A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
מחקרים של אינטראקציות מלווה-Cochaperone באמצעות אסאי הומוגני מבוסס חרוזים
In This Article
Summary
פרוטוקול זה מציג טכניקה לבדיקת אינטראקציות חלבון-חלבון באמצעות חרוזי תורם הקשורים גלוטתיון עם מלווים שותפים TPR-מוטיב מותך GST וחרוזים מקובלים יחד עם פפטיד שמקורו Hsp90. השתמשנו בטכניקה זו כדי לסנן מולקולות קטנות כדי לשבש Hsp90-FKBP51 או Hsp90-FKBP52 אינטראקציות וזיהינו מעכבי אינטראקציה Hsp90-FKBP51 חזקים וסלקטיביים.
Abstract
מיקוד חלבון הלם חום 90 (Hsp90)-cochaperone אינטראקציות מספק את האפשרות לווסת באופן ספציפי תהליכים תאיים תלוי Hsp90. המחומש המשומר MEEVD בטרמינל C של Hsp90 אחראי לאינטראקציה עם מוטיב חוזר טטרטריקופטיד (TPR) של מלווים משותפים. FK506-מחייב חלבון (FKBP) 51 ו FKBP52 הם שני מלווים שותפים TPR-מוטיב דומה המעורבים במחלות תלויות הורמונים סטרואידים עם פונקציות שונות. לכן, זיהוי מולקולות במיוחד חסימת אינטראקציות בין Hsp90 ו FKBP51 או FKBP52 מספק פוטנציאל טיפולי מבטיח עבור מספר מחלות אנושיות. כאן, אנו מתארים את הפרוטוקול עבור קרבה זוהרת מוגברת כדי לחקור אינטראקציות בין Hsp90 ושותפו מלווים FKBP51 ו- FKBP52. ראשית, טיהרנו את החלבונים המכילים מוטיב TPR FKBP51 ו- FKBP52 בצורת גלוטתיון S-transferase (GST). באמצעות חרוזי התורם הקשורים לגלוטתיון עם חלבוני TPR-מוטיב מותכים GST וחרוזים מקבלים יחד עם פפטיד מסוף C 10-mer של Hsp90, חקרנו אינטראקציות חלבון חלבון בסביבה הומוגנית. השתמשנו בבדיקה זו כדי לסנן מולקולות קטנות כדי לשבש את האינטראקציות Hsp90-FKBP51 או Hsp90-FKBP52 וזיהינו מעכבי אינטראקציה חזקים וסלקטיביים Hsp90-FKBP51.
Introduction
מלווים מולקולריים תורמים להומאוסטזיס של חלבונים, כולל קיפול חלבונים, הובלה והשפלה. הם מווסתים מספר תהליכים תאיים ומקושרים למחלות רבות כגון סרטן ומחלות ניווניות1. חלבון הלם חום 90 (Hsp90) הוא אחד המלווים החשובים ביותר שתפקידם תלוי בשינויים קונפורמיטיביים המונעים על ידי הידרוליזה ATP וקשירה עם חלבוני לקוח בתיווך המלווים השותפים שלה2. למרות הפוטנציאל הברור של Hsp90 כיעד הטיפולי, כוונון עדין של תפקידו מהווה אתגר גדול. ישנם מספר מעכבי Hsp90 המתמקדים באזור איגוד ה- ATP של N-terminal, אשר הוערכו בניסויים קליניים, אך אף אחד מהם לא אושר לשיווק3. בשל היעדר כיס4, המתמקדבאזור ה-C-terminal של Hsp90, זכה להצלחה מוגבלת4. לאחרונה, הפרעה של אינטראקציות Hsp90-cochaperone על ידי מולקולות קטנות נחקרה כאסטרטגיהחלופית 5. מיקוד אינטראקציות Hsp90-cochaperone לא לעורר תגובת מתח תא כללי ומספק את האפשרות לווסת באופן ספציפי תהליכים תאיים שונים. המחומש המשומר MEEVD בטרמינל C של Hsp90 אחראי לאינטראקציה עם מוטיב חוזר טטראטריאופפטייד (TPR) של מלווים משותפים6. מתוך 736 חלבונים המכילים מוטיב TPR המובאים במאגר החלבון האנושי, ~ 20 חלבונים שונים אינטראקציה עם Hsp90 באמצעות פפטידזה 7. מולקולות המתחרות על כריכת הפפטיד של MEEVD ישבשו את האינטראקציות בין Hsp90 למלווים שותפים המכילים תחום TPR. אתר כריכת הפפטידים בעל מבנה שלישוני דומה, אך ההומולוגיה הכוללת בין תחומי מוטיב TPR שונים נמוכה יחסית7, ומספקת הזדמנות לזהות מולקולות המסוגלות במיוחד לחסום אינטראקציות בין Hsp90 לבין מלווים משותפים מסוימים של TPR-מוטיב. בין אלה TPR-מוטיב מלווים משותפים, FK506 מחייב חלבון (FKBP) 51 ו FKBP52 הם רגולטורים של קולטן הורמון סטרואידים (SHR) איתות מעורב במספר מחלות תלויות הורמונים סטרואידים כולל סרטן, מחלות הקשורות ללחץ, מחלות מטבוליות, ומחלתאלצהיימר 8. למרות FKBP51 ו- FKBP52 מניות > דמיון רצף 80%, הפונקציות שלהם שונים: FKBP52 הוא רגולטור חיובי של פעילות SHR, בעוד FKBP51 הוא רגולטור שלילי ברוב המקרים8. לכן, זיהוי מולקולות, במיוחד חסימת אינטראקציות בין Hsp90 ו- FKBP51 או FKBP52, מספק פוטנציאל טיפולי מבטיח למחלות קשורות.
Luminescent Proximity Hאומוגנוס Assay (אלפא מסך) פותח לראשונה בשנת 1994 על ידי אולמן EF ואח'9. עכשיו הוא נמצא בשימוש נרחב כדי לזהות סוגים שונים של אינטראקציות ביולוגיות, כגון פפטיד10, חלבון11, DNA12,RNA13, וסוכר14. בטכניקה זו, ישנם שני סוגים של חרוזים (קוטר 200 ננומטר), אחד הוא חרוז התורם והשני הוא חרוז הקבלה. הביומולקולים משותקים על החרוזים האלה; האינטראקציות הביולוגיות שלהם מביאות את החרוזים התורמים והמקבלים לקרבה. ב 680 ננומטר, רגישות בצלם בחרוז התורם מאירה וממירה חמצן לחמצן יחיד. מכיוון שלחמצן הסינגל יש חיים קצרים, הוא יכול לפזר רק עד 200 ננומטר. אם חרוז הקבלה נמצא בסמיכות, נגזרת תיוקסן שלה מגיבה עם חמצן יחיד יצירת chemiluminescence ב 370 ננומטר. אנרגיה זו מפעילה עוד יותר פלואורופור באותו חרוז מקבל לפלוט אור ב 520-620 ננומטר15. אם האינטראקציות הביולוגיות משובשות, החרוז המקבל וחרוז התורם אינם יכולים להגיע לקרבה, וכתוצאה מכך דעיכה בחמצן יחיד ואות המיוצר נמוך.
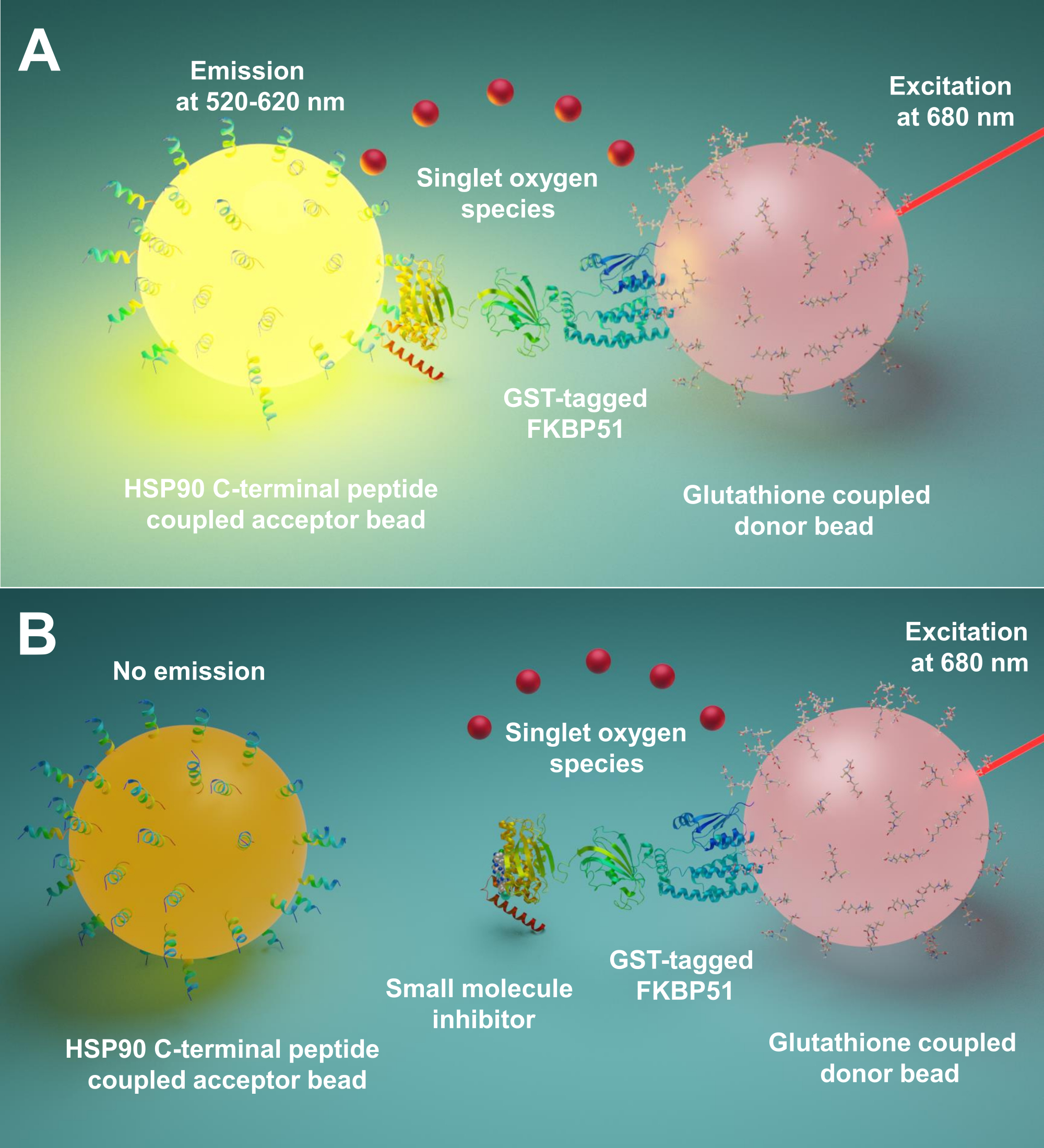
כאן אנו מתארים פרוטוקול באמצעות טכניקה זו להקרנת מולקולות קטנות המעכבות אינטראקציות בין Hsp90 ל- TPR מלווים משותפים, במיוחד FKBP51 ו- FKBP52. 10 פפטידים ארוכים של חומצות אמינו המתאימים ל- Hsp90 C-terminus קיצוני מחוברים לחרוזים מקובלים. מלווים שותפים TPR מתויגים GST אינטראקציה עם חרוזי תורם הקשורים גלוטתיון. כאשר האינטראקציה בין פפטידים שמקורם ב-Hsp90 לבין מלווים שותפים של TPR-מוטיב מאחדת את החרוזים, נוצר אות מוגבר(איור 1A). אם המולקולות הקטנות המוקרנות יכולות לעכב את האינטראקציות בין Hsp90 ל-TPR-מוטיב שותפים, האות המוגבר הזה יקטן (איור 1B). ניתן לחשב את IC50 שלהם לפי מדידה כמותית. פרוטוקול זה יכול להיות מורחב לכל מלווה - אינטראקציות שיתוף המלווה TPR-מוטיב של עניין והוא בעל חשיבות רבה בפיתוח של מולקולות חדשניות, במיוחד חסימת האינטראקציה בין Hsp90 ו- FKBP51 או FKBP52.

איור 1: העיקרון הבסיסי של ההסתה הזו. (A)GST-FKBP51 מטוהר מקיים אינטראקציה עם חרוזי תורמים הקשורים לגלוטתיון. 10 פפטידים ארוכים של חומצת אמינו המתאימים לטרמינל C הקיצוני של Hsp90 מחוברים לחרוזים מקובלים. האינטראקציה בין פפטידים שמקורם ב- Hsp90 לבין תחום TPR של FKBP51 מביאה את החרוזים התורמים והמקבלים לקרבה. ב 680 ננומטר, רגישות בצלם בחרוז התורם מאירה וממירה חמצן לחמצן יחיד. נגזרת תיוקסן על חרוז הקבלה מגיבה עם חמצן יחיד ומייצר chemiluminescence ב 370 ננומטר. אנרגיה זו מפעילה עוד יותר פלואורופור באותו חרוז קבלה לפלוט אור ב 520-620 ננומטר. (B)כאשר מולקולות קטנות מעכבות את האינטראקציות בין Hsp90 ל- FKBP51, החרוזים התורמים והמקבלים אינם יכולים להגיע לקרבה. ואז החמצן היחיד עם ריקבון תוחלת חיים קצרה, ולא מיוצר אות לגילוי. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Protocol
הערה: מבט כולל על פרוטוקול זה מוצג באיור 2.
1. ביטוי וטיהור של GST-FKBP51 ו- GST-FKBP52 (איור 2A)
-
פלסמידים
הערה: להשיג שיבוטים cDNA עבור FKBP51 אנושי (מזהה שיבוט: 5723416) עבור FKBP52 אנושי (מזהה שיבוט: 7474554) מקונסורציום IMAGE.- להגביר את ה- DNA FKBP51 האנושי על ידי PCR עם פריימרים (קדימה; 5'GGATCCATGAACTACTACTGATGAAGGT-3', הפוך; 5'-CTCGAGCTATGCTTCTCTCCAC-3') המכילים אוברהי באמי ו- XhoI ושיבוט לווקטור pGEX6-1 באתרי ההגבלה של באמהי / XhoI.
- להגביר את ה- DNA האנושי FKBP52 על ידי PCR עם פריימרים (קדימה; 5'-GAATTCATGACAGCCGAGGAGATG-3', הפוך; 5'-CTCGAGCTATATGCTTCTCTCTCCAC-3') המכיל אוקרי ו XhoI overhangs ושיבוט לתוך וקטור pGEX6-2 באתרי הגבלה EcoRI / XhoI.
הערה: הגדרת תגובת PCR והתנאים מוצגים בטבלה 1 ובטבלה 2. - לאמת את הרצף שהוכנס ולהפוך את plasmids לתוך E. coli מוסמך כימית על פי פרוטוקול הייצור.
-
ביטוי וטיהור חלבונים
- הוסיפו 25 גרם לבסיס מרק לוריא (LB) ב-1 ליטר מים מזוקקים כדי להפוך את פתרון ה-LB. תבודד אותו אוטומטית ב-121 מעלות צלזיוס למשך 15 דקות. לאחר הקירור, מוסיפים אמפיצ'ילין של 50 מיקרוגרם/מ"ל.
- קח מושבה של חיידקים המבטאות GST-FKBP51 או GST-FKBP52 ולערבב עם 500 μL של פתרון LB בצינור 1.5 מ"ל. מערבולת.
- מוסיפים את התערובת של "1.2.2" לתוך 1 L של פתרון LB בבקבוק Erlenmeyer מכוסה רדיד אלומיניום. לדגור על בקבוקון ארלנמייר בשייקר לילה ב 37 °C (50 °F).
- לגרום ביטוי חלבון על ידי הוספת 1 mM איזופרופיל-β-D-thiogalactoside (IPTG) לבקבוק ארלנמאייר ולהמשיך את הדגירה במשך 2 שעות נוספות.
- כדי לקבל כדורי תא, צנטריפוגה ב 5,000 x g במשך 15 דקות. הסר את סופר-טבעי.
הערה: כדורי התא ניתן לאחסן ב -20 °C (70 °F). - resuspend כדורי התא ב 40 מ"ל של PBS ו sonicate 3 x 20 s על קרח. יש להוסיף 1 מ"מ PMSF, 1 mM EDTA וקוקטייל מעכב פרוטאז (טבליה אחת) כדי למנוע פרוטאוליזיס.
- צנטריפוגה ההשעיה במשך 30 דקות 50,000 x g כדי להסיר פסולת תא ולהחיל את supernatant על 5 מ"ל GST מלכודת לעמודה.
- לאחר שטיפת העמוד עם 30 מ"ל PBS, יש לברוח GST-FKBP51 ו- GST-FKBP52 עם 5 מ"ל של גלוטתיון 10 מ"מ ב- PBS.
- מרכז חלבונים ביחידת צנטריפוגה של 15 מ"ל 10,000 MWCO. כדי להסיר גלוטתיון בחינם, מעבירים תרכיזים דרך עמודת PD-10 משוויבים עם PBS 0.5x ושוב מתרכזים בהתקן הצנטריפוגה של המסנן.
- לאסוף שברים המכילים חלבון. אמת את החלבונים ב-SDS-PAGE והתאם את ריכוזי החלבון ל-1 מ"ג/מ"ל.
הערה: תפוקת חלבון טיפוסית היא 2-5 מ"ג / L תרבית. ניתן לאחסן את החלבון ב-20 מעלות צלזיוס.
2. צימוד של Hsp90 C-terminal פפטיד לחרוזים המקובלים (איור 2B)
-
הכנת פפטיד Hsp90
- לסנתז עשר חומצות אמינו פפטיד NH2-EDASRMEEVD-COOH המתאים לחומצות אמינו 714-724 של איזופורם בטא Hsp90 אנושי (מזהה UniProt: P08238) על ידי שירות סינתזת פפטיד.
- לדלל את הפפטיד Hsp90 ב- PBS לריכוז של 1 מ"ג/מ"ל.
-
הכנת חרוזי קבלה
- לדלל את חרוזי הקבלה ללא מנוול PBS לריכוז 1 מ"ג / מ"ל ולהעביר לצינור 1.5 מ"ל.
- בצע את הכביסה על ידי צנטריפוגה ב 16,000 x g במשך 15 דקות. הסר בזהירות את סופר-טבעי.
-
הטיות
- הגדר את היחס בין חרוזים ופפטיד כ- 10:1. בצינור 1.5 מ"ל המכיל 1 מ"ג של כדור חרוזים מקובל (מוכן כפי שתואר לעיל), להוסיף 1 מ"ל של PBS (pH 7.4), 0.1 מ"ג של פפטיד מדולל, 1.25 μL של Tween-20, 10 μL של פתרון 400 mM של נתרן cyanoborohydride (NaBH3CN) במים.
זהירות: NaBH3CN רעיל; השתמש ברדס אדים וכפפות. פתרון NaBH3CN צריך להיות מוכן טרי. - דגירה במשך 24 שעות ב 37 °C (37 °F) עם תסיסה סוף סוף (10-20 סל"ד) על שייקר סיבובי.
- הגדר את היחס בין חרוזים ופפטיד כ- 10:1. בצינור 1.5 מ"ל המכיל 1 מ"ג של כדור חרוזים מקובל (מוכן כפי שתואר לעיל), להוסיף 1 מ"ל של PBS (pH 7.4), 0.1 מ"ג של פפטיד מדולל, 1.25 μL של Tween-20, 10 μL של פתרון 400 mM של נתרן cyanoborohydride (NaBH3CN) במים.
-
מרווה תגובה ושטיפת חרוזים
- הוסף 20 μL של פתרון 1 M Tris-HCl (pH 8.0) לתגובה לחסימת אתרים שלא הוחזרו. דגירה במשך 1 שעות ב 37 °C (50 °F).
- צנטריפוגה ב 16,000 x g (או מהירות מקסימלית) במשך 15 דקות ב 4 °C (70 °F). הסר את supernatant ו resuspend את גלולה חרוזים 1 מ"ל של פתרון Tris-HCl (100 mM, pH 8.0).
- חזור על שלב הכביסה שלוש פעמים.
- לאחר הצנטריפוגה האחרונה, resuspend החרוזים ב 1 מ"ג / מ"ל במאגר אחסון (1 מ"ל של 0.5 × PBS עם 0.01% נתרן אזיד כחומר משמר). יש לאחסן את פתרון חרוז הקבלה המצומד ב-4 °C (4 °F) מוגן.
זהירות: נתרן אזיד רעיל; השתמש ברדס אדים וכפפות.
3. ההבחנה בוחנת את האינטראקציה בין GST-FKBP51 או GST-FKBP52 ו- Hsp90 C-terminal פפטיד, ועיכוב עם תרכובות מסה מולקולרית קטנות(איור 2C)
-
חלבונים מתויגים GST אינטראקציה עם חרוזי תורם גלוטתיון
- הגדר את התגובות בצלחות 384-well.
- הכן את הפתרון המכיל 10 מיקרוגרם / מ"ל של חרוזי תורם גלוטתיון ב 0.5x PBS, pH 7.4.
הערה: לאחר אחסון ממושך, החרוזים מתיישבים וצריכים להיות מערבולת. - הוסף GST-FKBP51 או GST-FKBP52 לריכוז סופי של 10 מיקרוגרם /מ"ל.
- דגירה בחושך ב 25 °C (5 °F) במשך 10 דקות.
הערה: בשלב זה, חלבונים מתויגים GST אינטראקציה עם גלוטתיון מחובר חרוזים. עבור כל באר, 22.5 μL של תערובת זו ישמש. ריכוז השותפים הכרוכות חייב להיקבע אמפירית. Titrate GST-FKBP51 ו- GST-FKBP52 ולבחור את הריכוז שנותן את האות הטוב ביותר.
-
תוספת מורכבת
- הפוך דילול טורי של תרכובות בדיקה ב- DMSO.
הערה: הריכוזים המשמשים הם בדרך כלל 10, 30, 100, 300, 1,000 ו- 3,000 מיקרומטר. - הוסף 0.25 μL של DMSO (שליטה שלילית) או Hsp90 C-מסוף פפטיד (שליטה חיובית, 30 מיקרומטר) או תרכובות ב- DMSO לפינה של כל באר של הצלחת. יש להשתמש בטריפליקאטים לכל ריכוז מורכב.
- הוסיפו 22.5 μL של הפתרון המכיל חרוזי תורם גלוטתיון עם חלבונים מתויגים GST לכל באר.
- לנער את הצלחת בעדינות עם היד אבל ביסודיות. דגירה בחושך ב 25 °C (5 °F) במשך 15 דקות.
הערה: במהלך תקופה זו, תרכובות יקיימו אינטראקציה עם תחום TPR באתר איגוד הפפטיד Hsp90 C-terminal.
- הפוך דילול טורי של תרכובות בדיקה ב- DMSO.
-
תוספת חרוזי קבלה
- לדלל את החרוזים הקבלים עם פפטיד Hsp90 C-מסוף מצורף ל 100 מיקרוגרם / מ"ל ב 0.5x PBS.
- הוסף 2.25 μL של חרוזי קבלה מדוללים לכל באר.
- מערבבים בעדינות אך ביסודיות. דגירה בחושך ב 25 °C (5 °F) במשך 15 דקות.
הערה: בשלב זה, חרוזים תורמים ומקבלים מובאים לקרבה על ידי אינטראקציות חלבון-פפטיד. הנפח הסופי של תערובת התגובה הוא 25 μL. לכן, הריכוזים הסופיים של תרכובות נע בין 0.1 ל 30 מיקרומטר.
-
קריאת לוחיות רישוי
הערה: קרא את הצלחת באמצעות קורא לוחות שהוגדר במצב הרלוונטי.- הפעל את המכשיר ופתח את התוכנה
- בחר את הפרוטוקול הרלוונטי.
- לחץ על ערוך מפת לוח ובחר את הרווחה המשמשת בלוח לצורך מדידה.
- לחץ על הבא כדי להמשיך ולהפעיל את הפרוטוקול הנבחר.
- לאחר המדידה, לחץ על הצג תוצאות כדי להציג תוצאות.
- יצא את הנתונים.
4. ניתוח נתונים
-
יחס גורם Z ויחס אות לרקע (S/B)
- חשב את יחס ה- Z של גורם ו- S/B עבור ה- assay באמצעות המשוואה הבאה:
Z'=1-(3σpos+3σשלילי)/│μpos-μשלילי│16
S/B=μשלילי/קופת μ
כאשר, σ ו- μ מייצגים את סטיות התקן והאמצעים של הפקדים החיוביים (Hsp90 C-terminal, 30 מיקרומטר) ושלילי (DMSO), בהתאמה. גורם Z > 0.5 יבטיח כי בדיקת ה- Assay חזקה מספיק להקרנה. כדי לעקוב אחר רגישות ה- Assay, גם יחס ה- S/B חושב.
- חשב את יחס ה- Z של גורם ו- S/B עבור ה- assay באמצעות המשוואה הבאה:
-
עקומת תגובת מינון ו-IC 50
הערה: השתמש בניתוח רגרסיה לא ליניארית כדי להתאים לנתונים של מעכבי PPI Hsp90-cochaperones על ידי תוכנה.- צרו טבלת נתונים XY בתיבת הדו-שיח 'ברוכים הבאים' ובחרו 'מספריX' ו- Y Enter 3 (אם טריפליקאטים) משכפלים ערכים בעמודות זו לצד זו.
- נרמל את נתוני האות של דגימות לקבוצת הביקורת השלילית. יבא ערכי ריכוז לעמודת X וערכי האות לעמודה Y.
- לחץ על נתח ובחר ריכוז שינוי צורה (X) תחת שינוי צורה | לנרמל. בחר המר ללוגריתמים.
הערה: פעולה זו תהפוך את הריכוז לקנה מידה של יומן רישום. אם ריכוז ההתחלה שלך הוא אפס, הגדר אותו למספר קטן מאוד שהוא למעשה אפס (למשל, 0.1 nM) כדי לא לאבד ערכים אלה מכיוון שהלוגריתם של אפס אינו מוגדר. - לחץ על נתח ובחר רגרסיה לא ליניארית (התאמת עקומה) תחת ניתוחי XY, פתח את המינון-תגובה-עיכוב ובחר Log(מעכב) לעומת תגובה - שיפוע משתנה.
- לחץ על אישור כדי להציג את התוצאות (המכילות ערך IC 50) וגרפים.

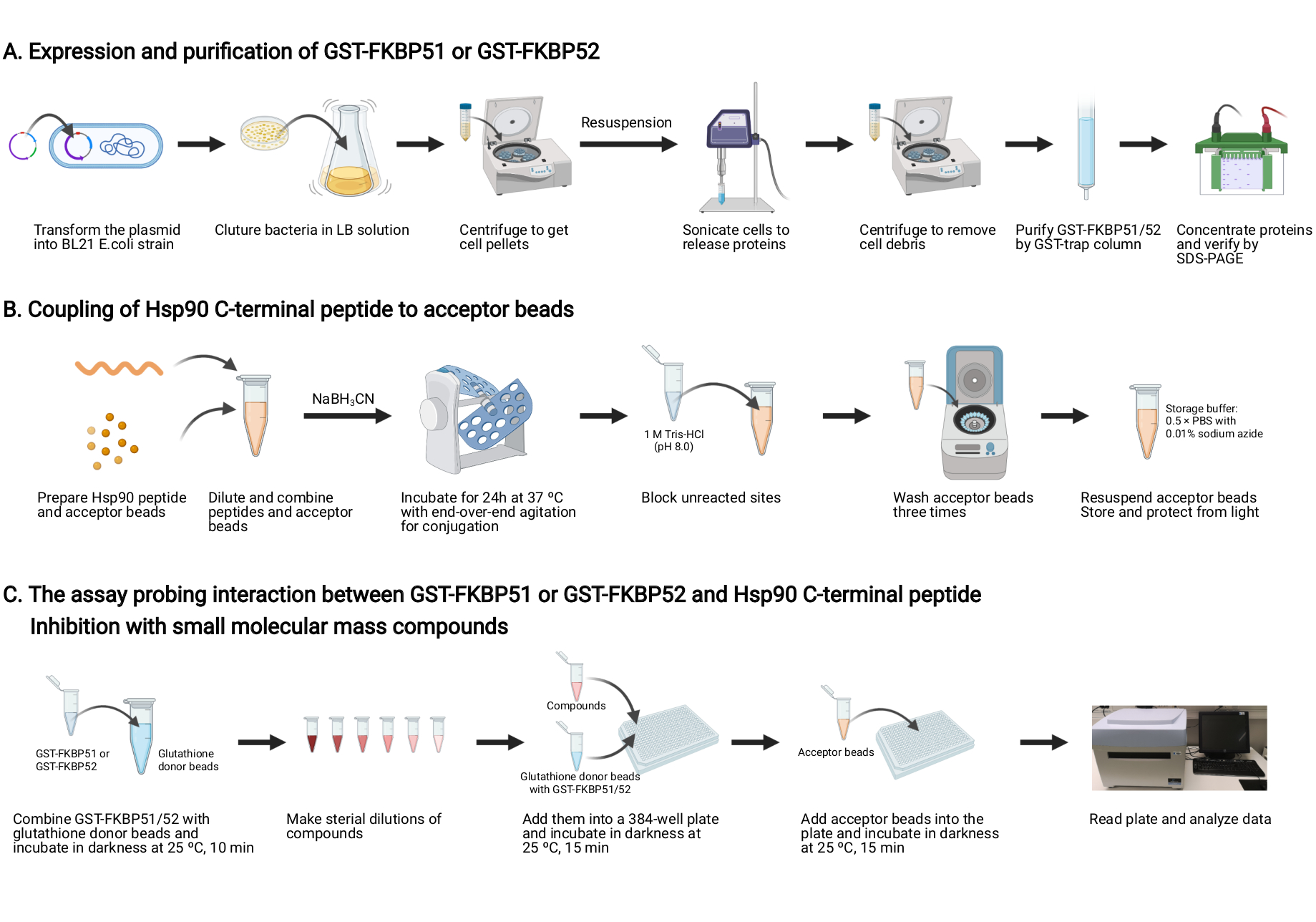
איור 2: שרטוט של פרוטוקול זה. (A)ביטוי וטיהור של GST-FKBP51 ו- GST-FKBP52. (B)צימוד של פפטיד Hsp90 C-מסוף לחרוזים המקובלים. (C)ההסתה בוחנת את האינטראקציה בין GST-FKBP51 או GST-FKBP52 ו- Hsp90 C-terminal פפטיד. עיכוב עם תרכובות מסה מולקולרית קטנות. נוצר עם BioRender.com אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
תוצאות
בבדיקה שלנו, יחס ה-Z וה-S/B הם 0.82 ו-13.35, בהתאמה (איור 3A),מה שמוכיח שבדיקת ה-Assay שלנו חזקה ואמינה להקרנה בתפוקה גבוהה. לאחר מכן השתמשנו בו כדי לסנן תרכובות מסה מולקולרית קטנות. איור 3B מציג עיכוב תלוי מינון של אינטראקציות קואצ'אפרון מלווה עם מולקולה קטנה שנבחרה (D10). ?...
Discussion
כאן אנו מתארים פרוטוקול באמצעות בדיקה לבדיקה עבור סינון מולקולות קטנות מעכב אינטראקציות בין Hsp90 ו TPR-מוטיב מלווים משותפים, במיוחד FKBP51 ו- FKBP52. ציון ה-Z הגבוה שלו (>0.8) מדגים את החוסן והאמינות לפורמט בעל תפוקה גבוהה. התוצאות ניתן להשיג בתוך שעה אחת, כמויות קטנות של חרוזים, חלבון ותרכובות נדרשים. ?...
Disclosures
המחברים לא מדווחים על ניגודי אינטרסים.
Acknowledgements
מחקר זה נתמך על ידי מענקים ממועצת המחקר השוודית (2018-02843), קרן המוח (Fo 2019-0140), הקרן למחלות גריאטריות במכון קרולינסקה, קרן גונוור ויוסף אנר, קרן מגנוס ברגוולס, קרן אקדח וברטל סטונס, קרן טורה נילסון למחקר רפואי, קרן מרגרטה af Ugglas והקרן למשרתים זקנים.
Materials
| Name | Company | Catalog Number | Comments |
| 384-well plates | Perkin Elmer | 6008350 | Assay volume 25 ml |
| Amicon 10.000 MWCO centrifugation unit | Millipore | UFC901008 | Concentrate protein |
| Ampicillin | Sigma | A0166 | Antibiotics |
| Bacteria shaker Unimax 1010 | Heidolph | Culture bacteria | |
| cDNA clones for human FKBP51 | Source BioScience | clone id: 5723416 | pCMV-SPORT6 vector |
| cDNA clones for human FKBP52 | Source BioScience | clone id: 7474554 | pCMV-SPORT6 vector |
| Chemically Competent E. coli | Invitrogen | C602003 | One Shot BL21 Star (DE3) |
| Data analysis software | GraphPad Prism | 9.0.0 | Analysis data and make figures |
| Data analysis software | Excel | Analysis data | |
| DMSO | Supelco | 1.02952.1000 | Dilute compounds |
| DPBS | Gibco | 14190-144 | Prepare solution |
| EDTA | Calbiochem | 344504 | Prevent proteolysis during sonication |
| Glutathione | Sigma | G-4251 | Elute GST-tagged proteins |
| Glutathione donor beads | Perkin Elmer | 6765300 | Donor bead |
| GST-trap column | Cytiva (GE Healthcare) | 17528201 | Purify GST-tagged proteins |
| Isopropyl-β-D-thiogalactoside | Thermo Fisher Scientific | R0392 | Induce protein expression |
| LB Broth (Miller) | Sigma | L3522 | Microbial growth medium |
| PCR instrument | BIO-RAD | S1000 Thermal Cycler | Amplification/PCR |
| PD-10 column | Cytiva (GE Healthcare) | 17085101 | Solution exchange |
| pGEX-6P-1 vector | Cytiva (GE Healthcare) | 28954648 | Plasmid |
| pGEX-6P-2 vector | Cytiva (GE Healthcare) | 28954650 | Plasmid |
| Plate reader | Perkin Elmer | EnSpire 2300 Multilabel Reader | Read alpha plate |
| Plate reader software | Perkin Elmer | EnSpire Manager | Plate reader software |
| Plate reader software protocol | Perkin Elmer | Alpha 384-well Low volume | Use this protocol to read plate |
| PMSF | Sigma | P7626 | Prevent proteolysis during sonication |
| protease inhibitor cocktail | Sigma | S8830 | Prevent proteolysis during sonication |
| Sodium azide | Sigma | S2002 | As a preservative |
| Sodium cyanoborohydride (NaBH3CN) | Sigma | 156159 | Activates matrix for coupling |
| Ten amino acid peptide NH2-EDASRMEEVD-COOH corresponding to amino acids 714-724 of human Hsp90 beta isoform | Peptide 2.0 inc | Synthesize Hsp90 C-terminal peptide | |
| Test-Tube Rotator | LABINCO | Make end-over-end agitation | |
| Tris-HCl | Sigma | 10708976001 | Block unreacted sites of acceptor beads |
| Tween-20 | Sigma | P1379 | Prevent beads aggregation |
| Ultra centrifuge Avanti J-20 XP | Beckman Coulter | Centrifuge to get bacteria cell pellets | |
| Ultrasonic cell disruptor | Microson | Sonicate cells to release protein | |
| Unconjugated acceptor beads | Perkin Elmer | 6762003 | Acceptor beads |
| XCell SureLock Mini-Cell and XCell II Blot Module | Invitrogen | EI0002 | SDS-PAGE |
References
- Muchowski, P. J., Wacker, J. L. Modulation of neurodegeneration by molecular chaperones. Nature Reviews Neuroscience. 6 (1), 11-22 (2005).
- Eckl, J. M., Richter, K. Functions of the Hsp90 chaperone system: lifting client proteins to new heights. International Journal of Biochemistry and Molecular Biology. 4 (4), 157-165 (2013).
- Yuno, A. Clinical evaluation and biomarker profiling of Hsp90 inhibitors. Methods in Molecular Biology. 1709, 426-441 (2018).
- Dutta Gupta, S., Bommaka, M. K., Banerjee, A. Inhibiting protein-protein interactions of Hsp90 as a novel approach for targeting cancer. European Journal of Medicinal Chemistry. 178, 48-63 (2019).
- Pavlov, P. F., Hutter-Paier, B., Havas, D., Windisch, M., Winblad, B. Development of GMP-1 a molecular chaperone network modulator protecting mitochondrial function and its assessment in fly and mice models of Alzheimer's disease. Journal of Cellular and Molecular Medicine. 22 (7), 3464-3474 (2018).
- Young, J. C., Obermann, W. M., Hartl, F. U. Specific binding of tetratricopeptide repeat proteins to the C-terminal 12-kDa domain of hsp90. Journal of Biological Chemistry. 273 (29), 18007-18010 (1998).
- Scheufler, C., et al. Structure of TPR domain-peptide complexes: critical elements in the assembly of the Hsp70-Hsp90 multichaperone machine. Cell. 101 (2), 199-210 (2000).
- Storer, C. L., Dickey, C. A., Galigniana, M. D., Rein, T., Cox, M. B. FKBP51 and FKBP52 in signaling and disease. Trends in Endocrinology & Metabolism. 22 (12), 481-490 (2011).
- Ullman, E. F., et al. Luminescent oxygen channeling immunoassay: measurement of particle binding kinetics by chemiluminescence. Proceedings of the National Academy of Sciences of the United States of America. 91 (12), 5426-5430 (1994).
- Wigle, T. J., et al. Screening for inhibitors of low-affinity epigenetic peptide-protein interactions: an AlphaScreen-based assay for antagonists of methyl-lysine binding proteins. Journal of Biomolecular Screening. 15 (1), 62-71 (2010).
- Guenat, S., et al. Homogeneous and nonradioactive high-throughput screening platform for the characterization of kinase inhibitors in cell lysates. Journal of Biomolecular Screening. 11 (8), 1015-1026 (2006).
- Sabatucci, A., et al. A new methodological approach for in vitro determination of the role of DNA methylation on transcription factor binding using AlphaScreen(R) analysis: Focus on CREB1 binding at hBDNF promoter IV. Journal of Neuroscience Methods. 341, 108720 (2020).
- Mills, N. L., Shelat, A. A., Guy, R. K. Assay Optimization and Screening of RNA-Protein Interactions by AlphaScreen. Journal of Biomolecular Screening. 12 (7), 946-955 (2007).
- Huang, X., et al. A competitive alphascreen assay for detection of hyaluronan. Glycobiology. 28 (3), 137-147 (2018).
- Principles of alphascreen amplified luinescent proximmity homogenous assay. PerkinElmer Life Sciences Available from: https://www.perkinelmer.com/lab-solutions/resources/docs/APP_AlphaScreen_Principles.pdf (2021)
- Zhang, J. H., Chung, T. D., Oldenburg, K. R. A Simple statistical parameter for use in evaluation and validation of high throughput screening assays. Journal of Biomolecular Screening. 4 (2), 67-73 (1999).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved