
Method Article
Isolement des bactéries de forme L de l’urine à l’aide de la méthode de filtration
Dans cet article
Résumé
Nous présentons ici un protocole pour l’isolement des bactéries de forme L de l’urine, à l’aide d’une méthode de filtration. Des méthodes complémentaires pour la préparation du milieu de forme L, l’observation des formes L par microscopie à contraste de phase et l’induction de formes L dans des conditions de laboratoire sont également décrites.
Résumé
On pense que la transition des bactéries vers l’état de forme L joue un rôle possible dans l’évasion immunitaire et la persistance bactérienne pendant le traitement avec des antibiotiques ciblant la paroi cellulaire. Cependant, l’isolement et la manipulation des bactéries de forme L sont difficiles, principalement en raison de leur grande sensibilité aux changements d’osmolarité. Nous décrivons ici des protocoles détaillés pour la préparation du milieu de forme L, l’isolement des formes L de l’urine à l’aide d’une méthode de filtration, la détection des formes L dans les échantillons d’urine par microscopie à contraste de phase et l’induction des formes L in vitro. Les exigences exactes pour la survie et la croissance des formes L peuvent varier d’une souche à l’autre. Par conséquent, les méthodes présentées ici sont destinées à servir de lignes directrices de base pour l’établissement de protocoles en forme de L au sein de laboratoires individuels, plutôt que d’instructions précises. La méthode de filtration peut entraîner une réduction du nombre de formes L dans un échantillon et ne doit pas être utilisée pour la quantification. Cependant, c’est la seule méthode utilisée jusqu’à présent pour séparer efficacement les variants déficients en paroi cellulaire de leurs homologues à paroi et pour identifier les souches bactériennes, capables de changer de forme L chez les patients atteints d’infections des voies urinaires. La méthode de filtration a le potentiel d’être adaptée pour l’isolement des formes L chez les patients atteints d’autres catégories d’infections bactériennes et dans des échantillons environnementaux.
Introduction
Pratiquement toutes les bactéries sont entourées d’une structure appelée paroi cellulaire. La paroi est importante pour la protection contre les stress environnementaux, aide à une division régulière et donne forme aux bactéries1. Cependant, la paroi est également une cible pour certaines parties du système immunitaire et certains des antibiotiques les meilleurs et les plus utilisés, y compris la pénicilline 2,3. Malgré son importance, les bactéries à Gram positif et à Gram négatif peuvent parfois survivre sans la paroi 4,5,6,7,8. Si les conditions environnantes offrent une osmoprotection suffisante pour les empêcher d’éclater et que des agents ciblant la paroi cellulaire sont également présents, les bactéries peuvent passer à un état sans paroi, appelé forme L 4,5,6,7,8.
De nombreux rapports indiquent que le passage à un état en forme L et le retour à un état clos peuvent être importants in vivo en tant que mécanisme permettant aux bactéries de survivre à la fois à l’attaque du système immunitaire de l’hôte et au traitement avec des antibiotiques ciblant la paroi cellulaire 9,10,11,12,13,14,15. Une telle transition fournit potentiellement une stratégie puissante pour la récurrence de l’infection bactérienne 9,10,11,12,13,14,15. Comprendre la biologie de base des bactéries de forme L et leurs interactions avec l’hôte est essentiel pour déchiffrer leur rôle dans la pathogenèse. Cependant, la manipulation des bactéries de forme L est difficile.
Tout d’abord, en raison de l’absence de paroi cellulaire, les bactéries de forme L ont tendance à éclater en réponse aux changements d’osmolarité. De plus, les formes L se divisent de manière très irrégulière, ont des modèles de croissance imprévisibles (généralement beaucoup plus lents que leurs homologues à parois) et, selon la déformation, peuvent mieux se propager sur des milieux semi-liquides, plutôt que solides ou liquides. Toutes ces considérations rendent difficiles la quantification et la comparaison des taux de croissance. Différentes espèces bactériennes (ou même souches) ont des exigences métaboliques diverses pour la commutation et la croissance de la forme L. Par exemple, les formes L de certaines bactéries à Gram positif, qui dépendent de la respiration aérobie, sont plus sensibles aux espèces réactives de l’oxygène que leurs homologues à parois16.
L’induction de formes L dans des conditions de laboratoire et dans l’hôte est généralement provoquée par des agents ciblant la paroi cellulaire, tels que les antibiotiques et le lysozyme9. Un tel traitement pourrait n’entraîner qu’une perte partielle de la paroi cellulaire et, par conséquent, certaines bactéries à paroi (ou partiellement à paroi) pourraient être présentes dans les échantillons, ce qui rend difficile de distinguer si les résultats expérimentaux observés sont dus à la présence de formes L ou de formes à paroi de bactéries. La fréquence des formes L induites in vivo a tendance à être faible, ce qui signifie qu’elles peuvent être difficiles à trouver et à isoler. Enfin, en raison de leur morphologie polymorphe, les formes L peuvent être facilement confondues in situ avec des structures d’origine eucaryote, telles que des corps apoptotiques ou des granules divers.
Depuis leur découverte en 193517, plusieurs méthodes ont été développées pour manipuler les formes en L en laboratoire. La plupart d’entre eux reposent sur l’ajout d’un agent osmoprotecteur au milieu de croissance ; généralement un sucre ou un sel 9,10,11,12,13,14,15,16,17,18. Comme mentionné ci-dessus, les formes L apparaissent souvent côte à côte avec des bactéries à paroi dans les échantillons de patients et il peut être difficile de séparer les deux populations. Cependant, il a été démontré que, contrairement aux bactéries à parois, les formes L peuvent passer à travers un filtre de 0,45 μm en raison de leur flexibilité et de leurs tailles variables. Une méthode utilisant un tel filtre a été utilisée pour isoler les formes L de l’urine 10,19,20,21.
Nous présentons ici un protocole d’isolement des bactéries de forme L dans l’urine, à l’aide d’une méthode de filtration (Figure 1). Des protocoles complémentaires pour la préparation du milieu de forme L, l’observation microscopique des formes L et l’induction de formes L in vitro sont également décrits.
Protocole
1. Préparation du milieu en forme de L (LM)
- Pesez le saccharose (quantité ajustée pour atteindre une concentration finale de 0,58 M), le MgSO4 (concentration finale de 8 mM) et la perfusion cerveau-cœur (BHI) (quantité recommandée par le fournisseur) dans une bouteille en verre deux fois plus grande que le volume de milieu souhaité. Pour préparer 0,5 L de milieu, pesez 100 g de saccharose, 1 g de MgSO4 et 18,5 g de BHI et placez-les dans une bouteille en verre de 1 L. Cela permettra une remise en suspension plus facile des ingrédients avant l’autoclavage.
- Si un milieu solide ou semi-solide est nécessaire, ajoutez la quantité souhaitée d’agar. Pour les milieux solides, ajouter 5 g de gélose pour obtenir une concentration finale de 1 % dans 0,5 L de milieu. Pour les milieux semi-solides, ajouter 1 g de gélose pour obtenir une concentration finale de 0,2 % dans 0,5 L de milieu.
- Complétez avec de l’eau déminéralisée (DI) jusqu’au volume final souhaité.
- Fermez la bouteille et mélangez bien en secouant. Il n’est pas nécessaire de remettre complètement les ingrédients en suspension, assurez-vous simplement qu’ils ne sont pas agglutinés au fond de la bouteille. Desserrez le capuchon et l’autoclave sur un cycle de support sensible (115 °C pendant 15 min). Il est recommandé de mélanger les ingrédients avant l’autoclave et d’utiliser un cycle de milieu sensible pour éviter l’agglutination des ingrédients et la caramélisation du saccharose.
- Examinez visuellement le milieu après l’autoclavage. Il ne doit pas y avoir de touffes et le milieu doit être de couleur ambrée (figure 2A).
- Après l’autoclavage, laissez le fluide refroidir à une température qui permet de tenir confortablement la bouteille à la main.
- Si le milieu est nécessaire pour l’induction des formes L, ajoutez un antibiotique et/ou un agent lytique à ce stade (par exemple, la phosphomycine à une concentration finale de 400 μg/mL, la pénicilline G à 200 μg/mL, la D-cyclosérine à 400 μg/mL, l’ampicilline à 100 μg/mL, la moénomycine à 50 μg/mL, le lysozyme à 100 μg/mL ou la lysostaphine à 5 μg/mL). Établir empiriquement le type d’agent inducteur de la forme L (ou les agents s’il en faut plusieurs) et sa concentration pour chacune des espèces bactériennes testées.
- Pour préparer des aliquotes individuelles, portez des gants et travaillez dans une enceinte de sécurité microbiologique ou utilisez un bec Bunsen. Les milieux peuvent nécessiter une incubation prolongée et la stérilité est d’une importance capitale.
- Pour préparer des aliquotes de milieu solide, transférez 25 mL dans des boîtes de Pétri individuelles de 92/16 mm et laissez-les prendre. L’utilisation d’un pipeteur plutôt que de verser directement les plaques est recommandée pour réduire le risque de contamination. Ne pas trop sécher les plaques.
- Pour préparer des aliquotes semi-liquides, transférez 5 ml de milieu dans plusieurs récipients universels en polystyrène de 30 ml à l’aide d’une pipette et laissez-les prendre.
- Utilisez le support immédiatement ou conservez-le à 4 °C jusqu’à une semaine avant de l’utiliser. Vérifiez les recommandations du fabricant avant de stocker un milieu contenant des antibiotiques, car la concentration peut diminuer, en raison de la dégradation du composé avec le temps.
2. Isolement des formes L de l’urine
REMARQUE : Portez des gants et une blouse de laboratoire pendant les procédures de laboratoire. Travaillez dans une enceinte de sécurité microbiologique ou utilisez un bec Bunsen. Portez des lunettes de sécurité pendant la filtration.
- Au moins 1 h avant l’arrivée prévue des échantillons d’urine, essuyez la zone de travail avec de l’éthanol à 70 % et indiquez le nombre requis d’aliquotes de milieu LM semi-liquide.
REMARQUE : Une suspension des formes L d’E. coli ST14410 induite en laboratoire dans un milieu LM liquide, plutôt que dans des échantillons d’urine humaine, sera utilisée pour démontrer le protocole. - Essuyez à nouveau la zone de travail avec de l’éthanol à 70 % avant de traiter les échantillons. D’un côté de la paillasse, disposez le nombre souhaité de filtres coupés de 0,45 μL, de seringues de 20 ml et de plusieurs récipients universels stériles vides de 30 ml en polystyrène correspondant au nombre d’échantillons. Ajoutez plusieurs filtres de rechange en plus du nombre d’échantillons d’urine attendus, au cas où plus d’un filtre serait nécessaire pour traiter certains échantillons. Placez les aliquotes moyennes LM et les récipients universels stériles vides de 30 ml en polystyrène dans un support stable au milieu de la paillasse. Si l’examen microscopique doit être effectué en parallèle, préparez également des lames de microscope en verre, des lamelles de 22 x 22 mm, des pipettes de 2 μL et 1 mL, des pointes stériles de 10 μL et 1 mL, des tubes stériles de 1,5 mL et assurez-vous qu’une microcentrifugeuse de paillasse est disponible.
- Transportez les échantillons d’urine du donneur au laboratoire dès que possible après le don, afin d’éviter la détérioration potentielle des bactéries de forme L dans l’échantillon.
- Mettez des gants et des lunettes de protection, retirez les échantillons d’urine du sac de sécurité, vaporisez-les d’éthanol, essuyez-les et placez-les dans le support, à proximité des aliquotes du milieu LM et des récipients stériles en polystyrène de rechange de 30 ml. À partir de ce point, traitez un échantillon à la fois.
- Desserrez les bouchons d’un tube contenant du fluide LM, un récipient universel en polystyrène de rechange et un tube contenant de l’urine.
- Retirez la seringue de l’emballage, retirez le piston et placez-la à votre droite.
- Retirez le papier de sécurité de l’arrière de l’emballage du filtre et fixez fermement la seringue au filtre (gardez le filtre face vers le bas dans le support en plastique du fabricant pour vous assurer qu’il reste stérile).
- En travaillant rapidement mais soigneusement, retirez et jetez le capuchon du tube de milieu LM et placez le filtre avec la seringue sur le dessus du tube.
- Retirez le capuchon de l’échantillon d’urine et versez doucement ~10 ml dans la seringue (seulement 2 ml seront filtrés, mais l’excès de volume empêche les éclaboussures), en veillant à ce qu’il reste plus de 1 ml pour un examen microscopique. Seulement 0,5 mL sera utilisé, mais il n’est pas pratique de mesurer avec précision un tel volume lors de la filtration, donc un excès approximatif de volume est conservé.
- Placez très soigneusement le piston à l’arrière de la seringue. Une petite résistance peut être ressentie avant que le piston ne soit placé en position complètement opérationnelle et il est facile de renverser l’échantillon au-delà de ce point. Entraînez-vous plusieurs fois à utiliser de l’eau avant de traiter les échantillons d’urine.
- Appuyez doucement sur le piston et faites passer 2 ml d’urine à travers le filtre jusqu’à ce qu’une résistance croissante se fasse sentir. Veillez à ne pas appliquer trop de pression pour éviter que l’échantillon ne déborde et que le filtre ne se brise. Si l’échantillon est particulièrement dense et génère trop de résistance, faites passer l’échantillon à travers plusieurs filtres, plutôt que de le forcer à travers un seul.
- Tapotez doucement le filtre contre le tube contenant le média LM pour déloger tout échantillon filtré restant, soulevez le filtre et la seringue et placez rapidement le capuchon du récipient universel en polystyrène de rechange de 30 ml sur le tube contenant le milieu LM et l’urine filtrée, en veillant à ne pas toucher l’intérieur du tube avec le bord du bouchon. Jetez en toute sécurité le filtre, la seringue et le récipient en polystyrène de rechange.
- Incuber les échantillons en position stationnaire à 30 °C pendant 1 mois maximum. Cela permettra aux formes en L qui sont passées à travers le filtre de régénérer leurs parois et de commencer à se développer sous forme de bactéries à parois.
- Observez les échantillons tous les jours pour détecter des signes de croissance, en tenant les tubes contenant des échantillons filtrés contre une source lumineuse. Pour les échantillons positifs, la croissance apparaît généralement dans les 3 à 7 jours.
- Une fois que l’excroissance a été détectée, transférez l’échantillon dans une enceinte de sécurité microbiologique ou à proximité d’un bec Bunsen. Ouvrez l’échantillon et plongez-le soigneusement dans une boucle en plastique de 5 μL, en visant la zone de croissance, puis striez les échantillons sur un milieu solide BHI standard sans osmoprotection, dans le but d’obtenir des colonies uniques22.
- En même temps, examinez au microscope une petite quantité de l’échantillon, ainsi qu’une fraction de milieu semi-liquide (~5 μL), comme décrit à l’étape 3. Le transfert de l’échantillon sur une lame microscopique avec le milieu garantira que toutes les formes en L qui peuvent encore être présentes dans l’échantillon n’éclateront pas et pourront être détectées. Cependant, à ce stade, on s’attend à ce que la majorité des bactéries soient revenues à des formes paroises et apparaissent sous forme de bâtonnets réguliers ou de coccis.
- Incuber les échantillons striés sur les plaques BHI en position stationnaire à 37 °C pendant la nuit.
- Examinez pour détecter tout signe de croissance, prélevez une seule colonie dans chaque plaque à l’aide d’une boucle de 5 μL et inculquez dans 5 mL de BHI liquide. Incuber à 37 °C pendant la nuit, en secouant.
- Utilisez les cultures de nuit pour préparer des stocks de glycérol ou pour isoler l’ADN afin d’identifier l’espèce à l’aide d’une méthode préférée (par exemple, Qiagen DNeasy Blood & Tissue Kit).
3. Examen des échantillons d’urine pour la présence de formes L par microscopie à contraste de phase
- Placez les 0,5 ml restants de l’échantillon d’urine dans un tube de microcentrifugation et faites tourner à 8 000 x g pendant 1 min dans une microcentrifugeuse de paillasse.
- Retirez délicatement le surnageant, en laissant environ 20 μL et remettez la pastille en suspension en pipetant doucement 3 fois de haut en bas.
- Placez 1 μL sur une lame de microscope en verre et recouvrez d’une lamelle de 22 x 22 mm (pour examiner un échantillon de 5 μL (voir l’étape 2.16), utilisez une lamelle de 22 x 50 mm pour vous assurer que la lamelle adhère bien à la lame). Appuyez doucement sur la lamelle à l’aide d’un coton stérile pour créer une étanchéité et vérifier que la lamelle ne peut pas bouger. Jetez le coton.
- Examinez la lame à l’aide d’un microscope équipé d’un objectif à contraste de phase 100x pour détecter la présence de structures de type L (Figure 3).
REMARQUE : Pour confirmer l’origine bactérienne de structures de type L dans les échantillons de patients, il est recommandé d’examiner les échantillons par hybridation in situ fluorescente (FISH) à l’aide de sondes oligonucléotidiques spécifiques pour les séquences bactériennes10,23.
4. Induction des formes L in vitro
- Disposez les bactéries à paroi striée de votre choix en colonies individuellesde 22 sur une plaque contenant un milieu non osmoprotecteur (par exemple BHI) et incubez stationnaire à 37 °C pendant la nuit.
- Le lendemain, préparez le milieu osmoprotecteur LM, contenant le(s) agent(s) inducteur(s) de forme L de votre choix, comme décrit à l’étape 1. Préparez le nombre souhaité de plaques avec un milieu solide.
- À l’aide d’une boucle en plastique stérile de 5 μL ou d’un cure-dent, prélevez une quantité généreuse (~4-5 colonies) de la plaque BHI non osmoprotectrice incubée la veille.
- Striez les bactéries sur la plaque LM en utilisant la surface plane de la boucle (plutôt que le bord). Striez les bactéries d’un mouvement continu sur le bord de la plaque, puis tournez la plaque à 90°. Touchez la boucle jusqu’au bord de la première traînée et, avec un mouvement continu, étalez la traînée sur un quadrant de la plaque, en diluant progressivement les bactéries avec plusieurs traînées qui se chevauchent (la plaque entière peut être utilisée si vous le souhaitez. Si vous n’utilisez qu’un quadrant de la plaque, d’autres quadrants peuvent être utilisés pour plusieurs inductions).
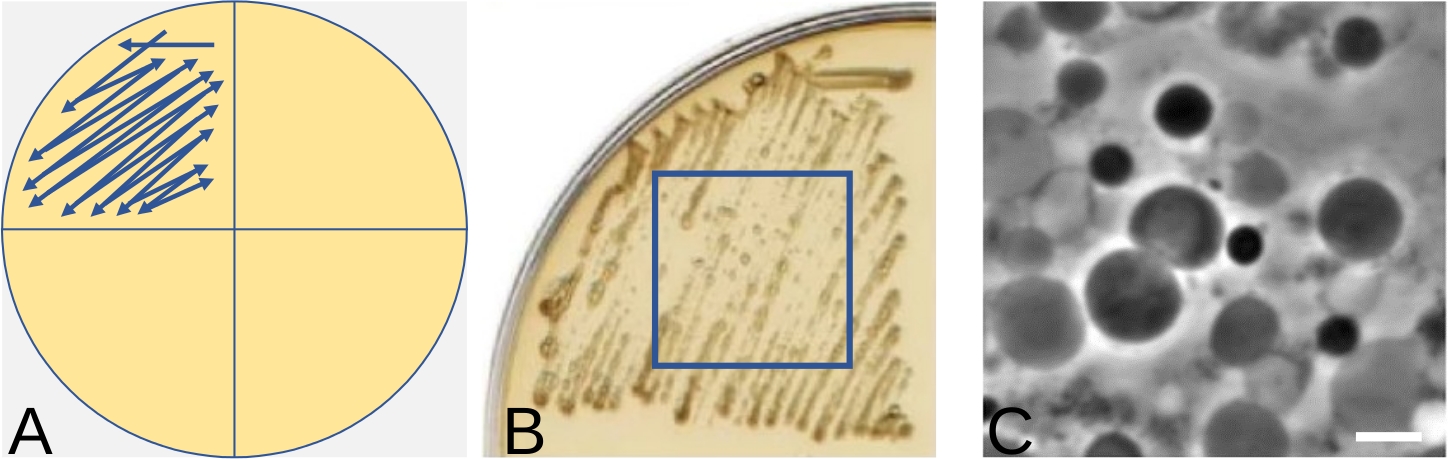
REMARQUE : La figure 4A montre schématiquement la direction des stries. L’objectif ici est d’obtenir une zone de croissance contenant autant de formes L pures que possible, plutôt que des colonies uniques. Pour cette raison, il n’est pas nécessaire de changer les boucles entre les stries. Il peut être très difficile d’obtenir des colonies uniques de formes L et l’utilisation d’un trop petit nombre de bactéries pourrait entraîner l’absence de croissance de la forme L. D’autre part, si trop de bactéries à paroi ont été utilisées pour l’induction, toutes les bactéries ne subiront pas le changement et certaines peuvent encore conserver la paroi. La meilleure zone de croissance en forme de L peut généralement être détectée au milieu de la strie. - Placez la plaque dans un sac en plastique ou un autre récipient pour éviter qu’elle ne sèche et incubez à 30 °C (les formes L de certaines espèces bactériennes peuvent mieux se développer dans des conditions anaérobies, de sorte qu’une chambre anaérobie peut être testée pour une croissance plus efficace de la forme L). Examinez la plaque à la recherche de signes de croissance tous les jours. Cela apparaît généralement dans les 3 à 7 jours. La quantité de croissance dépend de la souche. La figure 4B illustre un exemple de croissance en forme de L.
- Confirmez la présence de formes en L à l’aide d’un microscope. Placez une gouttelette de 2 μL de milieu LM sur une lame microscopique et, à l’aide d’une pointe de pipette, prélevez une petite quantité de cellules au milieu de la traînée et remettez-les en suspension dans la gouttelette.
- Recouvrez d’une lamelle de 22 x 22 mm et examinez la présence de formes en L à l’aide d’un microscope avec un objectif à contraste de phase 100x. La figure 4C en donne un exemple représentatif.
Résultats
Les milieux en forme de L contenant du saccharose peuvent subir divers degrés de caramélisation après l’autoclavage, qui sont associés à un changement de couleur. La figure 2 montre les résultats représentatifs des milieux d’autoclave contenant du saccharose. La figure 2A montre un exemple typique de milieu LM, qui a pris une couleur ambrée après l’autoclavage. La figure 2B montre un exemple typique d’une solution de saccharose 1,16 M (2 fois la concentration requise pour la fabrication du milieu LM) après autoclavage. Parfois, le milieu LM ou le saccharose peuvent subir une caramélisation importante pendant l’autoclavage, et il n’est pas recommandé d’utiliser le milieu si cela se produit. La figure 2C montre un exemple de milieu saccharose trop caramélisé.
Les bactéries en forme de L peuvent être très hétérogènes et la figure 3 montre des exemples de structures de type L observables dans les échantillons d’urine des patients. Pour confirmer l’origine bactérienne des structures de type L trouvées dans les échantillons d’urine, il est recommandé d’utiliser des sondes fluorescentes ciblant les séquences bactériennes10,23.
Les niveaux de croissance des formes L induits dans des conditions de laboratoire sont spécifiques à la souche. La figure 4B montre un exemple de croissance de la forme L de Bacillus subtilis et la figure 4C montre l’aspect microscopique des formes L induit dans la figure 4B.

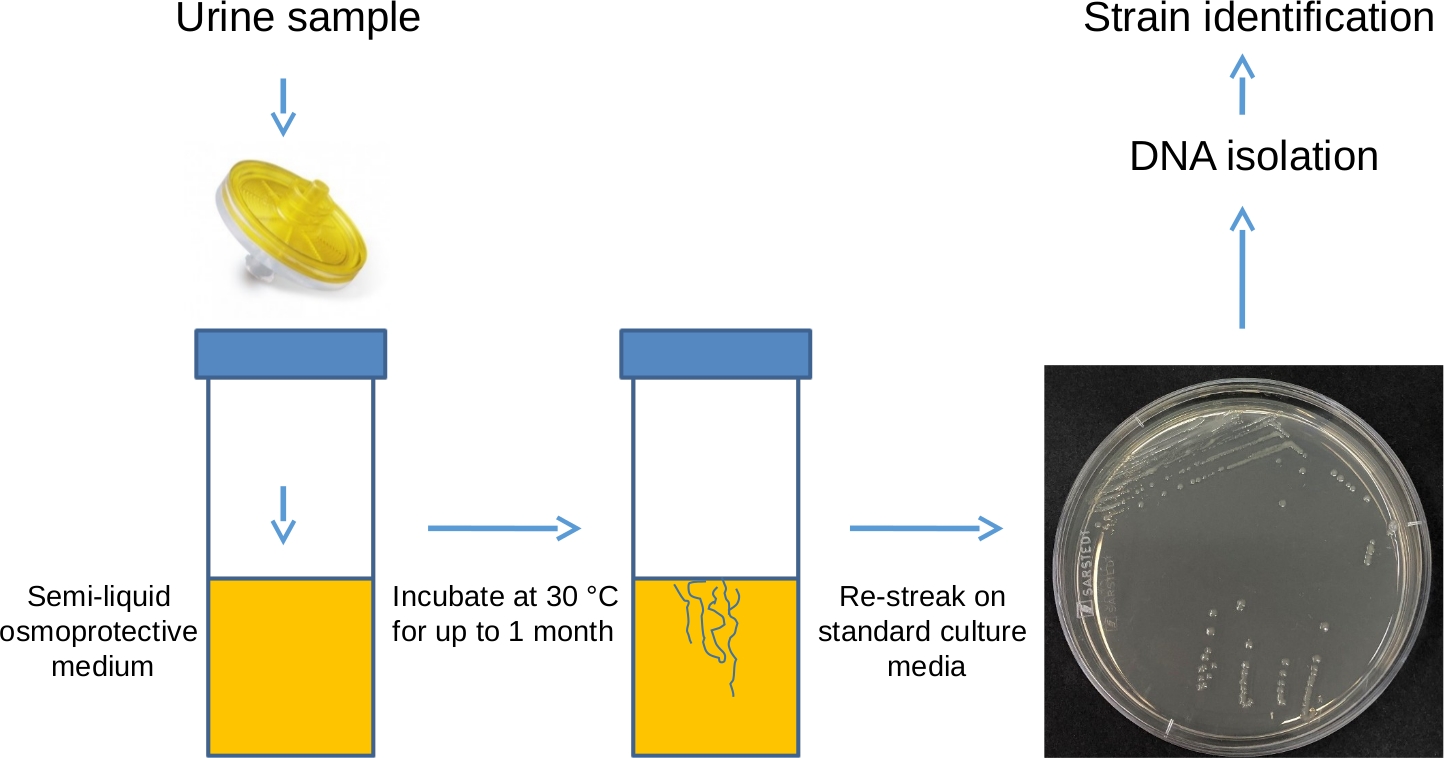
Figure 1 : Isolation des formes en L à l’aide de la méthode de filtration – représentation schématique. L’échantillon d’urine est passé à travers un filtre de 0,45 μm dans un récipient universel en polystyrène, contenant un milieu LM osmoprotecteur complété par 0,2 % d’agar. L’échantillon est ensuite incubé dans une position stationnaire à 30 °C pendant un mois et vérifié visuellement quotidiennement pour détecter tout signe de croissance. Cette période d’incubation permet à toutes les formes L présentes dans l’échantillon de régénérer leurs parois cellulaires. Une fois la croissance détectée, les bactéries peuvent être re-stries sur une plaque contenant un milieu régulier, solide et non osmoprotecteur (comme une gélose nutritive ou une perfusion cerveau-cœur) pour isoler des colonies uniques, qui peuvent ensuite être soumises à l’extraction et au séquençage de l’ADN pour identifier les espèces bactériennes isolées. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : support en forme de L. (A) Milieu LM après autoclavage. (B) 1,16 M de saccharose (2 x concentration) après autoclavage. (C) Milieu LM largement caramélisé. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Exemples de structures de type L observables dans l’urine de patients atteints d’infections urinaires récurrentes par microscopie à contraste de phase. (A,B) Structures typiques de la division des formes L bactériennes en suspension dans l’urine. (C, D) Structures typiques des formes L bactériennes en division associées aux cellules eucaryotes. (E, F) Structure en forme de croissant caractéristique des formes en L à Gram négatif (flèche rouge). (F,G) Structures typiques des étapes intermédiaires de transition entre les cellules paroises et les formes en L (flèche rouge en G). (H) Vésicules intracellulaires typiques des grandes formes en L. Barre d’échelle = 5 μm. Cette figure a été modifiée au lieu de10. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Technique de la plaque striée pour l’induction de formes en L sur des supports solides. (A) Représentation schématique de bactéries striées à l’aide d’un quart de plaque pour l’induction de formes en L. La flèche indique la direction dans laquelle strier les bactéries. (B) Un exemple de croissance de la forme L de Bacillus subtilis après stries, comme le montre (A), après 3 jours d’incubation à 30 °C. (C) Formes L induites dans (B) visualisées par microscopie à contraste de phase. Barre d’échelle = 5 μm. Les panneaux B et C ont été modifiés de16 Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
Discussion
Les protocoles décrits dans ce manuscrit ont été utilisés pour isoler et manipuler les formes L de diverses espèces bactériennes dans l’urine humaine, notamment E. coli, Streptococcus, Staphylococcus, Klebsiella, Pseudomonas, Enterococcus et Enterobacter spp, qui sont tous généralement associés aux infections urinaires10,24. Cependant, les méthodes de manipulation des formes L, établies dans un laboratoire, peuvent ne pas fonctionner immédiatement dans un autre et ce qui fonctionne pour une souche bactérienne peut ne pas fonctionner pour une autre. Par conséquent, les protocoles décrits ici sont susceptibles de nécessiter plusieurs tentatives et une optimisation supplémentaire. En particulier, il peut être nécessaire de tester les besoins en nutriments, la disponibilité en oxygène, la concentration en osmoprotecteur et la fluidité du milieu. Les conditions optimales pour la transition et la croissance de la forme L pourraient être différentes des conditions optimales pour la croissance des formes à parois de la même espèce. De plus, plusieurs problèmes techniques pourraient être rencontrés lors de la tentative des protocoles.
Un problème courant associé à la préparation de milieux en forme de L est la caramélisation du saccharose après autoclavage. Si le milieu devient brun foncé, il doit être jeté et un nouveau lot doit être préparé. Il peut être nécessaire de préparer le saccharose dans de l’eau dans une bouteille et les autres ingrédients dans une autre, tous deux à une concentration de 2x. 2x solutions concentrées peuvent ensuite être combinées dans un rapport de 1:1 après l’autoclavage, pour obtenir les concentrations finales souhaitées. Le saccharose autoclavé séparément peut devenir légèrement jaune (figure 2B), ce qui est acceptable, mais s’il devient brun foncé (figure 2C), il ne doit pas être utilisé pour des expériences. Il peut être nécessaire d’ajuster la durée et la température du cycle d’autoclave pour atténuer la caramélisation du saccharose. Il est recommandé de tester la stérilité des milieux préparés à l’aide d’un cycle d’autoclave ajusté, en incubant une aliquote de chacun des milieux pendant au moins 3 jours à 37 °C. Enfin, il peut être nécessaire de tester un agent osmoprotecteur alternatif, comme le sel, si le problème de caramélisation persiste18.
Plusieurs problèmes peuvent également être rencontrés lors de l’isolement des formes L à partir d’échantillons de patients. Comme mentionné dans le protocole, les formes L présentes dans les échantillons peuvent potentiellement se détériorer ; Par conséquent, il est important de transporter les échantillons au laboratoire le plus rapidement possible après le don. Pour la même raison, il est fortement déconseillé de stocker les échantillons à basse température ou de modifier leur composition (par exemple par l’ajout de PBS).
La méthode de filtration elle-même a ses limites. Certaines formes en L peuvent être déchirées en raison des forces de cisaillement générées par l’écoulement du média à travers le filtre. Dans l’étude de Mickiewicz et al., seulement 41 % des formes L dans les échantillons témoins, qui contenaient des formes L d’E. coli induites en laboratoire, sont passées à travers le filtre10. Cela démontre que la méthode de filtration peut conduire à une sous-estimation du nombre d’échantillons positifs et n’est pas recommandée pour les études quantitatives.
En de très rares occasions, après la filtration, une croissance significative peut être observée le lendemain dans les aliquotes de milieux semi-liquides utilisés pour l’isolement de la forme L. Cela pourrait indiquer que le filtre s’est brisé pendant le protocole, permettant le passage des bactéries à parois, ou que l’échantillon a été contaminé. Il est recommandé de jeter ces échantillons ou au moins de les traiter avec prudence. L’examen visuel quotidien des échantillons est essentiel pour éviter les faux positifs. Avant de traiter des échantillons d’urine par filtration, il est recommandé de faire passer plusieurs échantillons de formes en L induites en laboratoire, de bactéries à paroi et de milieu stérile à travers le filtre de votre choix, afin de contrôler l’efficacité de la séparation de la forme L, le passage potentiel des bactéries à paroi dû à la rupture du filtre et de s’assurer que la technique stérile utilisée fonctionne bien.
Dans de rares cas, les formes L stables peuvent être isolées par filtration, sans qu’aucune forme à paroi ne soit détectable par microscopie. Pour maintenir des bactéries de forme L viables, il est recommandé de transférer plusieurs « boucles pleines » de l’échantillon dans un tube contenant un milieu LM semi-liquide frais ou de retracer sur un milieu osmoprotecteur solide tous les 3 à 7 jours, en fonction de l’efficacité de la croissance. Si aucune forme paroi n’apparaît après plusieurs passages, on pourrait tenter de congeler des échantillons dans du glycérol à 40 % à –80 °C pour préserver l’isolat ; cependant, certaines formes en L peuvent ne pas bien tolérer une telle procédure.
Malgré ses limites, la méthode de filtration est la seule utilisée à ce jour pour séparer les formes en L des formes à paroi dans les échantillons de patients. Il permet d’identifier des souches bactériennes capables de commuter en forme L in vivo. Il est possible de mettre au point et d’adapter la méthode de filtration pour isoler les formes L d’autres types d’échantillons humains ou environnementaux (par exemple du sang ou des plantes).
Compte tenu de toutes les considérations ci-dessus, nous recommandons les protocoles décrits dans ce manuscrit comme un bon point de départ pour développer des protocoles en forme de L sur mesure dans des laboratoires individuels, plutôt que comme des instructions rigides. Travailler avec des formes en L nécessite beaucoup de soin, de dévouement et de patience, mais avec de la pratique, cela peut être extrêmement gratifiant. Nous espérons que les lignes directrices décrites dans ce manuscrit deviendront une référence pour le développement de protocoles de forme L et encourageront davantage de groupes de recherche fondamentale et clinique à étudier ces formes bactériennes fascinantes.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Ce travail a été financé par un Conseil européen de la recherche (numéro de subvention 670980) à Jeff Errington (directeur du Centre de biologie cellulaire bactérienne, Université de Newcastle).
matériels
| Name | Company | Catalog Number | Comments |
| 0.45 µL cut off filters | Sarstedt | 83.1826 | |
| 20 mL syringes | Fisher | 17955460 | |
| 30 mL polystyrene universal tubes | Starlab | E1412-3010 | |
| 92 x 16 mm Petri Dishes | Starstedt | 82.1473 | |
| Agar | Oxoid | LP0011 | |
| Ampicillin | Sigma-Aldrich | A9518 | |
| Brain Heart Infusion | Oxoid | CM1135 | |
| Cover slips (22 x 22 mm, 22 x 50 mm) | VWR | 631-0137/-0125 | |
| D-cycloserine | Sigma-Aldrich | C6880 | |
| Glass microscope slides | VWR | 631-1550P | |
| Lysostaphin | Sigma-Aldrich | L7386 | |
| Lysozyme | Sigma-Aldrich | L4919 | |
| MgSO4 | VWR | 25165.26 | |
| Moenomycin | Sigma-Aldrich | 32404 | |
| Penicillin G | Sigma-Aldrich | 13752 | |
| Phosphomycin | Sigma-Aldrich | P5396 | |
| Sucrose | Sigma-Aldrich | 84100 |
Références
- Typas, A., Banzhaf, M., Gross, C. A., Vollmer, W. From the regulation of peptidoglycan synthesis to bacterial growth and morphology. Nature Reviews Microbiology. 10, 123-136 (2011).
- Wolf, A. J., Underhill, D. M. Peptidoglycan recognition by the innate immune system. Nature Reviews Immunology. 18, 243-254 (2018).
- Kohanski, M. A., Dwyer, D. J., Collins, J. J. How antibiotics kill bacteria: from targets to networks. Nature Reviews Microbiology. 8 (6), 423-435 (2010).
- Leaver, M., et al. Life without a wall or division machine in Bacillus subtilis. Nature. 457, 849-853 (2009).
- Mercier, R., Kawai, Y., Errington, J. General principles for the formation and proliferation of a wall-free (L-form) state in bacteria. eLife. 3, 642 (2014).
- Cross, T., et al. Spheroplast-Mediated Carbapenem Tolerance in Gram-Negative Pathogens. Antimicrobial Agents and Chemotherapy. 63 (9), 00756 (2019).
- Ramijan, K., et al. Stress-induced formation of cell wall-deficient cells in filamentous actinomycetes. Nature Communications. 9, 5164 (2018).
- Errington, J., et al. L-form bacteria, chronic diseases and the origins of life. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 371, 20150494 (2016).
- Kawai, Y., Mickiewicz, K., Errington, J. Lysozyme counteracts β-Lactam antibiotics by promoting the emergence of L-form bacteria. Cell. 172, 1038-1049 (2018).
- Mickiewicz, K. M., et al. Possible role of L-form switching in recurrent urinary tract infection. Nature Communications. 10, 4379 (2019).
- Domingue, G. J., Woody, H. B. Bacterial persistence and expression of disease. Clinical Microbiology Reviews. 10, 320-344 (1997).
- Clasener, H. Pathogenicity of the L-phase of bacteria. Annual Review of Microbiology. 26, 55-84 (1972).
- Onwuamaegbu, M. E., Belcher, R. A., Soare, C. Cell wall-deficient bacteria as a cause of infections: a review of the clinical significance. Journal of International Medical Research. 33, 1-20 (2005).
- Allan, E. J., Hoischen, C., Gumpert, J. Bacterial L-forms. Advances in Applied Microbiology. 68, 1-39 (2009).
- Domingue, G. J. Demystifying pleomorphic forms in persistence and expression of disease: Are they bacteria, and is peptidoglycan the solution. Discovery Medicine. 10, 234-246 (2010).
- Kawai, Y., et al. Crucial role for central carbon metabolism in the bacterial L-form switch and killing by β-lactam antibiotics. Nature Microbiology. 4, 1716-1726 (2019).
- Klieneberger, E. The natural occurrence of pleuropneumonia-like organisms in apparent symbiosis with Streptobacillus moniliformis and other bacteria. The Journal of Pathology and Bacteriology. 40, 93-105 (1935).
- Osawa, M., Erickson, H. P. L form bacteria growth in low-osmolality medium. Microbiology. 165 (8), 842-851 (2019).
- Domingue, G. J., Schlegel, J. U. The possible role of microbial L-forms in pyelonephritis. The Journal of Urology. 104, 790-798 (1970).
- Braude, A. I., Siemienski, J., Jacobs, I. Protoplast formation in human urine. Transactions of the Association of American Physicians. 74, 234-245 (1961).
- Kalmanson, G. M., Hubert, E. G., Guze, L. B. Production and therapy of Proteus mirabilis pyelonephritis in mice undergoing chronic diuresis. Antimicrobial Agents and Chemotherapy. 9, 458-462 (1969).
- Sanders, E. R. Aseptic Laboratory Techniques: Plating Methods. Journal of Visualized Experiments. 63, e3064 (2012).
- Takada, T., Matsumoto, K., Nomoto, K. Development of multi-color FISH method for analysis of seven Bifidobacterium species in human feces. Journal of Microbiological Methods. 58, 413-421 (2004).
- Drage, L. K., et al. Elevated urine IL-10 concentrations associate with Escherichia coli persistence in older patients susceptible to recurrent urinary tract infections. Immunity & Ageing. 16, 1-11 (2019).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.