hasta la adquisición de muestras.
NOTA: Rocíe los tubos con un 70% de etanol antes de sacarlos del laboratorio BSL-3. El formaldehído es tóxico (cancerígeno) y debe manipularse en un armario de bioseguridad de clase II. Deseche los residuos de formaldehído en un residuo químico separado.
7. Adquisición y análisis de datos citométricos de flujo de células derivadas de monocitos infectadas por Mtb
NOTA: Los pasos 7.1–7.2 deben realizarse antes de la tinción de citometría de flujo descrita anteriormente. Para evitar problemas con el aglutinamiento celular y la disociación de tintes tándem después de la fijación celular, la adquisición de muestras de células infectadas por Mtb y no infectadas se realiza dentro de 4-10 h después de la tinción primaria de anticuerpos.
- Antes de la tinción de citometría de flujo descrita anteriormente, compense la señal fluorescente de cada anticuerpo fluorocromo-conjugado que aparece en el panel de tinción(Tabla 1)utilizando cuentas de compensación (tanto positivas como negativas).
- Valorar la dilución de anticuerpos para la tinción de macrófagos humanos para obtener la señal óptima para cada fluorocromo.
- Utilice células sin manchar para determinar el nivel de fluorescencia de fondo necesario para establecer la puerta para la población celular negativa, lo que permite visualizar las células manchadas (los macrófagos son altamente fluorescentes automáticos).
- Adquiera un mínimo de 50.000 células/muestra en el cytómetro de flujo utilizando el software recomendado para la adquisición de datos.
- Exporte los archivos de adquisición desde el cytómetro de flujo en formato estándar de citometría de flujo (FCS) 3.1.
- Analice los archivos FCS en el software de análisis de citometría de flujo.
- Los macrófagos de compuerta de acuerdo con sus características de dispersión hacia delante y hacia el lado (FSC y SSC) y excluyen las células muertas mediante gating de células vivas /muertas usando el tinte de viabilidad Zombie-UV.
- Visualice los macrófagos infectados con H37Rv-GFP en el canal FITC.
- Identificar la frecuencia de las células manchadas positivamente y la intensidad media geométrica de fluorescencia (MFI) para todos los marcadores (Tabla 1).
8. Tinción de inmunofluorescencia de células derivadas de monocitos infectadas por Mtb
NOTA: La infección por Mtb debe realizarse en una instalación de BSL-3.
- Para inmunostaining, semilla 2 x 105 PBMCs/pozo en 500 μL de medio RPMI libre de suero en 8 toboganes de cámara de pozo para obtener 2 x 104 monocitos/pozo. Después de la diferenciación y la polarización M1/M2 de monocitos, proceda con la infección por Mtb como se describió anteriormente. Corrija las diapositivas después de 24 h de infección por Mtb con búfer de fijación durante 30 min. Las diapositivas fijas se almacenan en el congelador a -20 °C hasta que se analicen más.
- Lave las células derivadas de monocitos dos veces con 200 μL de PBS durante 10 minutos cada una.
- Permeabilizar las células con 200 μL de amortiguador de permeabilización durante 5 min en RT.
- Lave las células 3 veces con 200 μL de PBS durante 5 minutos cada una.
- Lave las células dos veces con 200 μL de tampón de lavado durante 5 minutos cada una.
- Bloquee la unión no específica con 200 μL de búfer de bloqueo durante 30 minutos en RT.
- Diluir los anticuerpos primarios 1:100 en tampones de tinción e incubar las células M1 con un anticuerpo CD64 no conjugado (Clone: 10.1) y las células M2 con un anticuerpo CD163 no conjugado (policlonal) durante 2 h en RT.
- A continuación, lave las células 3 veces con 200 μL de tampón de lavado durante 10 minutos cada una.
- Diluir los anticuerpos secundarios etiquetados fluorescentes 1:1,000 en tampones de tinción e incubar las células M1 con un IgG-Alexa Fluor 594 anti-ratón y las células M2 con un anti-conejo IgG-Alexa Fluor 594 para 1 h en RT.
- Lave las células 3 veces con el búfer de lavado de 200 μL durante 10 minutos cada una.
- Retire la rejilla de la cámara y agregue ~20 μL de medio de montaje DAPI en cada pozo y coloque un tubo de cubierta de 1,5 mm en cada diapositiva.
- Sella el cubrebocas con una capa de esmalte de uñas.
- Adquirir imágenes utilizando un microscopio confocal con láseres que emiten a 486 nm para excitación de GFP (canal verde), 402 nm para DAPI (azul) y 560 nm para anticuerpo secundario (rojo) respectivamente.
Resultados
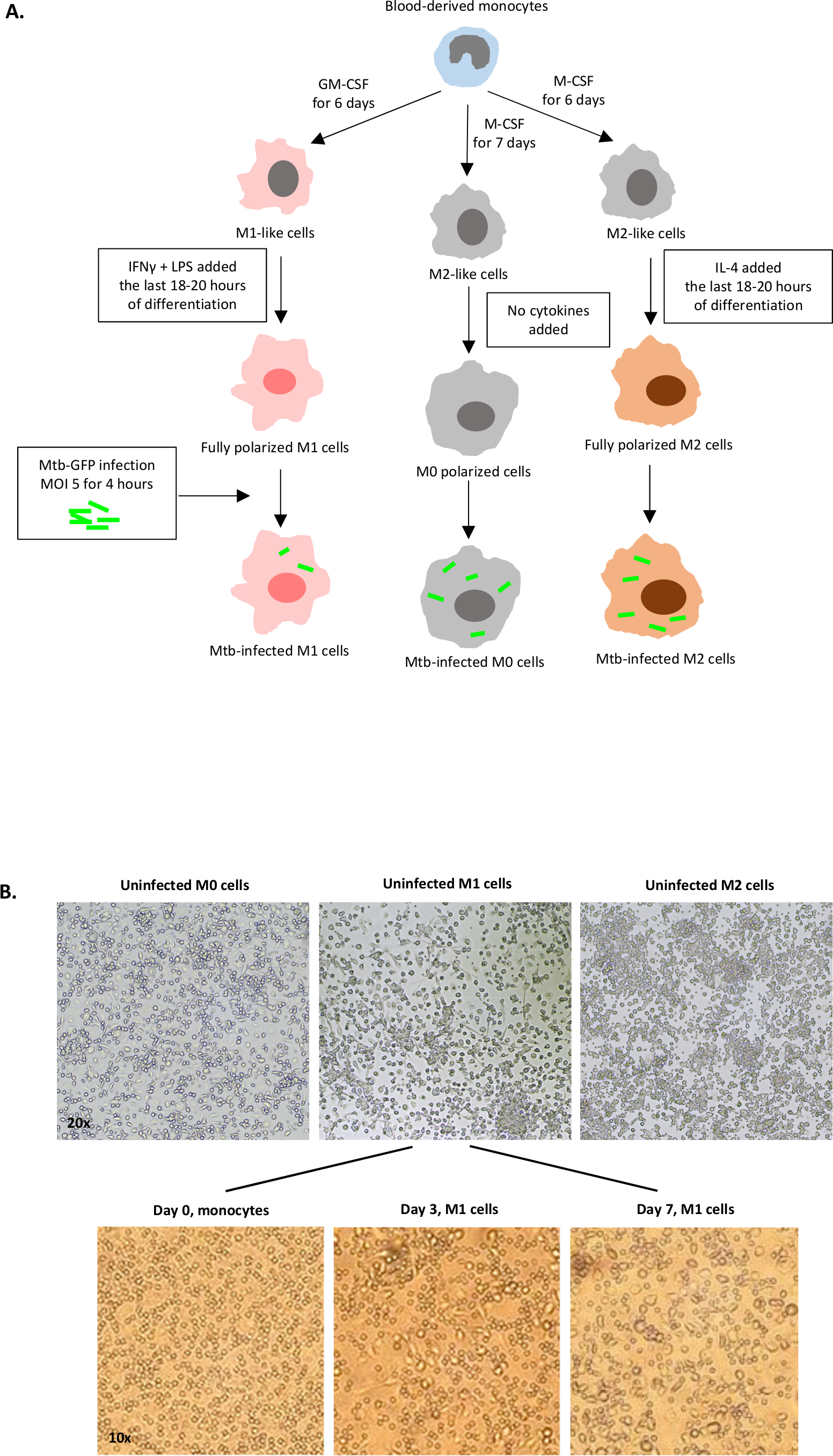
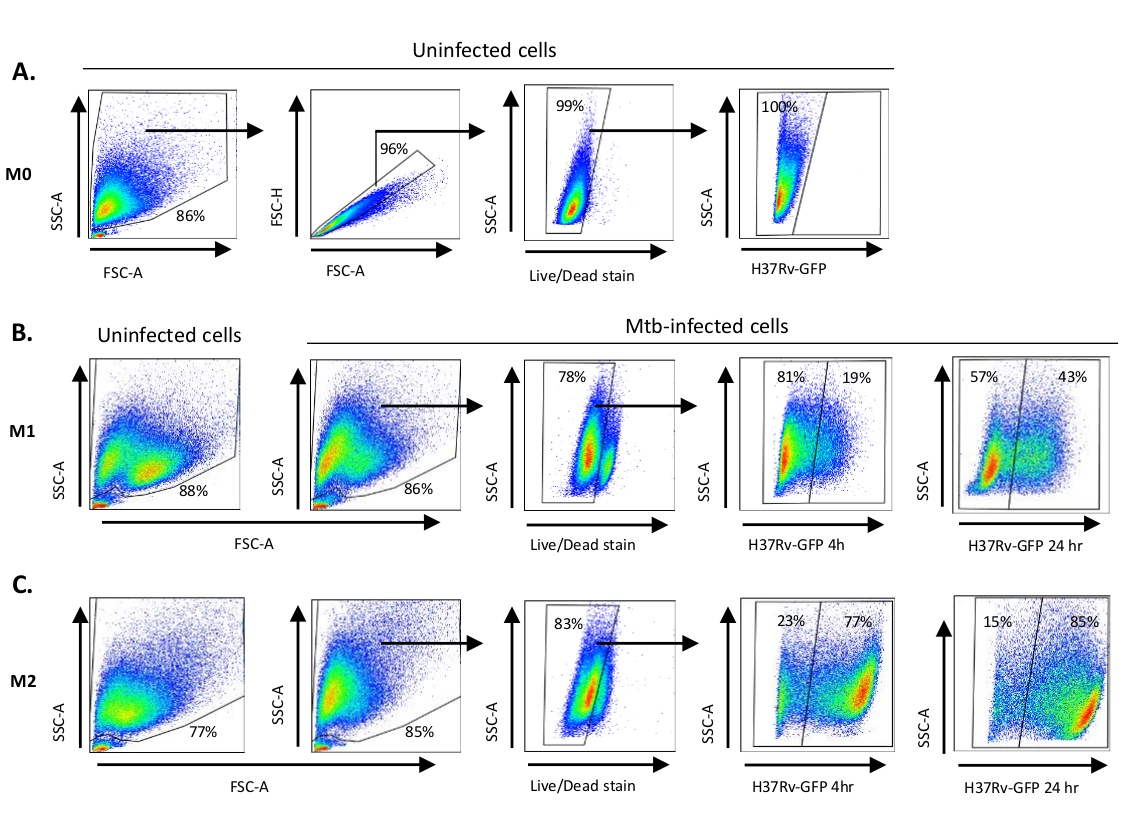
Una ilustración esquemática de las estimulaciones de citoquinas utilizadas para la polarización de células derivadas de monocitos a M0 (células similares a M2), M1 (células M1 totalmente polarizadas) y M2 (células M2 totalmente polarizadas) se presenta en la Figura 1A,mientras que las imágenes representativas de cultivos celulares M0, M1 y M2, así como cultivos M1 en los días 0, 3 y 7, se muestran en la Figura 1B. Las células M0 no infectadas se utilizaron para demostrar la estrategia básica de gating (Figura 2A). Inicialmente, las células mieloides (~85%) fueron cerradas de acuerdo con sus propiedades de dispersión hacia adelante (FSC) y dispersión lateral (SSC), incluyendo las celdas más grandes con granularidad alta y excluyendo los desechos de tamaño pequeño con un SSC y FSC bajos que se encuentran en la esquina inferior izquierda de la parcela de puntos. En la segunda gráfica, los dobles (es decir, los grupos de celdas) se definieron como tener un área aumentada pero una altura similar en comparación con las celdas individuales y se excluyeron de un análisis posterior. Por lo tanto, sólo las células proporcionadas entre fsc-área y altura FSC (células individuales) se incluyeron dentro de la puerta de forma inclinada. A continuación, el tinte de viabilidad Zombie-UV que mancha las proteínas citoplasmáticas dentro de las células muertas, se utilizó para excluir las células muertas del análisis posterior. Como era de esperar, las células M0 no infectadas viables eran negativas para la expresión Mtb-GFP visualizada en el canal FITC.
A continuación, aplicamos la misma estrategia de gating a macrófagos M1 y M2 infectados por Mtb a las 4 horas posteriores a la infección(Figura 2B,C). Se detectaron dos subpoblaciones en la puerta FCS/SSC de macrófagos polarizados M1 no infectados; una población con un tamaño más pequeño (FCS) y mayor granularidad (SSC) y la otra población con un tamaño más grande y menor granularidad(Figura 2B),mientras que la puerta principal de las células M2 no infectadas parecía más homogénea(Figura 2C). Las células derivadas de monocitos M1 y M2 mostraron un cambio vertical a mayor granularidad y menor tamaño celular sobre la infección por Mtb, lo que puede reflejar una mayor complejidad dentro de las células causada por la absorción de bacterias Mtb intracelulares(Figura 2B,C). Además, la mancha de viabilidad reveló una muerte celular mejorada (17-22%) entre las células M1 y M2 infectadas por Mtb a un MOI de 5, en comparación con las células M0 no infectadas (99%) (Figura 2A-C) o celdas M1 y M2 no infectadas (datos no mostrados). Los datos representativos mostraron que la expresión Mtb-GFP (es decir, infectividad Mtb) fue sustancialmente mayor en M2 (77% células positivas de GFP) en comparación con las células M1 (19% células GFP positivas) después de 4 horas de infección(Figura 2B,C). Después de 24 horas de infección, la expresión Mtb-GFP fue del 43% y del 85% en las células M1 y M2 respectivamente, lo que sugiere que las células M1 tuvieron un aumento relativamente mayor en la expresión de GFP de 4 a 24 horas después de la infección por Mtb en comparación con las células M2, 126% frente a 10,4% de aumento en la expresión de GFP en células M1 y M2 de 4 a 24 horas, respectivamente.
Para caracterizar la eficacia de la polarización M1/M2 en células derivadas de monocitos no infectadas, se utilizaron gráficas de puntos para identificar células M1 que eran doblemente positivas para CD64 y CD86 (CD64+CD86+) y células M2 que eran doblemente positivas para CD163 y CD200R (CD163+CD200R+; Figura 3A,B). La selección de marcadores M1/M2 se realizó principalmente en base a los resultados de nuestro trabajo anterior25, pero también de otros estudios26,27,28,29. Los cuadrantes para las celdas manchadas, se establecieron utilizando las puertas correspondientes para las células M1/M2 sin manchar (Figura 3A). Ninguno de estos marcadores es expresado exclusivamente por células M1 o M2, pero la proporción de células positivas, así como la intensidad de la expresión superficial es diferente. Esto fue particularmente evidente de la mancha M1 donde alrededor del 95% de las células M1 y el 79% de las células M2 eran CD64+CD86+, pero la intensidad de tinción fue sustancialmente mayor en el subconjunto M1 (Figura 3A). Mientras que el 27% de las células M1 fueron positivas para las células CD200R de marcador M2, sólo el 1% fueron positivas para CD163, proporcionando 0.5% CD163+CD200R+ células M1 en comparación con 63% CD163+CD200R+ células M2 (Figura 3A). Después de 4 horas de infección por Mtb, se observó un aumento en la frecuencia de las células CD200R+ en células polarizadas Mtb-GFP-positivas M1 (16%), mientras que la expresión CD163 se redujo en células M2(Figura 3B). El mapa de calor muestra una alta intensidad de expresión GFP en células CD163+CD200R+ M2, pero también en el subconjunto CD64+CD86+ M2 en comparación con los subconjuntos de celdas M1 correspondientes (Figura 3B). En general, el cambio de expresión de los marcadores M1 y M2 respectivos también se visualiza en los histogramas de la Figura 3C. Además, las bacterias Mtb-GFP también se visualizaron en células CD64+ M1 y en células CD163+ M2 mediante microscopía confocal, que apoyó una absorción intracelular mejorada y/o crecimiento de Mtb dentro de M2 en comparación con las células M1(Figura 3D).
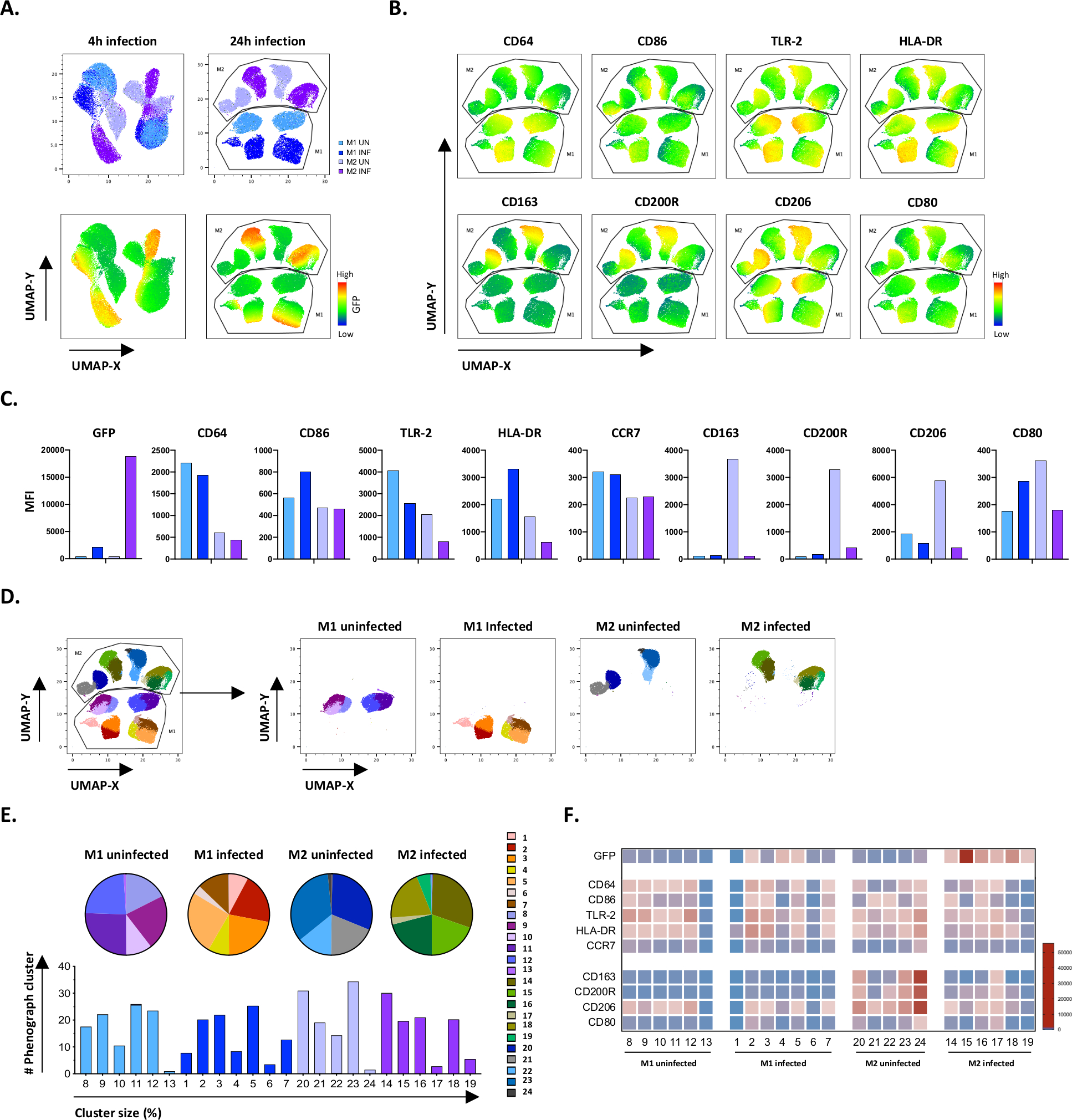
Para verificar los resultados de la gating manual, aplicamos la reducción de dimensionalidad utilizando aproximación y proyección uniforme de colectores (UMAP). El análisis de UMAP mostró que la infección por Mtb durante 4 horas no era suficiente para afectar la polarización de los macrófagos, a diferencia de las 24 horas de infección, lo que dio lugar a racimos claramente separados de células M1 y M2 no infectadas e infectadas(Figura 4A). Los macrófagos M1 no infectados mostraron una mayor expresión de CD64, CD86, TLR2, HLA-DR y CCR7 en comparación con los macrófagos M2, mientras que las células M2 no infectadas mostraron una fuerte regulación de los marcadores de fenotipo M2 CD163, CD200R, CD206 y CD80 (Figura 4B,C). De acuerdo con la gating manual, la infección por Mtb después de 24 horas causó una clara regulación descendente de CD163, CD200R y CD206 en células M2 y la regulación de CD86 y HLA-DR en células M1(Figura 4B,C),lo que sugiere que Mtb puede modular la polarización del macrófago. El análisis posterior del fenógrafo (Figura 4D-F) identificó 24 grupos diferentes de diferentes tamaños que se distribuyeron de forma única entre las células infectadas por M1 y M2 y Mtb, como se ilustra en los gráficos UMAP (Figura 4D), gráficos circulares (Figura 4E) y mapas de calor (Figura 4F). En total, estos resultados muestran una eficiencia prometedora de este protocolo para generar células polarizadas M1 y M2 fenotípica y funcionalmente diversas in vitro que se modulan aún más por la infección por Mtb.

Figura 1: Ilustración esquemática de la diferenciación in vitro y la polarización de las células derivadas del mieloide humano. (A) Se representan células M0 (M2-like), M1 (activadas clásicamente) y M2 (alternativamente activadas). Los monocitos obtenidos de donantes sanos de sangre fueron polarizados con diferentes citoquinas como se describe en el protocolo e infectados con la cepa Mtb etiquetada por GFP, H37Rv, durante 4 horas antes del análisis con citometría de flujo de 10 colores. Las células polarizadas M1 suelen contener menos bacterias en comparación con las células polarizadas por M2. (B) Imágenes microscópicas de células M0, M1 y M2 totalmente polarizadas y no infectadas en las placas de 6 pozos en el día 7, e imágenes representativas de la diferenciación celular M1 de monocitos en los días 0, 3 y 7. La ampliación es de 20x (panel superior) y 10x (panel inferior). Tenga en cuenta que las células M1 son más alargadas y estiradas en comparación con las células M0 y M2 más redondeadas (panel superior). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Estrategia de Gating de células derivadas del mieloide polarizadas diferencialmente. Gráficas de puntos representativas que muestran (A) Propiedades de dispersión hacia delante (FSC) y dispersión lateral (SSC) de macrófagos M0 no infectados. La gráfica FSC-A/FSC-H muestra la gating manual de células individuales proporcionadas para el área y la altura. La puerta de celda en vivo excluyó las células que eran positivas para Zombie-UV (tinte de viabilidad). El Mtb intracelular fue detectado por la expresión GFP en células vivas observadas en el canal FITC. (B) Gating de macrófagos M1 y (C) M2 que muestran gráficas de puntos FCS/SSC tanto en células no infectadas como en células infectadas por Mtb 4 h y 24 h después de la infección. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Eficacia del protocolo de polarización M1/M2 in vitro. Gráficas representativas de puntos y gating de cuadrante que muestran frecuencias de subconjunto de células polarizadas M1 y M2 utilizando células no infectadas CD64 y CD86 (M1) o CD163 y CD200R (M2) en (A) células no manchadas y manchadas no infectadas y (B) células manchadas infectadas con Mtb 4 h después de la infección. Las gráficas de puntos en (B) ilustran la intensidad de fluorescencia de la expresión GFP (mapa de calor) en macrófagos polarizados M1 y M2 obtenidos de diferentes sub-puertas. (C) La media geométrica de intensidad de fluorescencia (IMF) se muestra en histogramas de un donante representativo después de 4 h de infección por Mtb. Los valores MFI en células M1 (azul claro) y M2 no infectadas (púrpura claro) se presentan en el panel superior y las células M1 infectadas por Mtb (azul profundo) y M2 (púrpura profundo) se presentan en el panel inferior. (D) Se muestran imágenes confocales representativas de células polarizadas M1 y M2 infectadas por M1 y Mtb. Las células M1 y M2 fueron manchadas para la expresión CD64 y CD163, respectivamente, utilizando inmunofluorescencia. La tinción superficial positiva se muestra en rojo y las bacterias intracelulares que expresan GFP se muestran en verde. Los núcleos manchados por DAPI se muestran en color azul. Escala – 10 μm. La ampliación de las imágenes a la derecha es 350x. Por favor, haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Reducción de la dimensionalidad con aproximación y proyección uniforme del colector (UMAP) y análisis fenógrafo de células M1 y M2 no infectadas por Mtb. (A) UMAP, creado concatenando 11000 células vivas de cultivos celulares M1 y M2 no infectados por Mtb a partir de dos donantes de sangre representativos, 4 h (gráficos izquierdos) o 24 h (gráficos derecho) después de la infección. El mapa de calor para la expresión GFP (panel inferior) indica células no infectadas e infectadas por Mtb. (B-C) MFI de marcadores expresados en células M1 y M2 no infectadas por Mtb 24 h después de la infección, que se muestran como gráficas de barras (B) o (C). (D-F) El análisis fenógrafo identificó 24 grupos que se distribuyen diferencialmente entre los cultivos M1 y M2 no infectados por Mtb y no infectados. Los racimos 8–13 son únicos en células M1 no infectadas, los racimos 1–7 son únicos en células M1 infectadas por Mtb, los racimos 20-24 son únicos en células M2 no infectadas y los racimos 14-19 son únicos en células M2 infectadas con Mtb. El MFI de cada marcador en cada cúmulo fenógrafo se muestra en (F). Los datos se presentan como células M1 (azul claro) y M2 no infectadas (púrpura claro) y células M1 infectadas por Mtb (azul profundo) y M2 (púrpura profundo). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Tabla 1: Lista de anticuerpos utilizados para la citometría de flujo.
| láser | filtro | fluorocromo | fenotipo | función | clon | Catálogo no. | compañía |
| 639 | 670/30 | AF647 | TLR2 | Receptor de reconocimiento de patógenos | TL2.1 | 309714 | BioLegend |
| 639 | 780/60 | APC-Cy7 | CD206 | Receptor de la nariz de mano | 15-2 | 321120 | BioLegend |
| 405 | 610/20 | BV605 | CD163 | Receptor carroñero | GHI/61 | 333616 | BioLegend |
| 405 | 670/30 | BV650 | CD80 | Molécula coestimuladora | 2D10 | 305227 | BioLegend |
| 405 | 710/50 | BV711 | CCR7 | Receptor de quimiocina | G043H7 | 353228 | BioLegend |
| 405 | 780/60 | BV785 | CD86 | Molécula coestimuladora | IT2.2 | 305442 | BioLegend |
| 488 | 530/30 | GFP | Mtb | Bacterias intracelulares | |||
| 561 | 586/15 | pei | CD200R | Receptor inhibitorio | OX-108 | 329306 | BioLegend |
| 561 | 620/14 | PE/DESLUMBRAR 594 | CD64 | Fc receptor gamma-I de IgG | 10.1 | 305032 | BioLegend |
| 561 | 661/20 | PE-Cy5 (PC5) | HLA-DR | Molécula de clase II de MHC | L243 | 307608 | BioLegend |
| 355 | 450/50 | BUV395 | Tinte de viabilidad | Marcador de celda vivo/muerto | Zombie UV | 423108 | Invitrogen |
Discusión
Este protocolo experimental describe la polarización efectiva de las células derivadas del mieloide en fenotipos M1 o M2, incluida la evaluación con un panel de citometría de flujo de 10 colores que permite visualizar y caracterizar profundamente los Mtb etiquetados con GFP en diversos subconjuntos de macrófagos. Aunque la tuberculosis es una enfermedad humana antigua, actualmente no existe un modelo estándar dorado para estudiar las interacciones Mtb-macrófago, y la citometría de flujo multicolor de macrófagos podría complicarse en comparación con los análisis de las respuestas a los linfocitos. Pocos protocolos disponibles para la diferenciación in vitro de monocitos humanos a macrófagos presentan un profundo conocimiento del tipo de macrófagos generados. Un protocolo básico para la polarización del macrófago y la evaluación citométrica del flujo de la activación del macrófago utilizando un panel sólido de marcadores probablemente puede facilitar dicha caracterización y ofrecer oportunidades para explorar características adicionales de las células polarizadas tratadas en diferentes condiciones. Esto incluye análisis de células cultivadas in vitro, así como análisis de células in vivo en muestras clínicas, es decir, suspensiones de PBMC y unicelulares de fluidos corporales (es decir, lavado broncoalveolar) o tejido homogeneizado. En consecuencia, la diferenciación y/o el estado de activación de monocitos y macrófagos obtenidos de los pacientes podrían estar relacionados con el resultado de la enfermedad. Se ha notificado una expansión de CD16+CD163+ monocitos en sangre periférica en pacientes con tuberculosis pulmonar30. También se detectó un aumento de la frecuencia de las células CD163+ en la piel inflamada de los pacientes con dermatitis atópica31. Del mismo modo, se ha demostrado que los macrófagos similares a los CD206+ M2 inhiben la proliferación y diferenciación de células en el microambiente del tejido adipocito32 y se enriquecen en muestras de médula ósea de pacientes con leucemia mieloide aguda (LMA)29. Se encontró que una proporción elevada de células CD64 (M1) a CD163 (M2) en sangre entera de pacientes con osteoartritis se asoció con la gravedad de la enfermedad33. Otro estudio utilizó CD86 (M1) y CD163 (M2) para demostrar que la alta expresión M1 en el tejido se correlacionaba con un peor resultado en un subgrupo de tumores cerebrales malignos34.
Hay varias ventajas significativas de este protocolo experimental de citometría de flujo M1/M2. Este modelo ofrece la oportunidad de estudiar las respuestas inmunes innatas a la infección por Mtb virulenta y se puede desarrollar para contener estudios de respuestas inmunes adaptativas mediante la adición de células T autólogas junto con macrófagos M1 o M2 en reacciones de linfocitos mixtos (MLR). El protocolo también es adecuado para la detección de fármacos y pruebas de diferentes compuestos inmunomoduladores y antimicrobianos. Aquí, hemos estudiado previamente los efectos de la vitamina D y el inhibidor de la deacetilasa de piedra histone fenilbutirato en células derivadas del mieloide después de la infección por Mtb25,35. La citometría de flujo M1/M2 también podría utilizarse para evaluar la activación del macrófago después de acondicionar con sobrenautas de cultivo celular o plasma del paciente. Si bien los estudios in vivo de coinfección de tuberculosis con VIH o helmintos o co-morbilidad tb-diabetes podrían ser difíciles, el modelo M1/M2 menos complejo puede facilitar estudios de co-morbilidades in vitro. Del mismo modo, el protocolo podría ser explotado para estudios de transmisión para examinar la infectividad Mtb de las células o para investigar la capacidad de presentación de fácticos y antígenos de células M1/M2 individuales. La citometría de flujo M1/M2 también es atractiva para su uso en estudios de biomarcadores y vacunas, para seguir el pronóstico de la enfermedad durante el tratamiento y para probar terapias dirigidas a células derivadas del mieloide. Es importante destacar que una serie de métodos diferentes podrían aplicarse en paralelo a la citometría de flujo para la evaluación simultánea de fenotipos de polarización de macrófagos y respuestas funcionales utilizando microscopía confocal(Figura 3D),PCR en tiempo real, mancha occidental, ensayos multiplex y ELISA de factores solubles en el sobrenadante de cultivo, así como la evaluación de la infectividad bacteriana intracelular y el crecimiento utilizando la expresión GFP (citometría de flujo y microscopía confocal) y unidades formadoras de colonias (CFU). La infección de células M1 o M2 con bacterias Mtb-GFP también permite ordenar las células no infectadas y infectadas por Mtb de la misma muestra para el análisis de secuenciación de ARN de una sola célula.
El protocolo descrito también tiene algunas limitaciones, incluidas las desventajas técnicas y científicas. El inconveniente que utiliza macrófagos derivados de monocitos de donantes de sangre humana es que la variabilidad del donante a menudo es alta y el hecho de que las células no están polarizadas en el entorno fisiológico de los tejidos humanos. La gran variabilidad en la eficacia de la polarización M1/M2 o la infectividad de Mtb entre donantes puede dar lugar a problemas con variaciones intra e interexperimentales, bajo poder estadístico y la necesidad de incluir a muchos donantes para obtener resultados fiables. Además, la adherencia plástica de los monocitos de los PBMCs da lugar a un número dependiente del donante de monocitos/pozo que eventualmente puede proporcionar un MOI arbitrario que podría afectar la polarización del macrófago y la viabilidad celular después de la infección por Mtb. Los pasos críticos en el protocolo implican un lavado adecuado para evitar que otros tipos de células contaminen los cultivos celulares que también podrían afectar a la polarización del macrófago. Mientras que un MOI demasiado bajo puede imitar la infección por tuberculosis latente, un MOI demasiado alto matará las células, destacando la importancia de usar un MOI apropiado. Además, podría ser difícil recuperar células firmemente adherentes tras el desprendimiento, lo que puede dar lugar a una representación sesgada de ciertos subconjuntos de macrófagos utilizados para los análisis de citometría de flujo. Un paso crucial en el análisis de la citometría de flujo implica el uso adecuado de la matriz de compensación de cuentas y controles negativos como células no manchadas o controles FMO (Fluorescencia Menos Uno) para garantizar una correcta gating manual.
Otra limitación consiste en la polarización de monocitos derivados de la sangre y no del entorno tisular local. El sello distintivo de la tuberculosis humana es la formación de granulomas en tejidos infectados por Mtb y, por lo tanto, la inmunopatología en la tuberculosis debe estudiarse preferentemente en el sitio del tejido local. Sin embargo, los monocitos se reclutan al pulmón a partir de sangre periférica tras la inflamación/infección, donde las células pueden diferenciarse en macrófagos en presencia de citoquinas inflamatorias como gm-CSF12. Es importante destacar que, en el entorno fisiológico del tejido in vivo, es probable que haya una gran heterogeneidad de polarización del macrófago incluyendo una mezcla y diferentes proporciones de diversas poblaciones de macrófagos similares a M1 y M2 que contribuyen al destino de la infección por tuberculosis36. Anteriormente hemos desarrollado un modelo de tejido pulmonar organotípico humano que permite estudios 3D de formación de granuloma mediado por macrófago en TB37. Podría ser interesante explotar el protocolo de polarización M1/M2 actual en combinación con el modelo de tejido pulmonar para seguir estudiando la formación de granuloma de tuberculosis, las funciones del efector y la relación M1/M2 en el tejido experimental.
Este protocolo de diagrama de flujo M1/M2 podría adaptarse fácilmente para incluir un panel extendido de marcadores mieloides útiles para la evaluación de características asociadas con respuestas inhibitorias e inflamatorias. Hay un gran interés de investigación en moléculas inhibitorias de punto de control inmune como PD-1, SIRP-α, IDO y arginases que podrían modular las respuestas de macrófago38. En este contexto, la polarización de las células mieloides también podría implicar otros estímulos que promuevan los macrófagos inmunoreguladores (Mreg) o las células supresoras derivadas del mieloide (MDSC) que se ha demostrado que están implicadas en varias enfermedades, incluida la tuberculosis38. Los paneles de citometría de flujo más avanzados de los subconjuntos de macrófagos M1/M2/Mreg también pueden incluir tinción intracelular de citoquinas/quimioquinas IL-1β, TNF-α, IL-10 y MCP-1 u otros factores solubles o moléculas efectores como óxido nítrico inducible (iNOS) y péptidos antimicrobianos. Esto podría mejorar las posibilidades de estudiar las respuestas de macrófago polifuncional, similar a lo que se ha descrito extensamente para las células T39.
Actualmente, los paneles de tinción de citometría de flujo pueden incluir hasta 30-40 colores, lo que proporciona la capacidad de inmunofenotipo de múltiples subconjuntos celulares y moléculas simultáneamente. La configuración experimental básica de este protocolo de citometría de flujo M1/M2 puede utilizarse como columna vertebral compatible con la mayoría de los cytómetros de flujo antiguos y nuevos y se puede construir y adaptar según las necesidades individuales, incluidos los desafíos planteados por el trabajo con Mtb virulenta en un entorno BSL-3. Hoy en día, técnicas de reducción de dimensionalidad como UMAP están disponibles en las nuevas versiones del software de citometría de flujo, lo que permite el análisis de un gran número de parámetros generados en estudios unicelulares que es esencial para mejorar la visualización e interpretación de datos en alta dimensión40. Las constantes mejoras tecnológicas en la citometría de flujo probablemente continuarán en los próximos años, incluyendo la combinación de fenotipado multiparamétrico junto con capacidades modernas de clasificación celular, donde este protocolo podría resultar útil en varios ensayos de infección por Mtb basados en macrófagos.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Agradecemos a nuestros colegas de la Agencia de Salud Pública de Suecia, Matilda Svensson y Solomon Ghebremichael por su asistencia en el laboratorio BSL-3.
Este trabajo fue apoyado por subvenciones de la Fundación Sueca del Corazón y el Pulmón (HLF) (2019-0299 y 2019-0302 a SB), el Consejo Sueco de Investigación (VR) (2014-02592, 2019-01744 y 2019-04720 a SB), la Fundación para Prevenir la Resistencia a los Antibióticos (Resistencia), las Fundaciones Karolinska Institutet y KID a SB (financiación parcial de la educación doctoral para Marco Loreti) del Instituto Karolinska. Ml recibió el apoyo de la Fundación Sueca contra el Cáncer Infantil (TJ2018-0128 y PR2019-0100).
Materiales
| Name | Company | Catalog Number | Comments |
| 8-well chamber slides | Lab-Tek | 154534 | |
| BD Comp bead plus | BD | 560497 | |
| Bovine Serum Albumin | Sigma-Aldrich | A7906 | |
| DAPI Mounting media | Vector Laboratories | H-1200-10 | |
| EDTA (0.5 M) | Karolinska University hospital, Huddinge | N/A | |
| Falcon 6-well Flat Bottom plates | Corning Life Sciences | 353046 | |
| Fetal bovine serum (FBS) | Sigma-Aldrich | F7524 | |
| Formaldehyde | Sigma-Aldrich | F8775 | |
| Glycerol (70%) | Karolinska University hospital, Huddinge | N/A | |
| GM-CSF | Peprotech | 300-03 | |
| Goat anti-mouse IgG Alexa Fluor 594 secondary antibody | Invitrogen | R37121 | Secondary antibody for CD64 |
| Goat anti-Rabbit IgG Alexa Fluor 594 secondary antibody | Invitrogen | A-11037 | Secondary antibody for CD163 |
| HEPES | GE Healthcare Life Sciences | SH30237.01 | |
| IFN-γ | Peprotech | 300-02 | |
| IL-4 | Peprotech | 200-04 | |
| L-Glutamine | GE Healthcare Life Sciences | SH30034.01 | |
| LPS (Escherichia coli O55:B5) | Sigma-Aldrich | L6529 | |
| Lymphoprep | Alere Technologies AS | 11508545 | |
| M-CSF | Peprotech | 300-25 | |
| Middle Brook 7H10 agar plates | Karolinska University hospital, Huddinge | N/A | |
| Middle Brook 7H9 media | Karolinska University hospital, Huddinge | N/A | |
| Mouse anti-human CD64 primary antibody | Bio-Rad | MCA756G | Clone: 10.1 |
| Na-pyruvate | GE Healthcare Life Sciences | SH300239.01 | |
| Normal goat serum | Jackson ImmunoResearch | 005-000-121 | |
| Rabbit anti-human CD163 primary antibody | GeneTex | GTX81526 | Polyclonal |
| RPMI 1640 | Life Technologies Corporation | SH30096.01 | |
| Triton X-100 | Sigma-Aldrich | X-100 | |
| TubeSpin bioreactor tubes | TPP Techno Plastic Products AG | 87050 | |
| Tween-20 | Sigma-Aldrich | P9416 | |
| Tween-80 | Sigma-Aldrich | P4780 |
Referencias
- Sica, A., Mantovani, A. Macrophage plasticity and polarization: in vivo veritas. Journal of Clinical Investigation. 122 (3), 787-795 (2012).
- Cassetta, L., Cassol, E., Poli, G. Macrophage polarization in health and disease. Scientific World Journal. 11, 2391-2402 (2011).
- Mills, C. D., Kincaid, K., Alt, J. M., Heilman, M. J., Hill, A. M. M-1/M-2 macrophages and the Th1/Th2 paradigm. Journal of Immunology. 164 (12), 6166-6173 (2000).
- Martinez, F. O., Gordon, S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000 Prime Reports. 6, 13 (2014).
- Atri, C., Guerfali, F. Z., Laouini, D. Role of human macrophage polarization in inflammation during infectious diseases. International Journal of Molecular Sciences. 19 (6), (2018).
- Flynn, J. L., Gideon, H. P., Mattila, J. T., Lin, P. L. Immunology studies in non-human primate models of tuberculosis. Immunological Reviews. 264 (1), 60-73 (2015).
- Mosser, D. M., Edwards, J. P. Exploring the full spectrum of macrophage activation. Nature Reviews Immunology. 8 (12), 958-969 (2008).
- Fleetwood, A. J., Lawrence, T., Hamilton, J. A., Cook, A. D. Granulocyte-macrophage colony-stimulating factor (CSF) and macrophage CSF-dependent macrophage phenotypes display differences in cytokine profiles and transcription factor activities: implications for CSF blockade in inflammation. Journal of Immunology. 178 (8), 5245-5252 (2007).
- Nathan, C. F., Murray, H. W., Wiebe, M. E., Rubin, B. Y. Identification of interferon-gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. Journal of Experimental Medicine. 158 (3), 670-689 (1983).
- Leidi, M., et al. M2 macrophages phagocytose rituximab-opsonized leukemic targets more efficiently than m1 cells in vitro. Journal of Immunology. 182 (7), 4415-4422 (2009).
- Stein, M., Keshav, S., Harris, N., Gordon, S. Interleukin 4 potently enhances murine macrophage mannose receptor activity: a marker of alternative immunologic macrophage activation. Journal of Experimental Medicine. 176 (1), 287-292 (1992).
- Italiani, P., Boraschi, D. From Monocytes to M1/M2 macrophages: Phenotypical vs. functional differentiation. Frontiers in Immunology. 5, 514 (2014).
- Verreck, F. A., et al. Human IL-23-producing type 1 macrophages promote but IL-10-producing type 2 macrophages subvert immunity to (myco)bacteria. Proceedings of the National Academy of Sciences. 101 (13), 4560-4565 (2004).
- Redente, E. F., et al. Differential polarization of alveolar macrophages and bone marrow-derived monocytes following chemically and pathogen-induced chronic lung inflammation. Journal of Leukocyte Biology. 88 (1), 159-168 (2010).
- Refai, A., Gritli, S., Barbouche, M. R., Essafi, M. Mycobacterium tuberculosis virulent factor ESAT-6 drives macrophage differentiation toward the pro-inflammatory M1 phenotype and subsequently switches it to the anti-inflammatory M2 phenotype. Frontiers in Cellular and Infection Microbiology. 8, 327 (2018).
- Kahnert, A., et al. Alternative activation deprives macrophages of a coordinated defense program to Mycobacterium tuberculosis. European Journal of Immunology. 36 (3), 631-647 (2006).
- Marino, S., et al. Macrophage polarization drives granuloma outcome during Mycobacterium tuberculosis infection. Infection and Immunity. 83 (1), 324-338 (2015).
- Erbel, C., et al. An in vitro model to study heterogeneity of human macrophage differentiation and polarization. Journal of Visualized Experiments. (76), e50332 (2013).
- Lee, C. Z. W., Kozaki, T., Ginhoux, F. Publisher Correction: Studying tissue macrophages in vitro: are iPSC-derived cells the answer. Nature Reviews Immunology. 18 (11), 726 (2018).
- Ying, W., Cheruku, P. S., Bazer, F. W., Safe, S. H., Zhou, B. Investigation of macrophage polarization using bone marrow derived macrophages. Journal of Visualized Experiments. (76), e50323 (2013).
- Van den Bossche, J., Baardman, J., de Winther, M. P. Metabolic characterization of polarized M1 and M2 bone marrow-derived macrophages using real-time extracellular flux analysis. Journal of Visualized Experiments. (105), e53424 (2015).
- McKinnon, K. M. Flow Cytometry: An Overview. Current Protocols in Immunology. 120, 1-11 (2018).
- Njoroge, J. M., et al. Characterization of viable autofluorescent macrophages among cultured peripheral blood mononuclear cells. Cytometry. 44 (1), 38-44 (2001).
- Li, F., et al. Autofluorescence contributes to false-positive intracellular Foxp3 staining in macrophages: a lesson learned from flow cytometry. Journal of Immunological Methods. 386 (1-2), 101-107 (2012).
- Rao Muvva, J., Parasa, V. R., Lerm, M., Svensson, M., Brighenti, S. Polarization of human monocyte-derived cells with vitamin D promotes control of Mycobacterium tuberculosis infection. Frontiers in Immunology. 10, 3157 (2019).
- Tarique, A. A., et al. functional, and plasticity features of classical and alternatively activated human macrophages. American Journal of Respiratory Cell and Molecular Biology. 53 (5), 676-688 (2015).
- Hristodorov, D., et al. Targeting CD64 mediates elimination of M1 but not M2 macrophages in vitro and in cutaneous inflammation in mice and patient biopsies. MAbs. 7 (5), 853-862 (2015).
- Jaguin, M., Houlbert, N., Fardel, O., Lecureur, V. Polarization profiles of human M-CSF-generated macrophages and comparison of M1-markers in classically activated macrophages from GM-CSF and M-CSF origin. Cellular Immunology. 281 (1), 51-61 (2013).
- Xu, Z. J., et al. The M2 macrophage marker CD206: a novel prognostic indicator for acute myeloid leukemia. Oncoimmunology. 9 (1), 1683347 (2020).
- Liu, Q., et al. Differential expression and predictive value of monocyte scavenger receptor CD163 in populations with different tuberculosis infection statuses. BMC Infectious Diseases. 19 (1), 1006 (2019).
- Sugaya, M., et al. Association of the numbers of CD163(+) cells in lesional skin and serum levels of soluble CD163 with disease progression of cutaneous T cell lymphoma. Journal of Dermatological Science. 68 (1), 45-51 (2012).
- Nawaz, A., et al. CD206(+) M2-like macrophages regulate systemic glucose metabolism by inhibiting proliferation of adipocyte progenitors. Nature Communications. 8 (1), 286 (2017).
- Liu, B., Zhang, M., Zhao, J., Zheng, M., Yang, H. Imbalance of M1/M2 macrophages is linked to severity level of knee osteoarthritis. Experimental and Therapeutic Medicine. 16 (6), 5009-5014 (2018).
- Lee, C., et al. M1 macrophage recruitment correlates with worse outcome in SHH Medulloblastomas. BMC Cancer. 18 (1), 535 (2018).
- Rekha, R. S., et al. Phenylbutyrate induces LL-37-dependent autophagy and intracellular killing of Mycobacterium tuberculosis in human macrophages. Autophagy. 11 (9), 1688-1699 (2015).
- Mattila, J. T., et al. Microenvironments in tuberculous granulomas are delineated by distinct populations of macrophage subsets and expression of nitric oxide synthase and arginase isoforms. Journal of Immunology. 191 (2), 773-784 (2013).
- Braian, C., Svensson, M., Brighenti, S., Lerm, M., Parasa, V. R. A 3D Human Lung Tissue Model for Functional Studies on Mycobacterium tuberculosis Infection. Journal of Visualized Experiments. (104), e53084 (2015).
- Brighenti, S., Joosten, S. A. Friends and foes of tuberculosis: modulation of protective immunity. Journal of Internal Medicine. , 12778 (2018).
- Chattopadhyay, P. K., Roederer, M. Good cell, bad cell: flow cytometry reveals T-cell subsets important in HIV disease. Cytometry Part A. 77 (7), 614-622 (2010).
- Becht, E., et al. Dimensionality reduction for visualizing single-cell data using UMAP. Nature Biotechnology. 37, 38-44 (2019).
Erratum
Formal Correction: Erratum: Polarization of M1 and M2 Human Monocyte-Derived Cells and Analysis with Flow Cytometry upon Mycobacterium tuberculosis Infection
Posted by JoVE Editors on 10/14/2020. Citeable Link.
An erratum was issued for: Polarization of M1 and M2 Human Monocyte-Derived Cells and Analysis with Flow Cytometry upon Mycobacterium tuberculosis Infection. Author and affiliation information was updated.
The author and affiliation information was updated from:
Akhirunnesa Mily1, Sadaf Kalsum1, Marco Giulio Loreti1, Rokeya Sultana Rekha2, Jagadeeswara Rao Muvva1, Magda Lourda1,3, Susanna Brighenti1
1Center for Infectious Medicine (CIM), Department of Medicine Huddinge, ANA Futura, Karolinska Institutet
2Clinical Microbiology, Department of Laboratory Medicine (Labmed), ANA Futura, Karolinska Institutet
3Childhood Cancer Research Unit, Department of Women's and Children's Health, Karolinska Institutet
to:
Akhirunnesa Mily1,2, Sadaf Kalsum1, Marco Giulio Loreti1, Rokeya Sultana Rekha3, Jagadeeswara Rao Muvva1, Magda Lourda1,4, Susanna Brighenti1
1Center for Infectious Medicine (CIM), Department of Medicine Huddinge, ANA Futura, Karolinska Institutet
2Infectious Diseases Division, International Centre for Diarrhoeal Disease Research, Bangladesh
3Clinical Microbiology, Department of Laboratory Medicine (Labmed), ANA Futura, Karolinska Institutet
4Childhood Cancer Research Unit, Department of Women's and Children's Health, Karolinska Institutet
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados