
Method Article
Isolement de neurones hypothalamiques ciblés pour l’étude de la régulation hormonale, métabolique et électrique
Dans cet article
Résumé
Nous présentons ici un protocole pour cultiver des sous-types spécifiques de cellules hypothalamiques en culture. Les cellules peuvent être sélectionnées sur la base de marqueurs membranaires opportuns/uniques et utilisées dans de nombreuses applications, notamment l’immunofluorescence, les tests électrophysiologiques et biochimiques.
Résumé
L’hypothalamus régule les processus métaboliques fondamentaux en contrôlant des fonctions aussi variées que la prise alimentaire, la température corporelle et la libération d’hormones. Comme les fonctions de l’hypothalamus sont contrôlées par des sous-ensembles spécifiques de populations neuronales, la capacité de les isoler fournit un outil majeur pour étudier les mécanismes métaboliques. À cet égard, la complexité neuronale de l’hypothalamus pose des défis exceptionnels.
Pour ces raisons, de nouvelles techniques, telles que le tri cellulaire activé par magnétisme (MACS), ont été explorées. Cet article décrit une nouvelle application du tri cellulaire activé par le magnétisme (MACS) utilisant la technologie des microbilles pour isoler une population neuronale ciblée à partir de cerveaux de souris prénatales. La technique est simple et garantit une culture de neurones hypothalamiques primaires très pure et viable avec une grande reproductibilité. L’hypothalamus est légèrement dissocié, les neurones sont isolés sélectivement et séparés des cellules gliales, et enfin, à l’aide d’un anticorps spécifique pour un marqueur de surface cellulaire, la population d’intérêt est sélectionnée.
Une fois isolés, les neurones ciblés peuvent être utilisés pour étudier leurs caractéristiques morphologiques, électriques et endocriniennes et leurs réponses dans des conditions normales ou pathologiques. De plus, étant donné les rôles variés de l’hypothalamus dans la régulation de l’alimentation, du métabolisme, du stress, du sommeil et de la motivation, un examen plus approfondi des neurones ciblés et spécifiques à une région peut donner un aperçu de leurs tâches dans cet environnement complexe.
Introduction
L’hypothalamus est une zone à plusieurs volets du cerveau qui intervient dans les fonctions endocriniennes, autonomes, viscérales et comportementales, notamment l’alimentation, le métabolisme, le sommeil, la température corporelle, le comportement social et la libido 1,2,3,4,5. L’hétérogénéité fonctionnelle est obtenue par une combinaison synergique de mécanismes biochimiques et électriques : les neurones hypothalamiques déclenchent des potentiels d’action et sécrètent et libèrent des hormones et des neuropeptides pour moduler les régions du cerveau et les organes du corps. Enfin, les neurones hypothalamiques traduisent les messages homéostatiques du corps, répondant par une rétroaction à long et à court terme et des régulations de rétroaction6.
L’environnement neuronal complexe de l’hypothalamus comprend des neurones endocriniens magnocellulaires, libérant de l’ocytocine et de la vasopressine ; les neurones parvocellulaires, principalement impliqués dans la régulation hormonale systémique, libérant par exemple l’hormone de libération de la thyrotropine (TRH) et l’hormone de libération de la corticotropine (CRH) à l’hypophyse ; de grands neurones de projection peptidergique, libérant de l’orexine et de l’hormone de concentration de la mélanine (MCH) ; et les neurones peptrgiques parvocellulaires du noyau arqué (ARC) libérant du POMC (proopiomélanocortine) et de l’AgRP (protéine liée à l’agouti), nommés respectivement ARCPOMC et ARCAgRP. Avec les cellules sécrétrices, d’autres neurones excitateurs et inhibiteurs, notamment les neurones dopaminergiques, glutaminergiques et GABAergiques 7, sont impliqués dans la formation de circuits intrahypothalamiques et extrahypothalamiques, créant ainsi des réseaux coordonnés à grande échelle d’une hétérogénéité cellulaire considérable8.
La diversité hypothalamique est un défi que les chercheurs tentent de surmonter depuis 50 ans. Pour étudier cette hétérogénéité dans les hypothalames en développement, matures et vieillissants, les chercheurs, d’une part, ont utilisé le séquençage de l’ARN unicellulaire pour explorer l’organisation neuronale, ainsi que les signatures moléculaires et transcriptomiques. Cet effort a fourni un regard perspicace sur les rôles variés des neurones hypothalamiques et a abordé les liens entre l’identité cellulaire et son rôle possible dans le système physiologique 8,9,10. D’autre part, les fonctions neuronales ont été étudiées par des manipulations optogénétiques et des approches comportementales de photométrie des fibres, ce qui a permis d’examiner de près la structure des circuits. Au cours des deux dernières décennies, la technologie de la Cre-recombinase a permis aux chercheurs de stimuler ou d’inhiber ontogénétiquement un groupe ciblé de neurones tout en observant les changements dans les comportements et les réponses de l’organisme 6,11,12.
Cependant, ces approches examinent les fonctions hypothalamiques d’un point de vue général sans approfondir les mécanismes cellulaires spécifiques ou la base biologique de leur rôle dans l’environnement hypothalamique complexe. Pour y remédier, très peu d’études se sont concentrées sur l’étude des propriétés moléculaires, biochimiques et électriques à l’aide de cultures hypothalamiques primaires hétérogènes. Ces études ont cherché à disséquer des processus neuronaux spécifiques dans un environnement complexe et ont généré des modèles intégratifs de mécanismes physiologiques13,14,15. Néanmoins, les cultures non spécifiques posent des défis importants. Par exemple, la connectivité physiologique et la distribution anatomique des neurones sont perturbées par le placage de neurones de différentes régions hypothalamiques qui n’interagiraient normalement pas, créant des effets confondants. De plus, chaque région a des rôles différents et des populations neuronales variées, ce qui rend difficile l’étude de processus biologiques simples.
Pour relever ces défis, au cours de la dernière décennie, de nouvelles approches ont été mises en œuvre pour isoler les neurones d’intérêt, telles que l’immunopanoramique, le tri cellulaire activé par fluorescence (FACS) et le tri cellulaire activé magnétiquement (MACS). L’immunopanoramique est une stratégie employée pour purifier les cellules ciblées à l’aide de boîtes enrobées d’anticorps pour une série de sélections non neuronales (négatives) et neuronales (positives). Bien que cette technique puisse, en principe, générer des cultures cellulaires purifiées à haut rendement, dans la pratique, elle est principalement utilisée pour les astrocytes et les oligodendrocytes, car ces cellules peuvent résister à des heures de manipulation16,17. La technologie FACS est un outil puissant pour trier les cellules en fonction des marqueurs fluorescents et des caractéristiques cellulaires à l’aide de la cytométrie en flux18,19,20. Cependant, très peu d’études ont utilisé cette méthode pour isoler des cellules en vue de leur culture. La technique est coûteuse et nécessite un personnel hautement qualifié pour être utilisée et entretenue. De plus, il est difficile de maintenir des cellules viables et stériles à la fin de la procédure de tri21. Dans l’ensemble, le MACS semble être une technique simple et peu coûteuse pour obtenir des cultures très pures et viables de neurones primaires hypothalamiques. La méthode utilise des billes magnétiques liées aux cellules par l’intermédiaire d’un anticorps. Cela permet d’isoler les cellules à l’aide du champ magnétique de la colonne.
Nous décrivons ici une méthode basée sur la technologie MACS, qui est généralement utilisée avec les neurones corticaux. Ce protocole permet d’isoler, en principe, des neurones hypothalamiques viables et d’une grande pureté. Dans cette étude, nous préparons des cultures primaires de neurones exprimant le récepteur de la leptine (LepR), tels que les neurones ARCPOMC et ARCAgRP , qui ne sont présents que dans le noyau arqué. Ces neurones réagissent à la leptine, une hormone anorexigène sécrétée par le tissu adipeux, de manière biochimique et électrique. Par conséquent, l’isolement de ce groupe de neurones en culture permet d’étudier leurs propriétés hormonales, métaboliques et électriques in vitro.
Protocole
NOTE : Une vue générale de la procédure expérimentale est illustrée graphiquement à la figure 1A. Toutes les expériences sur des souris réalisées dans le cadre de cette étude ont été approuvées par le Comité de protection et d’utilisation des animaux (IACUC) de notre établissement. Nous avons utilisé des souris C57BL6/J âgées de 3 mois, qui ont été hébergées dans un vivarium approuvé par l’Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC), sous les soins d’un vétérinaire. Les souris vivaient dans de grandes cages, avec un cycle lumière-obscurité de 12 heures, et étaient nourries ad libitum.
1. Vérification de la conception et de la grossesse
- Placez des souris de n’importe quel milieu et génotype d’intérêt pour la reproduction. Notez la date et le poids de la femelle avant la conception.
- Après 6 h, inspectez la femelle à la recherche d’une plaque à l’aide d’une sonde. Si la plaque est présente, séparez la femelle du mâle. Si la plaque n’est pas présente, gardez la femelle dans la cage jusqu’au lendemain, puis séparez les souris.
- Aux jours 7, 10 et 14 après la conception, pesez la femelle pour confirmer la grossesse.
2. Préparation des médias, des plaques à 24 puits et des matériaux
- Le jour de l’isolement des cellules, placez les lamelles en verre recouvertes de poly-D-lysine prêtes à l’emploi (voir le tableau des matériaux) dans une plaque à 24 puits comme suit :
- Sous une hotte biologique, stérilisez un seul paquet contenant 15 lamelles avec 70% d’éthanol et laissez sécher. Ouvrez l’emballage et placez les lamelles dans une plaque de 60 mm. Secouez la plaque horizontalement pour séparer les lamelles. Ensuite, retournez la plaque pour ramasser des lamelles simples à placer dans les puits d’une plaque à 24 puits.
- Lavez les lamelles une fois avec 1,0 mL de solution saline équilibrée Hank’s (HBSS) stérile pendant 5 minutes.
- Dans l’intervalle, préparez 20,0 mL de substrat de placage comme suit : à 18,31 mL de BME (Basal Medium Eagle, + Sels d’Earle), complété par 1,0 mL de sérum de veau fœtal inactivé par la chaleur (FBS), ajoutez 200 μL de pyruvate de sodium (à partir d’un stock 100x), 200 μL de glutamine (à partir d’un stock de 200 nM) et 100 μL de pénicilline/streptomycine (à partir d’un stock 200x).
- Remplacez le HBSS dans les puits par 1,0 mL de média de placage et placez le plateau dans l’incubateur à 37 °C.
- À l’aide d’un bec Bunsen, polir au feu trois pipettes Pasteur de diamètres décroissants. En tenant la pipette d’une main, insérez l’embout dans la flamme et retirez-le rapidement. Répétez le processus jusqu’à ce que la pointe lisse et que le diamètre soit réduit au diamètre souhaité (évalué à l’œil).
3. Préparation des réactifs pour la dissociation des tissus neuraux, en suivant les instructions du kit de dissociation neurale
- Après avoir réchauffé le tampon de digestion 1 à température ambiante, préparez le mélange enzymatique 1 en mélangeant 50 μL d’enzyme 1 avec 1,91 mL de tampon 1 et le mélange enzymatique 2 en mélangeant 15 μL d’enzyme 2 avec 30 μL de tampon de digestion 2. Les mélanges sont suffisants pour être utilisés pour tous les tissus cérébraux des embryons.
- Préparez de l’albumine sérique bovine (BSA) à 0,5 % dans HBSS, par exemple, 0,25 g dans 50,0 mL d’HBSS.
4. Extraction d’embryons
- Autoclavez deux pinces fines droites, une pince à pointe incurvée et une paire de ciseaux chirurgicaux fins et stérilisez-les avec de l’éthanol à 70 % avant utilisation. Ensuite, remplissez les boîtes de Pétri avec HBSS.
- Euthanasier une mère enceinte E14-E16 dans la chambre CO2 et effectuer une luxation cervicale.
REMARQUE : Les étapes suivantes doivent être effectuées sous la hotte dans des conditions stériles : - Stériliser l’abdomen avec de l’éthanol à 70 %. Coupez la cavité abdominale de la symphyse pubienne à l’apophyse xiphoïde de la cage thoracique avec des ciseaux chirurgicaux et des pinces.
- Extrayez la corne utérine et placez-la dans une assiette de 100 mm remplie d’HBSS glacée et lavez-la soigneusement.
- Extrayez et séparez tous les embryons de l’utérus à l’aide d’une pince fine. Décapitalisez rapidement les embryons à l’aide de ciseaux chirurgicaux fins et/ou de pinces. Placez les têtes dans la boîte de Pétri de 60 mm remplie de HBSS.
5. Extraction, collecte et dissociation tissulaire de l’hypothalamus
- Placez une pince fine dans la cavité de l’œil pour maintenir le cerveau. À l’aide de l’autre pince fine, retirez la peau et le crâne en pelant jusqu’à ce que le cerveau soit visible. Distinguer le cerveau des autres tissus en fonction de son apparence blanche. La peau et le crâne sont roses et riches en vascularisation.
- Retirez le cerveau du crâne à l’aide de la pince à pointe incurvée et retirez le cerveau des bulbes olfactifs, en le retournant à l’envers.
- Maintenant, le cortex est ventral, et l’hypothalamus est visible dorsalement sur la surface supérieure (Figure 1B). À l’aide de la pince incurvée, retirez la couche de méninges et de vaisseaux sanguins jusqu’à ce que le cerveau apparaisse blanc et clair.
- À l’aide de la pince incurvée, séparez la zone hypothalamique du reste du cerveau.
- Coupez l’hypothalamus en 3-4 petits morceaux et à l’aide d’une pipette, transférez les morceaux dans un tube de 15 ml.
- Répétez les étapes pour les autres embryons pendant que le tube est sur de la glace.
- Remplissez le tube avec 6,0 mL d’HBSS et laissez le tissu se déposer, retirez le surnageant et ajoutez le mélange d’enzymes 1. Mélangez et agitez doucement le tube pour éviter que le tissu ne se détache.
- Incuber le tube dans un bain-marie à 37 °C pendant 15 min et agiter le tissu toutes les 5 min pour le remettre en suspension.
- Au bout de 15 min, ajouter 30 μL de mélange enzymatique 2. Dissocier le tissu cérébral à l’aide de la pipette Pasteur du plus grand diamètre (<1 mm). Pipeter de haut en bas 10x sans former de bulles.
- Incuber pendant 10 min à 37 °C au bain-marie. Agitez doucement le tube pour remettre le tissu en suspension toutes les 5 minutes.
- Au bout de 10 min, ajouter les 15 μL restants du mélange enzymatique 2. Dissociez le tissu 10x avec les deux autres pipettes polies au feu de diamètre décroissant, de haut en bas sans former de bulles.
- Dans le tube contenant le tissu dissocié, ajouter immédiatement 10,0 mL de HBSS-0,5 % BSA et centrifuger à 300 × g pendant 10 min à température ambiante.
- Aspirer le surnageant et remettre en suspension la pastille cellulaire dans 1,0 mL de HBS-0,5 % BSA.
6. Comptage cellulaire
- Diluer la suspension cellulaire 1 :5 en utilisant HBSS-0,5% BSA.
- Placer 10 μL de la suspension cellulaire diluée dans une chambre de comptage Neubauer.
- À l’aide d’un microscope à fond clair, ne comptez que les cellules qui se trouvent dans les carrés d’angle des quatre chambres. Calculez la moyenne et multipliez-la par 5 × 104.
REMARQUE : Assurez-vous qu’il y a >106 cellules pour procéder à l’isolement cellulaire ; Le nombre optimal de cellules est 107.
7. Sélection négative
REMARQUE : La sélection négative permet aux utilisateurs d’obtenir une culture de neurones primaires purs en séparant les cellules neuronales et non neuronales. Utilisez des solutions prérefroidies.
- Centrifuger la suspension cellulaire à 300 × g pendant 3 min (la centrifugation peut être prolongée jusqu’à 10 min). Aspirer doucement le surnageant et remettre la pastille en suspension à une concentration de 10à 7 cellules dans 80 μL de HBSS-0,5 % BSA.
- Ajouter 20 μL de Cocktail Biotine-Anticorps pour Cellules Non Neuronales et incuber pendant 5 min à 4 °C.
- Lavez les cellules pour éliminer les anticorps libres avec 2,0 mL de HBSS-0,5 % BSA et centrifugez à 300 × g pendant 3 min.
- Aspirer doucement le surnageant et remettre la pastille en suspension dans 80 μL de HBSS-0,5 % BSA. Ajouter 20 μL de microbilles anti-biotine, bien mélanger et incuber pendant 10 min à 4 °C.
- Ajouter 0,5 mL de HBSS-0,5 % BSA pour un maximum de 10à 7 cellules et attendre que la colonne magnétique soit prête.
8. Séparation magnétique, sélection négative
REMARQUE : La séparation magnétique est une étape cruciale qui permet la séparation des cellules non neuronales des cellules neuronales. L’échantillon contenant des cellules neuronales et non neuronales est passé à travers le champ magnétique et les cellules non neuronales, qui sont liées à un complexe biotine-anticorps-billes magnétiques, sont piégées dans la colonne (Figure 1C). Les cellules neuronales libres sont éluées à travers la colonne et sont recueillies dans un tube de 15 ml.
- Préparez le support (inclus dans le kit) avec le séparateur et la colonne MS comme illustré à la Figure 1C.
- Ouvrez la colonne et installez le support uniquement lorsque les cellules sont prêtes à être séparées.
- Rincez la colonne avec 0,5 mL de HBSS-0,5 % BSA. Attendez que la solution cesse de couler.
- Pour prélever des cellules neuronales, placez un tube de 15 mL sous la colonne et faites passer 0,5 mL de la suspension cellulaire à travers la colonne. Collectez l’éluat dans le tube jusqu’à ce qu’il cesse de couler. Pour capturer les cellules neuronales résiduelles, laver la colonne 3 x 0,5 mL de HBSS-0,5 % BSA.
- Pour prélever des cellules non neuronales, retirez la colonne de l’aimant et placez-la dans un nouveau tube de 15 ml. Ajouter 1,0 mL de HBSS--0,5% BSA dans la colonne et utiliser le piston pour collecter les cellules non neuronales marquées magnétiquement.
- Centrifuger les cellules neuronales et non neuronales à 300 × g pendant 3 min. Aspirer doucement le surnageant et remettre les cellules en suspension dans 1,0 mL de HBSS-0,5 % BSA. Comptez les cellules comme décrit précédemment dans la section 6.
- Si nécessaire, plaquez les cellules non neuronales dans une plaque à 24 puits ; sinon, jetez-les.
9. Sélection positive
REMARQUE : Une fois qu’une suspension de cellules neuronales pures est obtenue, une sélection positive est effectuée pour isoler les cellules ciblées. Les cellules peuvent être isolées à l’aide d’un anticorps spécifique conjugué à la biotine pour un antigène de surface. L’anticorps est reconnu par des billes magnétiques anti-biotine. En faisant circuler la suspension cellulaire à travers la colonne, seules les cellules d’intérêt sont piégées dans le champ magnétique.
- Centrifuger la suspension de cellules neuronales pures à 300 × g pendant 3 min. Aspirer doucement le surnageant et remettre la pastille en suspension dans 80 μL de HBSS-0,5 % BSA. Ajouter l’anticorps spécifique en suivant les instructions du fabricant et incuber à 4 °C pendant 10 min.
REMARQUE : Si l’on recherche des cellules exprimant le LepR, nous suggérons un anticorps biotinylé à la leptine R de souris (voir le tableau des matériaux) à une concentration de 0,50 μg/106 cellules. - Rincer l’excès d’anticorps avec 2,0 mL de HBSS-0,5 % BSA et centrifuger à 300 × g pendant 3 min.
- Retirer le surnageant, remettre la pastille en suspension dans 80 μL de HBSS-0,5 % BSA et ajouter 20 μL de microbilles anti-biotine. Incuber à 4 °C pendant 10 min.
- Pour chaque tranche de 10à 7 cellules, ajoutez 0,5 mL de HBSS-0,5 % BSA et attendez que la colonne magnétique soit prête.
10. Séparation magnétique, sélection positive
- Préparez le support avec le séparateur et la colonne MS. Rincez la colonne MS avec 0,5 mL de HBSS-0,5 % BSA. Attendez que l’égouttement s’arrête.
- Placez un tube de 15 mL sous la colonne, faites passer 0,5 mL de la suspension cellulaire à travers la colonne et collectez l’éluat comprenant les cellules neuronales non spécifiques. Pour nettoyer la colonne de cellules neuronales résiduelles non spécifiques, laver avec 3 x 0,5 mL de HBSS-0,5% BSA.
- Retirez la colonne de l’aimant, placez-la dans un nouveau tube de 15 ml et ajoutez 1,0 ml de HBSS-0,5 % BSA. Utilisez le piston pour rincer les cellules ciblées.
- Centrifuger les deux tubes à 300 × g pendant 3 min. Retirer délicatement le surnageant et le remettre en suspension dans 0,5 mL de milieu de placage.
- Comptez les cellules comme décrit précédemment dans la section 6.
- Plaquer à la fois les cellules ciblées en tant que témoins positifs et les cellules non spécifiques en tant que témoins négatifs à une densité de 120 000 à 200 000 cellules/mm3 dans la plaque à 24 puits préalablement préparée comme décrit à la section 2 et incuber à 37 °C dans 5 % de CO 2, 9 % d’O2 et 95 % d’humidité pendant 12 h.
11. Entretien de la culture cellulaire
- Préparez 20 mL de milieu de culture avec 19,2 mL de milieu de culture neuronale, 400 μL de supplément de vitamine B27 (à partir d’un stock 50x), 200 μL de glutamine (à partir d’un stock de 200 μM) et 100 μL de pénicilline/streptomycine (à partir d’un stock 200x).
- Remplacez le support de placage de la plaque à 24 puits contenant des cellules neuronales ou non neuronales.
- Laver avec 2 x 1,0 mL de HBSS.
- Ajouter 1,0 mL de milieu de culture.
- Rafraîchissez le support tous les 2/3 jours en remplaçant 0,5 mL d’ancien support par 0,5 mL de support neuf.
REMARQUE : Les cellules peuvent être maintenues en culture et utilisées jusqu’à 21 jours in vitro (DIV21).
12. Coloration par immunofluorescence neuronale
- Douze heures avant la coloration, préparez une solution composée de méthanol 50/50 et d’acétone et refroidissez-la à -20 °C pendant la nuit.
- Lavez les neurones dans la plaque à 24 puits avec 2 x 1,0 mL de solution saline tamponnée au phosphate (PBS) 1x pendant 5 min.
- Remplacer la solution de PBS par 1,0 mL de la solution 50/50 et incuber dans de la glace pendant 20 minutes.
- Laver avec 1x PBS pendant 3 x 5 min.
- Bloquez les neurones avec 3% de BSA dans 1x PBS pendant 1 h à température ambiante.
- Préparez la solution primaire d’anticorps dans 3 % de BSA dans 1x PBS, en utilisant la concentration d’anticorps mentionnée dans les instructions du fabricant. Les anticorps et les concentrations utilisés sont indiqués dans le tableau des matériaux.
- Remplacez la solution bloquante par la solution d’anticorps primaire et incubez à 4 °C pendant la nuit.
- Lavez les cellules pendant 3 x 10 min avec 1x PBS.
- Préparez la solution d’anticorps secondaire avec 3 % de BSA dans 1x PBS, en utilisant les concentrations des anticorps en suivant les instructions du fabricant. Les anticorps secondaires utilisés sont énumérés dans le tableau des matériaux.
- Incuber les cellules avec la solution d’anticorps secondaire à température ambiante pendant 1 h.
- Lavez les cellules pendant 3 x 10 min avec 1x PBS.
- Laissez les neurones dans 1x PBS pendant la procédure de montage. Placez une petite goutte de milieu de montage (avec ou sans 4',6-diamidino-2-phénylindole pour l’identification nucléaire) sur la lame de microscope. Extrayez une lamelle de verre contenant des neurones à l’aide d’une pince et tapotez le côté de la lamelle sur un mouchoir en papier pour sécher l’excès de PBS. Retournez la lamelle sur le support de montage, en vous assurant que les neurones font face aux lames du microscope ; Pressez délicatement et retirez l’excédent de support de montage avec du papier de soie.
- Les lames de microscope sont prêtes à être analysées avec un microscope à fond clair ou un microscope confocal.
Résultats
Cet article décrit un protocole d’isolement des neurones ciblés de l’hypothalamus (Figure 1). L’objectif de la méthode est d’étudier des caractéristiques neuronales spécifiques dans un contexte contrôlé et isolé. Ainsi, des embryons de souris ont été extraits des mères gravides à E14-E16. Les méninges ont été enlevées et l’hypothalamus a été isolé du reste du cerveau. Le tissu a été délicatement dissocié avec deux mélanges d’enzymes fraîchement préparés à l’aide du kit de dissociation référencé. Tout d’abord, les cellules non neuronales ont été séparées des cellules neuronales - glie, microglie et neurones ont été collectés dans la même suspension unicellulaire. À cette fin, les cellules non neuronales ont été marquées à l’aide d’un cocktail d’anticorps reconnaissant les épitopes de surface non neuronaux. Après incubation, le complexe anticorps-cellule a été conjugué avec des microbilles magnétiques, puis passé à travers une colonne magnétique pour piéger les cellules non neuronales.
Cette étape a donné lieu à deux suspensions cellulaires, l’une contenant des cellules non neuronales et l’autre contenant des cellules neuronales. Les deux suspensions pouvaient être plaquées immédiatement. Alternativement, la suspension des cellules neuronales pourrait être manipulée davantage pour séparer une sous-population neuronale (suspension ciblée) du reste en utilisant la même stratégie. Sur la base de l’expérience qui nous intéresse, les cellules neuronales peuvent être plaquées de 125 000 à 200 000 cellules/mm3. Les cultures moins denses peuvent être utilisées pour analyser les neurones à la résolution d’une seule cellule : du développement axonal, de la formation synaptique et de la transmission à l’électrophysiologie. Les cultures plus denses peuvent être utilisées pour des analyses biochimiques, y compris l’extraction d’ADN et d’ARN, le transfert Western, le transfert de Southern, le transfert de Nord, la PCR en temps réel et le séquençage de l’ARN.
Dans cette étude, LepR a été ciblé pour isoler les neurones impliqués dans le système de mélanocortine, tels que les neurones ARCPOMC et ARCAgRP. Les cellules ont été plaquées à des densités allant de 120 000 cellules/mm 3, pour les neurones LepR+, à 200 000 cellules/mm3 pour les populations neuronales génériques. Après 48 h, les neurones LepR+ ont commencé à former des neurites (Figure 2). À DIV4, les extensions axonales ont progressé, tandis que les processus dendritiques ont commencé à apparaître. À DIV6, les neurones étaient suffisamment développés et étaient donc prêts à être analysés. Des expériences d’immunofluorescence sur des neurones LepR+ ont montré une expression de 99% de LepR (vert, Figure 3A). Aucune cellule gliale ou autre cellule non neuronale n’a été observée, ce qui confirme la pureté de la culture neuronale primaire. La nature neuronale des cellules a été confirmée par la coloration de la protéine 2 associée aux microtubules (MAP2), avec l’identification d’axones et de protubérances dendritiques (Figure 3B). À DIV10, 30 % des cellules LepR+ exprimaient POMC (rouge). On s’y attend car la majorité des cellules LepR+ expriment soit POMC, soit AgRP. Les figures 3C et D illustrent la colocalisation entre les signaux POMC et LepR. Il est à noter que la colocalisation était importante au niveau et autour du noyau, comme on pouvait s’y attendre.
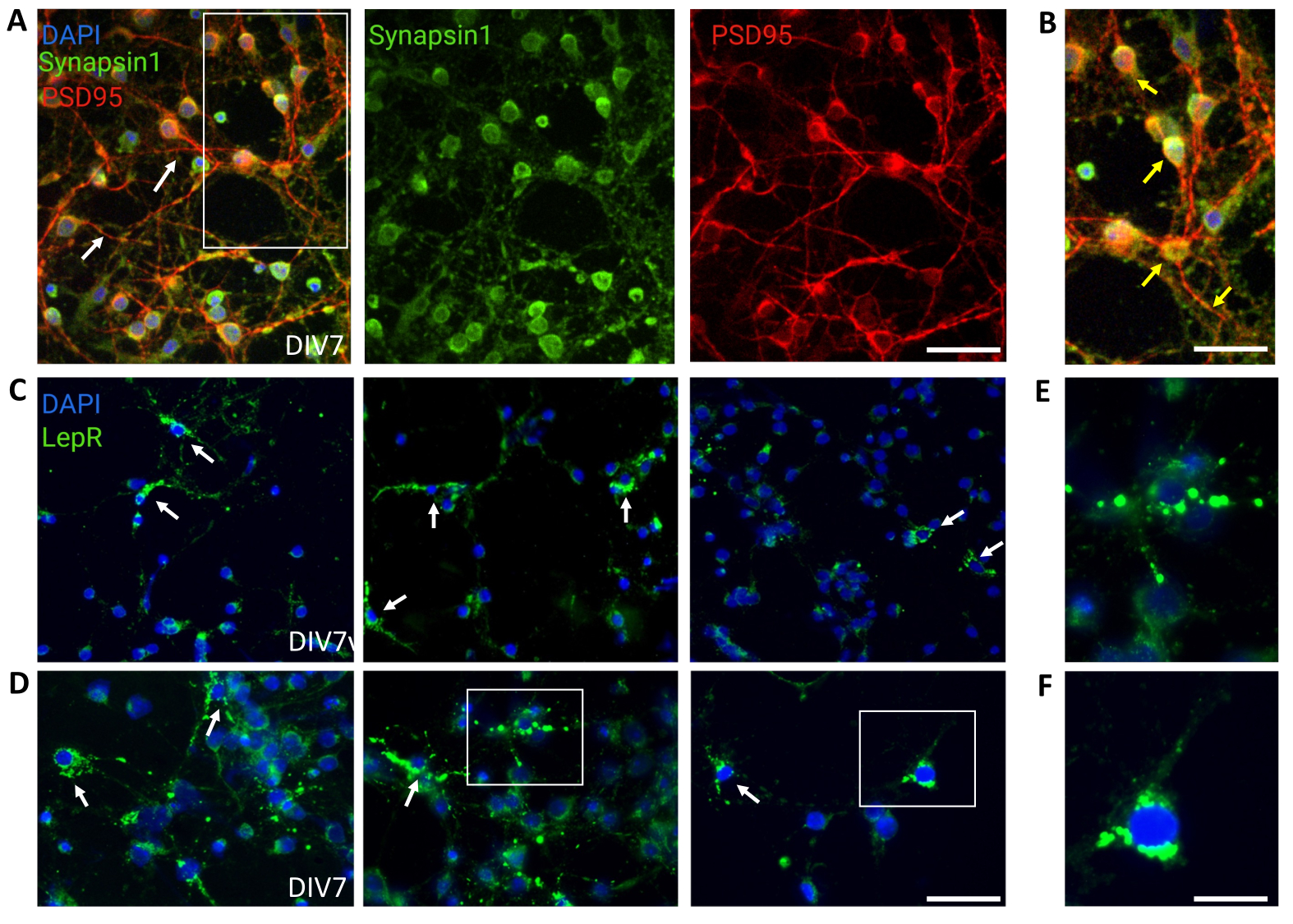
Des cultures générales contenant des populations neuronales hypothalamiques hétérogènes ont été utilisées pour le contrôle. L’immunofluorescence a démontré une connectivité et une fonctionnalité synaptiques, telles qu’évaluées par la co-coloration de la synapsine-1 (vert) et de la PSD 95 (rouge) (Figure 4A,B). Le nombre de neurones LepR+ présents dans la culture générale était de ~5%, un pourcentage cohérent avec l’idée que la majorité des neurones exprimant LepR avaient été sélectionnés au cours du processus de séparation magnétique (les cellules LepR+ représentatives sont illustrées à la figure 4C,D). Toutes les données générées ou analysées au cours de cette étude sont disponibles à https://doi.org/10.5061/dryad.cnp5hqc9c.

Figure 1 : Organigramme expérimental et mise en place. (A) Représentation graphique de la procédure expérimentale. Go-no-go : ≥106 cellules sont nécessaires pour procéder à l’isolement cellulaire ; Le nombre optimal de cellules est 107. (B) Image représentative d’un cerveau d’embryon E16. L’hypothalamus et les méninges sont indiqués. (C) Configuration MACS utilisée pour la séparation et l’isolement des cellules ciblées. Le support magnétique, le séparateur magnétique et la colonne sont indiqués. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Culture neuronale entre DIV2 et DIV6. (A) Les cellules LepR+ obtenues à partir de la sélection positive montrent une densité cellulaire et une connectivité réduites, mais un développement normal des neurites (flèches rouges), des axones (flèches bleues) et des dendrites (flèches vertes). Les cellules ont été plaquées à une densité de 120 000 cellules/mm3. Barre d’échelle = 100 μm. (B) Les neurones hypothalamiques génériques, plaqués à une densité de 200 000 cellules/mm3, présentent des caractéristiques normales de développement et de croissance ainsi qu’une connectivité (flèches noires). Barre d’échelle = 100 μm. Abréviations : DIV = jours in vitro ; LepR = récepteur de la leptine. Toutes les données générées ou analysées pour construire ce chiffre sont disponibles à https://doi.org/10.5061/dryad.cnp5hqc9c. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Les neurones in vivo sont récapitulés par des neurones LepR+ en culture. (A) Images représentatives d’une culture de neurones exprimant le LepR (vert ; DAPI est bleu). Quatre-vingt-dix-neuf pour cent des cellules exprimaient le LepR. Les neurones ont été plaqués à 120 000 cellules/mmde densité 3. À DIV7, les neurones présentaient des axones allongés, une maturation dendritique et une connectivité neuronale (flèches). Barre d’échelle = 40 μm. (B) L’immunofluorescence avec anti-MAP2 a été utilisée pour confirmer la nature neuronale des cellules. Les morphologies spécifiques aux neurones telles que les axones, les dendrites et les protubérances sont démontrées (flèches). Barre d’échelle = 40 μm. (C) Grossissement des cellules LepR+. LepR en vert et DAPI en bleu. Barre d’échelle = 10 μm. (D) Co-coloration avec LepR (vert) et POMC (rouge). Environ 30 % des neurones LepR+ étaient co-immunoréactifs à la POMC, qui a été détectée au niveau du noyau (flèches rouges). Barre d’échelle = 10 μm. Abréviations : DAPI = 4',6-diamidino-2-phénylindole ; MAP2 = protéine 2 associée aux microtubules ; POMC = proopiomélanocortine ; LepR = récepteur de la leptine. Toutes les données générées ou analysées pour construire ce chiffre sont disponibles à https://doi.org/10.5061/dryad.cnp5hqc9c. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Les cellules in vivo sont récapitulées par des neurones hypothalamiques génériques en culture. (A) Images représentatives d’une culture neuronale hypothalamique générique colorée à la synapsine 1 (vert), à la PSD95 (rouge) et au DAPI (bleu). Les neurones présentaient une connectivité et une fonctionnalité synaptique bien développées (flèches). Barre d’échelle = 40 μm. (B) Grossissement de la case en (A), montrant la colocalisation de la synapsine 1 (vert) et de la DSP95 (rouge, flèches). Barre d’échelle = 20 μm. (C,D) Images représentatives montrant des cellules LepR+ dans une culture générale. Les cellules LepR+ (en vert) représentaient ~5% du total. Les cellules LepR+ représentatives sont indiquées par des flèches. Barre d’échelle = 40 μm. (E,F) Grossissements des cases en (C,D) montrant les récepteurs de la leptine puncta verts localisés au niveau du soma. Barre d’échelle = 20 μm. Toutes les données générées ou analysées pour construire ce chiffre sont disponibles à https://doi.org/10.5061/dryad.cnp5hqc9c. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
L’étude des propriétés biochimiques et électriques des neurones hypothalamiques est essentielle pour comprendre les bases moléculaires du métabolisme, de la thermorégulation, de la gestion de l’humeur, du comportement alimentaire, etc. Cependant, l’hétérogénéité neuronale de l’hypothalamus rend cet effort difficile, et des méthodes d’isolement et d’étude de sous-populations hypothalamiques spécifiques sont nécessaires.
Les techniques in vivo utilisent la CRE-recombinase, l’optogénétique, la photométrie des fibres et l’imagerie calcique. Ces approches permettent principalement d’étudier les propriétés électriques des neurones hypothalamiques, et très peu de méthodes sont actuellement disponibles pour étudier leurs attributs non électriques. La technologie MACS développée dans cette étude pourrait fournir une technique permettant d’isoler in vitro des sous-populations neuronales hypothalamiques spécifiques, permettant ainsi des traitements et des analyses ciblés. Les cultures neuronales sont plus simples à gérer que les co-cultures de différentes populations neuronales. De plus, les cultures pures évitent les effets confondants découlant de la présence de cellules gliales et de microglies. Ainsi, des neurones de la même région hypothalamique et du même type pourraient être étudiés en réponse à des entrées métaboliques et hormonales spécifiques.
Dans ce protocole, nous avons sélectionné des neurones hypothalamiques exprimant la LepR. Des cellules LepR+ isolées ont été cultivées pour étudier leurs caractéristiques cellulaires, morphologiques et moléculaires qui sont difficiles à étudier in vivo. La pureté des cultures était de 99 %, ce qui confirme la précision de la méthode. De plus, les cellules LepR+ étaient saines et viables à DIV7 jusqu’à DIV21.
Cette technique présente cependant certaines limites. Les cultures de neurones purs E18 ou plus anciennes sont difficiles à maintenir. Par conséquent, la fenêtre d’extraction est limitée à E14-E16. Cela implique que les changements cellulaires survenant après E16 ne sont pas pris en compte. Par exemple, l’expression du récepteur de la leptine dans les neurones ARC augmente au début de la période postnatale22. La procédure d’isolement doit être effectuée le plus rapidement possible pour réduire le stress cellulaire et la mort et améliorer le rendement. La procédure peut prendre jusqu’à 5 h ; Par conséquent, il est essentiel de maintenir des conditions stériles et de réduire les manipulations au minimum nécessaire. La sélection positive peut conduire à un faible rendement en raison des faibles quantités de tissu disponible, ce qui limite le nombre d’expériences pouvant être effectuées avec une seule préparation. Une mortalité neuronale élevée a été observée, probablement due à une faible densité cellulaire et à une connectivité neuronale et à un soutien intraneuronal réduits.
De plus, l’anticorps ciblant l’antigène d’intérêt doit se lier à la surface de la cellule pour garantir une séparation correcte ; Habituellement, les anticorps utilisés pour la cytométrie en flux conviennent à la technique MACS. Si l’anticorps n’a jamais été utilisé auparavant dans des méthodes de séparation cellulaire, des expériences de validation et de titrage sont nécessaires pour déterminer l’utilisation et la concentration idéales. L’extraction des cellules ciblées nécessite un marqueur de surface cellulaire. Ici, nous avons utilisé un anticorps biotinylé, mais en principe, des anticorps conjugués avec d’autres molécules, telles que le FITC (isothiocyanate de fluorescéine) et le PE (anti-phycoérythrine purifiée), pourraient également être utilisés. La technologie MACS pourrait également être appliquée aux neurones exprimant un fluorophore, tel que la GFP ou une autre protéine Tag, ce qui pourrait augmenter la spécificité et le rendement. Si un fluorophore n’est pas utilisé, l’alternative serait de confirmer l’expression de la molécule d’intérêt par immunofluorescence avant d’effectuer des expériences sur des cellules vivantes. Des études futures testeront la validité de ces alternatives.
Un aspect important que cette étude n’a pas abordé concerne la « fidélité » des populations sous-neuronales. Nous avons constaté que les neurones LepR+ cultivés exprimaient POMC, qui est une signature des neuronesPOMC ARC natifs. Cependant, d’autres tests seront nécessaires pour conclure que les cultures neuronales LepR+ récapitulent leurs homologues in vivo natifs. Dans l’ensemble, le protocole d’isolement neuronal MACS présenté ici peut fournir une méthode valide et efficace pour étudier les mécanismes hypothalamiques in vitro qui seraient autrement difficiles à étudier in vivo.
Déclarations de divulgation
Les auteurs déclarent n’avoir aucun conflit d’intérêts.
Remerciements
Des figures graphiques ont été créées avec BioRender.com. Ce travail a été soutenu par une subvention de la NIA (R01AG060919) et une subvention de la NSF (2030348) à FS.
matériels
| Name | Company | Catalog Number | Comments |
| Embryo extraction | |||
| 1 curved point forceps | Fine Science Tools | 11270-20 | Dumont |
| 1 fine surgical scissor | Fine Science Tools | 14058-11 | Dumont |
| 100 mm Petri dish | Corning | 430167 | |
| 2 straight fine forceps | Fine Science Tools | 11254-20 | Dumont |
| 60 mm Petri dish | Corning | 430196 | |
| 70% ethanol | Decon Laboratories, INC. | 2801 | Ethanol 190 Proof |
| Anti-Biotin MicroBeads 1mL | Miltenyi Biotec | 130-115-390 | |
| Anti-MAP2 antibody | Abcam | ab5392 | 1 : 800 |
| Bench pads | |||
| Bovine Serum Albumin | Sigma-Aldrich | A9418-50G | |
| Buffer Y | Miltenyi Biotec | 130-094-802 | |
| Buffer Z | Miltenyi Biotec | 130-094-802 | |
| Cell Culture | |||
| Anti-Biotin MicroBeads 1mL | Miltenyi Biotec | 130-115-390 | |
| Bovine Serum Albumin | Sigma-Aldrich | A9418-50G | |
| Buffer Y | Miltenyi Biotec | 130-094-802 | |
| Buffer Z | Miltenyi Biotec | 130-094-802 | |
| Enzyme A | Miltenyi Biotec | 130-094-802 | |
| Enzyme P | Miltenyi Biotec | 130-094-802 | |
| GG-12-1.5, 12 mm dia.#1.5 thick 100 pc cell culture tested German coverglasses | Neuvitro Corporation | GG-12-15 | |
| Gibco B-27 Supplement 10 mL | ThermoFisher | 17504-044 | |
| Gibco Basal Medium Eagle (BME) 500 mL | ThermoFisher | 21010046 | (+) Earle's Salts, (-) L-Glutamine |
| Gibco HBBS (1x) Hanks' Balanced Salt Solution 500 mL | ThermoFisher | 14025092 | Calcium, Magnesium, No phenol red |
| Gibco HI FBS 100 mL | ThermoFisher | 16140-063 | |
| Gibco L-Glutamine 200 mM (100x) | ThermoFisher | 25030-081 | |
| Gibco Penicilline/Streptomicine | ThermoFisher | 15140-122 | 10,000 U/mL |
| Gibco Sodium Pyruvate (100 mM) 100 mL | ThermoFisher | 11360070 | |
| MiniMACS Separator and Starting Kit | Miltenyi Biotec | 130-042-102 | |
| Mouse Leptin R Biotinylated Antibody | R&D Systems | ABAF497 | 0.25 μg/106 cells |
| MS Column | Miltenyi Biotec | 130-042-201 | |
| Neaubeaur-Improved Brightline 100 µm Chamber | Hausser Scientific | 3120 | |
| Neural Tissue Dissociation Kit - Postnatal Neurons | Miltenyi Biotec | 130-094-802 | |
| Neuronal Culture Medium 500 mL | ThermoFisher | 88283 | |
| Non-Neuronal Cell Biotin-Antibody Cocktail mouse 1 mL | Miltenyi Biotec | 130-115-389 | |
| Olympus SZ61 Zoom Stereomicroscope | Olympus Life Science | SZ61/SZ51 | |
| Pierce Primary Neuron Isolation Kit | ThermoFisher | 88280Y | |
| Staining | |||
| Anti-MAP2 antibody | Abcam | ab5392 | 1 : 800 |
| Donkey anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 | ThermoFisher | A32766 | 1 : 500 |
| Donkey anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 | ThermoFisher | A32790 | 1 : 500 |
| Dulbecco's Phosphate Buffered Saline (DPBS) | Sigma Aldrich | MFCD00131855 | |
| Goat anti-Chicken IgY (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 647 | ThemoFisher | A32933 | 1 : 500 |
| Goat anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor 594 | ThermoFisher | A11037 | 1 : 200 |
| Invitrogen Leptin Receptor Recombinant Rabbit Monoclonal Antibody (JA73-01) | ThermoFisher | MA5-32685 | 1 : 500 |
| Mouse Leptin R Biotinylated Antibody | R&D Systems | ABAF497 | 1 : 500 |
| POMC Rabbit mAb | Cell Signaling Technology | D3R1U | 1 : 500 |
| PSD95 (D74D3) XP Rabbit mAb | Cell Signaling Technology | D74D3#3409 | 1 : 500 |
| Streptavidin, Alexa Fluor 594 conjugate | ThermoFisher | S11227 | 1 : 500 |
| Synapsin 1 Monoclonal Antibody (7H10G6) | ThermoFisher | MA5-31919 | 1 : 500 |
| Vectashield Plus Antifade Mountina Medium with DAPI 10 mL | Vector Laboratories | H-2000 |
Références
- Cone, R. D. Anatomy and regulation of the central melanocortin system. Nature Neuroscience. 8 (5), 571-578 (2005).
- Clarke, I. J. Hypothalamus as an endocrine organ. Comprehensive Physiology. 5 (1), 217-253 (2015).
- Mignot, E., Taheri, S., Nishino, S. Sleeping with the hypothalamus: emerging therapeutic targets for sleep disorders. Nature Neuroscience. 5 Suppl, 1071-1075 (2002).
- Baird, A. D., Wilson, S. J., Bladin, P. F., Saling, M. M., Reutens, D. C. Neurological control of human sexual behaviour: insights from lesion studies. Journal of Neurology, Neurosurgery, and Psychiatry. 78 (10), 1042-1049 (2007).
- Caria, A., Dall, O. G. Functional neuroimaging of human hypothalamus in socioemotional behavior: a systematic review. Brain Sciences. 12 (6), 707(2022).
- Andermann, M. L., Lowell, B. B. Toward a wiring diagram understanding of appetite control. Neuron. 95 (4), 757-778 (2017).
- Romanov, R. A., Alpar, A., Hokfelt, T., Harkany, T. Unified classification of molecular, network, and endocrine features of hypothalamic neurons. Annual Review of Neuroscience. 42, 1-26 (2019).
- Hajdarovic, K. H., Yu, D., Webb, A. E. Understanding the aging hypothalamus, one cell at a time. Trends in Neurosciences. 45 (12), 942-954 (2022).
- Zhang, Y. H., et al. Cascade diversification directs generation of neuronal diversity in the hypothalamus. Cell Stem Cell. 28 (8), 1483-1499 (2021).
- Chen, R., Wu, X., Jiang, L., Zhang, Y. Single-cell RNA-seq reveals hypothalamic cell diversity. Cell Reports. 18 (13), 3227-3241 (2017).
- Ma, C., et al. Neural pathways from hypothalamic orexin neurons to the ventrolateral preoptic area mediate sleep impairments induced by conditioned fear. Frontiers in Neuroscience. 17, 1122803(2023).
- Wang, F., et al. A parabrachial to hypothalamic pathway mediates defensive behavior. Elife. 12, e85450(2023).
- Cowley, M. A., et al. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature. 411 (6836), 480-484 (2001).
- Parekh, R. U., et al. Hypothalamic kinin B1 receptor mediates orexin system hyperactivity in neurogenic hypertension. Scientific Reports. 11 (1), 21050(2021).
- Schmidt, C. X., Tsang, A. H., Oster, H. Generation of mouse primary hypothalamic neuronal cultures for circadian bioluminescence assays. Bio-protocol. 11 (5), e3944(2021).
- Foo, L. C. Purification of rat and mouse astrocytes by immunopanning. 2013 (5), Cold Spring Harbor Protocols. 421-432 (2013).
- Emery, B., Dugas, J. C. Purification of oligodendrocyte lineage cells from mouse cortices by immunopanning. 2013 (9), Cold Spring Harbor Protocols. 854-868 (2013).
- Zhao, H., et al. Changes of constituents and activity to apoptosis and cell cycle during fermentation of tea. International Journal of Molecular Sciences. 12 (3), 1862-1875 (2011).
- Zhang, Z. M., et al. Down-regulation of human leukocyte antigens class I on peripheral T lymphocytes and NK cells from subjects in region of high-incidence gastrointestinal tumor. Chinese Medical Journal. 124 (12), 1813-1817 (2011).
- Drake, S. S., Zaman, A., Simas, T., Fournier, A. E. Comparing RNA-sequencing datasets from astrocytes, oligodendrocytes, and microglia in multiple sclerosis identifies novel dysregulated genes relevant to inflammation and myelination. WIREs Mechanisms of Disease. 15 (2), e1594(2023).
- Mattanovich, D., Borth, N. Applications of cell sorting in biotechnology. Microbial Cell Factories. 5, 12(2006).
- Cottrell, E. C., et al. Developmental changes in hypothalamic leptin receptor: relationship with the postnatal leptin surge and energy balance neuropeptides in the postnatal rat. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 296 (3), R631-R639 (2009).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.