
Method Article
Suivi des particules sensibles à la masse pour caractériser la dynamique des macromolécules associées à la membrane
Dans cet article
Résumé
Ce protocole décrit une approche de traitement d’image et de suivi d’une seule particule basée sur iSCAT qui permet d’étudier simultanément la masse moléculaire et le comportement diffusif des macromolécules interagissant avec les membranes lipidiques. Des instructions étape par étape pour la préparation des échantillons, la conversion masse-contraste, l’acquisition de films et le post-traitement sont fournies ainsi que des instructions pour éviter les pièges potentiels.
Résumé
Les interactions de courte durée ou transitoires des macromolécules au niveau et avec les membranes lipidiques, une interface où se produisent une multitude de réactions biologiques essentielles, sont intrinsèquement difficiles à évaluer avec des méthodes biophysiques standard. L’introduction du suivi des particules sensibles à la masse (MSPT) constitue une étape importante vers une caractérisation quantitative approfondie de ces processus. Techniquement, cela a été rendu possible grâce à l’avènement de la photométrie de masse (MP) basée sur la microscopie à diffusion interférométrique (iSCAT). Lorsque la stratégie d’élimination de fond est optimisée pour révéler le mouvement bidimensionnel des particules associées à la membrane, cette technique permet l’analyse en temps réel de la diffusion et de la masse moléculaire des macromolécules non marquées sur les membranes biologiques. Ici, un protocole détaillé pour effectuer et analyser le suivi des particules sensibles à la masse des systèmes associés à la membrane est décrit. Les mesures effectuées sur un photomètre de masse commercial atteignent une résolution temporelle de l’ordre de la milliseconde et, selon le système MP, une limite de détection de masse jusqu’à 50 kDa. Pour mettre en valeur le potentiel de la MSPT pour l’analyse approfondie de la dynamique des macromolécules catalysées par membrane en général, les résultats obtenus pour des systèmes protéiques exemplaires tels que l’interacteur membranaire natif annexine V sont présentés.
Introduction
Autrefois simplement perçues comme une barrière contre le large éventail de conditions physiques ambiantes, les membranes biologiques sont aujourd’hui considérées comme des entités fonctionnelles etdes plates-formes catalytiques 1,2. En raison de leur capacité à localiser, amplifier et diriger les signaux en réponse aux réactions de macromolécules associées à la membrane, les interfaces lipidiques constituent un élément crucial pour une grande variété de processus cellulaires tels que le trafic membranaire et les cascades de signalisation 3,4,5. Servant de site de nucléation pour l’assemblage de complexes stables, la fixation membranaire repose souvent sur un équilibre dynamique entre les formes de macromolécules associées à la membrane et cytosoliques et est donc de nature transitoire 6,7.
Malgré leur grande importance en biologie, il a jusqu’à présent été difficile de développer des méthodes capables de donner accès aux hétérogénéités compositionnelles, spatiales et temporelles des réactions de macromolécules associées à la membrane en temps réel 7,8. Pour résoudre les processus moléculaires sous-jacents, deux aspects expérimentaux sont décisifs : une résolution temporelle suffisante et une sensibilité à une seule particule. Par conséquent, les techniques moyennes d’ensemble telles que la récupération de fluorescence après photoblanchiment (FRAP) mais aussi la spectroscopie de corrélation de fluorescence (FCS), beaucoup plus sensible, ont des limites, car elles dissocient largement les informations spatiales des informationstemporelles 9. Une étape importante vers la caractérisation de la dynamique des molécules individuelles a donc été l’avènement du suivi des particules uniques (SPT) en combinaison avec la microscopie hautement sensible. En particulier, deux approches SPT se sont avérées efficaces à cet égard. Tout d’abord, l’utilisation de fluorophores comme étiquettes et les systèmes de détection de fluorescence correspondants ont ouvert la voie à une précision nanométrique et à une résolution temporelle milliseconde 10,11,12. Deuxièmement, la détection basée sur la diffusion à l’aide de nanoparticules d’or a amélioré à la fois la précision de localisation et la résolution temporelle dans la plage sub-nanométrique et microseconde, respectivement 13,14,15,16. Malgré les nombreux avantages des deux approches et leurs contributions significatives concernant la compréhension mécaniste des systèmes membranairesassociés 17,18, les deux techniques ont jusqu’à présent été limitées : elles nécessitent le marquage des molécules d’intérêt, ce qui perturbe potentiellement leur comportement natif et est insensible à la composition moléculaire des particules associées à la membrane19,20.
Ces deux limites ont récemment été surmontées par l’introduction d’une nouvelle approche basée sur la diffusion interférométrique (iSCAT) appelée photométrie de masse (MP)21,22,23. Cette technique permet de déterminer les distributions de masse en solution des biomolécules en fonction de leur contraste iSCAT lors de l’atterrissage sur une interface en verre. Cependant, pour la détection et la caractérisation des molécules mobiles diffusant sur les membranes lipidiques, une approche d’analyse d’image plus sophistiquée a dû être développée. Cela a entre-temps été mis en œuvre avec succès et permet de détecter, de suivre et de déterminer la masse moléculaire de biomolécules uniques non marquées diffusant sur une interface lipidique24,25. Appelée photométrie de masse dynamique ou suivi des particules sensibles à la masse (MSPT), cette technique permet désormais d’évaluer les interactions complexes des macromolécules en enregistrant directement les changements dans la masse moléculaire des entités suivies et ouvre ainsi de nouvelles possibilités pour l’analyse mécaniste de la dynamique moléculaire associée à la membrane.
Ici, un protocole détaillé pour la préparation des échantillons, l’imagerie et le pipeline d’analyse des données requis pour MSPT est présenté. En particulier, les exigences en matière d’échantillons et les problèmes potentiels pouvant survenir lors de la mesure et de l’analyse sont discutés. En outre, le potentiel inégalé d’analyse des systèmes de macromolécules interagissant avec la membrane est mis en évidence par divers résultats représentatifs.
Protocole
1. Préparation de l’échantillon
- Génération de vésicules multilamellaires (VLM)
- Calculer la quantité de lipides dissous dans le chloroforme en fonction du mélange lipidique souhaité et du volume de suspension requis.
REMARQUE: Une concentration finale de vésicule de 4 mg / mL lipides est recommandée pour le tampon de remise en suspension (réaction). - Pipeter le volume calculé de lipides dans un flacon en verre de 1,5 mL à l’aide de pipettes à déplacement positif équipées d’embouts en verre.
- Évaporer le solvant lipidique sous un léger jet d’azote et faire tourner constamment le flacon pour assurer une distribution égale des lipides sur les parois vitrées.
- Assurer l’évaporation complète du solvant en plaçant le flacon sous un flux constant d’azote pendant 15 min.
- Éliminer les traces résiduelles de chloroforme en séchant sous vide dans un dessiccateur sous vide pendant une heure supplémentaire.
- Réhydrater le mélange lipidique dans le tampon de remise en suspension (réaction) souhaité et bien vortexer la suspension jusqu’à ce que le film lipidique ait été dissous des parois du flacon.
REMARQUE: Le tampon de réaction doit assurer l’activité et la stabilité des protéines. Le tampon réactionnel utilisé dans cette étude contient 50 mM de Tris-HCl (pH = 7,5), 150 mM de KCl et 5 mM de MgCl2. Notez que tout tampon utilisé pour diluer les lipides ou les protéines doit être filtré pour éliminer les impuretés particulaires interférentes (voir l’étape 5).
- Calculer la quantité de lipides dissous dans le chloroforme en fonction du mélange lipidique souhaité et du volume de suspension requis.
- Génération de petites vésicules unilamellaires (VUS)
- Pour les cycles consécutifs de congélation-décongélation de la remise en suspension lipidique (étape 1.1.6), faire bouillir 500 mL d’eau dans un bécher sur une plaque chauffante (entre 70 °C et 99 °C) et préparer un récipient avec de l’azote liquide.
- Gel par choc la remise en suspension lipidique dans l’azote liquide. Transférer le flacon dans le bécher avec de l’eau chaude jusqu’à ce que la solution soit complètement décongelée. Répétez ce cycle de congélation-décongélation 8 à 10 fois ou jusqu’à ce que le mélange précédemment trouble apparaisse clair.
ATTENTION : Utilisez des vêtements et de l’équipement de sécurité appropriés tels que des lunettes, des gants et une pince à épiler pour éviter tout contact direct avec l’azote liquide, le flacon lipidique congelé ou l’eau bouillante. - Pour la génération d’une distribution de vésicule monodispersée, assemblez une extrudeuse lipidique et testez son intégrité avec un tampon de réaction pour vous assurer qu’elle ne fuit pas.
REMARQUE: Si une fuite est observée, réassemblez soigneusement l’extrudeuse lipidique jusqu’à ce qu’aucun déversement de tampon ne soit évident. - Extruder la suspension lipidique pendant 37 passages à travers une membrane nucléopore avec une taille de pore de 50 nm26. Le nombre de passages doit être inégal pour s’assurer que le mélange final de SUV a traversé la membrane nucléopore et est donc exempt d’agrégats lipidiques ou de vésicules multilamellaires. Les vésicules extrudées seront utilisées ultérieurement pour former des bicouches lipidiques supportées (voir étapes 6 et 7).
REMARQUE: Les SUV peuvent également être formés par sonication du mélange lipidique réhydraté. Cependant, la préparation par extrusion fournit une distribution plus monodispersée des VUS, ce qui facilite la rupture des vésicules lors de la formation de bicouche lipidique soutenue. Les vésicules extrudées peuvent être conservées au réfrigérateur pendant un maximum de 3 jours.
2. Nettoyage des lames de microscope
- Distribuer un nombre égal de lames de microscope (n° 1.5; épaisseur de 0,17 mm) avec des dimensions de 24 mm x 60 mm et 24 mm x 24 mm dans des porte-microscopes en polytétrafluoréthylène (PTFE).
- Transférer les porte-PTFE dans des béchers contenant de l’eau ultrapure et les soniquer pendant 15 min à température ambiante.
REMARQUE: Selon le bécher, le volume d’eau doit être ajusté pour couvrir complètement le support en PTFE. - Utilisez une pince à épiler pour retirer les supports du bécher et remplacez l’eau par de l’isopropanol ultrapur. Insérez le support dans le bécher contenant de l’isopropanol et sonicez à nouveau pendant 15 min.
REMARQUE: Selon le bécher, le volume de l’isopropanol doit être ajusté pour couvrir complètement le support en PTFE. - Remplacer l’isopropanol par de l’eau ultrapure et soniquer le bécher contenant les supports pendant 15 min.
- Retirez les supports en PTFE des béchers et séchez les lames du microscope dans le support sous un flux constant d’azote gazeux ou d’air comprimé.
REMARQUE: Assurez-vous d’un nettoyage approprié des glissières de couverture en utilisant des gants, des béchers propres et un film de paraffine pour couvrir chaque bécher. Sinon, la poussière résiduelle peut provoquer des fluctuations de fond importantes lors des mesures MSPT.
3. Hydrophilisation des lames de microscope
REMARQUE: Pour obtenir une bicouche lipidique homogène et fluide, l’hydrophilisation des lames est essentielle et doit être effectuée juste avant l’assemblage de la chambre d’écoulement.
- Placez les supports en PTFE contenant seulement des lames de microscope de 24 mm x 60 mm dans un nettoyeur plasma avec de l’oxygène comme gaz de procédé et nettoyez les lames de microscope avec du plasma (paramètres utilisés dans ce travail: puissance de 30%, pression d’oxygène de 0,3 mbar pendant 30 s; voir tableau des matériaux pour plus de détails sur le nettoyeur plasma utilisé).
REMARQUE: Pour obtenir des membranes fluides, les paramètres de nettoyage plasma tels que la puissance, la pression d’oxygène et le temps de nettoyage doivent être ajustés pour chaque instrument. À cette fin, l’utilisation de lipides marqués par fluorescence est recommandée pour assurer la fluidité de la membrane, qui peut être quantifiée avec des expériences de récupération de fluorescence après photoblanchiment (FRAP)27. Si les paramètres ne sont pas optimisés pour la configuration respective, la diffusion de la membrane peut être altérée en raison de la fluidité réduite de la membrane.
4. Assemblage des chambres d’écoulement
- Avant l’assemblage de la chambre d’écoulement, gardez les composants suivants prêts : lames de microscope nettoyées (24 mm x 24 mm), lames de microscope hydrophilisées (24 mm x 60 mm), feuille d’aluminium, carton plat, scalpel et ruban adhésif double face.
- Enveloppez le carton plat avec du papier d’aluminium.
- Étalez les lames de microscope nettoyées de 24 mm x 24 mm sur la feuille d’aluminium avec une distance suffisante entre elles.
- Fixez des bandes de ruban adhésif double face aux bords supérieur et inférieur des glissières.
- Retirez chaque lame de microscope avec le scalpel, de sorte qu’elle puisse être retirée de la feuille d’aluminium. Par conséquent, chaque diapositive doit être munie de bandes de ruban adhésif double face attachées aux bords supérieur et inférieur de la diapositive (voir la figure 1).
- Fixez la lame de 24 mm x 24 mm avec les deux bandes de ruban adhésif double face à la lame hydrophilisée de 24 mm x 60 mm pour former un chemin d’écoulement entre les plus petites et les plus grandes lames de microscope.
REMARQUE: Pour assurer la propreté des chambres d’écoulement, portez constamment des gants et assurez-vous que l’établi est exempt de poussière.
5. Filtration des tampons de réaction
- Filtre stérile tous les tampons de réaction à travers des membranes d’acétate de cellulose de 0,45 μm pour assurer un signal de fond minimal lors des mesures MSPT.
REMARQUE: Si la présence de nucléotides, tels que l’ATP, est essentielle pour une expérience réussie, soyez conscient d’une augmentation potentielle du signal de fond. Il est recommandé de n’utiliser que des quantités minimales qui assurent encore l’activité des protéines.
6. Formation de bicouche lipidique (SLB) prise en charge
REMARQUE: Il est recommandé d’effectuer la formation de bicouches lipidiques supportées sur le photomètre de masse pour assurer visuellement une propagation réussie des vésicules et l’élimination complète des vésicules non fusionnées.
- Diluer les VUS fraîchement extrudés (voir l’étape 1 pour plus de détails) jusqu’à une concentration finale de 0,4 mg/mL dans le tampon de réaction requis. Éventuellement, pour favoriser la rupture des vésicules, ajouter 2 mM de CaCl2 à la suspension de la vésicule.
REMARQUE: Les cations divalents peuvent provoquer l’agrégation de certains lipides tels que PiP2. Pour les mélanges contenant de tels lipides, s’abstenir d’utiliser CaCl2 pour favoriser la rupture des vésicules ou d’autres cations divalents dans le tampon de remise en suspension. Si nécessaire pour l’expérience, des cations divalents peuvent être ajoutés après la formation réussie de la bicouche lipidique supportée. - Rincer 50 μL de la suspension de vésicule dans la chambre d’écoulement (étape 4) et incuber la chambre pendant 2 min.
REMARQUE: Les tampons, les vésicules ou les solutions protéiques peuvent être rincés à travers la chambre d’écoulement avec un petit morceau de tissu de trempage. Cependant, il est également possible d’utiliser un système de pompe mécanique. - Retirer les vésicules non fusionnées par lavage répété (au moins trois fois) de la chambre d’écoulement avec 200 μL du tampon de réaction à chaque fois.
REMARQUE: Les vésicules doivent être soigneusement lavées de la chambre d’écoulement pour assurer un signal de fond stable pendant les mesures MSPT.
7. Génération de la courbe d’étalonnage
REMARQUE: Pour convertir le contraste des particules détectées en masse moléculaire, leur signal doit être étalonné à l’aide de protéines de tailles connues. Il est recommandé d’ajuster le régime standard de taille des protéines pour couvrir la gamme de masses moléculaires attendues pour le système d’intérêt.
- Biotinylation de protéines standard avec un résidu de cystéine
- Calculer la quantité appropriée de maléimide-biotine pour la protéine standard conformément aux instructions du fabricant.
- Incuber la protéine standard avec le volume déterminé de maléimide-biotine pendant 1 h à température ambiante.
- Pour éliminer la maléimide-biotine non conjuguée du complexe conjugué biotine-protéine, effectuez une chromatographie d’exclusion de taille sur une colonne adaptée à la protéine d’intérêt.
- Déterminez la concentration en protéines à l’aide d’un test de Bradford.
REMARQUE: Pour stocker la protéine standard pour d’autres mesures, congeler la protéine dans des aliquotes à usage unique dans de l’azote liquide et les stocker à -80 ° C.
- Mesure des protéines étalons pour la courbe d’étalonnage
- Dans une chambre d’écoulement, préparer une bicouche lipidique supportée avec des VUS extrudés de 0,4 mg/mL (voir les étapes 1 et 6 pour plus de détails) contenant 0,01 mol% (v/v) biotinyl Cap PE (1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-cap biotinyl).
- Ajouter 50 μL de streptavidine divalente de 2,5 nM à la bicouche dans la chambre d’écoulement et incuber pendant 10 min.
REMARQUE: La streptavidine divalente a été exprimée et purifiée comme indiqué dans Howarth et al.28. La streptavidine tétravalente peut également être utilisée. Cependant, l’utilisation de streptavidine divalente peut réduire les stœchiométries de réaction possibles entre les lipides biotinylés et les protéines standard qui sont conjuguées à une fraction de biotine afin de faciliter l’attribution des espèces. - Éliminer la streptavidine divalente non liée avec 100 μL de tampon réactionnel.
- Ajouter 50 μL de protéine étalon conjuguée à la biotine de 100 nM à la bicouche dans la chambre d’écoulement et incuber pendant 2 min.
REMARQUE: Selon l’efficacité de la biotinylation et si la streptavidine di- ou tétravalente est utilisée, les concentrations optimales de protéine standard conjuguée à la biotine et de streptavidine peuvent varier. - Effectuez la mesure MSPT selon les détails décrits à l’étape 8.
ATTENTION : Les conditions d’imagerie doivent être identiques pour les étalons d’échantillonnage et d’étalonnage.
8. Imagerie
- Formation de SLB et préparation des échantillons
- Comme décrit plus en détail à l’étape 6, introduisez les VUS du mélange lipidique souhaité (25 μL) dans la chambre d’écoulement de l’échantillon et formez une bicouche lipidique supportée. Lavez soigneusement la chambre (trois fois) avec 100 μL de tampon de réaction pour éliminer toutes les vésicules non fusionnées.
- Ajouter 50 μL de la protéine d’intérêt à la chambre d’échantillonnage.
REMARQUE: Comme la MSPT est une méthode à particule unique, la concentration en protéines doit être maintenue dans la plage pM à nM pour permettre la détection et le suivi des particules non perturbées.
- Acquisition vidéo
- Définissez les conditions d’imagerie souhaitées telles que la taille du champ de vision (FOV), la fréquence d’images, le temps d’exposition et le temps d’acquisition dans le logiciel d’acquisition.
REMARQUE: Les paramètres suivants se sont avérés efficaces pour MSPT sur un photomètre de masse commercial (voir tableau des matériaux): FOV de 128 pixels x 35 pixels, une fréquence d’images de 1 kHz résultant en environ 200 images par seconde après une moyenne d’images 5 fois supérieure et un temps d’exposition de 0,95 ms. - Réglez la mise au point automatiquement ou manuellement. Si nécessaire, déplacez le champ de vision dans une position avec une membrane homogène à l’aide de la commande latérale.
- Créez un dossier de projet et commencez à enregistrer le film. Une fois l’enregistrement terminé, spécifiez un nom de fichier dans la boîte de dialogue demandée par le logiciel d’acquisition. Le film est ensuite automatiquement enregistré dans le dossier du projet en tant que fichier MP pour une analyse ultérieure.
REMARQUE : Enregistrez au moins trois répétitions dans différentes chambres d’écoulement pour assurer l’intégrité des membranes individuelles et la reproductibilité des résultats. La durée du film peut être définie à l’avance et dépend du type d’expérience. Dans la plupart des cas, un temps d’acquisition compris entre 5 min et 7 min est recommandé.
ATTENTION : Par défaut, les enregistrements vidéo sur le logiciel commercial d’acquisition de photomètres de masse sont compressés avant d’être enregistrés afin de réduire l’espace de stockage. Toutefois, la compression de fichiers doit être désactivée pour permettre l’analyse personnalisée des données comme décrit dans ce protocole. Vous trouverez des détails sur la façon de désactiver la compression de fichiers dans le manuel d’utilisation du fabricant.
- Définissez les conditions d’imagerie souhaitées telles que la taille du champ de vision (FOV), la fréquence d’images, le temps d’exposition et le temps d’acquisition dans le logiciel d’acquisition.
9. Analyse des données
REMARQUE : Le pipeline d’analyse de données est accompagné de deux blocs-notes Jupyter interactifs (MSPT analysis.ipynb, Movie visualization.ipynb). Les blocs-notes Jupyter et les modules Python personnalisés associés nécessaires pour effectuer l’analyse MSPT décrite ci-dessous sont disponibles dans un référentiel public : https://github.com/MSPT-toolkit/MSPT-toolkit. Pour des instructions détaillées sur l’analyse ci-dessous, les lecteurs sont référés à MSPT analysis.ipynb accessible en utilisant le lien ci-dessus.
- Traitement vidéo
- Supprimez la diffusion statique dominante de la lumière à l’aide de l’algorithme d’estimation d’arrière-plan par pixel à l’aide de la fonction image_processing.mp_reader .
- Pour appliquer la suppression de l’arrière-plan, choisissez l’option continuous_median pour le mode paramètre et définissez une longueur appropriée pour la fenêtre médiane coulissante (window_length) dans la section B.1 du bloc-notes. Si vous le souhaitez, enregistrez les films après la suppression de l’arrière-plan à utiliser pour la détection des particules et la liaison de trajectoire (en définissant le paramètre save_processed_movies sur True).
REMARQUE: Ajustez la taille de la fenêtre (window_length) à des valeurs comprises entre 101 et 2001 en fonction de la densité de particules sur la membrane, du coefficient de diffusion attendu, de la fréquence d’images d’acquisition et de la vitesse de traitement requise.
ATTENTION : La stratégie d’enlèvement de fond fonctionne bien si la membrane n’est pas trop dense et si la diffusion des particules est suffisamment rapide (c’est-à-dire que chaque pixel n’est la plupart du temps pas occupé par une particule). Sinon, le contraste des particules sera systématiquement sous-estimé car elles ne peuvent pas être correctement distinguées du signal de fond. Cela peut être compensé en augmentant la taille médiane de la fenêtre au détriment de la vitesse de calcul. Cependant, sachez que le fait de définir la taille de la fenêtre trop grande peut influencer négativement la sortie en raison de la dérive de l’échantillon. Une inspection visuelle des vidéos traitées est cruciale.
- Pour appliquer la suppression de l’arrière-plan, choisissez l’option continuous_median pour le mode paramètre et définissez une longueur appropriée pour la fenêtre médiane coulissante (window_length) dans la section B.1 du bloc-notes. Si vous le souhaitez, enregistrez les films après la suppression de l’arrière-plan à utiliser pour la détection des particules et la liaison de trajectoire (en définissant le paramètre save_processed_movies sur True).
- Détectez les particules et leur position respective tout au long du film à l’aide de la fonction particle_fitting.particle_fitter (voir la section B.2 du bloc-notes).
- Réglez la sensibilité de la détection des particules avec le paramètre de seuil (battage; voir la section B.1 du carnet), qui est utilisé pour mettre en évidence les points candidats par binarisation d’image. L’effet de la variation des paramètres de seuil sur la sensibilité de détection ponctuelle peut être examiné dans un bloc-notes séparé (Movie visualization.ipynb). Les résultats de la détection de particules sont automatiquement enregistrés dans des fichiers CSV dans un sous-répertoire du fichier vidéo.
REMARQUE: Il n’est pas recommandé de définir le paramètre de seuil arbitrairement bas (par exemple, pour les films pris avec le photomètre de masse utilisé, un paramètre de seuil inférieur à 0,0005) car les spots candidats seront dominés par un bruit fallacieux et prolongeront donc le temps de traitement.
- Réglez la sensibilité de la détection des particules avec le paramètre de seuil (battage; voir la section B.1 du carnet), qui est utilisé pour mettre en évidence les points candidats par binarisation d’image. L’effet de la variation des paramètres de seuil sur la sensibilité de détection ponctuelle peut être examiné dans un bloc-notes séparé (Movie visualization.ipynb). Les résultats de la détection de particules sont automatiquement enregistrés dans des fichiers CSV dans un sous-répertoire du fichier vidéo.
- Supprimez la diffusion statique dominante de la lumière à l’aide de l’algorithme d’estimation d’arrière-plan par pixel à l’aide de la fonction image_processing.mp_reader .
- Liez des particules dans des images consécutives dans des trajectoires à l’aide du package Python trackpy (v.0.5.0)29.
REMARQUE: La liaison de trajectoire est effectuée à la volée après la détection de spot. Par conséquent, un fichier CSV supplémentaire contenant les informations de trajectoire est stocké dans un sous-répertoire du fichier CSV de détection de particules.- Supprimez les trajectoires avec trop peu de points à l’aide du paramètre minimum_trajectory_length (voir la section B.1 du cahier) pour permettre une détermination robuste des coefficients de diffusion. Pour des explications détaillées concernant les autres paramètres des fonctions trackpy , reportez-vous à la documentation de trackpy .
- Analyse de trajectoire
- Dans la section C.1 du bloc-notes, spécifiez la fréquence d’images (frame_rate) et la taille des pixels en nm (pixel_size), qui ont été utilisées pour l’acquisition de films. Créez une liste de fichiers CSV contenant les informations de trajectoire renvoyées par trackpy (voir étape 9.2) avec la fonction trajectory_analysis.get_csv_files (section C.2 du bloc-notes).
- En outre, spécifiez un nom de fichier de sortie pour le conteneur HDF5, qui est utilisé pour stocker les résultats d’ajustement sur le disque (section C.3 du bloc-notes). Analysez toutes les trajectoires avec la fonction trajectory_analysis.fit_trajectories dans la section C.4 du bloc-notes, qui parcourt la liste des fichiers CSV. Cette fonction utilise la distribution de distance de saut (JDD)30 et l’analyse du déplacement moyen au carré (MSD)31 pour estimer le coefficient de diffusion de chaque trajectoire.
- Convertissez le contraste médian de chaque trajectoire en masse correspondante en utilisant la relation contraste-masse obtenue à partir de l’étalonnage MSPT (voir section 7). Spécifiez la pente (pente) et l’interception y (décalage) de la ligne d’étalonnage, qui relie le contraste iSCAT à la masse moléculaire (fonction trajectory_analysis.apply_calibration; voir la section C.5 du cahier). Cette fonction ajoute une colonne avec la masse médiane de la trajectoire à chaque bloc de données.
- Évaluez la densité apparente des particules sur la membrane avec la fonction trajectory_analysis.membrane_density, qui renvoie la valeur de densité médiane en termes de particules détectées et présente les trajectoires au cours de chaque image (voir la section C.6 du bloc-notes) sous forme de colonnes supplémentaires dans le bloc de données.
REMARQUE: Comme une partie des particules sera perdue pendant le processus de détection et de liaison de trajectoire, les densités réelles de particules peuvent être plus élevées. Pour des résultats fiables concernant les densités de particules ainsi que les histogrammes de masse, inspectez visuellement des instantanés de film représentatifs pour vérifier que les conditions de mesure sont plausibles pour le suivi d’une seule particule (voir l’étape 9.1.1).
10. Visualisation des données
- Illustrer la corrélation du coefficient de masse et de diffusion avec l’estimation bidimensionnelle de la densité du noyau (KDE), qui est basée sur le paquet Python fastkde (v.1.0.19; https://pypi.org/project/fastkde/).
- Pour générer le tracé, spécifiez le fichier HDF5 contenant les résultats MSPT (voir l’étape 9.3.2 et la section D.1 du bloc-notes) et sélectionnez un seul (bloc-notes section D.2) ou un bloc de données concaténé (section D.3 du bloc-notes) comme données d’entrée pour la fonction plotting.generate_2D_KDE (section D.4 du bloc-notes).
REMARQUE: Chaque jeu de données tracé doit contenir idéalement plus de 1 000 trajectoires pour un 2D-KDE fiable.
Résultats
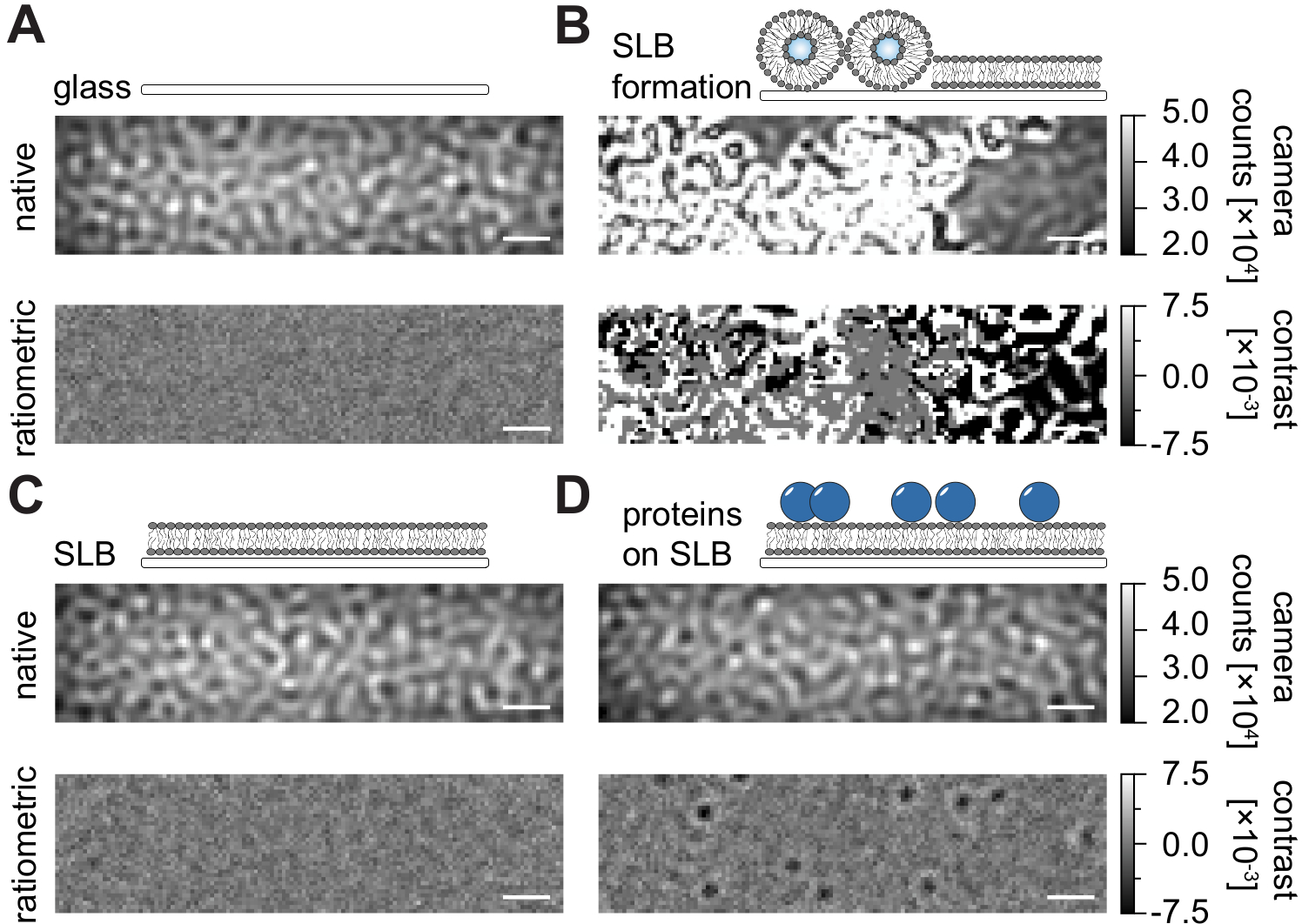
En suivant le protocole détaillé ici pour la préparation des bicouches lipidiques (SLB) supportées dans les chambres d’écoulement (Figure 1), on peut clairement reconnaître un motif ressemblant à une moucheture dans la vue native de toutes les conditions affichées (Figure 2). Cet effet est causé par la rugosité de surface du verre, qui domine généralement le signal de diffusion et conduit à des conditions visuellement indiscernables (verre, verre avec SLB ou verre avec SLB et protéines attachées). La présence de vésicules, cependant, est clairement distincte en raison de la grande section de diffusion des vésicules et permet l’observation de la rupture et de la fusion des vésicules en membranes homogènes (Figure 2B et Film supplémentaire 1). En enlevant le signal de diffusion statique de la surface du verre avec une approche ratiométrique qui met l’accent sur les éléments dynamiques dans le champ de vision24,25, on peut découvrir des protéines non marquées diffusant sur la membrane (Figure 2D) tandis qu’un SLB vide (Figure 2C) ou le verre lui-même (Figure 2A) apparaît comme une image bruyante.
L’arrière-plan inhérent aux mesures MSPT peut être estimé localement en divisant chaque valeur de pixel par la médiane de n pixels précédents et suivants du film à la même position d’image (Figure 3). En conséquence, les macromolécules apparaissent comme des fonctions isotropes d’étalement ponctuel (PSF) dont le mouvement sur la membrane peut être observé, suivi et quantifié. En fait, la disponibilité du contraste et du comportement dynamique permet la relation directe de la taille moléculaire d’une particule à son comportement diffusif respectif, le tout sans qu’il soit nécessaire de marquer la particule. Néanmoins, pour interpréter le contraste iSCAT déterminé lors des expériences MSPT, il est essentiel d’effectuer un étalonnage qui traduit l’amplitude du signal en masse moléculaire. Ceci peut être réalisé en attachant des biomolécules de masse connue à un SLB via un complexe biotine-streptavidine-biotine (Figure 4A). Comme stratégie exemplaire, on peut utiliser des variantes biotinylées de l’albumine sérique bovine (BSA), de la protéine A (prA), de la phosphatase alcaline (AP) et de la fibronectine (FN), qui se lient à la streptavidine (STP) qui est elle-même liée aux lipides contenant de la biotine (Biotinyl Cap PE) dans la membrane. Comme le montre la figure 4A, le contraste de plus en plus prononcé de ces macromolécules exemplaires reflète le poids moléculaire croissant des étalons biotinylés respectifs. En attribuant chaque pic des histogrammes de contraste (Figure 4B) à la masse correspondante de l’état oligomère de la protéine standard, une relation linéaire entre le contraste et la masse est révélée21,22 et peut ensuite être utilisée pour l’analyse de systèmes de macromolécules inconnus (Figure 4C).
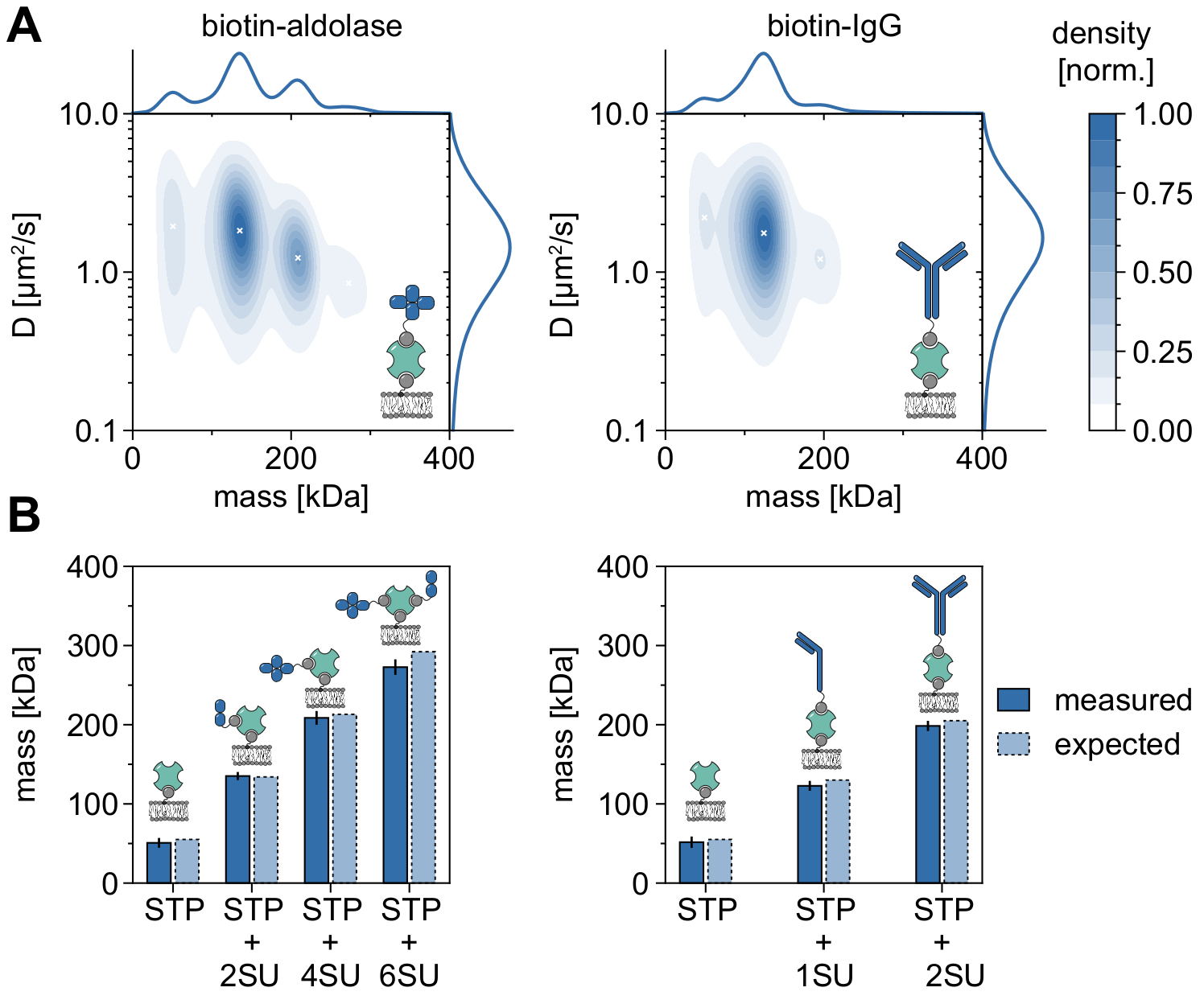
Un bon exemple démontrant l’applicabilité et les capacités de la MSPT pour analyser les poids moléculaires et donc étudier les états des oligomères et les événements d’oligomérisation est la prise en compte de l’aldolase biotinylée et des IgG biotinylées (Figure 5). L’aldolase est généralement considérée comme un homotétramère32. Cependant, la distribution de masse résolue par mspt comporte quatre pics distincts, ce qui met en évidence la présence de plusieurs populations (figure 5A). Alors que le premier pic mineur correspond à la streptavidine inoccupée et peut être attendu en raison de la configuration dans ce type d’expérience, des complexes d’aldolases avec seulement deux sous-unités (2SU) ou six sous-unités (6SU) peuvent également être détectés (Figure 5B). Fait intéressant, les complexes tétra- et hexamériques aldolase-streptavidine présentent un coefficient de diffusion réduit par rapport à l’aldolase dimérique et à la streptavidine seule, ce qui indique une traînée visqueuse accrue, par exemple via la fixation d’un deuxième lipide biotinylé à la streptavidine. De même, les IgG biotinylées présentent trois pics dans la distribution de masse, le premier pic correspondant à nouveau à la masse d’une seule streptavidine. La masse du pic le plus abondant correspond à la masse d’une chaîne légère et d’une chaîne lourde (1SU), c’est-à-dire la moitié d’un anticorps IgG. L’anticorps complet avec deux moitiés identiques (2SU) est détecté dans environ 11% des cas. La diminution du coefficient de diffusion avec l’augmentation des tailles complexes indique des interactions de la streptavidine avec plus d’un lipide biotinylé ou une traînée supplémentaire causée par les IgG attachées, ou les deux.
Outre la seule analyse des états oligomères dépendants de la membrane, la MSPT confère également l’avantage particulier de corréler le comportement diffusif d’une macromolécule d’intérêt avec son état oligomère. Des résultats représentatifs pour ce type d’analyse sont présentés pour l’annexine V (AnV) et la sous-unité B de la toxine cholérique (CTxB), qui se lient à la dioléoylphosphatidylsérine (DOPS) ou aux glycosphingolipides (GM1), respectivement, incorporés dans la membrane (figure 6A). Les deux estimations de densité du noyau (KET) présentent des distributions unimodales de masse et de diffusion, indiquant une seule espèce abondante ayant un comportement diffusif similaire. La position maximale de la masse moléculaire et du coefficient de diffusion s’est avérée être de 49,8 ± 2,2 kDa et 1,4 ± 0,1 μm2/s, respectivement, pour l’AnV ainsi que de 62,7 ± 3,1 kDa et 0,4 ± 0,1 μm2/s, respectivement, pour CTxB. Les coefficients de diffusion mesurés sont comparables aux valeurs précédemment rapportées obtenues à partir de l’AFM à grande vitesse et du FRAP33,34. La masse légèrement réduite par rapport à la masse de la macromolécule attendue (52 kDa pour un trimère AnV, 65 kDa pour un pentamère CTxB) peut indiquer la présence de complexes plus petits avec moins de sous-unités dans l’ensemble. Bien que la différence de masse entre les protéines soit faible et proche de la limite de détection spécifiée au microscope (≈50 kDa), leurs coefficients de diffusion diffèrent considérablement. Dans un mélange équimolaire, par exemple, en comparant la diffusion du mélange à la distribution de l’AnV et du CTxB seul, on peut conclure que l’AnV est plus abondant sur la membrane que le CTxB (Figure 6B). Cependant, si la concentration de CTxB est doublée par rapport à la concentration d’AnV, l’équilibre est déplacé vers CTxB comme protéine prédominante sur la membrane. Comme illustré pour les mélanges d’AnV et de CTxB, le MSPT permet non seulement de discriminer les macromolécules associées à la membrane en fonction de leur poids moléculaire, mais permet également de discriminer différentes populations de macromolécules en fonction de leur comportement diffusif.
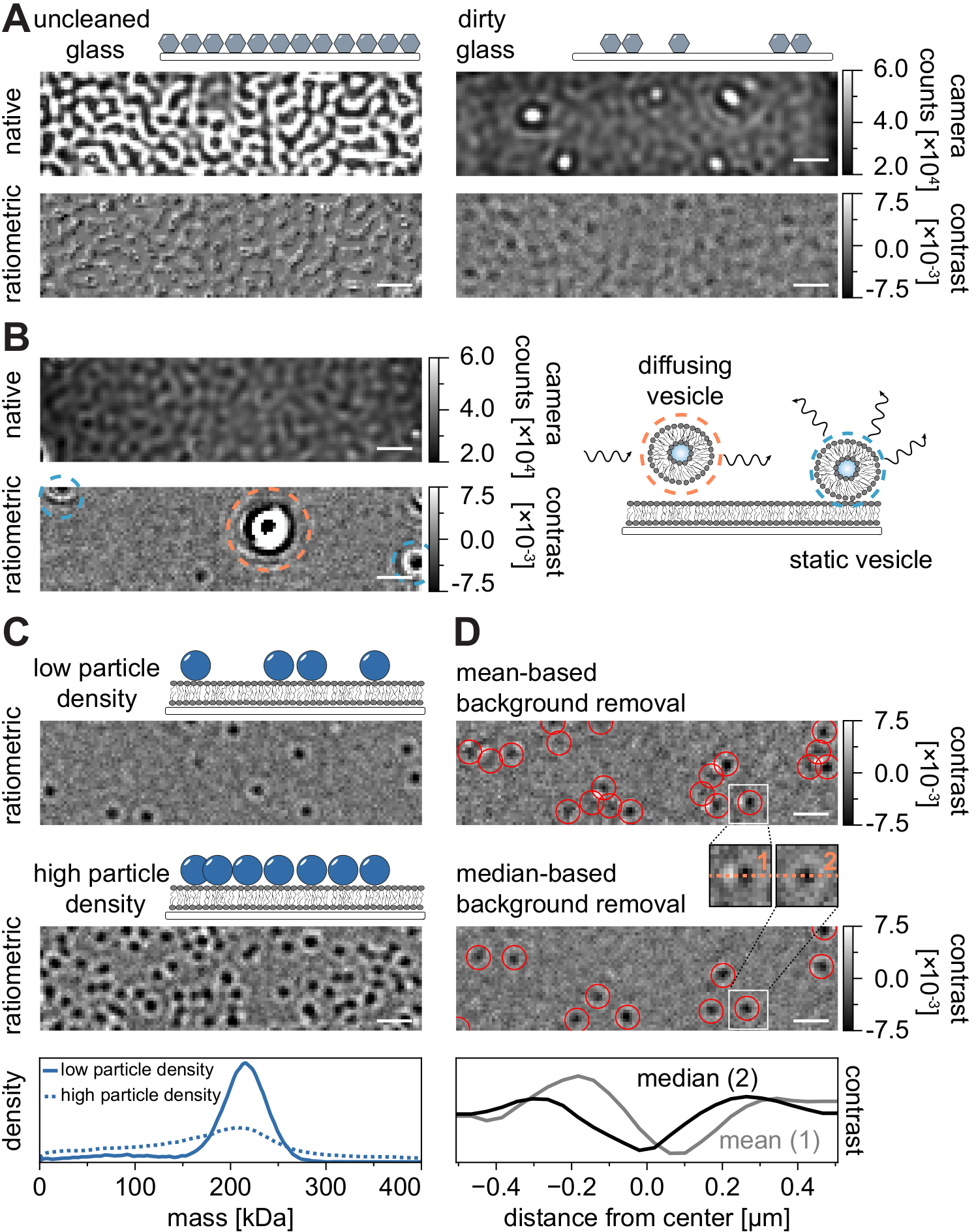
Comme pour toutes les techniques de microscopie, certaines exigences expérimentales sont cruciales pour atteindre la qualité souhaitée des données. Un exemple important dans ce contexte est le nettoyage minutieux des couvercles. En général, cela est considéré comme une condition préalable aux expériences à molécule unique liées à la microscopie, mais le MSPT est particulièrement sensible aux impuretés des échantillons. La diffusion accrue provenant de la surface vitrée des couvercles non nettoyés empêche toute mesure quantitative iSCAT. Notamment, même les particules de saleté ou de poussière résiduelles sur un verre insuffisamment nettoyé peuvent provoquer des distorsions d’image notables, reconnaissables comme des points lumineux en mode d’imagerie natif (Figure 7A). Bien que ces défauts soient éliminés par l’estimation de fond en raison de leur nature statique, la détermination précise du contraste d’une particule peut être altérée et donc influencer négativement son analyse quantitative. Un autre problème courant rencontré dans les expériences MSPT sont les vésicules restantes qui flottent (encerclées en orange) à travers le champ de vision ou les vésicules non fusionnées qui sont coincées (encerclées en bleu) à une position spécifique sur la membrane et apparaissent comme de grands diffuseurs pulsés (Figure 7B). Pour minimiser leur apparition et leur interférence avec l’acquisition de films, il est recommandé de bien laver le SLB avant d’ajouter la protéine et d’utiliser des mélanges fraîchement préparés de petites vésicules unilamellaires (VUS) et de cations divalents.
Un facteur qui doit être pris en compte pour la conception d’expériences de suivi des particules sensibles à la masse est la densité des macromolécules associées à l’interface membranaire. Des densités de particules élevées sur la membrane peuvent en effet causer des problèmes pour deux raisons : i) La liaison des détections de particules à partir de trames consécutives à des trajectoires devient ambiguë et augmente donc la probabilité d’erreurs et de coefficients de diffusion mal évalués. ii) La masse des particules, qui est extraite de l’amplitude de leur ajustement PSF correspondant, devient systématiquement sous-estimée et les pics de masse s’élargissent car la séparation du signal de fond statique du signal de particules dynamiques est de plus en plus difficile (Figure 7C). Actuellement, l’évaluation visuelle de la qualité des données dans le processus d’acquisition de vidéos MSPT est difficile sur les microscopes commerciaux disponibles car la vue ratiométrique implémentée dans le logiciel d’acquisition utilise la suppression d’arrière-plan établie pour la photométrie de masse21 au lieu de l’algorithme basé sur la médiane décrit ici et dans les références24,25 (Figure 7D ). L’élimination continue de l’arrière-plan basée sur la moyenne utilisée pour visualiser les molécules d’atterrissage en photométrie de masse fait apparaître les particules diffusantes comme des fronts sombres avec des queues brillantes, ce qui rend les taches très anisotropes et interfère avec l’ajustement PSF pendant la procédure de détection. Ainsi, l’utilisation du traitement d’image basé sur la moyenne mis en œuvre dans le logiciel d’acquisition ne convient pas à l’analyse de biomolécules diffusantes sur des membranes.

Figure 1 : Diagramme de flux de processus des différentes étapes requises pour analyser les interactions protéine-membrane avec le suivi des particules sensibles à la masse (MSPT). Pour préparer les échantillons pour les mesures MSPT, les lames de couverture en verre doivent être soigneusement nettoyées et activées avec un plasma d’oxygène. Après leur assemblage dans des chambres d’écoulement d’échantillons, de petites vésicules unilamellaires (VUS) sont préparées pour la formation de bicouche lipidique (SLB) soutenue et tous les tampons de réaction sont filtrés pour réduire la diffusion de fond. Des VUS sont ajoutés pour former des bicouches lipidiques dans les chambres d’écoulement. En option, des cations divalents tels que les ions Ca2+ peuvent être ajoutés aux VUS pour favoriser la rupture des vésicules. Enfin, de faibles concentrations de la protéine d’intérêt sont évacuées dans la chambre de réaction. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Vue native et ratiométrique de surfaces exemplaires pertinentes pour les mesures MSPT. Images représentatives de la rugosité de surface d’une lame de couverture de verre (A), lors de la formation d’une bicouche lipidique supportée (B), avec une bicouche lipidique supportée intacte (C) et de protéines exemplaires reconstituées sur un SLB (D). Les quatre exemples sont affichés en mode natif, auquel on peut accéder pendant la mesure elle-même, et en tant qu’images ratiométriques traitées après suppression de l’arrière-plan basée sur la médiane. Les barres d’échelle représentent 1 μm. Pour l’analyse des données (voir le bloc-notes Jupyter accompagné; étape 9), les paramètres suivants ont été utilisés: taille médiane de la fenêtre (window_length) = 1001. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

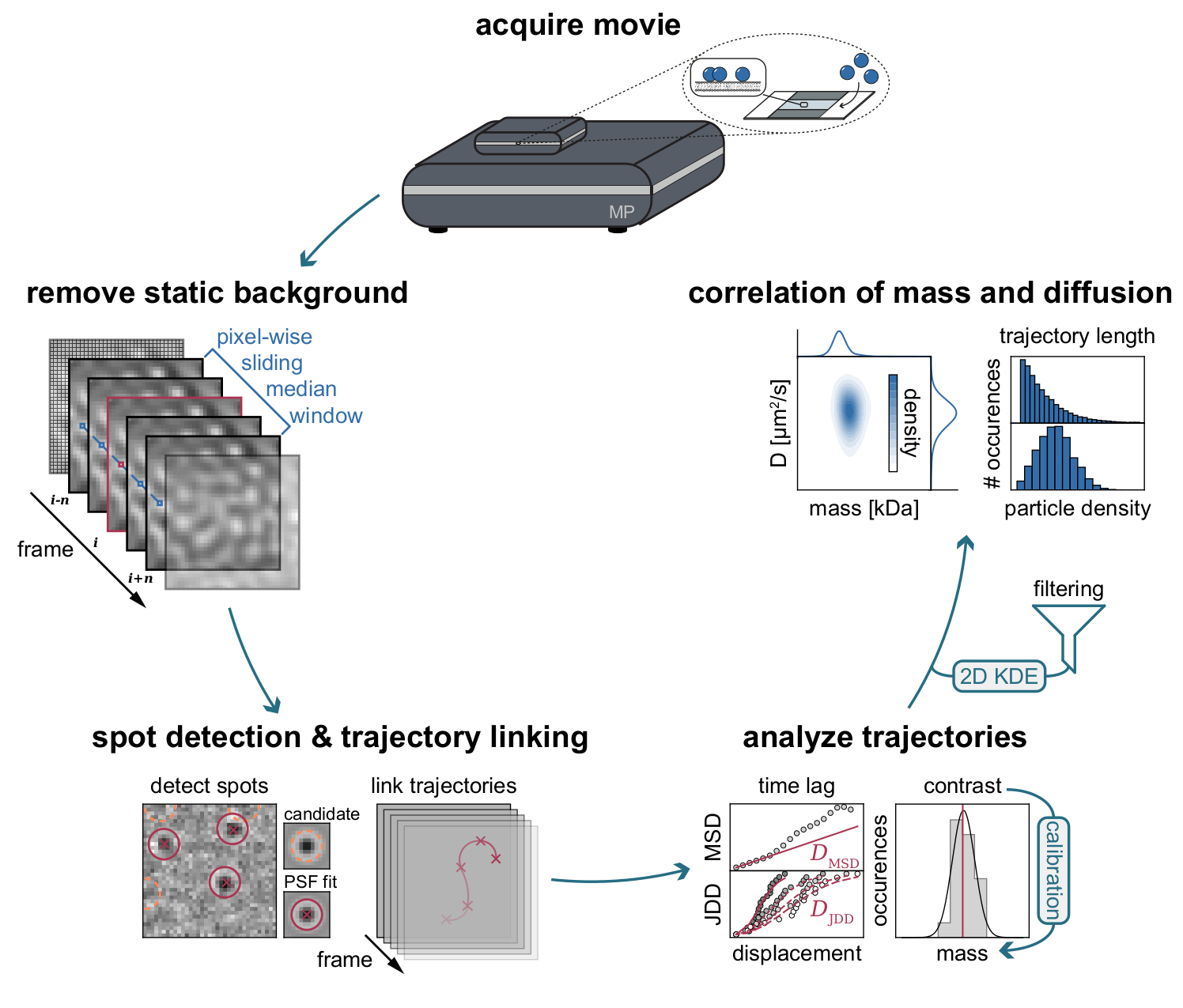
Figure 3 : Diagramme étape par étape des étapes requises pour la collecte et l’analyse des données MSPT. Après l’acquisition des données pour l’échantillon d’intérêt sur le photomètre de masse, les films sont traités pour supprimer l’arrière-plan statique grâce à une approche médiane coulissante au pixel. Par la suite, les particules candidates sont identifiées et ajustées par une fonction d’étalement ponctuel (PSF) avant leur liaison dans les trajectoires des particules. Pour permettre la détermination du coefficient de diffusion pour chaque particule, une analyse du déplacement au carré moyen (MSD) ou de la distribution de distance de saut (JDD) est utilisée. À ce stade, les valeurs de contraste peuvent être transformées en masses moléculaires selon la relation contraste-masse déterminée par la stratégie d’étalonnage. En dernière étape, les trajectoires peuvent être filtrées en fonction de leur longueur ou de la densité des particules de la membrane et visualisées par une estimation bidimensionnelle de la densité du noyau (2D-KDE). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Étalonnage de la relation masse-contraste pour les mesures MSPT. (A) Cadres ratiométriques représentatifs obtenus pour des complexes protéiques étalons de streptavidine exemplaires diffusant sur une bicouche lipidique supportée contenant un faible pourcentage de lipides biotinylés (rapport DOPC:DOPG:Biotinyl Cap PE de 70:29,99:0,01 mol%). En tant que modèles d’étalons de poids moléculaire, la streptavidinemonovalente 28 (STP uniquement) ou la streptavidinedivalente 28 en complexe avec l’albumine sérique bovine biotinylée (BSA), la protéine A biotinylée (prA), la phosphatase alcaline biotinylée (AP) ou la fibronectine biotinylée (FN) sont montrées. Les taches candidates sont surlignées en orange (cercles pointillés) et les détections de particules réussies sont surlignées en rouge (cercles pleins). Les barres d’échelle représentent 1 μm. (B) Distributions de densité de probabilité des valeurs de contraste obtenues pour les cinq protéines standard du modèle. Toutes les données affichées représentent des distributions groupées de trois expériences indépendantes par condition : STP seulement n = 82 719 ; BSA n = 9 034; prA n = 22 204; AP n = 69 065 et FN n = 71 759 trajectoires. Par rapport au nombre de particules déterminé pour les membranes contenant des protéines, le nombre de particules détectées sur une bicouche vide est négligeable à des densités membranaires modérées (figure supplémentaire 1). Les pics de contraste pris en compte pour l’étalonnage de masse sont marqués par des lignes continues tandis que les pics en pointillés représentent des états oligomères non pris en compte. (C) Courbe d’étalonnage contraste-masse dérivée des contrastes de crête dans le panneau D et des masses de séquence respectives des complexes. Les barres d’erreur affichent l’erreur-type des emplacements de pointe estimés par amorçage (100 rééchantillons de 1 000 trajectoires chacun). Pour l’analyse des données (voir bloc-notes Jupyter ; étape 9), les paramètres suivants ont été utilisés : taille médiane de la fenêtre (window_length) = 1 001 images, seuil de détection (thresh) = 0,00055, plage de recherche (dmax) = 4 pixels, mémoire (max_frames_to_vanish) = 0 images, longueur de trajectoire minimale (minimum_trajectory_length) = 7 images (STP uniquement), 9 images (BSA/FN), 15 images (prA), 10 images (AP). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 5 : Déchiffrement des états oligomères des protéines associées à la membrane. (A) Estimations 2D de la densité du noyau de la masse et du coefficient de diffusion de la streptavidine tétravalente en complexe avec de l’aldolase biotinylée (panneau de gauche) ou avec un anticorps IgG de chèvre modifié par la biotine (panneau de droite). La reconstitution des deux complexes a été réalisée sur une bicouche lipidique supportée contenant du DOPC, du DOPG et du Biotinyl Cap PE dans un rapport de 70:29,99:0,01 mol%, respectivement. Au total, 116 787 trajectoires de trois réplicats indépendants ont été incluses pour le complexe streptavidine-aldolase (densité de particules de 0,1 μm-2) et 348 405 pour le composite streptavidine-IgG (densité de particules de 0,1 μm-2). Seules les particules d’une longueur de piste d’au moins cinq images ont été incluses. Les distributions de probabilité marginale de la masse moléculaire (en haut) et du coefficient de diffusion (à droite) sont présentées. Le x noir dans les deux panneaux marque les maxima locaux respectifs du KDE. (B) Comparaison des masses oligomères déterminées pour le complexe de streptavidine tétravalente avec l’aldolase modifiée par la biotine (panneau de gauche) ou les IgG biotinylées (panneau de droite) avec, selon les masses de séquence, les poids moléculaires attendus. L’abréviation SU est introduite au nom de la sous-unité de la protéine des intérêts. Les barres d’erreur affichent l’erreur-type des emplacements de pointe estimés par amorçage (100 rééchantillons de 1 000 trajectoires chacun). Pour l’analyse des données (voir bloc-notes Jupyter accompagné; étape 9), les paramètres suivants ont été utilisés : taille médiane de la fenêtre (window_length) = 1 001 images, seuil de détection (thresh) = 0,00055, plage de recherche (dmax) = 4 pixels, mémoire (max_frames_to_vanish) = 0 images, longueur de trajectoire minimale (minimum_trajectory_length) = 5. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 6 : Dissolution du comportement diffusif des protéines natives interagissant avec la membrane annexine V (AnV) et sous-unité B (CTxB) de la toxine cholérique. (A) Estimations 2D de la densité du noyau de la masse et du coefficient de diffusion de l’annexine V (panneau de gauche) et de la sous-unité B de la toxine cholérique (panneau de droite). Pour la reconstitution des membranes AnV et CTxB, des compositions lipidiques de 80:20 mol% DOPC à DOPS et de 99,99:0,01 mol% DOPC à GM1 ont été utilisées, respectivement. Au total, 206 819 trajectoires de trois réplicats indépendants ont été incluses pour AnV (densité de particules de 0,1 μm-2) et 142 895 trajectoires pour CTxB (densité de particules de 0,2 μm-2). (B) Estimations 2D de la densité du noyau des mélanges CTxB et AnV dans un rapport de 1:1 (panneau de gauche) ou de 2:1 (panneau de droite), respectivement. La reconstitution des mélanges de protéines a été réalisée sur une bicouche lipidique supportée contenant des lipides DOPC, DOPS et GM1 dans un rapport de 80:19,99:0,01 mol%. Au total, 42 696 trajectoires de trois répétitions indépendantes ont été incluses pour le mélange 1:1 (densité de particules de 0,1 μm-2) et 264 561 trajectoires pour le rapport 2:1 (densité de particules de 0,3 μm-2). Pour (A) et (B), seules les particules d’une longueur de piste d’au moins cinq images ont été incluses. Les distributions de probabilité marginale de la masse moléculaire (en haut) et du coefficient de diffusion (à droite) sont présentées. Le x blanc dans chaque panneau marque le maximum global respectif du KDE. Pour l’analyse des données (voir bloc-notes Jupyter accompagné; étape 9), les paramètres suivants ont été utilisés : taille médiane de la fenêtre (window_length) = 1 001 images, seuil de détection (thresh) = 0,00055, plage de recherche (dmax) = 4 pixels, mémoire (max_frames_to_vanish) = 0 images, longueur de trajectoire minimale (minimum_trajectory_length) = 5. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 7: Complications potentielles au cours des mesures MSPT ou lors de l’analyse des données. (A) Images représentatives de la rugosité de surface affichées à la fois dans la vue ratiométrique native et traitée (suppression médiane de l’arrière-plan) d’une lame de verre de couverture non nettoyée. Dans les deux cas, les points lumineux constituent des impuretés de surface résiduelles qui entravent les mesures sans artefacts. (B) Images exemplaires de vésicules résiduelles dans le champ de vision après un lavage insuffisant de la membrane. Les vésicules statiques (surlignées en bleu) et diffusantes (surlignées en orange) nuiront à la qualité de mesure, soit en raison de pulsations et de remuements, soit en raison de leur mouvement directionnel, respectivement. (C) En tant que technique monoparticulaire, la MSPT nécessite de faibles densités de particules (image représentative, panneau supérieur) pour permettre une liaison et une détermination de masse appropriées de chaque particule. Dans le cas de densités de particules membranaires élevées (panneau central), l’ajustement des particules est altéré, ce qui affecte la détermination de la masse (voir panneau inférieur). (D) Images ratiométriques représentatives de particules diffusant sur une interface membranaire après élimination de fond basée sur la moyenne (panneau supérieur) ou sur la médiane. Pour les particules diffusantes, la stratégie d’élimination de l’arrière-plan basée sur la moyenne produit des images déformées du PSF de la particule, comme on peut le voir dans les petits encarts entre le panneau supérieur et le panneau central. En revanche, les PSF de particules non déformées peuvent être obtenues grâce à l’approche basée sur la médiane. Panneau inférieur : Comparaison des profils de ligne à travers le centre de la FSP obtenue après suppression de l’arrière-plan moyen ou médian. Pour toutes les images natives et ratiométriques affichées dans cette figure, les barres d’échelle représentent 1 μm. Pour l’analyse des données (voir bloc-notes Jupyter accompagné; étape 9), les paramètres suivants ont été utilisés : taille médiane de la fenêtre (window_length) = 1 001 images, seuil de détection (thresh) = 0,00055, plage de recherche (dmax) = 4 pixels, mémoire (max_frames_to_vanish) = 0 images, longueur de trajectoire minimale (minimum_trajectory_length) = 5. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Figure supplémentaire 1 : Comparaison des membranes exemptes de protéines et occupées. Images représentatives d’une bicouche lipidique soutenue intacte avant (A) et après (B) l’ajout de streptavidine purifiée (STP). Les taches candidates qui ont été ajustées avec succès au modèle PSF sont encerclées en rouge. (C) Distributions de probabilité de contraste des particules détectées sur une membrane vide (fond membranaire, gris) et sur une bicouche avec des particules de streptavidine diffusantes (bleu). Les deux distributions de probabilité représentent les données regroupées de trois expériences indépendantes avec des paramètres d’acquisition et d’analyse de films identiques. Pour l’analyse des données (voir bloc-notes Jupyter accompagné; étape 9), les paramètres suivants ont été utilisés : taille médiane de la fenêtre (window_length) = 1 001 images, seuil de détection (thresh) = 0,00055, plage de recherche (dmax) = 4 pixels, mémoire (max_frames_to_vanish) = 0 images, longueur de trajectoire minimale (minimum_trajectory_length) = 7 images. Veuillez cliquer ici pour télécharger ce fichier.
Film supplémentaire 1 : Film exemplaire montrant la rupture et la fusion des vésicules en une membrane homogène enregistrée avec le photomètre de masse. Taille médiane de la fenêtre de traitement d’image (window_length) = 1 001 images. Barre d’échelle: 1 μm. Plage de comptage de la caméra: noir = 16 892; blanc = 65 408. Veuillez cliquer ici pour télécharger ce film.
Film supplémentaire 2: Films exemplaires montrant la diffusion de complexes d’annexine V (en haut) et d’aldolase biotinylée (en bas) sur une bicouche obtenue à partir de mesures MSPT. Taille médiane de la fenêtre de traitement d’image (window_length) = 1 001 images. Barre d’échelle: 1 μm. Plage de contraste de diffusion interférométrique: noir = -0,0075; blanc = 0,0075. Veuillez cliquer ici pour télécharger ce film.
Discussion
Le protocole présenté étend la photométrie de masse21, une technique qui analyse la masse de biomolécules uniques adsorbant sur le verre, à un outil encore plus polyvalent capable de mesurer simultanément la masse et la diffusion de biomolécules membranaires non marquées. Cette extension d’analyse est obtenue grâce à la mise en œuvre d’une stratégie d’enlèvement de fond modifiée adaptée au mouvement latéral des molécules24,25. En général, l’élimination de l’arrière-plan est de la plus haute importance pour les approches basées sur iSCAT, car la forte diffusion de la rugosité de la surface du verre représente le principal obstacle à l’analyse, et la détermination précise du fond local de chaque pixel est essentielle pour la quantification de la masse et de l’emplacement des particules. Outre l’analyse d’images adaptée au mouvement des particules, la détection ultérieure des particules, la liaison de trajectoire et l’analyse des données complètent la nouvelle expansion de la MP dans le suivi des particules sensibles à la masse (MSPT).
En général, des lames de couverture en verre soigneusement nettoyées et un environnement de travail propre sont des exigences essentielles pour la réussite des expériences MSPT. En raison de l’absence de marquage des macromolécules, le signal acquis est intrinsèquement non sélectif. Des échantillons propres, ainsi qu’une manipulation appropriée des échantillons, sont donc essentiels pour s’assurer que les observations ne peuvent pas être mal interprétées. En particulier, lorsque des molécules de faible poids moléculaire sont examinées, des mesures de contrôle de membranes sans protéines sont approuvées pour évaluer les contributions de fond (figure supplémentaire 1). Outre l’inclusion de mesures de contrôle, il est donc recommandé de suivre les étapes de préparation indiquées à la figure 2 pour chaque chambre d’écoulement. Lorsqu’elles sont combinées, ces mesures de sécurité garantiront que le signal détecté provient de la biomolécule d’intérêt et non, par exemple, d’une chambre d’écoulement, d’un tampon ou d’une membrane contaminée.
Outre les précautions concernant la conception expérimentale, des précautions doivent également être prises lors du traitement de l’image MSPT. Pendant le traitement vidéo, la valeur de trois paramètres doit être choisie avec soin pour garantir des résultats corrects: i) la longueur de la fenêtre médiane pour la suppression de l’arrière-plan, ii) le seuil de détection des particules et iii) le rayon de recherche maximal pendant l’affectation de liaison. Une fenêtre médiane plus grande (i) facilite généralement la séparation des particules diffusantes de l’arrière-plan quasi constant superposé. Cependant, pour les fenêtres de trop grande taille, la dérive de l’échantillon finira par devenir perceptible et diminuera la précision de l’estimation de fond. Les réglages optimaux dépendent fortement des propriétés de l’échantillon et des conditions de mesure. Néanmoins, une valeur de 1 001 peut être utilisée comme point de départ robuste. Le paramètre de seuil (ii) doit être réglé en fonction de la masse moléculaire la plus faible attendue dans l’échantillon. Une valeur inférieure à 0,0005 n’est pas recommandée pour les mesures prises avec le photomètre de masse utilisé dans cette étude. Pour accélérer les temps d’analyse, des valeurs plus élevées peuvent être choisies si un échantillon de poids moléculaire élevé est attendu. Le rayon de recherche dans la liaison de trajectoire (iii) spécifie la distance radiale maximale en pixels dans laquelle l’emplacement décalé de la particule sera recherché dans des images consécutives. Il devrait être adapté à la particule la plus rapide de l’échantillon et, s’il était favorisé, une plage de recherche adaptative (voir la documentation de trackpy) pourrait être utilisée à la place pour réduire le temps de calcul. Surtout pendant la phase initiale d’un projet, il est recommandé de réanalyser les films avec des paramètres variables pour valider les résultats obtenus.
Compte tenu de la nature monomoléculaire de la MSPT, il convient d’éviter de mesurer à des densités de particules membranaires élevées, car celles-ci peuvent interférer avec un contraste précis et une détermination de masse. Il a été démontré que des densités inférieures à une particule par micromètre carré sont favorables aux mesures MSPT24. Une considération supplémentaire est les coefficients de diffusion attendus dans l’échantillon. Bien qu’applicable à une large gamme de coefficients de diffusion, le MSPT a une limite inférieure de coefficients de diffusion accessibles. Le confinement local dans une région de quelques pixels pendant une partie importante de la période médiane de la fenêtre fusionne la particule avec l’arrière-plan statique. Pour les conditions d’imagerie utilisées dans ce protocole, la mesure de coefficients de diffusion inférieurs à 0,01 μm2/s n’est pas recommandée. À cette vitesse de diffusion, par exemple, le déplacement moyen au carré d’une particule pendant la demi-taille de la fenêtre médiane est d’environ 4 pixels et donc de taille similaire à l’étendue du PSF. En conséquence, l’estimation statique du fond est susceptible de contenir des contributions de signal de la particule elle-même, ce qui entraîne un contraste apparemment réduit de la particule jusqu’à ce qu’elle se rapproche finalement du niveau de bruit. Cependant, des coefficients de diffusion de macromolécules compris entre 0,05 et 10 μm2/s peuvent être clairement résolus.
Pour étendre davantage la gamme des applications MSPT, on peut envisager une avancée de l’algorithme d’arrière-plan basé sur la médiane par l’élimination des pixels temporairement occupés par une particule, ou par la correction de la dérive de l’échantillon permettant des tailles de fenêtre médianes plus grandes. Les deux approches permettraient d’atténuer les problèmes liés aux mesures à des densités de particules élevées et à diffusion lente. Des améliorations en termes de sensibilité de masse plus faible sont à l’horizon avec une nouvelle génération de photomètres de masse, qui pourraient donner accès à des biomolécules inférieures à 50 kDa. Par conséquent, les futures expériences MSPT seront en mesure d’étudier la dynamique d’une seule molécule et les interactions liées à la membrane pour une gamme encore plus large de mimétismes membranaires tels que les bicouches amorties et les systèmes macromoléculaires.
Déclarations de divulgation
Les auteurs n’ont aucun conflit d’intérêts.
Remerciements
Nous apprécions sincèrement le soutien de Philipp Kukura, Gavin Young et de l’équipe du logiciel Refeyn et reconnaissons leur aide en partageant des parties du code d’analyse d’image. Nous remercions le Cryo-EM MPIB Core Facility d’avoir donné accès au photomètre de masse commercial Refeyn. F.S. remercie Jürgen Plitzko et Wolfgang Baumeister pour leur soutien et leur financement. T.H. et P.S. ont reçu un financement de la Deutsche Forschungsgemeinschaft (DFG, Fondation allemande pour la recherche) - Project-ID 201269156 - SFB 1032 (A09). N.H. a été soutenu par une subvention de retour DFG HU 2462/3-1. P.S. reconnaît le soutien du réseau de recherche MaxSynBio via l’initiative de financement conjointe du ministère fédéral allemand de l’Éducation et de la Recherche (BMBF) et de la Société Max Planck.
matériels
| Name | Company | Catalog Number | Comments |
| annexin V | Sigma Aldrich | #SRP8026 | examplary membrane-interacting protein |
| Bio-Rad Protein Assay | Bio-Rad Laboratories Inc. | #5000006 | bradford assay kit to determine protein stock concentrations |
| biotin labeled bovine albumin | Sigma Aldrich | #A8549 | examplary protein that can be used as standard protein for MSPT |
| cholera toxin subunit B | Sigma Aldrich | #SAE0069 | examplary membrane-interacting protein |
| cover glasses, #1.5, 24 x 24 mm | Paul Marienfeld GmbH & Co. KG | #0102062 | |
| cover glasses, #1.5, 24 x 60 mm | Paul Marienfeld GmbH & Co. KG | #0102242 | |
| dioleoyl-sn-glycero-3-phosphocholine (DOPC) | Avanti Polar Lipids | #850375 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| dioleoyl-sn-glycero-3-phosphoethanolamine-N-cap biotinyl (18:1 Biotinyl Cap PE | Avanti Polar Lipids | #870273 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| dioleoyl-sn-glycero-3-phosphoglycerol (DOPG) | Avanti Polar Lipids | #840475 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| dioleoyl-sn-glycero-3-phospho-L-serine (DOPS) | Avanti Polar Lipids | #840035 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| double-sided tape | tesa | #57912-00000-02 | needed for the assembly of glass sample chambers |
| Extruder | Avanti Polar Lipids | #610023 | Lipid extruder to enable monodisperse vesicle distributions |
| EZ-Link Maleimide-PEG2-Biotin | Thermo Fisher Scientific | #A39261 | maileimide-fused biotin that can be used to biotinylate standard proteins for MSPT |
| Fibronectin (Biotinylated) | Cytoskeleton Inc. | #FNR03-A | examplary protein that can be used as standard protein for MSPT |
| Gel Filtration HMW Calibration Kit | Cytiva | #28403842 | standard proteins, e.g. aldolase that can be biotinylated and used as molecular weight standards for MSPT |
| GM1 Ganglioside (Brain, Ovine-Sodium Salt) | Avanti Polar Lipids | #860065 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| Goat anti-Rabbit IgG (H+L) Secondary Antibody, Biotin | Thermo Fisher Scientific, Waltham, USA) | #31820 | examplary protein to highlight the existence of different protein states |
| Isopropanol, 99.5%, for spectroscopy | Thermo Fisher Scientific | #10003643 | |
| Low Autofluorescence Immersion Oil | Olympus K.K. | #IMMOIL-F30CC | |
| pET21a-Streptavidin-Alive | Addgene | #20860 | required to express and purify divalent streptavidin in combination with each other |
| pET21a-Streptavidin-Dead | Addgene | #20859 | required to express and purify divalent streptavidin in combination with each other |
| Pierce Alkaline Phosphatase, biotinylated | Thermo Fisher Scientific | #29339 | examplary protein that can be used as standard protein for MSPT |
| Pierce Protein A, Biotinylated | Thermo Fisher Scientific | #29989 | examplary protein that can be used as standard protein for MSPT |
| Refeyn Acquire | Refeyn Ltd. | control software for Refeyn OneMP | |
| Refeyn One | Refeyn Ltd. | - | mass photometer |
| sterile syringe filters 0.45 µm cellulose acetate membrane | VWR International | #514-0063 | needed to filter particles from the buffer of interest |
| tetravalent streptavidin | Thermo Fisher Scientific | #SNN1001 | tetravalent streptavidin to enable the presence of several biotin binding sites |
| Whatman Nuclepore Hydrophilic Membrane, 0.05 µm Pore Size, 25 mm Circle | Cytiva | #110603 | a pore size of 50 nm is recommended for supported lipid bilayer formation in the context of MSPT |
| Zepto model 2 plasma cleaner | Diener electronic GmbH | - |
Références
- Robertson, J. L. The lipid bilayer membrane and its protein constituents. Journal of General Physiology. 150 (11), 1472-1483 (2018).
- Grecco, H. E., Schmick, M., Bastiaens, P. I. H. Signaling from the Living Plasma Membrane. Cell. 144 (6), 897-909 (2011).
- Cho, W., Stahelin, R. V. Membrane-protein interactions in cell signaling and membrane trafficking. Annual Review of Biophysics and Biomolecular Structure. 34, 119-151 (2005).
- Whited, A. M., Johs, A. The interactions of peripheral membrane proteins with biological membranes. Chemistry and Physics of Lipids. 192, 51-59 (2015).
- Gonzalez, L., Scheller, R. H. Regulation of membrane trafficking: Structural insights from a Rab/effector complex. Cell. 96 (6), 755-758 (1999).
- Sezgin, E., Levental, I., Mayor, S., Eggeling, C. The mystery of membrane organization: composition, regulation and roles of lipid rafts. Nature Reviews Molecular Cell Biology. 18 (6), 361-374 (2017).
- Bagheri, Y., Ali, A. A., You, M. Current methods for detecting cell membrane transient interactions. Frontiers in Chemistry. 8, 603259 (2020).
- Miller, H., Zhou, Z., Shepherd, J., Wollman, A. J. M., Leake, M. C. Single-molecule techniques in biophysics: A review of the progress in methods and applications. Reports on Progress in Physics. 81 (2), 024601 (2018).
- Manzo, C., Garcia-Parajo, M. F. A review of progress in single particle tracking: From methods to biophysical insights. Reports on Progress in Physics. 78 (12), 124601 (2015).
- Gelles, J., Schnapp, B. J., Sheetz, M. P. Tracking kinesin-driven movements with nanometre-scale precision. Nature. 331 (6155), 450-453 (1988).
- Funatsu, T., Harada, Y., Tokunaga, M., Saito, K., Yanagida, T. Imaging of single fluorescent molecules and individual ATP turnovers by single myosin molecules in aqueous solution. Nature. 374 (6522), 555-559 (1995).
- Schmidt, T., Schütz, G. J., Baumgartner, W., Gruber, H. J., Schindler, H. Imaging of single molecule diffusion. Proceedings of the National Academy of Sciences of the United States of America. 93 (7), 2926-2929 (1996).
- Taylor, R. W., et al. Interferometric scattering microscopy reveals microsecond nanoscopic protein motion on a live cell membrane. Nature Photonics. 13 (7), 480-487 (2019).
- Kukura, P., et al. High-speed nanoscopic tracking of the position and orientation of a single virus. Nature Methods. 6 (12), 923-927 (2009).
- Jacobsen, V., Stoller, P., Brunner, C., Vogel, V., Sandoghdar, V. Interferometric optical detection and tracking of very small gold nanoparticles at a water-glass interface. Optics Express. 14 (1), 405 (2006).
- Ueno, H., et al. Simple dark-field microscopy with nanometer spatial precision and microsecond temporal resolution. Biophysical Journal. 98 (9), 2014-2023 (2010).
- Loose, M., Fischer-Friedrich, E., Herold, C., Kruse, K., Schwille, P. Min protein patterns emerge from rapid rebinding and membrane interaction of MinE. Nature Structural and Molecular Biology. 18 (5), 577-583 (2011).
- Bezeljak, U., Loya, H., Kaczmarek, B., Saunders, T. E., Loose, M. Stochastic activation and bistability in a Rab GTPase regulatory network. Proceedings of the National Academy of Sciences of the United States of America. 117 (12), 6504-6549 (2020).
- Ha, T., Tinnefeld, P. Photophysics of fluorescent probes for single-molecule biophysics and super-resolution imaging. Annual Review of Physical Chemistry. 63 (1), 595-617 (2012).
- Garcia-Parajo, M. F., Segers-Nolten, G. M. J., Veerman, J. A., Greve, J., Van Hulst, N. F. Real-time light-driven dynamics of the fluorescence emission in single green fluorescent protein molecules. Proceedings of the National Academy of Sciences of the United States of America. 97 (13), 7237-7242 (2000).
- Young, G., et al. Quantitative mass imaging of single biological macromolecules. Science. 360 (6387), 423-427 (2018).
- Piliarik, M., Sandoghdar, V. Direct optical sensing of single unlabelled proteins and super-resolution imaging of their binding sites. Nature Communications. 5, 4495 (2014).
- Cole, D., Young, G., Weigel, A., Sebesta, A., Kukura, P. Label-free single-molecule imaging with numerical-aperture-shaped interferometric scattering microscopy. ACS Photonics. 4 (2), 211-216 (2017).
- Heermann, T., Steiert, F., Ramm, B., Hundt, N., Schwille, P. Mass-sensitive particle tracking to elucidate the membrane-associated MinDE reaction cycle. Nature Methods. 18 (10), 1239-1246 (2021).
- Foley, E. D. B., Kushwah, M. S., Young, G., Kukura, P. Mass photometry enables label-free tracking and mass measurement of single proteins on lipid bilayers. Nature Methods. 18 (10), 1247-1252 (2021).
- Voss, O. H., Lee, H. N., Tian, L., Krzewski, K., Coligan, J. E. Liposome preparation for the analysis of lipid-receptor interaction and efferocytosis. Current Protocols in Immunology. 120, 1-21 (2018).
- Pincet, F., et al. FRAP to characterize molecular diffusion and interaction in various membrane environments. PLoS ONE. 11 (7), 0158457 (2016).
- Howarth, M., et al. A monovalent streptavidin with a single femtomolar biotin binding site. Nature Methods. 3 (4), 267-273 (2006).
- Allan, D., et al. soft-matter/trackpy: Trackpy v.0.5.0. Zenodo. , (2021).
- Weimann, L., et al. A quantitative comparison of single-dye tracking analysis tools using Monte Carlo simulations. PLoS ONE. 8 (5), 64287 (2013).
- Michalet, X. Mean square displacement analysis of single-particle trajectories with localization error: Brownian motion in an isotropic medium. Physical Review E - Statistical, Nonlinear, and Soft Matter Physics. 82 (4), 041914 (2010).
- Sygusch, J., Beaudry, D., Allaire, M. Molecular architecture of rabbit skeletal muscle aldolase at 2.7-A resolution. Proceedings of the National Academy of Sciences of the United States of America. 84 (22), 7846-7850 (1987).
- Heath, G. R., Scheuring, S. High-speed AFM height spectroscopy reveals µs-dynamics of unlabeled biomolecules. Nature Communications. 9 (1), 4983 (2018).
- Day, C. A., Kenworthy, A. K. Mechanisms underlying the confined diffusion of cholera toxin B-subunit in intact cell membranes. PLOS ONE. 7 (4), 34923 (2012).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.