
Method Article
Analyse vidéomorphométrique de la vasoconstriction pulmonaire hypoxique des artères intra-pulmonaires à l’aide de tranches pulmonaires coupées de précision murine
Dans cet article
Résumé
La vasoconstriction pulmonaire hypoxique (HPV) est un phénomène physiologique important par lequel à l’hypoxie alvéolaire la perfusion pulmonaire est assortie à la ventilation. Le segment vasculaire principal contribuant au VPH est l’artère intra-acinnaire. Ici, nous décrivons notre protocole pour l’analyse du VPH des vaisseaux pulmonaires murins d’un diamètre de 20 à 100 μm.
Résumé
L’hypoxie alvéolaire aiguë provoque une vasoconstriction pulmonaire (VPH) - également connue sous le nom de mécanisme de von Euler-Liljestrand - qui sert à faire correspondre la perfusion pulmonaire à la ventilation. Jusqu’à présent, les mécanismes sous-jacents ne sont pas entièrement compris. Le segment vasculaire principal contribuant au VPH est l’artère intra-acinnaire. Cette section des vaisseaux est responsable de l’approvisionnement en sang d’un acinus individuel, qui est défini comme la partie du poumon distale à une bronchiole terminale. Les artères intra-acinaire sont principalement situées dans la partie du poumon qui ne peut pas être atteinte sélectivement par un certain nombre de techniques couramment utilisées telles que la mesure de la pression artérielle pulmonaire dans des poumons perfusés isolés ou des enregistrements de force à partir de segments d’artère pulmonaire proximaux disséqués1,2. L’analyse des vaisseaux sous-pléuraux par microscopie à luminescence confocale à balayage laser en temps réel est limitée aux vaisseaux ayant jusqu’à 50 μm de diamètre3.
Nous fournissons une technique pour étudier le VPH des artères intra-pulmonaires murines de l’ordre de 20 à 100 μm de diamètre intérieur. Il est basé sur l’analyse vidéomorphométrique des artères transversales en tranches pulmonaires coupées avec précision (PCLS). Cette méthode permet la mesure quantitative de la vasoreactivité de petites artères intra-acinaires d’un diamètre intérieur compris entre 20 et 40 μm qui sont situées à des goussets de septa alvéolaires à côté de conduits alvéolaires et d’artères pré-acinaires plus grandes de diamètre intérieur compris entre 40 et 100 μm qui courent à côté des bronches et des bronchioles. Contrairement à l’imagerie en temps réel des vaisseaux sous-phénolaires chez les souris anesthésiées et ventilées, l’analyse vidéomorphométrique de PCLS se produit dans des conditions exemptes d’effort de cisaillement. Dans notre modèle expérimental les deux segments artériels montrent un HPV monophasique une fois exposés au milieu gazé avec 1% O2 et la réponse s’estompe après la minute 30-40 à l’hypoxie.
Introduction
Dans la plupart des lits vasculaires systémiques, l’hypoxie induit une vasodilatation, en comparaison de la vasoconstriction causée par l’hypoxie dans la vascularisation pulmonaire. Cette réponse spécifique aux poumons à la baisse de la tension de l’oxygène est appelée vasoconstriction pulmonaire hypoxique (VPH), qui se manifeste en quelques secondes et s’inverse rapidement après le retour à la ventilation normoxique. Bien que le VPH soit connu depuis plus de 60 ans, les capteurs cellulaires d’oxygène et les cascades de signalisation entraînant une vasoconstriction font toujours l’objet de débats. Il existe un consensus relativement large selon lequel les changements redox et ROS évoqués par l’hypoxie sont essentiels pour le VPH et le développement de l’hypertension pulmonaire (examinés dans Sylvester et coll. 4 et Schumacker et al. 5). Nos propres données soutiennent un rôle central du complexe II de la chaîne respiratoire mitochondriale dans HPV6,7. Récemment, Wang et al. a présenté un concept complètement nouveau pour la détection de l’oxygène et le HPV: Sur la base de leurs données, ils proposent que l’hypoxie alvéolaire soit détectée par les capillaires adjacents provoquant la dépolarisation membranaire des cellules endothéliales. La réponse se propage via la connexion 40 jonctions intersitaires des cellules endothéliales conduisant à la constriction des cellules musculaires lisses des artérioles amont8.
Les artères du poumon longent les voies respiratoires, se ramifient avec elles, diminuent continuellement de diamètre et fournissent enfin du sang au système capillaire situé dans les parois alvéolaires. Cette circulation artérielle est composée de segments anatomiquement et fonctionnellement distincts. Les artères proximales de conduit, caractérisées par une abondance de fibres élastiques dans les murs, sont suivies des artères intra-pulmonaires entièrement musclées qui commandent en grande partie la résistance vasculaire pulmonaire. Étape par étape, ces artères transitent en segments où la couche musculaire devient incomplète, et enfin les vaisseaux sont exempts de cellules actine-immunoréactives de muscle lisse. L’artère intra-acinaire alimentant un acinus pulmonaire individuel avec du sang représente un segment partiellement musculaire6. De même, le système artériel pulmonaire ne représente pas une structure uniforme concernant la réponse hypoxique mais présente une diversité régionale marquée9,10. Par exemple, dans les artères pulmonaires proximales isolées des poumons du rat, l’hypoxie induit une réponse biphasique, présentant une contraction initiale rapide de courte durée qui - après relaxation incomplète - est suivie d’une seconde contraction lente mais soutenue11. Dans les artères de résistance isolées du parenchyme pulmonaire du rat en tant que quatrième et cinquième divisions des artères pulmonaires (diamètre extérieur <300 μm), l’hypoxie provoque une constriction monophasique9. Déjà en 1971, Glazier et Murray ont conclu à partir de mesures des changements dans la concentration en globules rouges capillaires dans les poumons de chiens ventilés avec des mélanges de gaz hypoxiques que l’augmentation induite par l’hypoxie de la résistance vasculaire s’est principalement produite en amont des capillaires12. De nos jours, la microscopie intravitale des poumons intacts de souris anesthésiées et ventilées mécaniquement représente un outil puissant pour l’analyse de la microvasculature pulmonaire13,14. L’excision d’une fenêtre circulaire dans la paroi thoracique donne un accès microscopique à la surface du poumon et permet l’analyse des vaisseaux pulmonaires sous-pléuraux de diamètre allant jusqu’à 50 μm. En combinant cette technique avec l’infusion de FITC-dextrane, Tabuchi et al. a démontré que seules les artérioles de taille moyenne d’un diamètre de 30 à 50 μm présentent une réponse marquée à l’hypoxie qui s’est maintenue sur une période de 60 minutes avec une atténuation mineure après 30 minutes. En revanche, les petites artérioles d’un diamètre de 20 à 30 μm ne présentaient qu’une réponse mineure à l’hypoxie3. Cependant, cette technique ne permet pas l’analyse d’artères d’un diamètre supérieur à 50 μm puisque ces vaisseaux sont situés trop profondément dans le tissu pulmonaire.
Afin de combler l’écart dans l’analyse des grandes et très petites artères pulmonaires (telles que les vaisseaux sous-plérurales) des poumons murins, nous avons adopté une méthode qui a été décrite par Martin et al. pour l’analyse de la réactivité des voies respiratoires15. Basé sur une technique d’instillation de gel d’agarose, il facilite la préparation de tranches pulmonaires coupées avec précision (PCLS) à partir de cet organe relativement doux et élastique. Dans le PCLS, la vasoreactivité des artères en coupe transversale d’un diamètre intérieur compris entre 20 et 100 μm peut être observée directement par vidéomicroscopie. L’application de médicaments lors de l’incubation hypoxique du PCLS permet l’analyse de leurs effets sur le VPH. Il est particulièrement important que cette technique puisse également être appliquée aux souris génétiquement modifiées. En fonction de leur emplacement dans le poumon, nous classons les artères comme des vaisseaux pré- et intra-acinaires, avec des diamètres intérieurs de 20-40 μm et 40-100 μm, respectivement. Sous une vue fonctionnelle l’artère intra-acinaire fournit à un acinus pulmonaire individuel avec le sang et l’artère pré-acinaire est les sections précédentes de navire. L’enregistrement d’images sur un appareil photo numérique permet la quantification ultérieure de la vasoreaction. Un attribut évident de ce modèle PCLS est l’absence de cisaillement-stress agissant sur l’endothélium. En revanche, dans les vaisseaux perfusés, le VPH aigu conduit à une augmentation du stress de cisaillement induisant ainsi des mécanismes secondaires tels que la libération de NO16. De plus, l’utilisation du PCLS permet de mesurer le VPH sans influences neuronales ou hormonales extrapulmonaires. Contrairement aux systèmes de culture cellulaire, par exemple préparés à partir de cellules musculaires lisses artérielles pulmonaires canines17,l’architecture histologique de la paroi des vaisseaux est presque entièrement préservée.
En résumé, ce protocole fournit une méthode utile pour l’analyse des capteurs moléculaires potentiels d’oxygène et/ou des voies cellulaires responsables du VPH des artères intra-pulmonaires dont les diamètres intérieurs se sont compris entre 20 et 100 μm dans des conditions exemptes de contrainte de cisaillement.
Protocole
1. Préparation de mélanges, d’équipements, d’instruments et de solutions de gaz
Cette section décrit l’équipement nécessaire et la configuration pour le protocole. Des détails supplémentaires et des informations sur le fabricant se trouvent dans le tableau ci-joint.
- Obtenir ou préparer les mélanges de gaz suivants:
- Deux bouteilles avec mélange de gaz normoxique composé de 21% d’O2,5,3% deCO2,73,7% deN2.
- Une bouteille avec mélange de gaz hypoxique composé de 1%d’O2,5,3% deCO2,93,7%N2.
- Rassemblez l’équipement suivant :
- Un four à micro-ondes, pour faire fondre l’agarose.
- Une armoire chauffante, pour laver l’agarose des fentes pulmonaires (voir ci-dessous). Insérez un tube connecté à une bouteille avec un mélange de gaz normoxique dans l’armoire.
- Un vibratome avec des lames de rasoir appropriées, pour couper les poumons en tranches de 200 μm d’épaisseur. Il est avantageux que le vibratome soit muni d’un ensemble réfrigérant pour refroidir le tampon dans le bassin du vibratome.
- Une chambre de superfusion d’écoulement-à travers montée sur un microscope inversé, pour l’analyse du HPV des artères intrapulmonaires.
- Pour faciliter la fixation des sections pulmonaires au fond de la chambre, connectez les cordes en nylon à un anneau en platine (auto-construit). Joindre la chambre de perfusion à une pompe péristaltique avec des débits ajustés à 0,7 ml/min et 6 ml/min, respectivement.
- Assembler l’équipement de manière à ce que, pendant les expériences, les milieux soient stockés dans un bain-marie à 37 °C et barbotés avec du gaz normoxique ou hypoxique à l’aide de canules de 21 G x 4 3/4. En outre, utilisez une deuxième connexion de la bouteille de gaz pour permettre aux mélanges de gaz normoxiques / hypoxiques d’être introduits dans l’espace d’air de la chambre de perfusion. Assurez-vous que tous les tubes de ce système sont étanches aux gaz.
- Une caméra CCD montée sur un microscope inversé vertical pour enregistrer les images de l’artère analysées.
- Préparer les instruments suivants :
- Pour la préparation des poumons: un ensemble de dissection stérile comprenant un ciseau rugueux et deux paires de pinces, un ciseau fin pour l’ouverture de la poitrine, un microscisseur pour percer un trou dans la trachée pour remplir l’agarose, et coudre du coton (environ 20 cm) pour la ligature de la trachée afin d’empêcher l’écoulement de l’agarose.
- Pour le remplissage des voies respiratoires avec de l’agarose: connectez une seringue de 2 ml au tuyau en plastique flexible d’une canule à demeure IV (20 G x 1 1/4).
- Pour la perfusion des poumons avec tampon: fixer une seringue de 50 ml comme réservoir tampon à environ 40 cm au-dessus du lieu de travail pour la préparation de la souris.
- Pour l’écoulement du tampon: connectez la seringue à un tube auquel une canule de 25 G x 1 est attachée. Utilisez une pince au niveau du tube pour ajuster le débit sortant à environ 1 goutte/s (environ 0,3-0,4 ml/min).
- Préparez les mémoires tampons et les médias suivants :
- Faire 1 000 ml de tampon HEPES-Ringer (10 mM HEPES, 136,4 mM NaCl, 5,6 mM KCl, 1 mMMgCl2•6H2O, 2,2 mM CaCl2•2H2 O, 11 mM glucose, pH 7,4). Conserver le tampon filtré stérile (taille des pores du filtre : 0,2 μm) à 4 °C.
- Environ 30 min avant de commencer l’isolement des poumons, préparez les solutions suivantes:
- Dissoudre 1,5 % p/v d’agarose à faible point de fusion dans un tampon HEPES-Ringer (volume total 10 ml) et le faire fondre en le cuisinant au four à micro-ondes. Par la suite, refroidissez-le à 37 °C en le rangeant dans une armoire chauffante. Pré-warm la seringue de 2 ml pour remplir l’agarose dans les voies respiratoires pulmonaires.
- Prendre 20 ml du tampon HEPES-Ringer, ajouter l’héparine à une concentration finale de 250 I.U./ml et chauffer le tampon à 37 °C. Immédiatement avant l’utilisation, ajouter le nitroprusside de sodium à une concentration finale de 75 μM. C’est le tampon de perfusion pour s’enfuir du sang de la vascularisation pulmonaire.
- Mettez 200 ml de tampon HEPES-Ringer dans un bécher en verre et conservez-le sur de la glace. Ce tampon sera nécessaire pour refroidir les poumons isolés remplis d’agarose.
- Remplissez environ 200 ml de MEM complétés par 1% de pénicilline/streptomycine dans un bécher en verre et conservez-le à 37 °C dans l’armoire chauffante. Faire bouillonner le milieu avec un mélange de gaz normoxique. Il sera utilisé pour enlever l’agarose des sections pulmonaires.
- Prérudent 2 bouteilles de MEM complétées avec 1% de pénicilline/streptomycine au bain-marie jusqu’à 37 °C et vaporisez-les avec un mélange de gaz normoxique et hypoxique, respectivement, pendant au moins 2 heures avant de commencer les mesures vidéomorphométriques. Environ 250 ml de MEM seront nécessaires par mesure.
- Rassemblez les documents supplémentaires suivants :
- 70% EtOH pour la désinfection.
- Solution mère d’héparine (25.000 I.U./5 ml), conservée à 4 °C (voir ci-dessus).
- Solution mère du nitroprusside sodique donneur de NO (Nipruss) : 10 mM dansH2O, stocké sur de la glace (voir ci-dessus).
- Solution mère d’analogue de thromboxane U46619 : 10 μM dans de l’éthanol, stocké sur de la glace.
- Superglue.
- Coudre du coton pour la ligature de la trachée après remplissage à l’agarose.
2. Animaux
Utilisez des souris (par exemple de la souche C57Bl6) des deux sexes à l’âge de 10 à 25 semaines. Le VPH peut également être analysé dans les souches knockout et les souches de type sauvage correspondantes.
Toutes les expériences ont été réalisées conformément aux lignes directrices des NIH pour le soin et l’utilisation des animaux de laboratoire, et ont été approuvées par les conseils institutionnels locaux.
3. Isolement des poumons murins et préparation de tranches pulmonaires coupées avec précision (PCLS)
- Tuer la souris par luxation cervicale. Immédiatement après la mise à mort, stérilisez la surface ventrale du corps avec 70% EtOH et utilisez le ciseau rugueux pour couper la peau le long de la ligne médiane ventrale du menton au bassin.
Remarque: Comme on sait que les anesthésiques inhalatifs tels que les isoflurane ont un impact sur le tonus vasculaire18,19, n’utilisez pas d’anesthésiques volatils. - Après l’ouverture de la cavité abdominale, mettez les boucles intestinales de côté et sectionz les gros vaisseaux abdominaux pour les saignements. Après avoir pénétré dans le diaphragme avec le ciseau fin, les poumons s’effondreront à la suite de l’entrée d’air dans la cavité pleurale. Utilisez le ciseau pour détacher le diaphragme de l’ouverture thoracique inférieure. Couper les côtes et la clavicule latéralement pour enlever la partie ventrale de la cage de déchirure.
Remarque: Il est important que les poumons ne soient pas endommagés dans cette étape car sinon les ballonnements des poumons avec de l’agarose seront impossibles! N’utilisez que des instruments stériles et de la verrerie de laboratoire. - Avant de commencer la perfusion de la vascularisation pulmonaire, coupez un petit trou dans le ventricule gauche du cœur pour la décharge du tampon. Remplissez le réservoir de seringue avec un tampon HEPES-Ringer chaud (37 °C) contenant de l’héparine et du nitroprusside de sodium (tampon de perfusion) et perfuser lentement la vascularisation pulmonaire via le ventricule droit.
Remarque: La perfusion est efficace lorsque les poumons changent de couleur et obtiennent un aspect blanc. Dans cette étape, il est important d’ajouter du nitroprusside de sodium dans le tampon de perfusion; cela empêche les artères pré-acinaires de s’arracher des tissus environnants. - Enlevez les glandes salivaires, les petits muscles et le tissu conjonctif de la trachée. Débranchez la trachée du tissu conjonctif environnant et fil cousez le coton entre l’œsophage et la trachée pour une ligature ultérieure.
- Utilisez le microscisseur pour couper un petit trou dans la partie supérieure de la trachée entre deux cartilages trachéaux voisins. Insérez maintenant le tuyau en plastique flexible d’une canule à demeure IV via le petit trou dans la trachée et fixez-le soigneusement avec le coton à coudre. Remplissez lentement les voies respiratoires avec l’agarose chaude (37 °C) à faible point de fusion. Observez les poumons: Au début, le poumon droit commence à se dilater, suivi du poumon gauche. Le remplissage est terminé lorsque les deux poumons sont gonflés à un volume comparable à la situation in vivo (environ 1,2 à 2,0 ml selon le sexe, l’âge et le poids).
Remarque : Si un seul poumon se dilate, le tuyau en plastique peut avoir été inséré trop profondément pour atteindre la bronche. Dans ce cas, il faut le retirer un peu. Gardez à l’esprit que l’agarose se solidifiera lorsqu’il se refroidira progressivement. - Lorsque les poumons sont pleins, retirez simultanément le tuyau en plastique et liez la trachée avec le coton à coudre pour empêcher l’écoulement de l’agarose. Par la suite, coupez la trachée au-dessus de la ligature et détachez les poumons et le cœur sur bloc de la poitrine.
Remarque: Pour les débutants, il peut être utile de demander de l’aide à un collègue à l’étape de ligature. - Transférer le paquet d’organes dans un tampon HEPES-Ringer glacé pour solidifier l’agarose. Cela se produit en quelques minutes.
- Séparez les lobes pulmonaires individuels et fixez un lobe avec une superglue sur le porte-échantillon du vibratome.
Remarque: Il est utile de coller un morceau de liège de champagne sur le support qui sert de skewback lors de la coupe du tissu pulmonaire élastique. En fonction du lobe pulmonaire utilisé et de son orientation sur le porte-échantillon, le PCLS obtenu peut être plus adapté à l’analyse de petits ou grands vaisseaux. La plupart du temps, nous utilisons le lobe gauche et le lobe crânien droit pour la préparation du PCLS. Pour obtenir de petites artères intra-acinaires en coupe transversale, collez le lobe droit crânien avec le hile au support et coupez de la périphérie. Pour obtenir des artères pré-acinaires sectionnés, alignez le hilum du lobe droit avec le liège de champagne. - Utilisez un vibratome équipé d’une lame de rasoir fraîche pour couper le lobe pulmonaire en tranches épaisses de 200 μm (vitesse : 12 = 1,2 mm/s ; fréquence : 100 ; amplitude : 1,0). Recueillir le PCLS dans le bassin vibratome rempli de 4 °C hepes froid-sonneur tampon.
Remarque: Le refroidissement de la sonnerie HEPES est recommandé mais non essentiel. - Pour l’élimination de l’agarose, transférer les sections d’organes dans un bécher en verre rempli d’environ 200 ml de MEM chaud à 37 °C. Mettez le bécher dans l’armoire chauffante dans laquelle est inséré un tube joint à une bouteille avec un mélange de gaz normoxique. Faites bouillonner le MEM avec le gaz normoxique de sorte que les sections pulmonaires se déplacent lentement dans le milieu. Après environ 2 heures, les « plaques d’agarose » remplissant les espaces aériens seront retirées du tissu pulmonaire. Cela peut être reconnu par le fait que les sections ne nagent plus sur le dessus du milieu mais s’installent sur le bas du bécher.
4. Analyse vidéomorphométrique des artères intrapulmonaires de PCLS
- Pour l’analyse vidéomorphométrique des artères intra-pulmonaires, transférer un PCLS dans la chambre de superfusion à écoulement traversant remplie de MEM gazé normoxique de 1,2 ml. Fixez le PCLS au bas de la chambre avec des cordes en nylon reliées à un anneau en platine (diamètre extérieur / intérieur: 14/10 mm).
- Balayer le PCLS au microscope pour les artères transversales avec des diamètres intérieurs compris entre 20-100 μm.
Remarque: La lumière des artères est bordée de cellules endothéliales plates. Les cellules musculaires lisses environnantes peuvent être identifiées dans l’image de contraste de phase comme un « anneau sombre » entourant la lumière (voir les images de contraste de phase dans Figure 2). En revanche, des voies aériennes peuvent être identifiées par l’épithélium co-co-laryngé initialement pseudostratified qui transite sur le chemin de la surface pleurale dans un épithélium conaire simple suivi d’un épithélium cubique simple.- Mesurer le diamètre intérieur au début de chaque expérience.
Remarque: Dans les vaisseaux légèrement coupés obliquement, le diamètre intérieur réel de la structure tubulaire peut être déterminé dans un angle de 90 ° par rapport à l’axe le plus long de la lumière. De grandes artères pré-acinaires de diamètre intérieur >40 μm sont adjacentes aux bronches et aux bronchioles. De petites artères intra-acinaires d’un diamètre intérieur <40 μm sont situées aux goussets de septa alvéolaires à côté des alvéoles et des conduits alvéolaires6.
- Mesurer le diamètre intérieur au début de chaque expérience.
- Conception expérimentale :
- Commencez chaque expérience par une « phase d’adaptation » dans laquelle la chambre est perfusée avec un milieu gazé normoxique (débit: 0,7 ml / min; 10 min). Ensuite, tester la viabilité du vaisseau : Analyser la contractilité de l’artère par l’ajout de 12 μl de 10 μM U46619 (concentration finale 0,1 μM ; 10 min ; pas d’écoulement).
Remarque : Pour ce travail, un récipient est défini comme viable lorsque la surface luminale est réduite d’au moins 30 % (pour la méthode de mesure, voir ci-dessous). - Après avoir lavé le médicament avec un milieu gazé normoxique (débit: 6 ml/min; 10 min), dilatez l’artère par application de 3 μl de Nipruss 10 mM (concentration finale 25 μM; 10 min; pas de débit).
- Encore une fois, retirez le médicament par un lavage de 10 min avec un milieu gazé normoxique (débit: 6 ml/min) suivi de 10 min à un débit de 0,7 ml/min.
- Induire une vasoconstriction pulmonaire hypoxique par incubation du PCLS avec milieu gazé hypoxique (débit : 0,7 ml/min ; 40 min). Par un système de tube supplémentaire, alimentez le mélange de gaz hypoxique dans l’espace d’air de la chambre de perfusion.
- Retirer le milieu hypoxique par un lavage de 20 min avec un milieu gazé normoxique (débit : 6 ml/min).
- À la fin de chaque expérience, ajouter 1,2 μl de 10 μM U46619 (concentration finale 0,01 μM; 20 min; pas de débit) à la chambre de perfusion pour induire une vasoconstriction.
Remarque: Cette dernière étape permet de déterminer si un changement dans la réponse hypoxique (par exemple induit par l’application simultanée d’un médicament dans la phase hypoxique ou observé dans un PCLS préparé à partir d’une souche de souris knockout) est spécifique à la constriction induite par l’hypoxie ou s’il reflète un impact général sur la contractilité. La concentration de 0,01 μM correspond à la valeur CE50 de l’U46619 qui a été estimée dans les mesures concentration-contraction précédentes (non publiées). - Utilisez un autre PCLS pour effectuer des expériences de contrôle avec le MEM gazé hypoxique au lieu du mélange de gazmeux hypoxique gazés et de gaz normoxique dans l’espace de l’air de la chambre de perfusion.
- Commencez chaque expérience par une « phase d’adaptation » dans laquelle la chambre est perfusée avec un milieu gazé normoxique (débit: 0,7 ml / min; 10 min). Ensuite, tester la viabilité du vaisseau : Analyser la contractilité de l’artère par l’ajout de 12 μl de 10 μM U46619 (concentration finale 0,1 μM ; 10 min ; pas d’écoulement).
5. Analyse de la vasoreactivité et présentation graphique
- À l’aide d’un logiciel approprié, prenez des photos de l’artère sectionnée toutes les minutes pendant toute l’expérience.
- À l’aide d’un logiciel approprié, évaluez les changements de la zone luminale des navires en tapissant les limites intérieures à la main à l’aide d’images en plein écran.
Remarque: Il suffit d’analyser une image sur deux pour obtenir des graphiques clairs. Cependant, lorsqu’une condition dans la chambre de superfusion à écoulement est remplacée par une autre, analysez chaque image. Les images restantes servent de sauvegarde au cas où une photographie ne peut pas être analysée.
Malheureusement, cette étape fastidieuse doit être faite à la main et non par des programmes appropriés car parfois les cellules sanguines sont attachées à la paroi vasculaire qui se déplacent pendant la mesure ou ce n’est pas une section transversale parfaite du vaisseau mais une section tangentielle dans laquelle une localisation fiable des limites internes ne peut être faite que sous contrôle visuel. - Définir la valeur obtenue pour la zone de la lumière du vaisseau au début de l’expérience comme étant de 100% et exprimer la vasoconstriction ou la dilatation comme diminution ou augmentation relative de cette valeur.
- Pour chaque expérience, tracez la zone luminale relative en fonction du temps à l’aide d’un logiciel approprié.
- Pour résumer plusieurs expériences, tracer les moyennes des aires luminales relatives +/- erreur type de la moyenne (MEB) par rapport au temps.
Remarque: Pour une présentation claire des effets de diverses substances sur l’hypoxie- ou U46619-induite vasoconstriction pulmonaire ou de la réponse hypoxique des souches de souris knockout, la phase initiale des expériences, dans lequel la viabilité du vaisseau est testée, peut être omise du graphique. Dans ce cas, les valeurs obtenues au début de l’exposition à l’oxygène réduit sont définies comme 100%.
6. Analyse statistique
- Analyser les différences entre les groupes expérimentaux avec le test de Kruskal-Wallis- et le test de Mann-Whitney, p≤0,05 étant considéré comme significatif, et p≤0,01 très significatif.
Note: Pour plus d’informations sur la préparation et l’utilisation opérationnelle de PCLS voir aussi20,21.
Résultats
Sur la figure 1, les résultats de la mesure du VPH d’une grande artère pré-acinnaire et sur la figure 2 de petites artères intra-acinaires sont donnés. Dans les images de contraste de phase(figures 1A et 2A),il devient clair qu’il est possible de distinguer ces 2 classes d’artères en fonction de leur emplacement dans le tissu pulmonaire: les artères pré-acinales courent à proximité des bronches et des bronchioles(figure 1A),tandis que les artères intra-acinaires sont situées à des goussets de septa alvéolaires et entourées d’alvéoles(figure 2A). Avec un peu de pratique, il est possible de voir les changements dans la zone luminale en réponse à U46619 sur les images de contraste de phase(figures 1A et 2A). Cependant, la vasoconstriction pulmonaire hypoxique n’est souvent pas aussi prononcée et ne devient claire qu’après une évaluation complète des changements des zones luminales(figures 1B, 1C, 2B et 2C). Pour une raison didactique, nous avons donné un exemple d’une artère pré-acinnaire qui montre une vasoconstriction inhabituellement prononcée. En moyenne, le VPH entraîne une réduction de 20 à 30 % de la région luminale.
Sur la figure 2, les enregistrements de petites artères intra-acillaires incubées avec un milieu hypoxique gazé avec ou sans pinacidil de 50 μM (un ouvreur non sélectif de canaux potassiques sensibles à l’ATP mitochondrial) sont montrés et l’effet inhibiteur du médicament devient clairement visible. La dernière partie de l’expérience démontre la sélectivité de l’action du médicament sur le VPH : La vasoconstriction induite par l’analogue du thromboxane U46619 n’est pas modifiée par l’ajout de pinacidil. En effet, la courbe de l’artère exposée au pinacidil est nettement inférieure à celle des deux autres, mais l’étendue de la réduction de la zone luminale par U46619 seul est comparable. Dans ce cas, c’est la réversion incomplète du VPH qui cause la différence entre les courbes. Dans ce graphique également une artère exposée au milieu gazé normoxique est incluse comme contrôle supplémentaire. Dans cette condition, aucun changement de la zone luminale n’est détectable.
La figure 3 montre les données de la série complète de mesures sur l’influence du pinacidil sur le VPH. Pour comparer les deux groupes pour les différences statistiques, les ensembles de données des points temporels indiqués ont été analysés avec le test de Kruskal-Wallis et le test de Mann-Whitney. HPV a été clairement supprimé en présence du pinacidil tandis que la contraction induite par U46619 était inchangée.
Alternativement, il est possible d’identifier les différences entre les groupes par comparaison de l’aire sous la courbe comme décrit dans Müller-Redetzky et al. 22 ans

Figure 1. Mesure du HPV d’une grande artère pré-acinnaire. (A) Images de contraste de phase d’une artère pré-acinnaire sectionnée (a) qui court dans le voisinage proche d’une bronche en coupe transversale(B). Les photos sont prises aux points de temps indiqués en(B)par des cercles : au début de la mesure (a), à la fin du traitement avec U46619 (b), à la fin de l’exposition à Nipruss (c), après 30 ou 40 min en milieu gazé hypoxique (d, e), après lavage avec un milieu gazé normoxique (f), et après l’application finale d’U46619 (g). Dans le graphique(B),les changements de la zone luminale sont tracés en fonction du temps, tandis que la zone luminale au début de l’expérience est définie comme 100% et la vasoconstriction / dilatation est donnée comme valeurs relatives. Dans ce cas, l’hypoxie induit une réduction de 60% de la zone luminale. (C) Pour une présentation plus claire de la réponse hypoxique, la phase initiale de l’expérience dans laquelle la vasoreactivity a été testée n’est pas incluse, mais la valeur obtenue immédiatement avant l’exposition à l’oxygène réduit est fixée à 100% (voir aussi figure 2). Cliquez ici pour agrandir l’image.
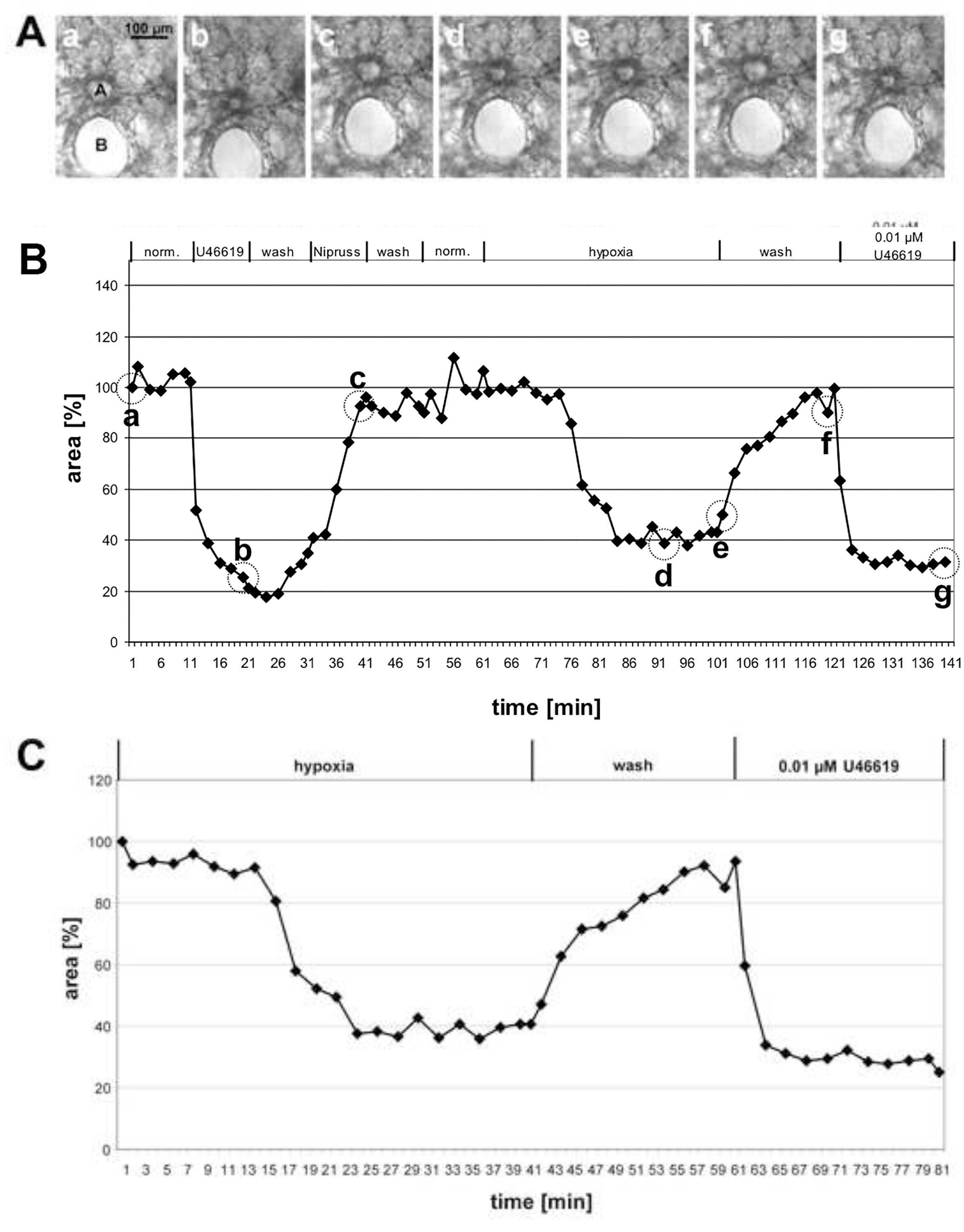{kind=link}

Figure 2. Impact du pinacidil (un ouvreur non sélectif des canaux potassiques sensibles à l’ATP mitochondrial; mitoKATP) sur le VPH de petites artères intra-acinaires. (A) Les artères intra-acinaires (a) sont situées au niveau des goussets des septa alvéolaires. Alv = alvéole. La séquence des conditions individuelles appliquées dans ces expériences est donnée dans le titre du graphique(B). L’exposition hypoxique est réalisée en présence ou en l’absence de pinacidil de 50 μM. Les incubations de contrôle sont effectuées avec un milieu gazé normoxique. Les photos représentées au point(A)sont prises au début de la mesure (a, a', a »), à la fin du traitement avec U46619 (b, b', b »), à la fin de l’exposition au Nipruss (c, c', c »), après 30 ou 40 min en milieu gazé hypoxique ou normoxique (d, d', d »; e, e', e »), après lavage avec un milieu gazé normoxique (f, f', f »), et après l’application finale d’U46619 (g, g', g »). Pour une présentation plus claire de la réponse à la normoxie/hypoxie/hypoxie+pinacidil, les valeurs obtenues immédiatement avant l’exposition au milieu normoxique/hypoxique gazé sont fixées à 100% (C). Cliquez ici pour agrandir l’image.
{kind=link}

Figure 3. Inhibition de HPV par pinacidil. Les enregistrements de petites artères intra-acinaires exposées à un milieu gazé hypoxique avec ou sans pinacidil de 50 μM sont résumés et présentés comme moyens ± MEB. Dans (A) les enregistrements complets sont donnés, dans (B) les données relatives par rapport à la valeur au début de l’incubation hypoxique sont indiquées. Aucune vasoreactivity n’est détectable dans les PCLS qui sont exposés à un milieu hypoxique gazé contenant du pinacidil. La vasoconstriction induite par U46619 n’est pas affectée par le médicament. « n » entre parenthèses fait référence au nombre d’artères/nombre d’animaux à partir desquels des PCLS ont été fabriqués. En d’autres termes, le premier chiffre décrit le nombre de sections pulmonaires analysées et le deuxième nombre donne le nombre de souris à partir desquelles ces sections ont été préparées. Aux moments donnés, la signification des différences entre les deux groupes est testée. n.s. : non significatif, *: p≤0,05, **: p≤0,01, *** : p≤0,001. Cliquez ici pour agrandir l’image.
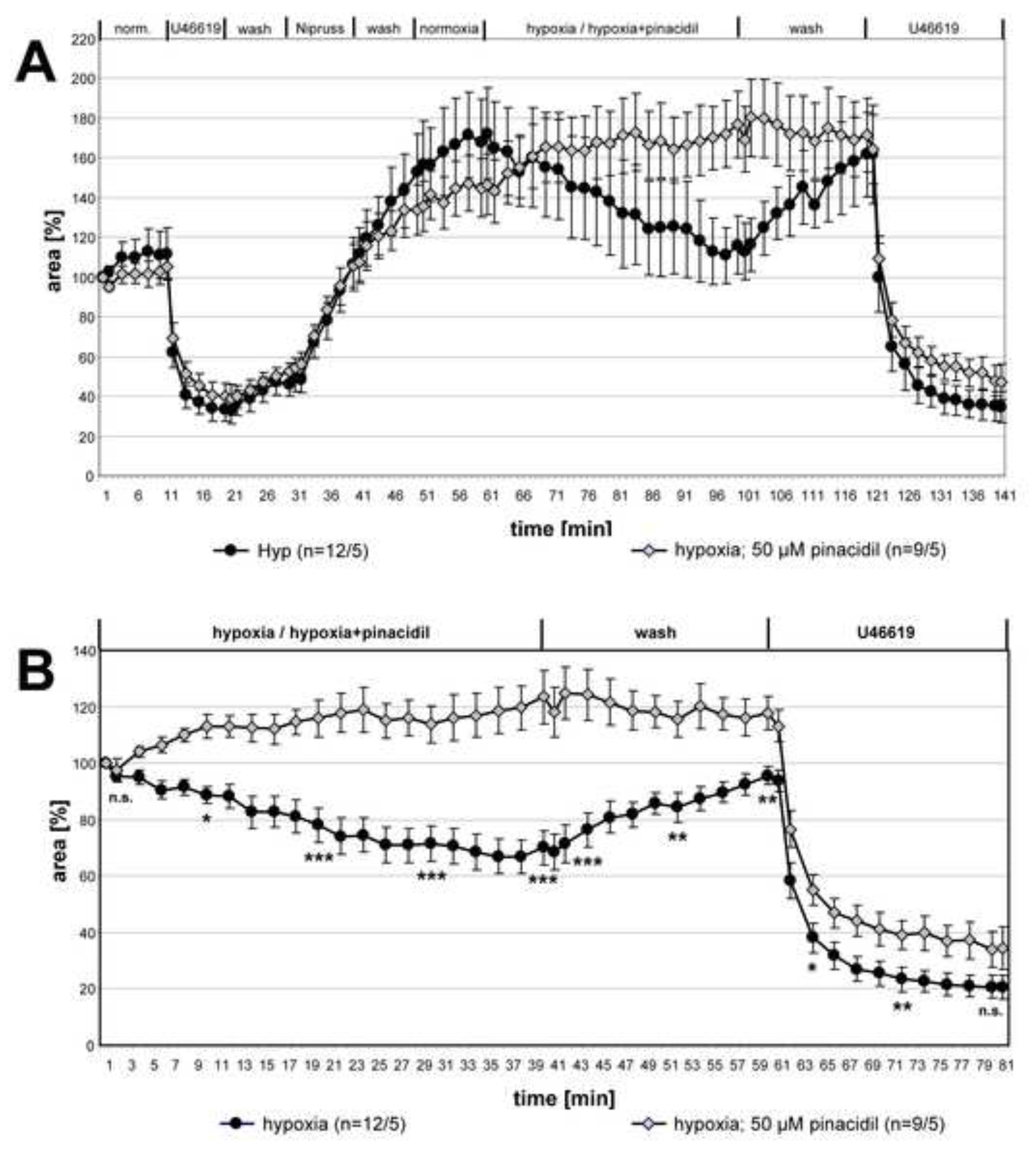{kind=link}

Figure 4. Vue d’ensemble schématique de la méthode. En bref, les souris sont tuées par luxation cervicale. Après l’ouverture de la poitrine, les poumons sont remplis d’agarose à faible point de fusion et après refroidissement, coupés en tranches pulmonaires coupées de précision de 200 μm d’épaisseur (PCLS). Après élimination de l’agarose à 37 °C, un PCLS est transféré dans la chambre de superfusion d’écoulement dans laquelle il est exposé au milieu normoxique ou au milieu gazé à 1 %d’O2. La vasoreactivity est enregistrée comme des changements dans la zone luminale. Cliquez ici pour agrandir l’image.
{kind=link}
Discussion
Le poumon de souris isolé ventilé et perfusé est un excellent modèle pour l’analyse de la réponse physiologique du système vasculaire pulmonaire sur les changements d’apport en oxygène et permet entre autres la mesure continue de la pression artérielle pulmonaire1. Cependant, ce modèle ne permet pas l’identification et l’analyse de ces segments vasculaires montrant la réponse la plus forte à l’hypoxie. C’est l’avantage de notre analyse vidéomorphométrique du PCLS qui facilite la mesure du VPH d’artères individuelles avec des diamètres intérieurs de 20-100 μm. Les PCLS représentent un modèle in vitro attrayant car ils ressemblent étroitement à l’organe à partir duquel ils sont préparés. Contrairement aux systèmes de culture cellulaire, tous les types de cellules sont présents dans leur configuration tissu-matrice d’origine. En outre, un poumon suffit pour la préparation de nombreux PCLS, de sorte qu’au moins partiellement les expériences peuvent être normalisées par l’utilisation de sections de la même souris. Selon le concept 3R (réduction, raffinement et remplacement des animaux de laboratoire dans les sciences de la vie) de Russell et Burch23, ce fait plaide également en faveur de l’utilisation de PCLS.
Cependant, il faut garder à l’esprit, que le tissu est endommagé par la coupe avec un vibratome et la signalisation longitudinale par exemple via les cellules endothéliales comme postulé par Kübler et al. 14 n’est plus possible.
Initialement, les PCLS ont été principalement appliqués pour des études biochimiques, pharmacologiques et toxicologiques, mais en attendant, ils sont également utilisés pour mesurer la contractilité bronchique, la fonction mucociliaire et les réponses vasculaires (pour les examens, voir Sanderson20 et Davies21). Held et al. ont réalisé une étude dans laquelle ils ont comparé les modèles de poumon de souris perfusé et ventilé isolé et de PCLS24. Ils ont constaté par analyse des réponses des voies aériennes et des vaisseaux pulmonaires à une série de médiateurs endogènes que des caractéristiques importantes du poumon entier ont été maintenues dans PCLS.
Dans PCLS, les conditions hypoxiques ne sont pas établies par l’intermédiaire des voies respiratoires comme dans le poumon intact mais par incubation de la section pulmonaire dans le milieu hypoxique-gazé. Nous avons analysé la pression partielle d’oxygène(pO2)du milieu prégazé avec 1%O2,5,3%CO2,93,7% N2 et avec 21%O2, 5,3%CO2,73,7%N2,respectivement, à l’aide d’un analyseur de gaz sanguin. Immédiatement avant de l’alimenter dans la chambre de perfusion, lepO2 du MEM gazé hypoxique était de 40 mmHg et celui du milieu gazé normoxique de 160 mmHg6. Dans le poumon intact, le VPH est induit lorsque le pO alvéolaire2 tombe en dessous de 50 mmHg25, une situation qui peut évidemment être imitée par l’application d’un milieu hypoxique gazé. Nos données sur l’étendue du VPH correspondent bien aux résultats obtenus avec une approche expérimentale différente. Yamaguchi et al. ont appliqué des poumons de rat isolés pour examiner des microvaisseaux d’un diamètre de 20-30 μm par microscopie à luminescence à balayage laser confocal en temps réel couplée à une caméra haute sensibilité avec un intensificateur d’image10. Ils ont observé une réduction moyenne du diamètre de 2,7 μm après exposition des poumons à l’hypoxie. On peut calculer qu’une réduction de 20% de la surface luminale telle que nous la mesurons dans notre système correspond à une diminution d’environ 15% du diamètre.
Dans nos expériences, nous avons classé les artères comme des vaisseaux pré- et intra-acinaires, respectivement, avec des diamètres intérieurs de 40-100 μm et 20-40 μm. Chez l’homme, la transition des artères musculaires aux artères non musculaires se produit dans la gamme de diamètre de 70-100 μm. Chez la souris, les cellules musculaires lisses sont présentes jusqu’à un diamètre extérieur de 20 μm26. Pour cette raison, il n’est pas possible d’analyser les artères avec des diamètres inférieurs à 20 μm car elles ne peuvent pas être identifiées de la fiabilité sur la base de l’image de contraste de phase. À l’autre extrémité de l’échelle, les vaisseaux d’un diamètre supérieur à 100 μm sont à peine à trouver dans le PCLS et généralement dépouillés des tissus environnants.
En fait, un certain nombre de candidats moléculaires sont discutés en tant que capteur(s) moléculaire(s) d’oxygène ou en tant que composant de la cascade de signalisation entraînant le VPH (pour une revue, voir Sylvester et al. 4). Une fois que les souris knockout appropriées sont disponibles, la vidéomorphométrie peut être utilisée pour l’analyse de la vasoreactivity des artères pré- et intra-acinaires par rapport aux animaux de type sauvage. Cependant, PCLS ont également été utilisés pour d’autres questions: Faro et al. on les employait pour caractériser le développement de la dilatation dépendante de l’endothélium dans le poumon après la naissance29 et le PCLS préparé à partir de cobayes exposés quotidiennement à la fumée ou à l’air pendant 2 semaines pour démontrer l’impact de la fumée de cigarette sur la vasoreactivity par induction d’un dysfonctionnement endothélial30.
Étapes critiques du protocole
Dans nos expériences, nous avons classé les artères comme pré-acinaires (diamètres intérieurs de 40-100 μm) et intra-acinaires (diamètres intérieurs de 20-40 μm). En particulier pour la préparation des sections pulmonaires qui doivent être utilisées pour l’analyse de vaisseaux plus gros, il est important d’ajouter du nitroprusside de sodium au tampon de perfusion. Ce médicament empêche la contraction des vaisseaux pendant la préparation de l’échantillon et donc leur arrachement du tissu environnant conduisant à une vasodilatation incomplète. Le nitroprusside de sodium dans le tampon de perfusion n’est pas si important pour la préparation de la section pulmonaire qui devrait être utilisée pour l’analyse des petites artères parce qu’elles sont fortement ancrées aux septa alvéolaires.
Toutes les expériences doivent être commencées par des incubations dans lesquelles la réactivité des artères est testée. Rarement, nous avons obtenu des préparations de poumon dans lesquelles aucune réponse des navires aux entrepreneurs ou aux dilatateurs n’était discernable. Nous ne connaissons pas la raison de cela: peut-être que le volume de l’agarose rempli dans les poumons était trop grand ou trop faible de sorte que la coupe de l’organe en PCLS n’était pas optimale. Alternativement, il est imaginable que l’agarose se refroidissait trop vite pendant la procédure d’instillation, ce qui entraînait un stress de cisaillement dommageable. Dans le cas où dans un PCLS individuel aucune artère viable n’est détectable, la section doit être écartée et remplacée par une autre.
La décision sur la viabilité d’une artère a été prise basée sur la réponse à U46619. L’application d’U46619 à une concentration de 0,1 μM induit une vasoconstriction qui - après un certain exercice - est visible directement dans la séquence d’images à l’écran. Puisqu’il y a quelques variances dans la vasoreactivity nous étudions l’impact d’un médicament sur HPV en mesurant la vasoresponse dans les sections pulmonaires exposées au médicament ou au milieu seul à son tour.
Le VPH d’une artère individuelle est souvent à peine détectable au microscope et, en moyenne, il en résulte une réduction de la zone luminale d’environ 20 à 30%. Cependant, de petits changements dans le diamètre d’une artère ont une entrée distincte sur la résistance à l’écoulement. Selon l’équation « R = 1/r4 »avec R=résistance et r=rayon, la résistance d’écoulement est inversement proportionnelle à la quatrième puissance du rayon. Permettez-moi de donner un exemple: Une « artère idéale » présentant une section circulaire d’un diamètre de 40 μm (r = 20 μm) a une surface luminale d’environ 1 260 μm2. Lorsque la surface luminale est réduite de 20%, nous pouvons calculer que le diamètre du vaisseau est réduit de 10,5% à 35,8 μm (r= 17,9 μm). Selon l’équation donnée ci-dessus, la résistance à l’écoulement de ce navire passerait de 6,25 x10 -6 à 9,71 x 10-6 soit environ 55%. En cas de réduction de la surface luminale de 30%, le rayon diminuerait d’environ 16%, mais la résistance à l’écoulement augmenterait d’environ 100%. Bien que ces calculs soient une simplification excessive dans laquelle un flux sanguin laminaire et une forme de vaisseau d’un tuyau rigide sont supposés, il est suggestif de l’impact de changements déjà mineurs du diamètre sur la résistance à l’écoulement.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Cette recherche est parrainée par le système cardio-pulmonaire Excellence Cluster.
matériels
| Name | Company | Catalog Number | Comments |
| Vibratome "Microm HM 650 V" | Microm/Thermo Fisher Scientific, Schwerte, Germany | ||
| Microwave oven | Bosch, Frankfurt, Germany | HMT 702C | |
| Heating cabinet | Heraeus/Thermo Fisher Scientific, Schwerte, Germany | ||
| Flow-through superfusion chamber | Hugo Sachs Elektronik, March, Germany | PCLS-Bath Type: 847 SN:4017 | |
| Upright inverted microscope equipped with 4X, 10X, 20X, and 40X objectives | Leica, Wetzlar, Germany | ||
| CCD-camera | Stemmer Imaging, Puchheim, Germany | ||
| Peristaltic pump Minipuls 3 | Gilson, Limburg-Offheim, Germany | ||
| Water bath “Universal Wasserbad Isotem 205” | Thermo Fisher Scientific, Schwerte, Germany | 9452450 | |
| Gas tight tubes Tygon R3603-13 Øi: 3/32 in, Øa: 5/32 in, wall: 1/32 in | VWR, Darmstadt, Germany | ||
| Various scissors and forceps | |||
| Sewing cotton | |||
| 2 ml Syringe | Braun-Melsungen AG, Melsungen, Germany | ||
| 50 ml Syringe | Braun-Melsungen AG, Melsungen, Germany | ||
| Flexible plastic pipe of an IV indwelling cannula “IntrocanR-W” (cannula 20 G x 1 ¼ in, 1.1 x 32 mm) | Braun-Melsungen AG, Melsungen, Germany | 4254112B | For instillation of the agarose into the lung |
| Cannula 21 G x 4 ¾ in; 0.8 x 120 mm | Braun-Melsungen AG, Melsungen, Germany | 4665643 | For bubbling of the medium |
| Cannula Nr. 17, 24 G x 1, 0.55 x 25 mm | Terumo, Eschborn, Germany | NN 2425 88DSF18 | For lung perfusion |
| Normoxic gas mixture (21% O2, 5.3% CO2, 73.7% N2) | Linde, Hildesheim, Germany | ||
| Hypoxic gas mixture (1% O2, 5.3% CO2, 93.7% N2) | Linde, Hildesheim, Germany | ||
| HEPES | Sigma, Deisenhofen, Germany | H 4034 | |
| NaCl | Roth, Karlsruhe, Germany | 3957.1 | |
| KCl | Merck, Darmstadt, Germany | 1.04936.0500 | |
| MgCl2•6H2O | Merck, Darmstadt, Germany | 1.05833.0250 | |
| CaCl2•2H2O | Merck, Darmstadt, Germany | 1.02382.0500 | |
| Glucose D-(+) | Sigma, Deisenhofen, Germany | G 7021 | |
| Low melting point agarose | Bio-Rad, Munich, Germany | 161-3111 | |
| Heparin-sodium | Ratiopharm, Ulm, Germany | 5120046 | |
| Phenolred-free minimal essential medium (MEM) | Invitrogen, Darmstadt, Germany | 5120046 | |
| 70% EtOH for desinfection | Stockmeier Chemie, Dillenburg, Germany | ||
| Superglue | UHU, Bühl/Baden, Germany or from a supermarket | ||
| U46619 (a thromboxane analog) | Calbiochem/Merck, Darmstadt, Germany | 538944 | |
| Sodium nitroprusside (Nipruss) | Schwarz Pharma, Monheim, Germany | 5332804 | |
| Optimas 6.5 software | Stemmer, Puchheim, Germany | ||
| SPSS 19 | AskNet, Karlsruhe, Germany |
Références
- Weissmann, N., Akkayagil, E., et al. Basic features of hypoxic pulmonary vasoconstriction in mice. Respir. Physiol. Neurobiol. 139, 191-202 (2004).
- Leach, R. M., Hill, H. M., Snetkov, V. A., Robertson, T. P., Ward, J. P. T. Divergent roles of glycolysis and the mitochondrial electron transport chain in hypoxic pulmonary vasoconstriction of the rat: identity of the hypoxic sensor. J. Physiol. 536, 211-224 (2001).
- Tabuchi, A., Mertens, M., Kuppe, H., Pries, A. R., Kuebler, W. M. Intravital microscopy of the murine pulmonary microcirculation. J. Appl. Physiol. 104 (2), 338-3346 (2008).
- Sylvester, J. T., Shimoda, L. A., Aaronson, P. I., Ward, J. P. Hypoxic pulmonary vasoconstriction. Physiol. Rev. 92 (1), 367-520 (2012).
- Schumacker, P. T. Lung cell hypoxia: role of mitochondrial reactive oxygen species signaling in triggering responses. Proc. Am. Thorac. Soc. 8 (6), 477-4784 (2011).
- Paddenberg, R., König, P., Faulhammer, P., Goldenberg, A., Pfeil, U., Kummer, W. Hypoxic vasoconstriction of partial muscular intra-acinar pulmonary arteries in murine precision cut lung slices. Respir. Res. 29 (7), 93-109 (2006).
- Paddenberg, R., et al. Mitochondrial complex II is essential for hypoxia-induced pulmonary vasoconstriction of intra- but not of pre-acinar arteries. Cardiovasc. Res. 93 (4), 702-710 (2012).
- Wang, L., Yin, J., et al. Hypoxic pulmonary vasoconstriction requires connexin 40-mediated endothelial signal conduction. J. Clin. Invest. 122 (11), 4218-4230 (2012).
- Archer, S. L., Huang, J. M., et al. Differential distribution of electrophysiologically distinct myocytes in conduit and resistance arteries determines their response to nitric oxide and. 78, 431-442 (1996).
- Yamaguchi, K., Suzuki, K., et al. Response of intra-acinar pulmonary microvessels to hypoxia, hypercapnic acidosis, and isocapnic acidosis. Circ. Res. 82, 722-728 (1998).
- Bennie, R. E., Packer, C. S., Powell, D. R., Jin, N., Rhoades, R. A. Biphasic contractile response of pulmonary artery to hypoxia. Am. J. Physiol. 261(2 Pt. 1, 156-163 (1991).
- Glazier, J. B., Murray, J. F. Sites of pulmonary vasomotor reactivity in the dog during alveolar hypoxia and serotonin and histamine infusion). J. Clin. Invest. 50 (12), 2550-2558 (1971).
- Bhattacharya, J., Staub, N. C. Direct measurement of microvascular pressures in the isolated perfused dog lung. Science. 210, 327-328 (1980).
- Kuebler, W. M. Real-time imaging assessment of pulmonary vascular responses. Proc. Am. Thorac. Soc. 8 (6), 458-4565 (2011).
- Martin, C., Uhlig, S., Ullrich, V. Videomicroscopy of methacholine-induced contraction of individual airways in precision-cut lung slices. Eur. Respir. J. 9 (12), 2479-2487 (1996).
- Grimminger, F., Spriestersbach, R., Weissmann, N., Walmrath, D., Seeger, W. Nitric oxide generation and hypoxic vasoconstriction in buffer-perfused rabbit lungs. J. Appl. Physiol. 78, 1509-1515 (1995).
- Ng, L. C., Kyle, B. D., Lennox, A. R., Shen, X. M., Hatton, W. J., Hume, J. R. Cell culture alters Ca2+ entry pathways activated by store-depletion or hypoxia in canine pulmonary arterial smooth muscle cells. Am. J. Physiol. Cell Physiol. 294 (1), 313-323 (2008).
- Fehr, D. M., Larach, D. R., Zangari, K. A., Schuler, H. G. Halothane constricts bovine pulmonary arteries by release of intracellular calcium. J. Pharmacol. Exp. Ther. 277 (2), 706-713 (1996).
- Oshima, Y., Ishibe, Y., Okazaki, N., Sato, T. Isoflurane inhibits endothelium-mediated nitric oxide relaxing pathways in the isolated perfused rabbit lung. Can. J. Anaesth. 44 (10), 1108-1114 (1997).
- Sanderson, M. J. Exploring lung physiology in health and disease with lung slices. Pulm. Pharmacol. Ther. 24 (5), 452-465 (2011).
- Davies, J. . Replacing Animal Models: A Practical Guide to Creating and Using Culture-based Biomimetic Alternatives (Google eBook. , 57-68 (2012).
- Müller-Redetzky, H. C., Kummer, W., et al. Intermedin stabilized endothelial barrier function and attenuated ventilator-induced lung injury in mice). PLoS One. 7 (5), (2012).
- Russell, W. M. S., Burch, R. L. The Principles of Humane Experimental Technique. Universities Federation for Animal Welfare. , (1959).
- Held, H. D., Martin, C., Uhlig, S. Characterization of airway and vascular responses in murine lungs. Br. J. Pharmacol. 126 (5), 1191-1199 (1999).
- Köhler, D., Schönhofer, B., Voshaar, T. Pneumologie: Ein Leitfaden für rationales Handeln in Klinik und Praxis. Thieme Verlag. , 198-19 (2009).
- Will, J. The Pulmonary Circulation. in Health and Disease. Elsevier Science, p49. , (1987).
- Faro, R., Moreno, L., Hislop, A. A., Sturton, G., Mitchell, J. A. Pulmonary endothelium dependent vasodilation emerges after birth in mice. Eur. J. Pharmacol. 567 (3), 240-244 (2007).
- Wright, J. L., Churg, A. Short-term exposure to cigarette smoke induces endothelial dysfunction in small intrapulmonary arteries: analysis using guinea pig precision cut lung slices. J. Appl. Physiol. 104, 1462-1469 (2008).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.