
Method Article
Tracciamento delle particelle sensibili alla massa per caratterizzare la dinamica delle macromolecole associate alla membrana
In questo articolo
Riepilogo
Questo protocollo descrive un approccio basato sull'elaborazione delle immagini e sul tracciamento di singole particelle basato su iSCAT che consente l'indagine simultanea della massa molecolare e del comportamento diffusivo delle macromolecole che interagiscono con le membrane lipidiche. Le istruzioni dettagliate per la preparazione del campione, la conversione da massa a contrasto, l'acquisizione di filmati e la post-elaborazione sono fornite insieme alle indicazioni per prevenire potenziali insidie.
Abstract
Le interazioni di breve durata o transitorie delle macromolecole a e con le membrane lipidiche, un'interfaccia in cui si verificano una moltitudine di reazioni biologiche essenziali, sono intrinsecamente difficili da valutare con metodi biofisici standard. L'introduzione del tracciamento delle particelle sensibili alla massa (MSPT) costituisce un passo importante verso una caratterizzazione quantitativa approfondita di tali processi. Tecnicamente, ciò è stato reso possibile attraverso l'avvento della fotometria di massa (MP) basata sulla microscopia a dispersione interferometrica (iSCAT). Quando la strategia di rimozione dello sfondo è ottimizzata per rivelare il movimento bidimensionale delle particelle associate alla membrana, questa tecnica consente l'analisi in tempo reale sia della diffusione che della massa molecolare di macromolecole non etichettate su membrane biologiche. Qui viene descritto un protocollo dettagliato per eseguire e analizzare il tracciamento delle particelle sensibili alla massa dei sistemi associati alla membrana. Le misurazioni eseguite su un fotometro di massa commerciale raggiungono una risoluzione temporale nel regime dei millisecondi e, a seconda del sistema MP, un limite di rilevamento di massa fino a 50 kDa. Per mostrare il potenziale dell'MSPT per l'analisi approfondita della dinamica delle macromolecole catalizzate da membrana in generale, vengono presentati i risultati ottenuti per sistemi proteici esemplari come l'annessina V dell'interattore di membrana nativo.
Introduzione
Un tempo semplicemente percepite come una barriera contro l'ampia gamma di condizioni fisiche ambientali, le membrane biologiche sono oggi considerate entità funzionali e piattaforme catalitiche 1,2. Sulla base della loro capacità di localizzare, amplificare e dirigere i segnali in risposta alle reazioni macromolecole associate alla membrana, le interfacce lipidiche costituiscono un elemento cruciale per un'ampia varietà di processi cellulari come il traffico di membrana e le cascate di segnalazione 3,4,5. Fungendo da sito di nucleazione per l'assemblaggio di complessi stabili, l'attaccamento della membrana si basa spesso su un equilibrio dinamico tra forme di macromolecole associate alla membrana e citosoliche ed è quindi di natura transitoria 6,7.
Nonostante la loro grande importanza in biologia, finora è stato difficile sviluppare metodi in grado di fornire l'accesso alle eterogeneità compositive, spaziali e temporali delle reazioni macromolecole associate alla membrana in tempo reale 7,8. Per risolvere i processi molecolari sottostanti, due aspetti sperimentali sono decisivi: una risoluzione temporale sufficiente e la sensibilità a singola particella. Pertanto, le tecniche medie di insieme come il recupero della fluorescenza dopo il fotosbiancamento (FRAP) ma anche la spettroscopia di correlazione a fluorescenza molto più sensibile (FCS) hanno dei limiti, poiché disaccoppiano in gran parte le informazioni spaziali dalle informazioni temporali9. Un passo importante verso la caratterizzazione della dinamica delle singole molecole è stato quindi l'avvento del tracciamento a singola particella (SPT) in combinazione con la microscopia altamente sensibile. In particolare, due approcci SPT si sono dimostrati efficaci in questo senso. In primo luogo, l'utilizzo dei fluorofori come etichette e i corrispondenti sistemi di rilevamento della fluorescenza hanno spianato la strada alla precisione nanometrica e alla risoluzione temporale al millisecondo 10,11,12. In secondo luogo, il rilevamento basato sullo scattering utilizzando nanoparticelle d'oro ha migliorato sia la precisione di localizzazione che la risoluzione temporale nell'intervallo sub-nanometro e microsecondo, rispettivamente 13,14,15,16. Nonostante i numerosi vantaggi di entrambi gli approcci e i loro contributi significativi per quanto riguarda la comprensione meccanicistica dei sistemi associati alla membrana17,18, entrambe le tecniche sono state finora limitate: richiedono l'etichettatura delle molecole di interesse, che potenzialmente perturba il loro comportamento nativo e sono insensibili alla composizione molecolare delle particelle associate alla membrana19,20.
Entrambe queste limitazioni sono state recentemente superate con l'introduzione di un nuovo approccio basato sullo scattering interferometrico (iSCAT) chiamato fotometria di massa (MP)21,22,23. Questa tecnica consente la determinazione di distribuzioni di massa in soluzione di biomolecole in base al loro contrasto iSCAT quando atterrano su un'interfaccia di vetro. Tuttavia, per il rilevamento e la caratterizzazione di molecole mobili che si diffondono sulle membrane lipidiche, è stato necessario sviluppare un approccio di analisi delle immagini più sofisticato. Questo nel frattempo è stato implementato con successo e consente di rilevare, tracciare e determinare la massa molecolare di singole biomolecole non etichettate che si diffondono su un'interfaccia lipidica24,25. Denominata fotometria di massa dinamica o tracciamento delle particelle sensibili alla massa (MSPT), questa tecnica consente ora la valutazione di complesse interazioni macromolecole registrando direttamente i cambiamenti nella massa molecolare delle entità tracciate e quindi apre nuove possibilità per l'analisi meccanicistica della dinamica molecolare associata alla membrana.
Qui viene presentato un protocollo dettagliato per la preparazione dei campioni, l'imaging e la pipeline di analisi dei dati necessaria per MSPT. In particolare, vengono discussi i requisiti del campione e i potenziali problemi che possono verificarsi durante la misurazione e l'analisi. Inoltre, l'impareggiabile potenziale di analizzare i sistemi di macromolecole che interagiscono con la membrana è mostrato attraverso vari risultati rappresentativi.
Protocollo
1. Preparazione del campione
- Generazione di vescicole multilamellari (MLV)
- Calcolare la quantità di lipidi disciolti dal cloroformio in base alla miscela lipidica desiderata e al volume di sospensione richiesto.
NOTA: Una concentrazione finale di vescicola di 4 mg/ml di lipidi è raccomandata per il tampone di risospensione (reazione). - Pipettare il volume calcolato di lipidi in un flaconcino di vetro da 1,5 ml utilizzando pipette a spostamento positivo dotate di punte di vetro.
- Evaporare il solvente lipidico sotto un debole flusso di azoto e ruotare costantemente la fiala per garantire un'equa distribuzione dei lipidi sulle pareti di vetro.
- Garantire la completa evaporazione del solvente posizionando il flaconcino sotto un flusso costante di azoto per 15 minuti.
- Rimuovere le tracce residue di cloroformio mediante essiccazione sottovuoto in un essiccatore sottovuoto per un'ora aggiuntiva.
- Reidratare la miscela lipidica nel tampone di risospensione (reazione) desiderato e ruotare accuratamente la sospensione fino a quando il film lipidico non è stato sciolto dalle pareti del flaconcino.
NOTA: Il tampone di reazione deve garantire l'attività e la stabilità delle proteine. Il tampone di reazione utilizzato in questo studio contiene 50 mM Tris-HCl (pH = 7,5), 150 mM KCl e 5 mM MgCl2. Si noti che qualsiasi tampone utilizzato per diluire lipidi o proteine deve essere filtrato per rimuovere le impurità particolato interferenti (vedere il passaggio 5).
- Calcolare la quantità di lipidi disciolti dal cloroformio in base alla miscela lipidica desiderata e al volume di sospensione richiesto.
- Generazione di piccole vescicole unilamellari (SUV)
- Per cicli consecutivi di congelamento-scongelamento della risospensione lipidica (fase 1.1.6), far bollire 500 ml di acqua in un becher su una piastra calda (tra 70 °C e 99 °C) e preparare un contenitore con azoto liquido.
- Shock-congelare la risospensione lipidica nell'azoto liquido. Trasferire il flaconcino nel becher con acqua calda fino a quando la soluzione non è completamente scongelata. Ripetere questo ciclo di congelamento-scongelamento 8-10 volte o fino a quando la miscela precedentemente torbida appare chiara.
ATTENZIONE: utilizzare indumenti e attrezzature di sicurezza adeguati come occhiali, guanti e pinzette per evitare qualsiasi contatto diretto con l'azoto liquido, la fiala lipidica congelata o l'acqua bollente. - Per la generazione di una distribuzione di vescicole monodisperse, assemblare un estrusore lipidico e testarne l'integrità con tampone di reazione per assicurarsi che non perda.
NOTA: se si osserva una perdita, riassemblare con cura l'estrusore lipidico fino a quando non è evidente alcuna fuoriuscita tampone. - Estrudere la sospensione lipidica per 37 passaggi attraverso una membrana nucleoporica con una dimensione dei pori di 50 nm26. Il numero di passaggi dovrebbe essere irregolare per garantire che la miscela SUV finale attraversi la membrana nucleoporica e sia quindi priva di aggregati lipidici o vescicole multilamellari. Le vescicole estruse saranno utilizzate in seguito per formare doppi strati lipidici supportati (vedere i passaggi 6 e 7).
NOTA: i SUV possono anche essere formati dalla sonicazione della miscela lipidica reidratata. Tuttavia, la preparazione tramite estrusione fornisce una distribuzione più monodispersa dei SUV, che facilita la rottura delle vescicole durante la formazione di doppio strato lipidico supportato. Le vescicole estruse possono essere conservate in frigorifero per un massimo di 3 giorni.
2. Pulizia dei vetrini del microscopio
- Distribuire un numero uguale di vetrini per microscopio (n. #1.5; spessore 0,17 mm) con dimensioni di 24 mm x 60 mm e 24 mm x 24 mm in portamicroscopio in politetrafluoretilene (PTFE).
- Trasferire i supporti in PTFE in bicchieri contenenti acqua ultrapura e sonicarli per 15 minuti a temperatura ambiente.
NOTA: A seconda del becher, il volume d'acqua deve essere regolato per coprire completamente il supporto in PTFE. - Utilizzare una pinzetta per rimuovere i supporti dal becher e sostituire l'acqua con isopropanolo ultrapuro. Inserire il supporto nel becher contenente isopropanolo e sonicare di nuovo per 15 min.
NOTA: A seconda del becher, il volume dell'isopropanolo deve essere regolato per coprire completamente il supporto in PTFE. - Sostituire l'isopropanolo con acqua ultrapura e sonicare il becher contenente i supporti per 15 min.
- Rimuovere i supporti in PTFE dai becher e asciugare i vetrini del microscopio nel supporto sotto un flusso costante di azoto gassoso o aria compressa.
NOTA: Garantire una corretta pulizia delle diapositive di copertura utilizzando guanti, bicchieri puliti e pellicola di paraffina per coprire ogni becher. In caso contrario, la polvere residua potrebbe causare fluttuazioni di fondo significative durante le misurazioni MSPT.
3. Idrofilizzazione dei vetrini del microscopio
NOTA: Per ottenere un doppio strato lipidico omogeneo e fluido, l'idrofilizzazione dei vetrini è essenziale e deve essere effettuata poco prima del montaggio della camera di flusso.
- Posizionare i supporti in PTFE contenenti solo vetrini per microscopio da 24 mm x 60 mm in un pulitore al plasma con ossigeno come gas di processo e pulire i vetrini del microscopio con plasma (parametri utilizzati in questo lavoro: potenza del 30%, pressione dell'ossigeno 0,3 mbar per 30 s; vedere Tabella dei materiali per i dettagli del pulitore al plasma utilizzato).
NOTA: Per ottenere membrane fluide, i parametri di pulizia al plasma come potenza, pressione dell'ossigeno e tempo di pulizia devono essere regolati per ogni strumento. A tale scopo, si raccomanda l'uso di lipidi marcati fluorescentemente per garantire la fluidità della membrana, che può essere quantificata con il recupero della fluorescenza dopo esperimenti di fotosbiancamento (FRAP)27. Se i parametri non sono ottimizzati per la rispettiva configurazione, la diffusione della membrana potrebbe essere compromessa a causa della ridotta fluidità della membrana.
4. Assemblaggio delle camere di flusso
- Prima del montaggio della camera di flusso, tenere pronti i seguenti componenti: vetrini per microscopio puliti (24 mm x 24 mm), vetrini per microscopio idrofilizzati (24 mm x 60 mm), foglio di alluminio, cartone piatto, bisturi e nastro biadesivo.
- Avvolgere il cartone piatto con un foglio di alluminio.
- Stendere i vetrini puliti per microscopio da 24 mm x 24 mm sul foglio di alluminio con una distanza sufficiente l'uno dall'altro.
- Attaccare strisce di nastro biadesivo ai bordi superiore e inferiore delle diapositive.
- Asportare ogni vetrino del microscopio con il bisturi, in modo tale che possa essere rimosso dal foglio di alluminio. Di conseguenza, ogni diapositiva dovrebbe avere strisce di nastro biadesivo attaccate ai bordi superiore e inferiore della diapositiva (vedere la Figura 1).
- Collegare il vetrino da 24 mm x 24 mm con le due strisce di nastro biadesivo al vetrino idrofilizzato da 24 mm x 60 mm per formare un percorso di flusso tra i vetrini del microscopio più piccoli e quelli più grandi.
NOTA: per garantire camere di flusso pulite, indossare costantemente guanti e assicurarsi che il banco di lavoro sia privo di polvere.
5. Filtrazione dei tamponi di reazione
- Filtrare sterili tutti i tamponi di reazione attraverso membrane di acetato di cellulosa da 0,45 μm per garantire un segnale di fondo minimo durante le misurazioni MSPT.
NOTA: Se la presenza di nucleotidi, come l'ATP, è essenziale per un esperimento di successo, essere consapevoli di un potenziale aumento del segnale di fondo. Si raccomanda di utilizzare solo quantità minime che garantiscano comunque l'attività proteica.
6. Formazione di doppio strato lipidico supportato (SLB)
NOTA: Si raccomanda di eseguire la formazione di doppi strati lipidici supportati sul fotometro di massa per garantire visivamente il successo della diffusione delle vescicole e la completa rimozione delle vescicole non fuse.
- Diluire i SUV appena estrusi (vedere il punto 1 per maggiori dettagli) a una concentrazione finale di 0,4 mg/mL nel tampone di reazione richiesto. Opzionalmente, per favorire la rottura delle vescicole, aggiungere 2 mM CaCl2 alla sospensione della vescicola.
NOTA: i cationi bivalenti potrebbero causare l'aggregazione di alcuni lipidi come il PiP2. Per le miscele contenenti tali lipidi, astenersi dall'utilizzare CaCl2 per la promozione della rottura delle vescicole o di altri cationi bivalenti nel tampone di risospensione. Se necessario per l'esperimento, i cationi bivalenti possono essere aggiunti dopo la formazione riuscita del doppio strato lipidico supportato. - Lavare 50 μL della sospensione delle vescicole nella camera di flusso (fase 4) e incubare la camera per 2 minuti.
NOTA: tamponi, vescicole o soluzioni proteiche possono essere lavati attraverso la camera di flusso con un piccolo pezzo di tessuto in ammollo. Tuttavia, è anche possibile utilizzare un sistema di pompaggio meccanico. - Rimuovere le vescicole non fuse attraverso il lavaggio ripetuto (almeno tre volte) della camera di flusso con 200 μL del tampone di reazione ogni volta.
NOTA: le vescicole devono essere accuratamente lavate fuori dalla camera di flusso per garantire un segnale di fondo stabile durante le misurazioni MSPT.
7. Generazione della curva di calibrazione
NOTA: Per convertire il contrasto delle particelle rilevate in massa molecolare, il loro segnale deve essere calibrato utilizzando proteine di dimensioni note. Si raccomanda di regolare il regime di dimensione standard delle proteine per coprire la gamma di masse molecolari previste per il sistema di interesse.
- Biotinilazione di proteine standard con un residuo di cisteina
- Calcolare la quantità appropriata di maleimide-biotina per la proteina standard secondo le istruzioni del produttore.
- Incubare la proteina standard con il volume determinato di maleimide-biotina per 1 ora a temperatura ambiente.
- Per rimuovere la maleimmide-biotina non coniugata dal complesso biotina-proteina coniugata, eseguire la cromatografia di esclusione dimensionale su una colonna adatta alla proteina di interesse.
- Determinare la concentrazione proteica utilizzando un saggio di Bradford.
NOTA: per conservare la proteina standard per ulteriori misurazioni, congelare le proteine in aliquote monouso in azoto liquido e conservarle a -80 °C.
- Misura di proteine standard per la curva di calibrazione
- In una camera di flusso, preparare un doppio strato lipidico supportato con SUV estrusi da 0,4 mg/mL (vedere i passaggi 1 e 6 per maggiori dettagli) contenente 0,01 mol% (v/v) Biotinyl Cap PE (1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-cap biotinyl).
- Aggiungere 50 μL di streptavidina bivalente da 2,5 nM al doppio strato nella camera di flusso e incubare per 10 minuti.
NOTA: La streptavidina bivalente è stata espressa e purificata come delineato in Howarth et al.28. Anche la streptavidina tetravalente può essere utilizzata. Tuttavia, l'uso di streptavidina bivalente può ridurre le possibili stechiometrie di reazione tra lipidi biotinilati e proteine standard che sono coniugate a una porzione di biotina al fine di facilitare l'assegnazione delle specie. - Rimuovere la streptavidina bivalente non legata con 100 μL di tampone di reazione.
- Aggiungere 50 μL di proteina standard coniugata con biotina 100 nM al doppio strato nella camera di flusso e incubare per 2 minuti.
NOTA: A seconda dell'efficienza della biotinilazione e se viene utilizzata streptavidina di- o tetravalente, le concentrazioni ottimali di proteina standard coniugata con biotina e streptavidina possono variare. - Eseguire la misurazione MSPT in base ai dettagli descritti nel passaggio 8.
ATTENZIONE: le condizioni di imaging devono essere identiche sia per gli standard di campionamento che per quelli di calibrazione.
8. Imaging
- Formazione di SLB e preparazione del campione
- Come descritto più dettagliatamente nella fase 6, introdurre i SUV della miscela lipidica desiderata (25 μL) nella camera di flusso del campione e formare un doppio strato lipidico supportato. Lavare accuratamente la camera (tre volte) con 100 μL di tampone di reazione per rimuovere tutte le vescicole non fuse.
- Aggiungere 50 μL della proteina di interesse alla camera del campione.
NOTA: poiché MSPT è un metodo a singola particella, la concentrazione proteica deve essere mantenuta nell'intervallo da pM a nM per consentire il rilevamento e il tracciamento indisturbati delle particelle.
- Acquisizione video
- Impostare le condizioni di imaging desiderate, ad esempio la dimensione del campo visivo (FOV), la frequenza dei fotogrammi, il tempo di esposizione e il tempo di acquisizione nel software di acquisizione.
NOTA: le seguenti impostazioni hanno dimostrato di funzionare per MSPT su un fotometro di massa commerciale (vedere Tabella dei materiali): FOV di 128 pixel x 35 pixel, frame rate di 1 kHz con conseguente circa 200 fotogrammi al secondo dopo la successiva media di fotogrammi 5 volte e un tempo di esposizione di 0,95 ms. - Regola la messa a fuoco automaticamente o manualmente. Se necessario, spostare il FOV in una posizione con una membrana omogenea utilizzando il controllo laterale.
- Creare una cartella di progetto e iniziare a registrare il filmato. Al termine della registrazione, specificare un nome di file nella finestra di dialogo richiesta dal software di acquisizione. Il filmato viene quindi salvato automaticamente nella cartella del progetto come file MP per la successiva analisi.
NOTA: Registrare almeno tre repliche in diverse camere di flusso per garantire l'integrità delle singole membrane e la riproducibilità dei risultati. La durata del film può essere impostata in anticipo e dipende dal tipo di esperimento. Nella maggior parte dei casi, si consiglia un tempo di acquisizione compreso tra 5 minuti e 7 minuti.
ATTENZIONE: per impostazione predefinita, le registrazioni di filmati sul software di acquisizione di fotometri di massa commerciali vengono compresse prima di essere salvate per ridurre lo spazio di archiviazione. Tuttavia, la compressione dei file deve essere disattivata per consentire l'analisi dei dati personalizzata come descritto in questo protocollo. I dettagli su come disattivare la compressione dei file sono disponibili nel manuale utente del produttore.
- Impostare le condizioni di imaging desiderate, ad esempio la dimensione del campo visivo (FOV), la frequenza dei fotogrammi, il tempo di esposizione e il tempo di acquisizione nel software di acquisizione.
9. Analisi dei dati
NOTA: la pipeline di analisi dei dati è accompagnata da due notebook Jupyter interattivi (MSPT analysis.ipynb, Movie visualization.ipynb). I notebook Jupyter e i moduli Python personalizzati associati necessari per eseguire l'analisi MSPT descritta di seguito sono disponibili in un repository pubblico: https://github.com/MSPT-toolkit/MSPT-toolkit. Per istruzioni dettagliate sull'analisi di seguito, i lettori sono indirizzati a MSPT analysis.ipynb accessibile utilizzando il link sopra.
- Elaborazione video
- Rimuovi la dispersione statica dominante della luce con l'algoritmo di stima dello sfondo in termini di pixel utilizzando la funzione image_processing.mp_reader .
- Per applicare la rimozione dello sfondo, scegliere l'opzione continuous_median per la modalità parametro e impostare una lunghezza appropriata per la finestra mediana scorrevole (window_length) nella sezione B.1 del blocco appunti. Facoltativamente, salvate i filmati dopo la rimozione dello sfondo da utilizzare per il rilevamento delle particelle e il collegamento della traiettoria (impostando il parametro save_processed_movies su True).
NOTA: regolare le dimensioni della finestra (window_length) su valori compresi tra 101 e 2001 a seconda della densità delle particelle sulla membrana, del coefficiente di diffusione previsto, della frequenza di acquisizione e della velocità di elaborazione richiesta.
ATTENZIONE: La strategia di rimozione dello sfondo funziona bene se la membrana non è troppo densamente imballata e se la diffusione delle particelle è sufficientemente veloce (cioè, ogni pixel non è il più delle volte occupato da una particella). In caso contrario, il contrasto delle particelle sarà sistematicamente sottovalutato in quanto non possono essere adeguatamente distinte dal segnale di fondo. Questo può essere compensato aumentando la dimensione mediana della finestra al costo della velocità computazionale. Tuttavia, tenere presente che l'impostazione di dimensioni troppo grandi della finestra può influire negativamente sull'output a causa della deriva del campione. Un'ispezione visiva dei video elaborati è fondamentale.
- Per applicare la rimozione dello sfondo, scegliere l'opzione continuous_median per la modalità parametro e impostare una lunghezza appropriata per la finestra mediana scorrevole (window_length) nella sezione B.1 del blocco appunti. Facoltativamente, salvate i filmati dopo la rimozione dello sfondo da utilizzare per il rilevamento delle particelle e il collegamento della traiettoria (impostando il parametro save_processed_movies su True).
- Rileva le particelle e la loro rispettiva posizione durante il filmato utilizzando la funzione particle_fitting.particle_fitter (vedere la sezione B.2 del notebook).
- Regolare la sensibilità del rilevamento delle particelle con il parametro di soglia (trebbiatura; vedere la sezione B.1 del notebook), che viene utilizzato per evidenziare i punti candidati mediante binarizzazione dell'immagine. L'effetto dei diversi parametri di soglia sulla sensibilità di rilevamento spot può essere esaminato in un notebook separato (Movie visualization.ipynb). I risultati del rilevamento delle particelle vengono automaticamente salvati in file CSV in una sottodirectory del file filmato.
NOTA: l'impostazione del parametro di soglia arbitrariamente bassa (ad esempio, per i filmati ripresi con il fotometro di massa utilizzato, un parametro di soglia inferiore a 0,0005) non è consigliabile in quanto i punti candidati saranno dominati da rumore spurio e quindi prolungheranno il tempo di elaborazione.
- Regolare la sensibilità del rilevamento delle particelle con il parametro di soglia (trebbiatura; vedere la sezione B.1 del notebook), che viene utilizzato per evidenziare i punti candidati mediante binarizzazione dell'immagine. L'effetto dei diversi parametri di soglia sulla sensibilità di rilevamento spot può essere esaminato in un notebook separato (Movie visualization.ipynb). I risultati del rilevamento delle particelle vengono automaticamente salvati in file CSV in una sottodirectory del file filmato.
- Rimuovi la dispersione statica dominante della luce con l'algoritmo di stima dello sfondo in termini di pixel utilizzando la funzione image_processing.mp_reader .
- Collega particelle in fotogrammi consecutivi in traiettorie usando il pacchetto Python trackpy (v.0.5.0)29.
NOTA: il collegamento della traiettoria viene eseguito al volo dopo il rilevamento dello spot. Di conseguenza, un file CSV aggiuntivo contenente le informazioni sulla traiettoria viene memorizzato in una sottodirectory del file CSV di rilevamento delle particelle.- Rimuovere le traiettorie con troppo pochi punti utilizzando il parametro minimum_trajectory_length (vedere la sezione B.1 del notebook) per consentire una solida determinazione dei coefficienti di diffusione. Per spiegazioni dettagliate sugli altri parametri delle funzioni trackpy , fare riferimento alla documentazione di trackpy .
- Analisi della traiettoria
- Nella sezione C.1 del blocco appunti, specificare la frequenza fotogrammi (frame_rate) e la dimensione dei pixel in nm (pixel_size), utilizzate per l'acquisizione di filmati. Creare un elenco di file CSV contenenti le informazioni sulla traiettoria restituite da trackpy (vedere il passaggio 9.2) con la funzione trajectory_analysis.get_csv_files (sezione notebook C.2).
- Inoltre, specificare un nome di file di output per il contenitore HDF5, che viene utilizzato per memorizzare i risultati di raccordo su disco (sezione notebook C.3). Analizzate tutte le traiettorie con la funzione trajectory_analysis.fit_trajectories nella sezione C.4 del blocco appunti, che scorre l'elenco dei file CSV. Questa funzione utilizza la distribuzione della distanza di salto (JDD)30 e l'analisi dello spostamento quadrato medio (MSD)31 per stimare il coefficiente di diffusione di ciascuna traiettoria.
- Convertire il contrasto mediano di ciascuna traiettoria nella massa corrispondente utilizzando la relazione contrasto-massa ottenuta dalla calibrazione MSPT (vedere paragrafo 7). Specificare la pendenza (pendenza) e l'intercetta y (offset) della linea di calibrazione, che mette in relazione il contrasto iSCAT con la massa molecolare (funzione trajectory_analysis.apply_calibration; vedere la sezione C.5 del notebook). Questa funzione aggiunge una colonna con la massa mediana della traiettoria a ciascun frame di dati.
- Valutare la densità apparente delle particelle sulla membrana con la funzione trajectory_analysis.membrane_density, che restituisce il valore di densità mediana in termini di particelle rilevate e traiettorie presenti durante ciascun fotogramma (vedere la sezione C.6 del notebook) come colonne aggiuntive nel frame di dati.
NOTA: poiché una parte delle particelle andrà persa durante il processo di rilevamento e di collegamento della traiettoria, le densità effettive delle particelle potrebbero essere più elevate. Per risultati affidabili per quanto riguarda le densità delle particelle e gli istogrammi di massa, ispezionare visivamente istantanee di filmati rappresentativi per verificare che le condizioni di misurazione siano plausibili per il tracciamento di singole particelle (vedere il passaggio 9.1.1).
10. Visualizzazione dei dati
- Illustrare la correlazione del coefficiente di massa e diffusione con la stima bidimensionale della densità del kernel (KDE), che si basa sul pacchetto Python fastkde (v.1.0.19; https://pypi.org/project/fastkde/).
- Per generare il plottaggio, specificare il file HDF5 contenente i risultati MSPT (vedere il passaggio 9.3.2 e la sezione notebook D.1) e selezionare un singolo (sezione notebook D.2) o un frame di dati concatenato (sezione notebook D.3) come dati di input per la funzione plotting.generate_2D_KDE (sezione notebook D.4).
NOTA: ogni set di dati tracciato dovrebbe contenere idealmente più di 1.000 traiettorie per un KDE 2D affidabile.
Risultati
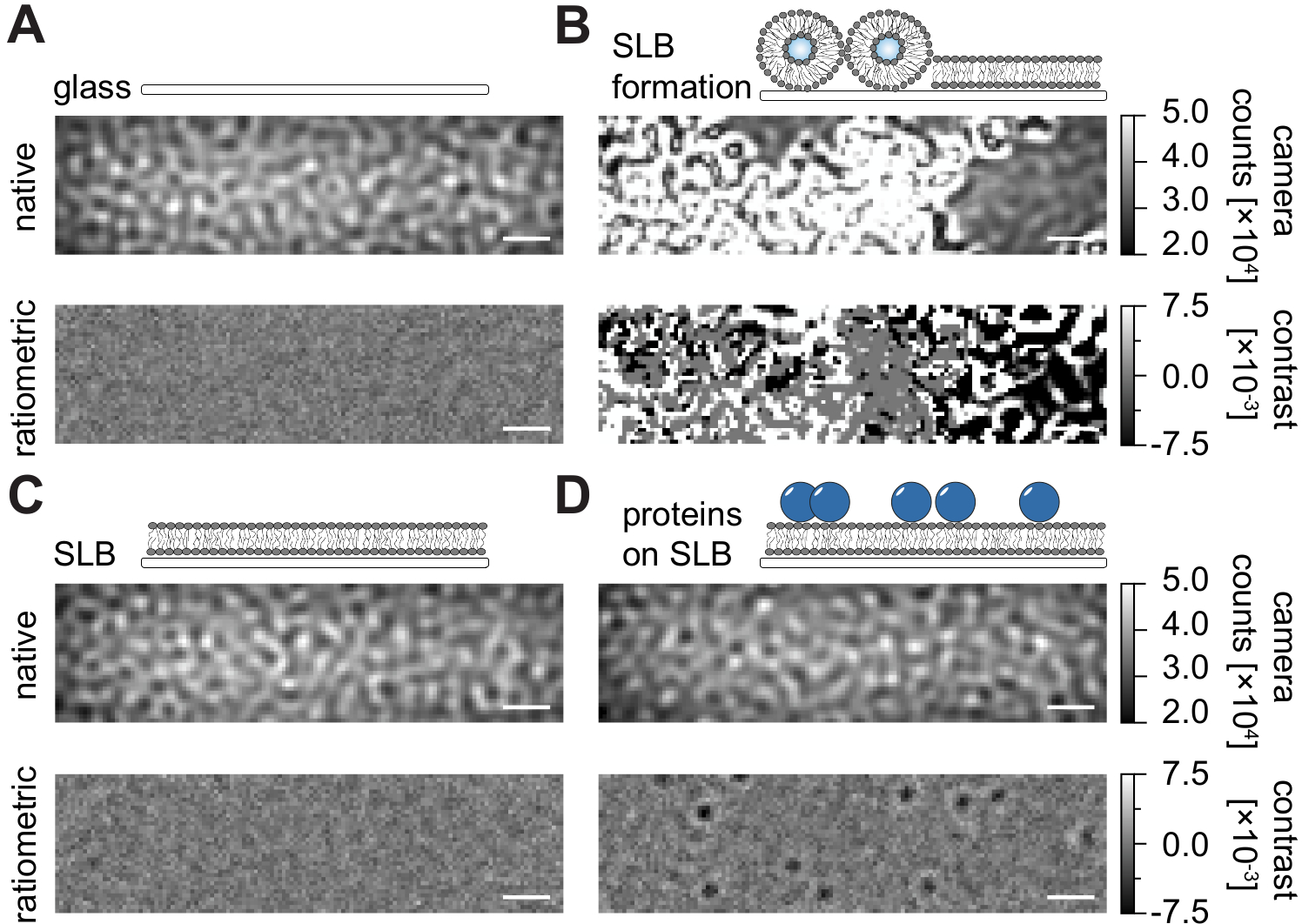
Seguendo il protocollo dettagliato qui riportato per la preparazione di doppi strati lipidici supportati (SLB) in camere di flusso (Figura 1), si può chiaramente riconoscere un modello simile a una macchia nella vista nativa di tutte le condizioni visualizzate (Figura 2). Questo effetto è causato dalla rugosità superficiale del vetro, che generalmente domina il segnale di dispersione e porta a condizioni visivamente indistinguibili (vetro, vetro con SLB o vetro con SLB e proteine attaccate). La presenza di vescicole, tuttavia, è chiaramente distinta a causa della grande sezione trasversale di dispersione delle vescicole e consente l'osservazione della rottura e della fusione delle vescicole in membrane omogenee (Figura 2B e Filmato supplementare 1). Quando si rimuove il segnale di scattering statico della superficie del vetro con un approccio raziometrico che enfatizza gli elementi dinamici all'interno del campo visivo 24,25, si possono scoprire proteine non etichettate che si diffondono sulla membrana (Figura 2D) mentre un SLB vuoto (Figura 2C) o il vetro stesso (Figura 2A) appare come un'immagine rumorosa.
Lo sfondo intrinseco delle misurazioni MSPT può essere stimato localmente dividendo ogni valore di pixel attraverso la mediana di n pixel precedenti e successivi del filmato nella stessa posizione dell'immagine (Figura 3). Di conseguenza, le macromolecole appaiono come funzioni isotrope a diffusione puntuale (PSF) il cui movimento sulla membrana può essere osservato, tracciato e quantificato. Infatti, la disponibilità sia del contrasto che del comportamento dinamico consente la relazione diretta della dimensione molecolare di una particella con il suo rispettivo comportamento diffusivo, il tutto senza la necessità di etichettare la particella. Tuttavia, per interpretare il contrasto iSCAT determinato durante gli esperimenti MSPT, è essenziale eseguire una calibrazione che traduca l'ampiezza del segnale in massa molecolare. Ciò può essere ottenuto collegando biomolecole di massa nota a un SLB tramite un complesso biotina-streptavidina-biotina (Figura 4A). Come strategia esemplare, si possono usare varianti biotinilate di albumina sierica bovina (BSA), proteina A (prA), fosfatasi alcalina (AP) e fibronectina (FN), che si legano alla streptavidina (STP) che a sua volta è legata ai lipidi contenenti biotina (Biotinyl Cap PE) nella membrana. Come mostrato nella Figura 4A, il contrasto sempre più pronunciato di queste macromolecole esemplari riflette il peso molecolare crescente dei rispettivi standard biotinilati. Assegnando ogni picco degli istogrammi di contrasto (Figura 4B) alla massa corrispondente dello stato oligomerico della proteina standard, viene rivelata una relazione lineare tra contrasto e massa21,22 e può essere successivamente utilizzata per l'analisi di sistemi macromolecolari sconosciuti (Figura 4C).
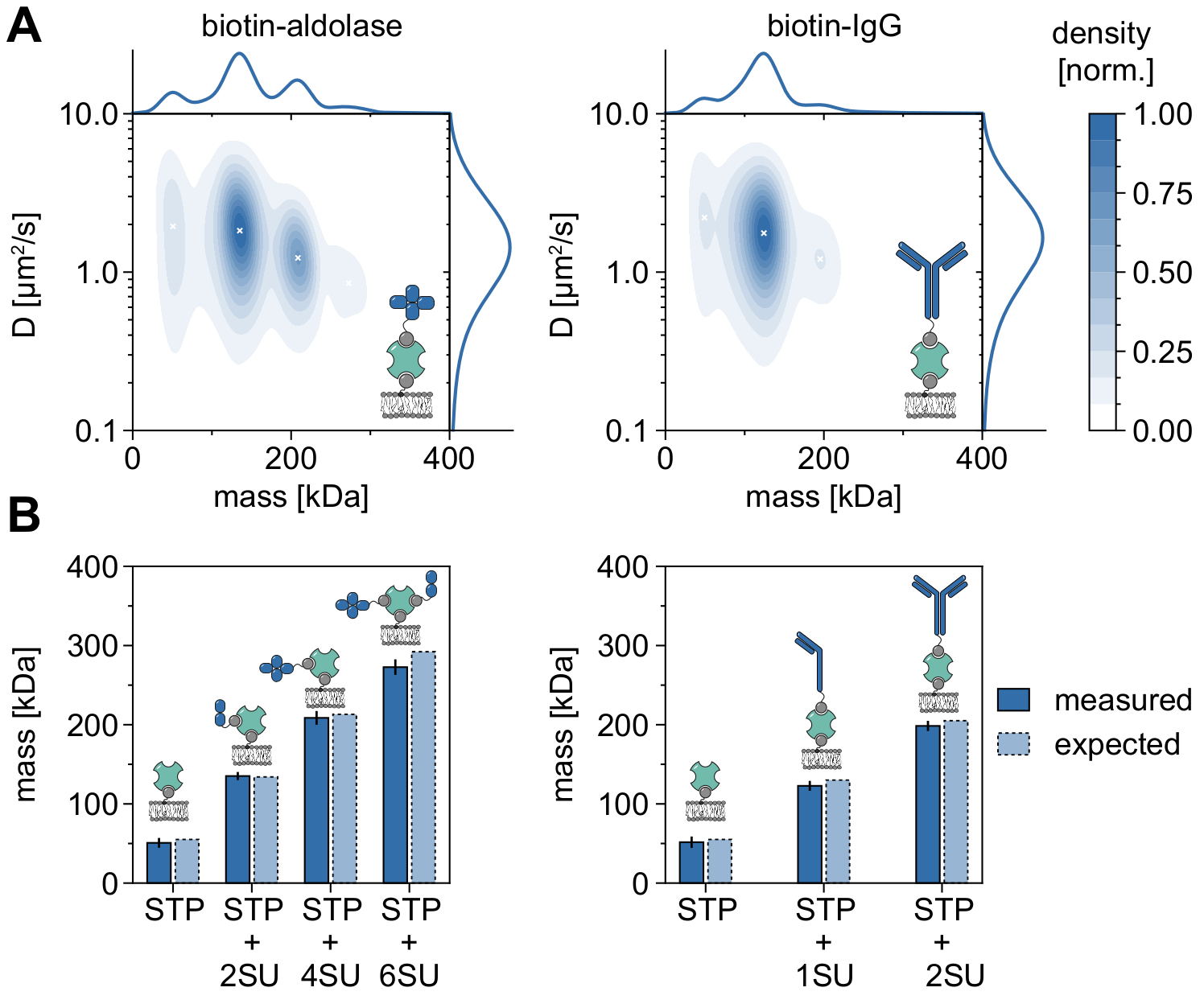
Un buon esempio che dimostra l'applicabilità e le capacità di MSPT per analizzare i pesi molecolari e quindi studiare gli stati oligomerici e gli eventi di oligomerizzazione è la considerazione dell'aldolasi biotinilata e delle IgG biotinilate (Figura 5). L'aldolasi è comunemente segnalata come un omotetramero32. Tuttavia, la distribuzione di massa risolta da MSPT presenta quattro picchi distinti, che evidenziano la presenza di più popolazioni (Figura 5A). Mentre il primo picco minore corrisponde alla streptavidina non occupata e può essere previsto a causa della configurazione in questo tipo di esperimento, possono essere rilevati anche complessi di aldolasi con solo due subunità (2SU) o sei subunità (6SU) (Figura 5B). È interessante notare che i complessi tetra- ed esamerici aldolasi-streptavidina mostrano un coefficiente di diffusione ridotto rispetto all'aldolasi dimerica e alla streptavidina da soli, indicando un aumento della resistenza viscosa, ad esempio, attraverso l'attaccamento di un secondo lipide biotinilato alla streptavidina. Allo stesso modo, le IgG biotinilate mostrano tre picchi nella distribuzione di massa, con il primo picco che corrisponde di nuovo alla massa di una singola streptavidina. La massa del picco più abbondante corrisponde alla massa di una catena leggera e una pesante (1SU), cioè la metà di un anticorpo IgG. L'anticorpo completo con due metà identiche (2SU) viene rilevato in circa l'11% dei casi. La diminuzione del coefficiente di diffusione con dimensioni complesse crescenti indica interazioni della streptavidina con più di un lipide biotinilato o resistenza aggiuntiva causata dalle IgG attaccate, o entrambe.
Oltre alla sola analisi degli stati oligomerici dipendenti dalla membrana, MSPT conferisce anche il particolare vantaggio di correlare il comportamento diffusivo di una macromolecola di interesse con il suo stato oligomerico. I risultati rappresentativi per questo tipo di analisi sono mostrati per l'annessina V (AnV) e la subunità B della tossina del colera (CTxB), che si legano rispettivamente alla dioleoilfosfatidilserina (DOPS) o ai glicosfingolipidi (GM1), incorporati nella membrana (Figura 6A). Entrambe le stime della densità del kernel (KDE) presentano distribuzioni unimodali di massa e diffusione, indicando una singola specie abbondante con comportamento diffusivo simile. La posizione di picco della massa molecolare e del coefficiente di diffusione è risultata essere rispettivamente di 49,8 ± 2,2 kDa e 1,4 ± 0,1 μm2/s per AnV e 62,7 ± 3,1 kDa e 0,4 ± 0,1 μm2/s, rispettivamente, per CTxB. I coefficienti di diffusione misurati sono paragonabili ai valori precedentemente riportati ottenuti da AFM ad alta velocità e FRAP33,34. La massa leggermente ridotta rispetto alla massa della macromolecola prevista (52 kDa per un trimero AnV, 65 kDa per un pentamero CTxB) può indicare la presenza di complessi più piccoli con meno subunità nell'insieme. Mentre la differenza di massa tra le proteine è piccola e vicina al limite di rilevamento specificato del microscopio (≈50 kDa), i loro coefficienti di diffusione differiscono considerevolmente. In una miscela equimolare, ad esempio, confrontando la diffusione della miscela con la distribuzione di AnV e CTxB da sola, si può concludere che AnV è più abbondante sulla membrana rispetto a CTxB (Figura 6B). Tuttavia, se la concentrazione di CTxB è raddoppiata rispetto alla concentrazione di AnV, l'equilibrio viene spostato verso CTxB come proteina predominante sulla membrana. Come illustrato per le miscele di AnV e CTxB, MSPT non solo consente di discriminare le macromolecole associate alla membrana in base al loro peso molecolare, ma consente anche la discriminazione di diverse popolazioni di macromolecole in base al loro comportamento diffusivo.
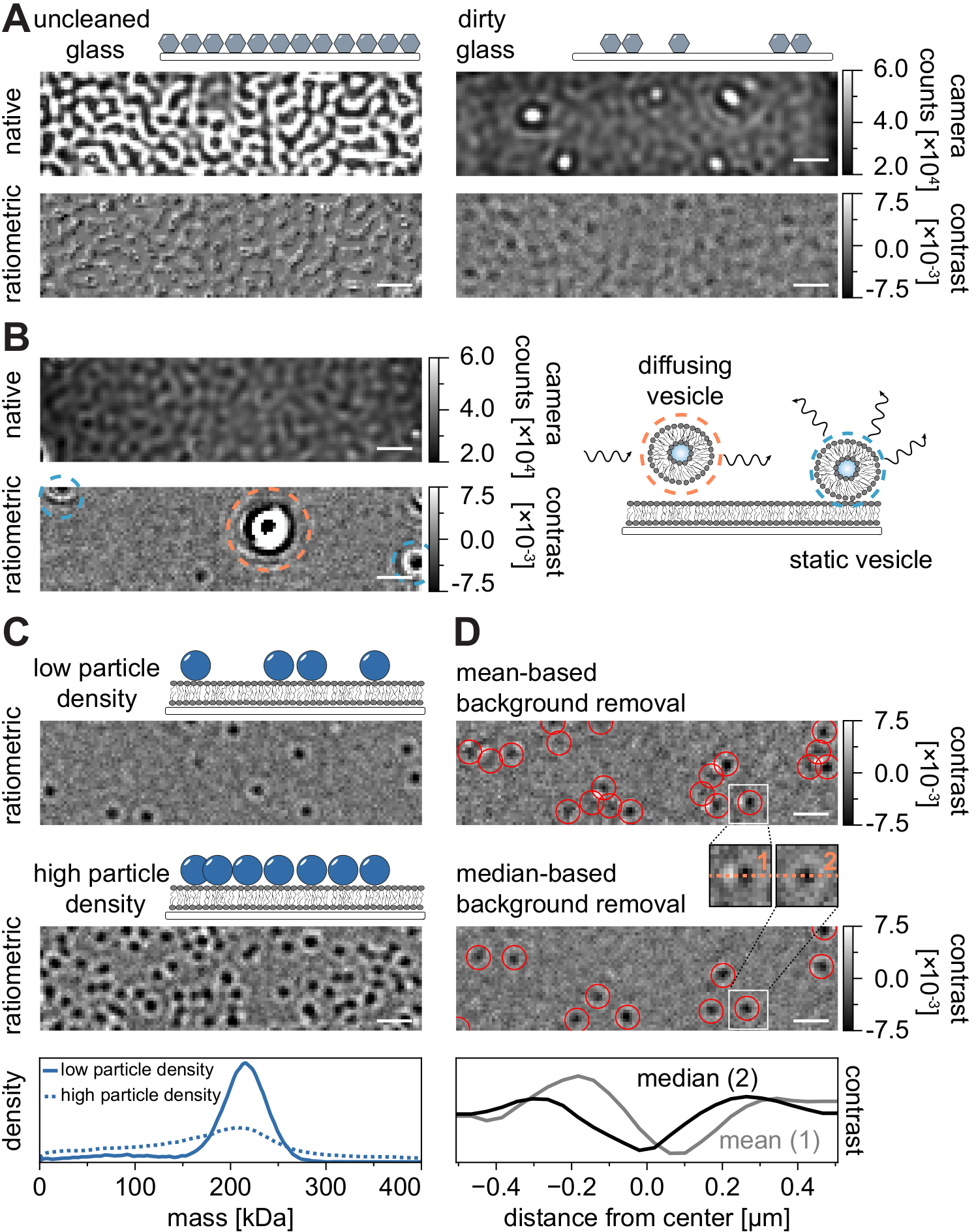
Come per tutte le tecniche di microscopia, alcuni requisiti sperimentali sono cruciali per ottenere la qualità desiderata dei dati. Un esempio importante in questo contesto sono le coperture accuratamente pulite. In generale, questo è considerato un prerequisito per gli esperimenti a singola molecola correlati alla microscopia, ma MSPT è particolarmente sensibile alle impurità del campione. L'aumento della dispersione proveniente dalla superficie vetrata delle coperture non pulite impedisce qualsiasi misurazione quantitativa dell'iSCAT. In particolare, anche lo sporco residuo o le particelle di polvere su vetro non sufficientemente pulito possono causare notevoli distorsioni dell'immagine, riconoscibili come punti luminosi nella modalità di imaging nativo (Figura 7A). Sebbene questi difetti siano rimossi dalla stima di fondo a causa della loro natura statica, la determinazione accurata del contrasto di una particella potrebbe essere compromessa e quindi influenzare negativamente la sua analisi quantitativa. Un altro problema comune riscontrato negli esperimenti MSPT sono le vescicole rimanenti che galleggiano (circondate in arancione) attraverso il campo visivo o vescicole non fuse che sono bloccate (circondate in blu) in una posizione specifica sulla membrana e appaiono come grandi scatterer pulsanti (Figura 7B). Per ridurre al minimo la loro presenza e interferenza con l'acquisizione del film, si consiglia di lavare accuratamente l'SLB prima di aggiungere la proteina e di utilizzare miscele appena preparate di piccole vescicole unilamellari (SUV) e cationi bivalenti.
Un fattore che deve essere preso in considerazione per la progettazione di esperimenti di tracciamento delle particelle sensibili alla massa è la densità delle macromolecole associate all'interfaccia di membrana. Elevate densità di particelle sulla membrana possono infatti causare problemi per due motivi: i) Il collegamento di rilevamenti di particelle da fotogrammi consecutivi a traiettorie diventa ambiguo e quindi aumenta la probabilità di errori e coefficienti di diffusione mal valutati. ii) La massa delle particelle, che viene estratta dall'ampiezza del loro corrispondente adattamento delle FPF, viene sistematicamente sottovalutata e i picchi di massa si allargano perché la separazione del segnale di fondo statico dal segnale dinamico delle particelle è sempre più difficile (Figura 7C). Attualmente, la valutazione visiva della qualità dei dati nel processo di acquisizione di video MSPT è difficile sui microscopi commerciali disponibili perché la vista raziometrica implementata nel software di acquisizione utilizza la rimozione dello sfondo stabilita per la fotometria di massa21 invece dell'algoritmo basato sulla mediana descritto qui e nei riferimenti 24,25 (Figura 7D ). La rimozione continua dello sfondo basata sulla media utilizzata per visualizzare le molecole di atterraggio nella fotometria di massa fa sì che le particelle diffuse appaiano come fronti scuri con code luminose, il che rende le macchie altamente anisotrope e interferisce con il montaggio della PSF durante la procedura di rilevamento. Pertanto, l'uso dell'elaborazione delle immagini basata sulla media implementata nel software di acquisizione non è adatto per l'analisi di biomolecole diffuse sulle membrane.

Figura 1: Diagramma di flusso del processo delle singole fasi necessarie per analizzare le interazioni proteina-membrana con il tracciamento delle particelle sensibili alla massa (MSPT). Per preparare i campioni per le misurazioni MSPT, i vetrini di copertura in vetro devono essere accuratamente puliti e attivati con un plasma di ossigeno. Dopo il loro assemblaggio in camere di flusso del campione, piccole vescicole unilamellari (SUV) vengono preparate per la formazione di doppio strato lipidico supportato (SLB) e tutti i tamponi di reazione vengono filtrati per ridurre la dispersione di fondo. I SUV vengono aggiunti per formare doppi strati lipidici nelle camere di flusso. Facoltativamente, i cationi bivalenti come gli ioni Ca2+ possono essere aggiunti ai SUV per promuovere la rottura delle vescicole. Infine, basse concentrazioni della proteina di interesse vengono scaricate nella camera di reazione. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Vista nativa e raziometrica di superfici esemplari rilevanti per le misurazioni MSPT. Immagini rappresentative della rugosità superficiale di un vetrino (A), durante la formazione di un doppio strato lipidico supportato (B), con un doppio strato lipidico supportato intatto (C) e di proteine esemplari ricostituite su un SLB (D). Tutti e quattro gli esempi vengono visualizzati in modalità nativa, a cui è possibile accedere durante la misurazione stessa, e come immagini ratiometriche elaborate dopo la rimozione dello sfondo basato sulla mediana. Le barre della scala rappresentano 1 μm. Per l'analisi dei dati (vedere il quaderno Jupyter accompagnato; passaggio 9), sono stati utilizzati i seguenti parametri: dimensione mediana della finestra (window_length) = 1001. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

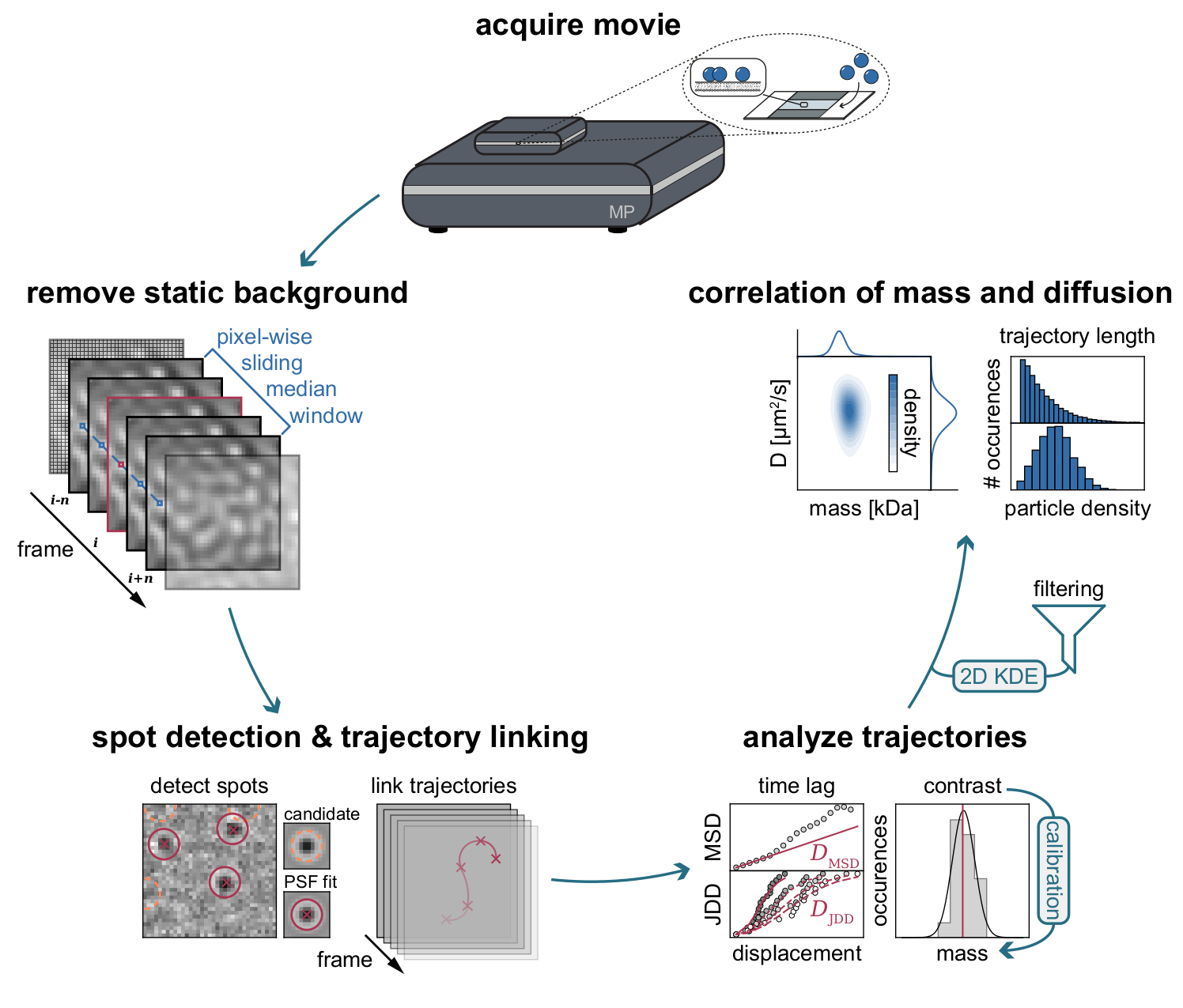
Figura 3: Diagramma dettagliato delle fasi necessarie per la raccolta e l'analisi dei dati MSPT. Dopo l'acquisizione dei dati per il campione di interesse sul fotometro di massa, i filmati vengono elaborati per rimuovere lo sfondo statico attraverso un approccio mediano scorrevole in termini di pixel. Successivamente, le particelle candidate vengono identificate e adattate da una funzione di diffusione puntuale (PSF) prima del loro collegamento nelle traiettorie delle particelle. Per consentire la determinazione del coefficiente di diffusione per ciascuna particella, viene utilizzata l'analisi dello spostamento quadrato medio (MSD) o della distribuzione della distanza di salto (JDD). In questa fase, i valori di contrasto possono essere trasformati in masse molecolari in base alla relazione di contrasto-massa determinata attraverso la strategia di calibrazione. Come passo finale, le traiettorie possono essere filtrate in base alla loro lunghezza o densità di particelle di membrana e visualizzate mediante stima bidimensionale della densità del kernel (2D-KDE). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Calibrazione della relazione massa-contrasto per le misurazioni MSPT. (A) Quadri raziometrici rappresentativi ottenuti per complessi proteici esemplari standard streptavidina che si diffondono su un doppio strato lipidico supportato contenente una piccola percentuale di lipidi biotinilati (rapporto DOPC:DOPG:Biotinyl Cap PE di 70:29.99:0.01 mol%). Come standard di peso molecolare modello, sono mostrati streptavidina28 monovalente (solo STP) o streptavidina28 bivalente in complesso con albumina sierica bovina biotinilata (BSA), proteina A biotinilata (prA), fosfatasi alcalina biotinilata (AP) o fibronectina biotinilata (FN). I punti candidati sono evidenziati in arancione (cerchi tratteggiati) e i rilevamenti di particelle di successo sono evidenziati in rosso (cerchi solidi). Le barre di scala rappresentano 1 μm. (B) Distribuzioni di densità di probabilità dei valori di contrasto ottenuti per le cinque proteine standard modello. Tutti i dati visualizzati rappresentano distribuzioni raggruppate di tre esperimenti indipendenti per condizione: STP solo n = 82.719; BSA n = 9.034; prA n = 22.204; AP n = 69.065 e FN n = 71.759 traiettorie. Rispetto al numero di particelle determinato per le membrane con proteine, il numero di particelle rilevate su un doppio strato vuoto è trascurabile a densità di membrana moderate (Figura supplementare 1). I picchi di contrasto considerati per la calibrazione della massa sono contrassegnati attraverso linee continue mentre quelli tratteggiati rappresentano stati oligomerici non considerati. (C) Curva di calibrazione contrasto-massa derivata dai contrasti di picco nel pannello D e dalle rispettive masse di sequenza dei complessi. Le barre di errore visualizzano l'errore standard delle posizioni di picco stimate mediante bootstrap (100 ricampioni con 1.000 traiettorie ciascuna). Per l'analisi dei dati (vedere Notebook Jupyter; passaggio 9), sono stati utilizzati i seguenti parametri: dimensione mediana della finestra (window_length) = 1.001 fotogrammi, soglia di rilevamento (thresh) = 0,00055, intervallo di ricerca (dmax) = 4 pixel, memoria (max_frames_to_vanish) = 0 fotogrammi, lunghezza minima della traiettoria (minimum_trajectory_length) = 7 fotogrammi (solo STP), 9 fotogrammi (BSA/FN), 15 fotogrammi (prA), 10 fotogrammi (AP). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Decifrare gli stati oligomerici delle proteine associate alla membrana. (A) Stime 2D della densità del kernel sia del coefficiente di massa che di diffusione della streptavidina tetravalente in complesso con aldolasi biotinilata (pannello sinistro) o con un anticorpo anti-Rabbit IgG di capra modificato con biotina (pannello destro). La ricostituzione di entrambi i complessi è stata eseguita su un doppio strato lipidico supportato contenente DOPC, DOPG e Biotinyl Cap PE in un rapporto di 70:29.99:0.01 mol%, rispettivamente. In totale, sono state incluse 116.787 traiettorie di tre repliche indipendenti per il complesso streptavidina-aldolasi (densità delle particelle di 0,1 μm-2) e 348.405 per il composito streptavidina-IgG (densità delle particelle di 0,1 μm-2). Sono state incluse solo particelle con una lunghezza della traccia di almeno cinque fotogrammi. Vengono presentate distribuzioni di probabilità marginali sia della massa molecolare (in alto) che del coefficiente di diffusione (a destra). La x nera in entrambi i pannelli segna i rispettivi massimi locali del KDE. (B) Confronto di masse oligomeriche determinate per il complesso di streptavidina tetravalente con aldolasi modificata da biotina (pannello di sinistra) o IgG biotinilato (pannello di destra) con, secondo le masse della sequenza, pesi molecolari attesi. L'abbreviazione SU viene introdotta per conto della subunità della proteina di interesse. Le barre di errore visualizzano l'errore standard delle posizioni di picco stimate mediante bootstrap (100 ricampioni con 1.000 traiettorie ciascuna). Per l'analisi dei dati (vedere il notebook Jupyter accompagnato; passaggio 9), sono stati utilizzati i seguenti parametri: dimensione mediana della finestra (window_length) = 1.001 fotogrammi, soglia di rilevamento (trebbiatura) = 0,00055, intervallo di ricerca (dmax) = 4 pixel, memoria (max_frames_to_vanish) = 0 fotogrammi, lunghezza minima della traiettoria (minimum_trajectory_length) = 5. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 6: Scioglimento del comportamento diffusivo delle proteine native che interagiscono con la membrana annessina V (AnV) e della subunità B (CTxB) della tossina del colera. (A) Stime 2D della densità del kernel sia del coefficiente di massa che di diffusione dell'annessina V (pannello sinistro) e della subunità B della tossina del colera (pannello destro). Per la ricostituzione della membrana AnV e CTxB, sono state utilizzate composizioni lipidiche di 80:20 mol% DOPC a DOPS e 99,99:0,01 mol% DOPC a GM1, rispettivamente. In totale sono state incluse 206.819 traiettorie di tre repliche indipendenti per AnV (densità delle particelle di 0,1 μm-2) e 142.895 traiettorie per CTxB (densità delle particelle di 0,2 μm-2). (B) Stime 2D della densità del kernel delle miscele CTxB e AnV in un rapporto di 1:1 (pannello di sinistra) o 2:1 (pannello di destra), rispettivamente. La ricostituzione di miscele proteiche è stata eseguita su un doppio strato lipidico supportato contenente lipidi DOPC, DOPS e GM1 in un rapporto di 80:19,99:0,01 mol%. In totale sono state incluse 42.696 traiettorie di tre repliche indipendenti per la miscela 1:1 (densità delle particelle di 0,1 μm-2) e 264.561 traiettorie per il rapporto 2:1 (densità delle particelle di 0,3 μm-2). Sia per (A) che per (B), sono state incluse solo particelle con una lunghezza della traccia di almeno cinque fotogrammi. Vengono presentate distribuzioni di probabilità marginali sia della massa molecolare (in alto) che del coefficiente di diffusione (a destra). La x bianca in ogni pannello segna il rispettivo massimo globale del KDE. Per l'analisi dei dati (vedere il notebook Jupyter accompagnato; passaggio 9), sono stati utilizzati i seguenti parametri: dimensione mediana della finestra (window_length) = 1.001 fotogrammi, soglia di rilevamento (trebbiatura) = 0,00055, intervallo di ricerca (dmax) = 4 pixel, memoria (max_frames_to_vanish) = 0 fotogrammi, lunghezza minima della traiettoria (minimum_trajectory_length) = 5. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 7: Potenziali complicazioni nel corso delle misurazioni MSPT o durante l'analisi dei dati. (A) Immagini rappresentative della rugosità superficiale visualizzate sia nella vista raziometrica nativa che in quella elaborata (rimozione dello sfondo basata sulla mediana) di una diapositiva di vetro di copertura non pulita. In entrambi i casi, i punti luminosi costituiscono impurità superficiali residue che impediscono misurazioni prive di artefatti. (B) Immagini esemplari di vescicole residue nel campo visivo dopo un insufficiente lavaggio della membrana. Sia le vescicole statiche (evidenziate in blu) che quelle diffuse (evidenziate in arancione) comprometteranno la qualità della misurazione a causa rispettivamente della pulsazione e della deformazione o del loro movimento direzionale. (C) Come tecnica a singola particella, MSPT richiede basse densità di particelle (immagine rappresentativa, pannello superiore) per consentire un corretto collegamento e determinazione della massa di ciascuna particella. Nel caso di alte densità di particelle di membrana (pannello centrale), il montaggio delle particelle è compromesso, il che influisce sulla determinazione della massa (vedi pannello inferiore). (D) Immagini raziometriche rappresentative di particelle che si diffondono su un'interfaccia di membrana dopo la rimozione dello sfondo a base media (pannello superiore) o mediana. Per la diffusione delle particelle, la strategia di rimozione dello sfondo basata sulla media produce immagini distorte della PSF della particella, come si può vedere nei piccoli inserti tra il pannello superiore e quello centrale. Al contrario, le PSF di particelle non distorte possono essere ottenute attraverso l'approccio basato sulla mediana. Pannello inferiore: confronto dei profili di linea attraverso il centro della FPF ottenuto dopo la rimozione dello sfondo basato sulla media o sulla mediana. Per tutte le immagini native e raziometriche visualizzate in questa figura, le barre della scala rappresentano 1 μm. Per l'analisi dei dati (vedere il notebook Jupyter accompagnato; passaggio 9), sono stati utilizzati i seguenti parametri: dimensione mediana della finestra (window_length) = 1.001 fotogrammi, soglia di rilevamento (trebbiatura) = 0,00055, intervallo di ricerca (dmax) = 4 pixel, memoria (max_frames_to_vanish) = 0 fotogrammi, lunghezza minima della traiettoria (minimum_trajectory_length) = 5. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Figura supplementare 1: Confronto tra membrane prive di proteine e occupate. Immagini rappresentative di un doppio strato lipidico supportato intatto prima (A) e dopo (B) l'aggiunta di streptavidina purificata (STP). I punti candidati che sono stati adattati con successo al modello PSF sono circondati in rosso. (C) Distribuzioni di probabilità di contrasto di particelle rilevate su una membrana vuota (fondo membrana, grigio) e su un doppio strato con particelle di streptavidina diffusive (blu). Entrambe le distribuzioni di probabilità rappresentano i dati aggregati di tre esperimenti indipendenti con identici parametri di acquisizione e analisi dei filmati. Per l'analisi dei dati (vedere il notebook Jupyter accompagnato; passaggio 9), sono stati utilizzati i seguenti parametri: dimensione mediana della finestra (window_length) = 1.001 fotogrammi, soglia di rilevamento (trebbiatura) = 0,00055, intervallo di ricerca (dmax) = 4 pixel, memoria (max_frames_to_vanish) = 0 fotogrammi, lunghezza minima della traiettoria (minimum_trajectory_length) = 7 fotogrammi. Fare clic qui per scaricare questo file.
Filmato supplementare 1: Film esemplare che mostra la rottura e la fusione delle vescicole in una membrana omogenea registrata con il fotometro di massa. Dimensione mediana della finestra di elaborazione dell'immagine (window_length) = 1.001 fotogrammi. Barra della scala: 1 μm. Gamma di conteggi della fotocamera: nero = 16.892; bianco = 65.408. Clicca qui per scaricare questo film.
Filmato supplementare 2: Filmati esemplari che mostrano la diffusione di complessi di annessina V (in alto) e aldolasi biotinilata (in basso) su un doppio strato come ottenuto dalle misurazioni MSPT. Dimensione mediana della finestra di elaborazione dell'immagine (window_length) = 1.001 fotogrammi. Barra della scala: 1 μm. Intervallo di contrasto di scattering interferometrico: nero = -0,0075; bianco = 0,0075. Clicca qui per scaricare questo film.
Discussione
Il protocollo presentato estende la fotometria di massa21, una tecnica che analizza la massa di singole biomolecole adsorbenti su vetro, a uno strumento ancora più versatile in grado di misurare contemporaneamente la massa e la diffusione di biomolecole che interagiscono con membrana non etichettate. Questa estensione dell'analisi è ottenuta attraverso l'implementazione di una strategia di rimozione dello sfondo modificata adattata al movimento laterale delle molecole24,25. In generale, la rimozione dello sfondo è della massima importanza per gli approcci basati su iSCAT, poiché la forte dispersione della rugosità superficiale del vetro rappresenta il principale ostacolo all'analisi e la determinazione accurata dello sfondo locale di ciascun pixel è essenziale per la quantificazione della massa e della posizione delle particelle. Oltre all'analisi delle immagini adattata al movimento delle particelle, il successivo rilevamento delle particelle, il collegamento della traiettoria e l'analisi dei dati completano la nuova espansione di MP nel tracciamento delle particelle sensibili alla massa (MSPT).
In generale, vetrine accuratamente pulite e un ambiente di lavoro pulito sono requisiti critici per il successo degli esperimenti MSPT. A causa dell'assenza di marcatura delle macromolecole, il segnale acquisito è intrinsecamente non selettivo. Campioni puliti, così come una corretta gestione dei campioni, sono quindi fondamentali per garantire che le osservazioni non possano essere interpretate erroneamente. In particolare, quando vengono esaminate molecole a basso peso molecolare, vengono approvate misure di controllo di membrane prive di proteine per valutare i contributi di fondo (Figura supplementare 1). Oltre all'inclusione delle misure di controllo, si raccomanda quindi di seguire le fasi di preparazione visualizzate nella Figura 2 per ciascuna camera di flusso. Se combinate, queste misure di sicurezza garantiranno che il segnale rilevato provenga dalla biomolecola di interesse e non, ad esempio, da una camera di flusso, un tampone o una membrana contaminati.
Oltre alle precauzioni relative alla progettazione sperimentale, è necessario prestare attenzione anche durante l'elaborazione delle immagini MSPT. Durante l'elaborazione video, il valore di tre parametri deve essere scelto con attenzione per garantire risultati corretti: i) la lunghezza della finestra mediana per la rimozione dello sfondo, ii) la soglia per il rilevamento delle particelle e iii) il raggio di ricerca massimo durante l'assegnazione del collegamento. Una finestra mediana più grande (i) generalmente facilita la separazione delle particelle diffuse dallo sfondo quasi-costante sovrapposto. Tuttavia, per finestre di dimensioni troppo grandi la deriva del campione alla fine diventerà evidente e diminuirà l'accuratezza della stima dello sfondo. Le impostazioni ottimali dipendono fortemente dalle proprietà del campione e dalle condizioni di misurazione. Tuttavia, un valore di 1.001 può essere utilizzato come punto di partenza robusto. Il parametro di soglia (ii) deve essere regolato in base alla massa molecolare più bassa prevista nel campione. Un valore inferiore a 0,0005 non è raccomandato per le misurazioni effettuate con il fotometro di massa utilizzato in questo studio. Per velocizzare i tempi di analisi, è possibile scegliere valori più elevati se si prevede un campione ad alto peso molecolare. Il raggio di ricerca nel collegamento della traiettoria (iii) specifica la distanza radiale massima in pixel in cui verrà ricercata la posizione spostata della particella in fotogrammi consecutivi. Dovrebbe essere adattato alla particella più veloce del campione e, se favorito, un intervallo di ricerca adattivo (vedi documentazione di trackpy) potrebbe essere utilizzato invece per ridurre il tempo di calcolo. Soprattutto durante la fase iniziale di un progetto, si consiglia di rianalizzare i filmati con parametri variabili per convalidare i risultati ottenuti.
Alla luce della natura a singola molecola di MSPT, si dovrebbe evitare di misurare ad alte densità di particelle di membrana in quanto possono interferire con un contrasto accurato e la determinazione della massa. È stato dimostrato che le densità inferiori a una particella per micrometro quadrato sono favorevoli per le misurazioni MSPT24. Un'ulteriore considerazione sono i coefficienti di diffusione attesi nel campione. Sebbene applicabile a una vasta gamma di coefficienti di diffusione, MSPT ha un limite inferiore di coefficienti di diffusione accessibili. Il confinamento locale in una regione di pochi pixel durante una porzione significativa del periodo della finestra mediana fonde la particella con lo sfondo statico. Per le condizioni di imaging utilizzate in questo protocollo, la misurazione di coefficienti di diffusione inferiori a 0,01 μm2/s non è raccomandata. A questa velocità di diffusione, ad esempio, lo spostamento quadrato medio di una particella durante la semidimensione della finestra mediana è di circa 4 pixel e quindi di dimensioni simili all'estensione della FPF. Di conseguenza, è probabile che la stima statica di fondo contenga contributi di segnale dalla particella stessa, il che si traduce in un contrasto apparentemente ridotto della particella fino a quando alla fine non si avvicina al livello di rumore. Tuttavia, i coefficienti di diffusione delle macromolecole compresi tra 0,05 e 10 μm2/s possono essere chiaramente risolti.
Per estendere ulteriormente la gamma di applicazioni MSPT, si può immaginare un avanzamento dell'algoritmo di sfondo basato sulla mediana attraverso l'eliminazione dei pixel temporaneamente occupati da una particella o mediante la correzione della deriva del campione che consente finestre di dimensioni mediane più grandi. Entrambi gli approcci allevierebbero i problemi relativi alle misurazioni ad alta densità di particelle e alla lenta diffusione. Miglioramenti in termini di minore sensibilità di massa sono all'orizzonte con una nuova generazione di fotometri di massa, che possono fornire l'accesso a biomolecole inferiori a 50 kDa. Pertanto, i futuri esperimenti MSPT saranno in grado di studiare la dinamica a singola molecola e le interazioni correlate alla membrana per una gamma ancora più ampia di mimetismo di membrana come doppi strati ammortizzati e sistemi macromolecolari.
Divulgazioni
Gli autori non hanno conflitti di interesse.
Riconoscimenti
Apprezziamo sinceramente il supporto di Philipp Kukura, Gavin Young e del team software Refeyn e riconosciamo la loro assistenza condividendo parti del codice di analisi delle immagini. Ringraziamo la Cryo-EM MPIB Core Facility per aver fornito l'accesso al fotometro di massa Commerciale Refeyn. F.S. riconosce con gratitudine il sostegno e i finanziamenti concessi da Jürgen Plitzko e Wolfgang Baumeister. T.H. e P.S. hanno ricevuto finanziamenti attraverso la Deutsche Forschungsgemeinschaft (DFG, Fondazione tedesca per la ricerca) - Project-ID 201269156 - SFB 1032 (A09). N.H. è stato supportato da una sovvenzione di ritorno DFG HU 2462/3-1. P.S. riconosce il sostegno attraverso la rete di ricerca MaxSynBio attraverso l'iniziativa di finanziamento congiunto del Ministero federale tedesco dell'Istruzione e della Ricerca (BMBF) e della Max Planck Society.
Materiali
| Name | Company | Catalog Number | Comments |
| annexin V | Sigma Aldrich | #SRP8026 | examplary membrane-interacting protein |
| Bio-Rad Protein Assay | Bio-Rad Laboratories Inc. | #5000006 | bradford assay kit to determine protein stock concentrations |
| biotin labeled bovine albumin | Sigma Aldrich | #A8549 | examplary protein that can be used as standard protein for MSPT |
| cholera toxin subunit B | Sigma Aldrich | #SAE0069 | examplary membrane-interacting protein |
| cover glasses, #1.5, 24 x 24 mm | Paul Marienfeld GmbH & Co. KG | #0102062 | |
| cover glasses, #1.5, 24 x 60 mm | Paul Marienfeld GmbH & Co. KG | #0102242 | |
| dioleoyl-sn-glycero-3-phosphocholine (DOPC) | Avanti Polar Lipids | #850375 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| dioleoyl-sn-glycero-3-phosphoethanolamine-N-cap biotinyl (18:1 Biotinyl Cap PE | Avanti Polar Lipids | #870273 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| dioleoyl-sn-glycero-3-phosphoglycerol (DOPG) | Avanti Polar Lipids | #840475 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| dioleoyl-sn-glycero-3-phospho-L-serine (DOPS) | Avanti Polar Lipids | #840035 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| double-sided tape | tesa | #57912-00000-02 | needed for the assembly of glass sample chambers |
| Extruder | Avanti Polar Lipids | #610023 | Lipid extruder to enable monodisperse vesicle distributions |
| EZ-Link Maleimide-PEG2-Biotin | Thermo Fisher Scientific | #A39261 | maileimide-fused biotin that can be used to biotinylate standard proteins for MSPT |
| Fibronectin (Biotinylated) | Cytoskeleton Inc. | #FNR03-A | examplary protein that can be used as standard protein for MSPT |
| Gel Filtration HMW Calibration Kit | Cytiva | #28403842 | standard proteins, e.g. aldolase that can be biotinylated and used as molecular weight standards for MSPT |
| GM1 Ganglioside (Brain, Ovine-Sodium Salt) | Avanti Polar Lipids | #860065 | lipid - in the form of extruded small unilamellar vesicles required for supported lipid bilayer formation |
| Goat anti-Rabbit IgG (H+L) Secondary Antibody, Biotin | Thermo Fisher Scientific, Waltham, USA) | #31820 | examplary protein to highlight the existence of different protein states |
| Isopropanol, 99.5%, for spectroscopy | Thermo Fisher Scientific | #10003643 | |
| Low Autofluorescence Immersion Oil | Olympus K.K. | #IMMOIL-F30CC | |
| pET21a-Streptavidin-Alive | Addgene | #20860 | required to express and purify divalent streptavidin in combination with each other |
| pET21a-Streptavidin-Dead | Addgene | #20859 | required to express and purify divalent streptavidin in combination with each other |
| Pierce Alkaline Phosphatase, biotinylated | Thermo Fisher Scientific | #29339 | examplary protein that can be used as standard protein for MSPT |
| Pierce Protein A, Biotinylated | Thermo Fisher Scientific | #29989 | examplary protein that can be used as standard protein for MSPT |
| Refeyn Acquire | Refeyn Ltd. | control software for Refeyn OneMP | |
| Refeyn One | Refeyn Ltd. | - | mass photometer |
| sterile syringe filters 0.45 µm cellulose acetate membrane | VWR International | #514-0063 | needed to filter particles from the buffer of interest |
| tetravalent streptavidin | Thermo Fisher Scientific | #SNN1001 | tetravalent streptavidin to enable the presence of several biotin binding sites |
| Whatman Nuclepore Hydrophilic Membrane, 0.05 µm Pore Size, 25 mm Circle | Cytiva | #110603 | a pore size of 50 nm is recommended for supported lipid bilayer formation in the context of MSPT |
| Zepto model 2 plasma cleaner | Diener electronic GmbH | - |
Riferimenti
- Robertson, J. L. The lipid bilayer membrane and its protein constituents. Journal of General Physiology. 150 (11), 1472-1483 (2018).
- Grecco, H. E., Schmick, M., Bastiaens, P. I. H. Signaling from the Living Plasma Membrane. Cell. 144 (6), 897-909 (2011).
- Cho, W., Stahelin, R. V. Membrane-protein interactions in cell signaling and membrane trafficking. Annual Review of Biophysics and Biomolecular Structure. 34, 119-151 (2005).
- Whited, A. M., Johs, A. The interactions of peripheral membrane proteins with biological membranes. Chemistry and Physics of Lipids. 192, 51-59 (2015).
- Gonzalez, L., Scheller, R. H. Regulation of membrane trafficking: Structural insights from a Rab/effector complex. Cell. 96 (6), 755-758 (1999).
- Sezgin, E., Levental, I., Mayor, S., Eggeling, C. The mystery of membrane organization: composition, regulation and roles of lipid rafts. Nature Reviews Molecular Cell Biology. 18 (6), 361-374 (2017).
- Bagheri, Y., Ali, A. A., You, M. Current methods for detecting cell membrane transient interactions. Frontiers in Chemistry. 8, 603259 (2020).
- Miller, H., Zhou, Z., Shepherd, J., Wollman, A. J. M., Leake, M. C. Single-molecule techniques in biophysics: A review of the progress in methods and applications. Reports on Progress in Physics. 81 (2), 024601 (2018).
- Manzo, C., Garcia-Parajo, M. F. A review of progress in single particle tracking: From methods to biophysical insights. Reports on Progress in Physics. 78 (12), 124601 (2015).
- Gelles, J., Schnapp, B. J., Sheetz, M. P. Tracking kinesin-driven movements with nanometre-scale precision. Nature. 331 (6155), 450-453 (1988).
- Funatsu, T., Harada, Y., Tokunaga, M., Saito, K., Yanagida, T. Imaging of single fluorescent molecules and individual ATP turnovers by single myosin molecules in aqueous solution. Nature. 374 (6522), 555-559 (1995).
- Schmidt, T., Schütz, G. J., Baumgartner, W., Gruber, H. J., Schindler, H. Imaging of single molecule diffusion. Proceedings of the National Academy of Sciences of the United States of America. 93 (7), 2926-2929 (1996).
- Taylor, R. W., et al. Interferometric scattering microscopy reveals microsecond nanoscopic protein motion on a live cell membrane. Nature Photonics. 13 (7), 480-487 (2019).
- Kukura, P., et al. High-speed nanoscopic tracking of the position and orientation of a single virus. Nature Methods. 6 (12), 923-927 (2009).
- Jacobsen, V., Stoller, P., Brunner, C., Vogel, V., Sandoghdar, V. Interferometric optical detection and tracking of very small gold nanoparticles at a water-glass interface. Optics Express. 14 (1), 405 (2006).
- Ueno, H., et al. Simple dark-field microscopy with nanometer spatial precision and microsecond temporal resolution. Biophysical Journal. 98 (9), 2014-2023 (2010).
- Loose, M., Fischer-Friedrich, E., Herold, C., Kruse, K., Schwille, P. Min protein patterns emerge from rapid rebinding and membrane interaction of MinE. Nature Structural and Molecular Biology. 18 (5), 577-583 (2011).
- Bezeljak, U., Loya, H., Kaczmarek, B., Saunders, T. E., Loose, M. Stochastic activation and bistability in a Rab GTPase regulatory network. Proceedings of the National Academy of Sciences of the United States of America. 117 (12), 6504-6549 (2020).
- Ha, T., Tinnefeld, P. Photophysics of fluorescent probes for single-molecule biophysics and super-resolution imaging. Annual Review of Physical Chemistry. 63 (1), 595-617 (2012).
- Garcia-Parajo, M. F., Segers-Nolten, G. M. J., Veerman, J. A., Greve, J., Van Hulst, N. F. Real-time light-driven dynamics of the fluorescence emission in single green fluorescent protein molecules. Proceedings of the National Academy of Sciences of the United States of America. 97 (13), 7237-7242 (2000).
- Young, G., et al. Quantitative mass imaging of single biological macromolecules. Science. 360 (6387), 423-427 (2018).
- Piliarik, M., Sandoghdar, V. Direct optical sensing of single unlabelled proteins and super-resolution imaging of their binding sites. Nature Communications. 5, 4495 (2014).
- Cole, D., Young, G., Weigel, A., Sebesta, A., Kukura, P. Label-free single-molecule imaging with numerical-aperture-shaped interferometric scattering microscopy. ACS Photonics. 4 (2), 211-216 (2017).
- Heermann, T., Steiert, F., Ramm, B., Hundt, N., Schwille, P. Mass-sensitive particle tracking to elucidate the membrane-associated MinDE reaction cycle. Nature Methods. 18 (10), 1239-1246 (2021).
- Foley, E. D. B., Kushwah, M. S., Young, G., Kukura, P. Mass photometry enables label-free tracking and mass measurement of single proteins on lipid bilayers. Nature Methods. 18 (10), 1247-1252 (2021).
- Voss, O. H., Lee, H. N., Tian, L., Krzewski, K., Coligan, J. E. Liposome preparation for the analysis of lipid-receptor interaction and efferocytosis. Current Protocols in Immunology. 120, 1-21 (2018).
- Pincet, F., et al. FRAP to characterize molecular diffusion and interaction in various membrane environments. PLoS ONE. 11 (7), 0158457 (2016).
- Howarth, M., et al. A monovalent streptavidin with a single femtomolar biotin binding site. Nature Methods. 3 (4), 267-273 (2006).
- Allan, D., et al. soft-matter/trackpy: Trackpy v.0.5.0. Zenodo. , (2021).
- Weimann, L., et al. A quantitative comparison of single-dye tracking analysis tools using Monte Carlo simulations. PLoS ONE. 8 (5), 64287 (2013).
- Michalet, X. Mean square displacement analysis of single-particle trajectories with localization error: Brownian motion in an isotropic medium. Physical Review E - Statistical, Nonlinear, and Soft Matter Physics. 82 (4), 041914 (2010).
- Sygusch, J., Beaudry, D., Allaire, M. Molecular architecture of rabbit skeletal muscle aldolase at 2.7-A resolution. Proceedings of the National Academy of Sciences of the United States of America. 84 (22), 7846-7850 (1987).
- Heath, G. R., Scheuring, S. High-speed AFM height spectroscopy reveals µs-dynamics of unlabeled biomolecules. Nature Communications. 9 (1), 4983 (2018).
- Day, C. A., Kenworthy, A. K. Mechanisms underlying the confined diffusion of cholera toxin B-subunit in intact cell membranes. PLOS ONE. 7 (4), 34923 (2012).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati