
Method Article
Quantificazione delle modificazioni post-traduzionali degli istoni globali utilizzando la citometria a flusso intranucleare nella microglia cerebrale di topo isolata
In questo articolo
Riepilogo
Questo lavoro descrive un protocollo per la quantificazione delle modificazioni istoniche globali utilizzando la citometria a flusso intranucleare in microglia cerebrali isolate. Il lavoro contiene anche il protocollo di isolamento della microglia che è stato utilizzato per la raccolta dei dati.
Abstract
Il controllo dell'espressione genica avviene in parte attraverso modifiche nella struttura della cromatina, inclusa l'aggiunta e la rimozione di modifiche post-traduzionali alle code istoniche. Le modificazioni post-traduzionali degli istoni (HPTM) possono facilitare l'espressione genica o la repressione. Ad esempio, l'acetilazione dei residui di lisina della coda istonica neutralizza la carica positiva e riduce le interazioni tra la coda e il DNA caricato negativamente. La diminuzione delle interazioni coda istonica-DNA si traduce in una maggiore accessibilità del DNA sottostante, consentendo un maggiore accesso al fattore di trascrizione. Il segno di acetilazione funge anche da sito di riconoscimento per gli attivatori trascrizionali contenenti bromodominio, che insieme si traducono in una maggiore espressione genica. I marcatori istonici possono essere regolati dinamicamente durante la differenziazione cellulare e in risposta a diversi ambienti e stimoli cellulari. Mentre gli approcci di sequenziamento di nuova generazione hanno iniziato a caratterizzare le posizioni genomiche per le singole modificazioni istoniche, solo una modifica può essere esaminata contemporaneamente. Dato che esistono centinaia di HPTM diversi, abbiamo sviluppato una misura quantitativa ad alto rendimento di HPTM globali che può essere utilizzata per lo screening delle modifiche istoniche prima di condurre approcci di sequenziamento del genoma più estesi. Questo protocollo descrive un metodo basato sulla citometria a flusso per rilevare HPTM globali e può essere condotto utilizzando cellule in coltura o cellule isolate da tessuti in vivo . Presentiamo dati di esempio da microglia cerebrali di topo isolate per dimostrare la sensibilità del test per rilevare cambiamenti globali negli HPTM in risposta a uno stimolo immunitario derivato da batteri (lipopolisaccaride). Questo protocollo consente la valutazione rapida e quantitativa degli HPTM e può essere applicato a qualsiasi regolatore trascrizionale o epigenetico che possa essere rilevato da un anticorpo.
Introduzione
L'epigenetica è lo studio dei meccanismi che regolano l'espressione genica senza alterare la sequenza di DNA sottostante. La regolazione epigenetica dell'espressione genica è dinamica all'interno delle cellule e può consentire risposte rapide e coordinate a vari stimoli ambientali. La regolazione dinamica avviene in parte a causa di cambiamenti nella struttura della cromatina a livello del nucleosoma, che è composto da proteine istoniche (H2A, H2B, H3, H4) assemblate in un nucleo di ottamero strettamente avvolto dal DNA1. Le interazioni tra le proteine istoniche e il DNA possono controllare l'accessibilità del DNA al meccanismo di trascrizione, che può in ultima analisi controllare l'espressione genica e altri aspetti della biologia della cromatina2. Le proteine istoniche hanno code non strutturate che presentano residui caricati positivamente che formano interazioni elettrostatiche con la spina dorsale del DNA caricata negativamente. Queste interazioni si traducono in un impacchettamento stretto del DNA e in una ridotta accessibilità del DNA. Le modificazioni covalenti delle code istoniche, denominate modificazioni post-traduzionali istoniche (HPTM), possono regolare queste interazioni 3,4. Alcuni degli HPTM più ben caratterizzati includono l'acetilazione e la metilazione della coda istonica, che può modificare l'affinità delle interazioni elettrostatiche tra le code istoniche e il DNA, con conseguente accessibilità differenziale al DNA sottostante e reclutamento di fattori di trascrizione che riconoscono questi HPTM in siti specifici. Gli HPTM sono regolati da tre importanti classi di enzimi denominati lettori, che riconoscono, scrittori, che depositano, e cancellatori, che rimuovono gli HPTM. Pertanto, il reclutamento o la dissoluzione di enzimi lettore, scrittore o cancellatore può in ultima analisi cambiare il panorama degli HPTM e governare la struttura e la funzione della cromatina, rendendo la loro regolazione e lettura essenziali per la comprensione della biologia cellulare e della funzione 3,4.
Le cellule del sistema nervoso centrale (SNC) sono epigeneticamente flessibili in quanto modificano il loro trascrittoma per adattarsi agli stimoli ambientali. Sempre più evidenze suggeriscono che i cambiamenti nell'epigenoma, come la metilazione del DNA, gli RNA non codificanti e gli HPTM, svolgono un ruolo essenziale nella formazione della memoria e nella funzione sinaptica5. L'interruzione delle dinamiche HPTM attraverso la manipolazione dei lettori, degli scrittori o dei cancellatori pertinenti può bloccare o migliorare l'apprendimento associativo e il potenziamento a lungo termine 6,7,8. La microglia, la cellula immunitaria residente del sistema nervoso centrale, regola rapidamente il loro trascrittoma in risposta alla stimolazione immunitaria attraverso cambiamenti dinamici nel loro epigenoma 9,10,11. Questo alto livello di adattamento all'ambiente cerebrale locale li rende difficili da esaminare in un contesto isolato, poiché gli studi hanno dimostrato che l'epigenoma e il trascrittoma della microglia vengono alterati dopo solo poche ore nei terreni di coltura dopo la rimozione dall'ambiente cerebrale11. Inoltre, poiché le microglia costituiscono solo il 10% delle cellule cerebrali, le misure che esaminano i cambiamenti a livello dell'intero tessuto mancano di sensibilità e specificità12,13. Di conseguenza, le microglia devono essere rapidamente isolate per esaminare i cambiamenti epigenetici come i livelli di HPTM, ex vivo.
I metodi comunemente utilizzati per esaminare gli HPTM includono il sequenziamento della cromatina-immunoprecipitazione (ChIP-seq) e il clivaggio sotto i bersagli e il sequenziamento di tagmentazione (CUT&Tag-seq)4. Sebbene queste tecniche siano altamente specifiche per un singolo HPTM e possano informare sulla presenza di HPTM all'interno di uno specifico contesto genomico, possono esaminare solo uno dei molti HPTM possibili all'interno di un singolo esperimento11,14 Pertanto, prima di procedere con tali esperimenti, che richiedono un investimento significativo in termini di tempo e denaro, è molto utile restringere l'elenco degli HPTM potenzialmente interessanti per ulteriori indagini, esaminando prima i cambiamenti nella globalizzazione livelli di HPTM. I due approcci principali per esaminare i livelli globali di HPTM sono l'immunoistochimica e l'analisi western blot, ma entrambi gli approcci sono solo semi-quantitativi, a basso rendimento e richiedono un gran numero di sezioni di tessuto o cellule isolate15,16. Pertanto, abbiamo mirato a sviluppare un metodo quantitativo altamente sensibile che potesse essere utilizzato per esaminare i livelli globali di HPTM rapidamente e a livello di singola cellula.
Il protocollo presentato consente di rilevare rapidamente i livelli globali di HPTM utilizzando la citometria a flusso intranucleare. Precedenti studi sulle cellule tumorali hanno giustificato l'importanza di esaminare i livelli globali da una prospettiva clinica17,18. È anche comune che gli studi utilizzino i livelli globali come metodo di screening prima di valutare la posizione genomica di specifici HPTM di interesse19,20. Per la microglia, la valutazione dei livelli globali dopo l'isolamento è difficile a causa della bassa resa cellulare; Pan et al. presentano livelli globali di HPTM da microglia isolate, in cui la microglia di tre animali è stata raggruppata per consentire il rilevamento del livello proteico mediante western blot19. Utilizzando il nostro protocollo, siamo in grado di rilevare i cambiamenti globali con input cellulari molto più bassi, consentendo lo screening di più marcature per animale ed eliminando la necessità di raggruppare i campioni.
In questo articolo, descriviamo un protocollo per rilevare rapidamente i livelli di HPTM tramite citometria a flusso intranucleare quantitativa in microglia isolate. Anche se per brevità ci concentriamo specificamente sulla quantificazione HPTM, questo protocollo può essere utilizzato allo stesso modo per quantificare i livelli globali di enzimi lettore, scrittore e cancellatore. Il protocollo è suddiviso in due parti: in primo luogo, il metodo di isolamento per la microglia e, in secondo luogo, il metodo basato sulla citometria a flusso per determinare i livelli di HPTM. Il metodo di isolamento produce cellule che possono essere utilizzate sia per l'isolamento dell'RNA che per la valutazione del livello di HPTM, che consente di valutare l'espressione genica e i livelli di HPTM dallo stesso campione. Inoltre, il metodo per la valutazione HPTM può essere utilizzato su altri tipi di cellule come indicato nel protocollo.
Protocollo
Tutti i protocolli per la cura degli animali sono stati approvati dal Comitato per la cura degli animali dell'Università della British Columbia in conformità con le linee guida del Consiglio canadese per la cura degli animali.
1. Digestione cerebrale per l'isolamento della microglia

Figura 1: Diagramma di flusso semplice del protocollo. I topi vengono prima perfusi per via transcardiaca con HBSS e il cervello viene sezionato. Il cervello viene quindi dissociato attraverso la digestione chimica e l'interruzione meccanica per ottenere un omogenato a singola cellula. La frazione arricchita immunitaria viene raccolta tramite gradiente di densità discontinuo, dopodiché le cellule vengono colorate per P2RY12. Le cellule colorate sono 1) ordinate tramite la selezione cellulare attivata fluorescente (FACS) per portare all'analisi dell'RNA o all'analisi delle proteine a valle e/o 2) fissate, permeabilizzate e colorate per le proteine intranucleari. Il livello proteico è quantificato in base all'intensità mediana della fluorescenza nel canale di interesse determinata dalla citometria a flusso. Le caselle colorate in blu fanno parte della fase 1 del protocollo) Digestione cerebrale per l'isolamento della microglia. I riquadri colorati in rosso fanno parte della fase 2 del protocollo) Colorazione a flusso intranucleare per l'analisi dell'espressione proteica. Creato con BioRender.com. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
- Preparazione dei reagenti
NOTA: Se si pianifica un'estrazione per raccogliere sia l'RNA che le cellule per l'analisi HPTM, fare riferimento alla sezione 1.7.1 per le modifiche che includono gli inibitori della trascrizione e della traduzione. Tuttavia, questo non è necessario se si valuta solo il segnale proteico, poiché le cellule sono in gran parte quiescenti quando vengono mantenute sul ghiaccio.- Tampone FACS (Fluorescence activated Cell Sorting) (20 mL per campione): sciogliere l'albumina sierica bovina (BSA) in 1x soluzione salina bilanciata Hanks (HBSS) per creare una soluzione BSA al 2%. Sciogliere l'EDTA ad una concentrazione finale di 1 mM nella soluzione di BSA al 2%. Filtrare: sterilizzare con un filtro da 0,2 μm e conservare a 4 °C per un massimo di 1 settimana prima dell'uso.
- Tampone di digestione (1 mL per campione): Ricostituire un flaconcino di papaina in HBSS a una concentrazione finale di 20 U/mL in 1 mM di L-cisteina con 0,5 mM di EDTA. Attivare a 37 °C per un minimo di 10 minuti o fino al momento di digerire il tessuto. Poco prima dell'uso, aggiungere DNasi I alla soluzione di papaina attivata fino a una concentrazione finale di 200 U/mL. Prepararlo il giorno dell'esperimento e non conservarlo.
- Soluzione isotonica a gradiente di densità (5,5 mL per campione): Aggiungere 10x HBSS al mezzo a gradiente di densità fredda fino a una concentrazione finale di 1x HBSS, ottenendo una densità finale di 1,117 g/mL. Frullare per almeno 30 secondi prima dell'uso. Mettere sul ghiaccio fino all'uso.
- Soluzione con gradiente di densità al 37% (4 mL per campione): aggiungere il gradiente di densità isotonica a 1x HBSS per ottenere una concentrazione finale del 37% con una densità finale di 1,043 g/mL. Aggiungere 20 μL di rosso fenolo per ogni mL di gradiente di densità del 37% per creare una soluzione rosa per la visualizzazione durante la stratificazione. Vortice per almeno 30 s prima dell'uso. Mettere sul ghiaccio fino all'uso.
- Soluzione con gradiente di densità al 70% (2 mL per campione): Aggiungere il gradiente di densità isotonico a 1x HBSS per ottenere una concentrazione finale del 70% con una densità finale di 1,082 g/mL. Aggiungere 5 μL di blu tripano per ogni mL di terreno con densità al 70% per ottenere una soluzione blu per la visualizzazione durante la stratificazione. Vortice per almeno 30 s prima dell'uso. Mettere sul ghiaccio fino all'uso.
- Perfusione e dissezione cerebrale
NOTA: Il protocollo di perfusione è simile a quello di Posel et al. che presenta una rappresentazione video della toracotomia del topo, della perfusione transcardica e della rimozione del cervello21. In questo caso, utilizziamo topi adulti C57BL/6J maschi e femmine (10-15 settimane, 20-30 g), ma questo protocollo può essere utilizzato per eseguire una toracotomia per qualsiasi topo. Tutte le procedure sugli animali devono essere approvate dal comitato etico istituzionale prima di condurre gli esperimenti.- Anestesia del topo: anestetizzare i topi con il 4% di isoflurano in ossigeno al 100% fino a oltre il piano dell'anestesia chirurgica, che può essere confermato con un pizzicamento del dito del piede o una mancanza di riflesso dopo aver pizzicato saldamente il piede del topo. Posiziona il topo sulla schiena e fissa saldamente le sue quattro zampe nella tavola di dissezione chirurgica posta inclinata in un vassoio di plastica, assicurandoti che il naso sia fissato al cono nasale in isofluorano. Dopo il trasferimento, assicurarsi che l'animale abbia ancora superato il piano chirurgico dell'anestesia prima di procedere.
- Toracotamia del topo: afferrare e sollevare la pelle addominale usando una pinza e praticare un'incisione poco profonda attraverso la pelle e la parete addominale per esporre lo xifoide senza danneggiare l'aorta discendente o gli organi sottostanti.
- Afferrare lo xifoide con una pinza e praticare incisioni laterali sotto la gabbia toracica per esporre il diaframma e il fegato. Eseguire tagli accurati e poco profondi attraverso il diaframma lungo la lunghezza della gabbia toracica usando forbici sottili e attraverso la gabbia toracica usando forbici per tessuti e fissare lo sterno alla stazione chirurgica vicino alla testa del topo per esporre il cuore e i polmoni per la perfusione transcardiaca.
- Perfusione transcardica: preparare una pompa peristaltica e collegare un ago da 26,5 G a un'estremità del tubo. Adescare il tubo per la procedura inserendo un'estremità del tubo in una fiala di 1x HBSS freddo e accendendo la pompa per riempire completamente il tubo con 1x HBSS.
- Mentre si tiene il cuore con una pinza smussata, inserire la punta di un ago da 26,5 G con il tubo di perfusione attaccato nel ventricolo sinistro del cuore e praticare una piccola incisione nell'atrio destro. Accendere la pompa di perfusione per perfondere con cura il topo a una velocità di ~2-4 mL/min con almeno 15-20 mL di HBSS 1x freddo.
NOTA: Una perfusione completa è spesso indicata quando il fegato inizia a pulire il sangue e diventa dello stesso colore del cuore. - Rimozione del cervello: decapitare il topo usando forbici da dissezione dei tessuti e praticare un'incisione sulla linea mediana del cuoio capelluto dal collo al naso. Staccare i lembi di pelle ai lati per esporre il cranio e rimuovere il tessuto e le ossa in eccesso all'estremità caudale del cranio con forbici da dissezione.
- Fai scorrere con cautela una lama delle forbici sotto il cranio nel forame magno con il lato affilato rivolto verso l'osso e taglia con cura la linea mediana verso il naso. Fai dei tagli laterali sia alla base del cranio che vicino al naso usando le forbici da dissezione. Usando una pinza sottile, fai vivere il cranio dalla linea mediana verso l'esterno per rompere i pezzi del cranio ed esporre il cervello. Sollevare delicatamente il cervello con una spatola e posizionarlo su carta assorbente.
- Dissezione cerebrale: posizionare il cervello su un pezzo di carta assorbente per dissezione bagnata con 1x HBSS sopra una capsula di Petri chiusa riempita di ghiaccio. Rimuovere il cervelletto e dividere in due gli emisferi cerebrali usando una lametta pulita.
- Rimuovi il tronco encefalico, lo striato e la materia bianca da ciascun emisfero, mantenendo intatto l'ippocampo e la corteccia sovrastante. Trasferire gli emisferi contenenti corteccia isolata e tessuto ippocampale in una provetta da 15 mL con 5 mL di HBSS 1x freddo e mantenere il ghiaccio.
NOTA: È importante eseguire le dissezioni il più rapidamente possibile in modo che il tessuto rimanga freddo con non più di 2 minuti tra la decapitazione e il posizionamento finale del tessuto sezionato in 1x HBSS su ghiaccio. Se si isolano le microglia da più animali, i cervelli possono essere conservati su ghiaccio in 1x HBSS per ~ 1 ora prima di procedere con l'elaborazione dell'intera coorte di animali per la digestione, ecc.
- Digestione e omogeneizzazione del cervello
- Dissociazione meccanica e chimica: Mettere il tessuto cerebrale di ciascun topo e 1 ml di tampone di digestione in singole piastre di Petri su ghiaccio. Usando una lama di bisturi pulita, tagliare accuratamente il cervello in piccoli pezzi (<1 mm).
- Tagliare la punta da una pipetta di trasferimento di plastica e trasferire con cura ciascuno dei cervelli tritati in pozzetti separati all'interno di una piastra da 24 pozzetti su ghiaccio. Coprire la piastra con pellicola flessibile trasparente e incubare su ghiaccio per 30 min.
NOTA: Se tritato correttamente, il tessuto cerebrale assomiglia all'aglio ben tritato. - Omogeneizzazione a spruzzo: Trasferire la soluzione cerebrale digerita da ciascun pozzetto in singoli omogeneizzatori in vetro da 7 mL su ghiaccio, ciascuno riempito con 5 ml di tampone FACS freddo. Far sbucare delicatamente ogni cervello con il pestello sciolto (A), circa 30-40 volte, fino ad ottenere una sospensione unicellulare. Dopo aver schiacciato con il pestello A, inzuppare delicatamente con il pestello stretto (B) 3-4 volte per garantire una sospensione a cellula singola.
NOTA: Non spingere il pestello per più di 3/4 verso il basso per evitare di schiacciare il tessuto sul fondo dell'omogeneizzatore. La soluzione finale dovrebbe essere opaca e lattiginosa.
NOTA: Se si digeriscono più cervelli in un singolo esperimento, cronometrare il trasferimento del cervello nel tampone FACS in modo che ogni campione rimanga nel tampone di digestione solo per 30 minuti. Una digestione eccessiva può provocare la scissione delle proteine di superficie, riducendo il legame e il segnale degli anticorpi a valle.
- Ottenimento di un frammento arricchito di immunità
- Stabilire il gradiente di densità: trasferire l'omogenato da ciascun cervello in provette di polipropilene separate da 15 mL e aggiungere 2,125 mL di gradiente di densità isotonica e completare a 8,5 mL con tampone FACS per ciascuno per ottenere una concentrazione finale del gradiente di densità del 25%. Capovolgere delicatamente le provette da 15 ml 20 volte per mescolare accuratamente.
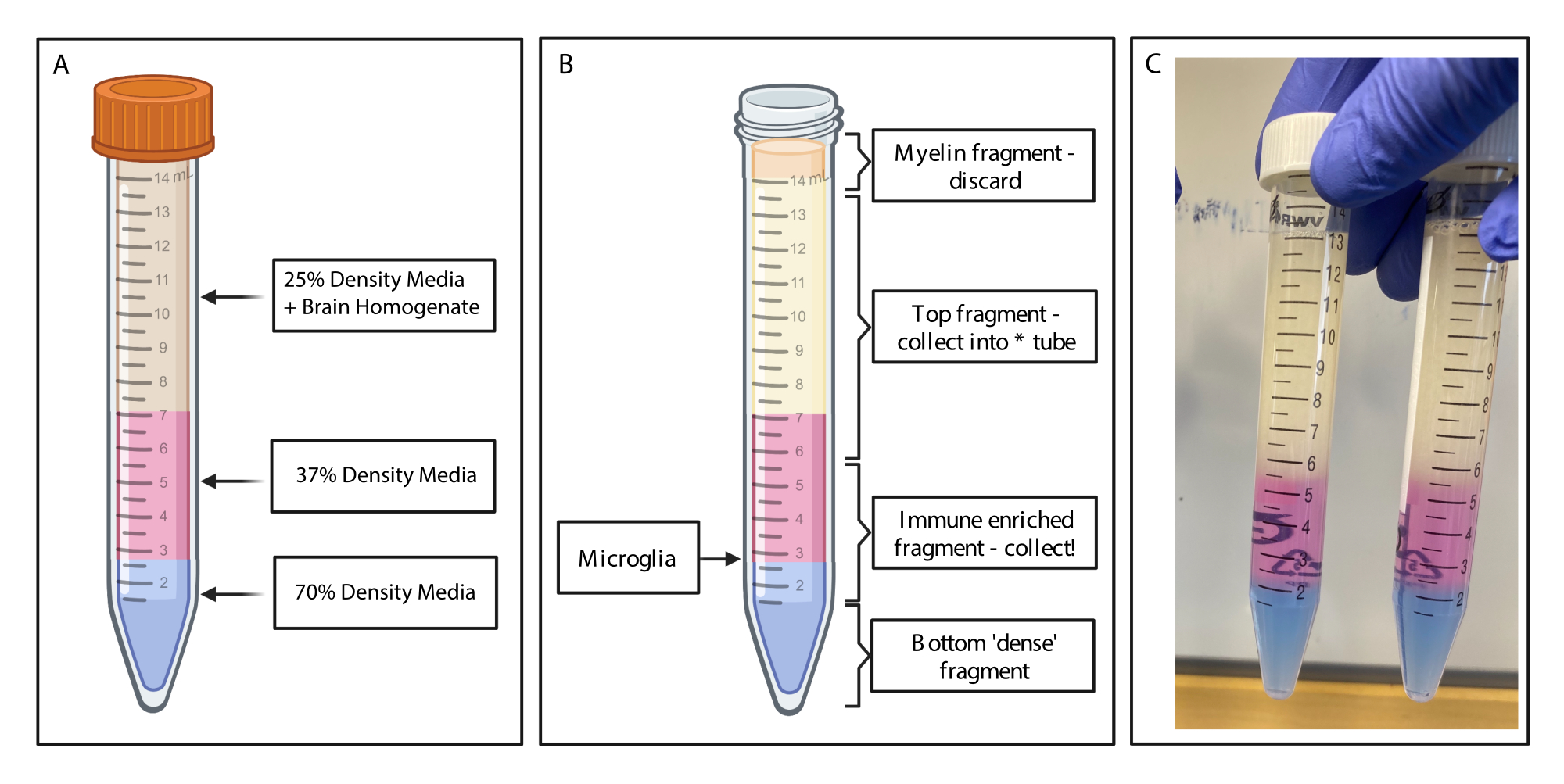
- Utilizzando una pipetta di trasferimento a graduazione stretta, sottoporre delicatamente 4 mL di gradiente di densità del 37% a ciascuna provetta, facendo molta attenzione a stabilire strati puliti. Cambiare le pipette di trasferimento e sottoporre delicatamente 2 mL di gradiente di densità al 70% (Figura 2A). Trasferire in una centrifuga raffreddata a 4 °C e centrifugare a 500 x g per 20 minuti con la rampa di frenatura impostata a zero.
- Raccolta di frammenti immunoarricchiti: utilizzando pipette di trasferimento pulite, aspirare delicatamente la mielina dalla parte superiore del volume nella provetta da 15 ml utilizzando una pipetta di trasferimento pulita ed eliminare. Raccogliere con cautela il frammento superiore del gradiente di densità in una provetta di polipropilene pulita da 15 mL utilizzando una pipetta di trasferimento.
- Raccogliere con cura il frammento arricchito di immunità (1,5 mL sopra e 1,5 mL sotto il punto in cui si incontrano gli strati di gradiente di densità del 70% e del 37%) in una nuova provetta di polipropilene da 15 mL (Figura 2B). Aggiungere 10 mL di tampone FACS al campione immunoarricchito per diluire il mezzo a gradiente di densità e capovolgere delicatamente la provetta 20 volte per mescolare accuratamente.
NOTA: Poiché le cellule tendono ad attaccarsi ai lati della provetta, assicurarsi di raccogliere tutte le cellule del campione durante le fasi di raccolta facendo girare lentamente la pipetta lungo i lati della provetta durante la raccolta del liquido. - Pellettare le cellule nel campione immunoarricchito centrifugando le provette da 15 mL in una centrifuga a 4 °C a 500 x g per 10 minuti con il freno della rampa in discesa impostato a zero. Subito dopo la fine della centrifuga, rimuovere con cura il surnatante, lasciando circa 300 μL di liquido nella provetta da 15 mL, facendo attenzione a non disturbare il pellet (che potrebbe non essere visibile).
- Raccogliere il surnatante in un'altra provetta da 15 mL per assicurarsi che le cellule siano state pellettate nello spin (scartare questa frazione una volta effettuata la verifica con la conta cellulare del pellet risospeso). Dopo aver risospeso il pellet cellulare nel volume di 300 μL utilizzando una pipetta P1000, contare le cellule con un ematotometro per stimare la resa cellulare totale.

Figura 2: Ottenimento del frammento arricchito di immunità mediante gradiente di densità discontinuo. (A) L'omogenato cerebrale è realizzato al 25% di densità media, sottostante 4 mL di densità al 37% di colore rosa medio tramite rosso fenolo e 2 ml di densità 70% di colore blu medio tramite blu tripano. (B) Dopo la centrifugazione, le frazioni si sono separate. La microglia si trova all'interfaccia dei frammenti di terreno con densità del 37% e del 70%. Il frammento di mielina si trova nella parte superiore della provetta da 15 mL e verrà scartato. Il frammento superiore viene raccolto come backup nel caso in cui la rotazione non riesca e non viene recuperata alcuna cella. In tal caso, il gradiente può essere ripetuto utilizzando questa frazione. La frazione immunitaria arricchita viene raccolta a valle. La frazione inferiore contenente i globuli rossi rimane nella provetta e viene scartata. (C) Figura esemplificativa raffigurante strati completi. Creato con BioRender.com. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
- Colorazione degli anticorpi extracellulari
- Blocco: Trasferire le cellule su una piastra a 96 pozzetti a fondo tondo su ghiaccio e centrifugare a 500 x g con freno per pellettizzare le cellule. Rimuovere rapidamente il surnatante nel lavandino muovendo la piastra per smaltire il surnatante, lasciando intatto il pellet cellulare sul fondo del pozzetto.
- Risospendere le cellule in 50 μL di tampone FACS con reagente bloccante il recettore FC CD16/32 anti-topo utilizzando una pipetta P200 (concentrazione finale 10 μg/mL, fattore di diluizione 1:50) per prevenire il legame non specifico degli anticorpi ai monociti o ad altre cellule portatrici di FcR. Incubare per 10 minuti con ghiaccio.
- Colorazione degli anticorpi: Preparare il volume appropriato di una miscela master 2x contenente P2RY12- Alloficocianina (APC; fattore di diluizione 1:50, concentrazione 4 μg/mL per una concentrazione finale del pozzetto di 1:100, concentrazione 2 μg/mL) e colorante viola 525 vivo morto (fattore di diluizione 1:50 per una concentrazione finale del pozzetto di 1:100). Aggiungere 50 μL della miscela master colorante alla sospensione cellulare (ottenuta dopo il blocco di cui al punto 1.5.1) e incubare la piastra per 30 minuti al buio su ghiaccio.
NOTA: Per questo protocollo, presentiamo la colorazione delle cellule con P2RY12. In primo luogo, P2RY12 è un marcatore omeostatico per la microglia che può essere sottoregolato in determinati contesti patologici. Ad esempio, i topi modello di Alzheimer 5XFAD hanno abbassato i livelli di P2RY12, il che potrebbe renderli difficili da identificare22. I coloranti alternativi che possono essere utilizzati per l'isolamento includono Tmem119, Cd11b e CD4523. In secondo luogo, il fluorocromo coniugato APC può essere regolato per adattarsi al pannello di anticorpi desiderato. Tuttavia, la scelta di un fluorocromo brillante, come APC o PE, contribuirà a garantire che le popolazioni positive e negative siano facilmente distinguibili24. - Dopo la colorazione, aggiungere 200 μL di tampone FACS direttamente a ciascun pozzetto per lavare le cellule. Centrifugare a 500 x g a 4 °C per rimuovere il surnatante sfiorando. Risospendere le cellule in 200 μL di tampone FACS con una pipetta P200, centrifugare a 500 x g a 4 °C e sbattere la piastra per rimuovere il tampone dai pozzetti.
- Preparazione dei controlli di flusso: prima della colorazione, separare i volumi necessari di cellule da ciascun campione dopo il blocco al punto 1.5.1 per i controlli di flusso richiesti.
NOTA: I controlli di flusso sono necessari per ogni esperimento per stabilire le porte. I controlli di flusso possono essere prelevati da un animale aggiuntivo o da una frazione di ciascuno dei pozzi sperimentali. Quando si suddividono le celle, assicurarsi di assegnare un numero sufficiente di celle per controllo, poiché sono necessarie 10.000-30.000 celle per controllo per stabilire i gate con un'elevata confidenza.- Ci sono tre controlli di flusso rilevanti: nessuna macchia, morti vivi e controllo dell'isotipo P2RY12. Per il controllo antimacchia, non aggiungere alcun anticorpo. Nel controllo dell'isotipo P2RY12, trattare le cellule con un colorante di vitalità (1:100) e un anticorpo di controllo dell'isotipo coniugato ad APC (1:100).
- Per preparare il controllo dei morti vivi, aliquotare le cellule in un pozzetto separato e spostare metà del volume della cellula in una provetta da 500 μL. Mettere la provetta da 500 μL nel congelatore a -80 °C per 5 minuti, quindi metterla in un incubatore a 37 °C per 5 minuti per uccidere le cellule. Riportare l'aliquota delle cellule morte nel pozzetto di controllo dei morti vivi e colorare con un colorante di vitalità legante l'ammina su viola 525 (fattore di diluizione 1:100) per contrassegnare le cellule morte.
NOTA: Il protocollo è scritto per la colorazione delle piastre con un metodo flick per la rimozione del surnatante. Tuttavia, ciò richiede che il surnatante venga rimosso immediatamente dopo il completamento della rotazione e il colpo deve essere eseguito con forza sufficiente per rimuovere rapidamente il surnatante senza disturbare il pellet. In alternativa, per la colorazione è possibile utilizzare provette prive di RNAsi/DNasi da 1,5 mL, con le seguenti modifiche: Trasferire le cellule in una provetta per microcentrifuga da 1,5 mL e pellet a 800 x g per 5 minuti a 4 °C. Aspirare il surnatante con pipette. Suggerimento: Per una maggiore rapidità, una pipetta di trasferimento da 5 mL con puntale P200 è in grado di aspirare il surnatante in modo rapido e preciso. Durante l'aspirazione, controllare la presenza di pellet. Se il pellet non è visibile, lasciare 50 μL di surnatante e regolare i calcoli di conseguenza. Quando si eliminano gli anticorpi, aggiungere ulteriore FACS per aumentare la diluizione degli anticorpi (1000 μL invece di 200 μL) per tenere conto della rimozione incompleta del surnatante. A seconda del citometro, utilizzare le provette da 1,5 mL per lo smistamento, riducendo la quantità di forniture necessarie.
- Smistamento FACS per microglia
- Preparazione: Risospendere ogni pozzetto in 200 μL di tampone FACS con una pipetta P200 e trasferire in provette di selezione a flusso etichettate e aggiungere il tampone FACS per un totale di 500 μL per una concentrazione di circa 5 x 105 eventi per mL. Conservare su ghiaccio al buio fino all'analisi. Preparare le provette post-sort aggiungendo 100 μL di tampone FACS come cuscino per le cellule in provette prive di RNAsi da 1,5 mL.
- Impostazioni del citometro: Ordina le cellule su un selezionatore di cellule con citometria a flusso impostato con l'ugello da 100 μm. Ordina le celle usando 18-20 psi.
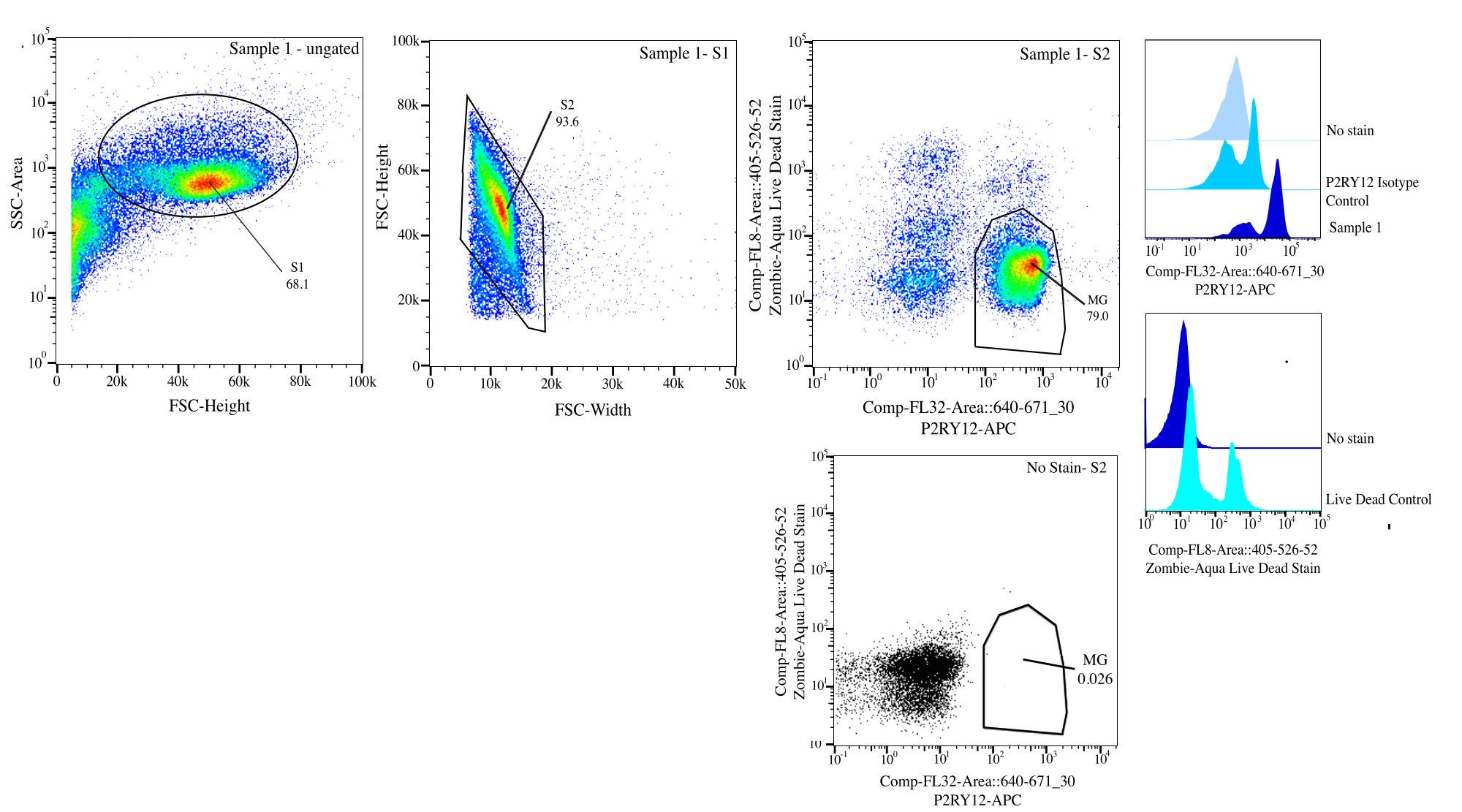
- Gating: sul citometro, gate per la dimensione della cellula utilizzando l'area side-scatter (SSC) rispetto all'altezza della diffusione diretta (FSC) utilizzando il controllo senza macchia per aiutare a distinguere i detriti, mettere SSC-A su un asse logaritmico per visualizzare una popolazione cellulare e gate da vicino per selezionare le cellule (gate S1; Figura 3). Per rimuovere eventuali doppiette, tracciare FSC-H vs FSC-W e gate strettamente attorno alla popolazione cellulare rimuovendo eventuali detriti e doppiette (gate S2). Utilizzando il controllo dell'isotipo P2RY12, esaminare le cellule nel canale APC e impostare il gate per l'autofluorescenza per determinare le cellule P2RY12+. Utilizzando i controlli senza macchia e morti vivi, cancello per le cellule che non sono fluorescenti su viola 525 nm come cellule vive.
- Ordinamento: Tracciare il colore viola 525 nm rispetto all'APC e determinare la popolazione che è P2RY12+ e vive in base agli FMO (MG). Ordinare tali cellule nella provetta di smistamento post-selezione etichettata (Figura 3). La percentuale di ordinamento finale è di circa il 50% degli eventi totali, con la maggior parte della perdita totale dell'evento dovuta alla rimozione dei detriti nel gate S1 (~70% degli eventi sono celle; Tabella 1).
- Isolamento e analisi dell'RNA
- Inibitori della trascrizione e della traduzione: se si pianifica l'estrazione dell'RNA, per eliminare il rischio di isolamento delle firme trascrittomiche associate, includere inibitori della traduzione e della trascrizione nelle fasi del tampone. Preparare il cocktail di inibitori come descritto da Marsh et al. che includa actinomicina D, anisomicina e triptolide25.
- Preparazione dell'inibitore: Ricostituire le scorte di inibitore e conservare come segue: Ricostituire l'actinomicina D in dimetilsolfossido (DMSO) a 5 mg/mL e conservare a -20 °C. Ricostituire il triptolide in DMSO a 10 mM e conservare a -20 °C, al riparo dalla luce. Ricostituire l'anisomicina in DMSO a 10 mg/mL e conservare a 4 °C, al riparo dalla luce. Conservare tutte le scorte di inibitori per non più di 1 mese dopo la ricostituzione.
- Modifiche del tampone: Aggiungere gli inibitori in quattro diversi tamponi nel protocollo come segue: Quando si esegue la perfusione transcardica, preparare HBSS con actinomicina D (5 μg/mL, 1:1000 da stock) e triptolide (10 μM, 1:1000 da stock). Dopo la perfusione, trasporta i cervelli in laboratorio in HBSS contenente actinomicina D (5 μg/mL, 1:1000 da magazzino), triptolide (10 μM, 1:1000 da magazzino) e anisomicina (27,1 μg/mL, 1:368,5 da magazzino). Preparare il tampone FACS con actinomicina D (5 μg/mL, 1:1000 da magazzino), triptolide (10 μM, 1:1000 da magazzino) e anisomicina (27,1 μg/mL, 1:368,5 da magazzino). Preparare il tampone di digestione con actinomicina D (5 μg/mL, 1:1000 da magazzino), triptolide (10 μM, 1:1000 da magazzino) e anisomicina (27,1 μg/mL, 1:368,5 da magazzino). Preparare il tampone di lavaggio post-selezione, con HBSS contenente actinomicina D (5 μg/mL, 1:1000 da magazzino), triptolide (10 μM, 1:1000 da magazzino) e anisomicina (27,1 μg/mL, 1:368,5 da magazzino).
NOTA: Quando si aggiungono gli inibitori, assicurarsi di aggiungerli immediatamente prima dell'uso e proteggere eventuali tamponi preparati dalla luce durante l'uso. Evitare il gelo-disgelo delle soluzioni madri.
- Lavaggi post-sorteggio: poiché le cellule sono state smistate in provette prive di RNasi da 1,5 mL in tampone FACS, che interferiscono con l'isolamento dell'RNA, è necessario lavare le cellule. Centrifugare le cellule a 1000 x g a 4 °C per 5 minuti e rimuovere il surnatante, lasciando circa 50 μL di liquido.
- Aggiungere 200 μL di 1x HBSS contenente actinomicina D (5 μg/mL, 1:1000 dalla cella), triptolide (10 μM, 1:1000 dalla riserva) e anisomicina (27,1 μg/mL, 1:368,5 dalla riserva) e mescolare accuratamente. Ripetere la centrifuga e rimuovere il surnatante lasciando 50 μL di liquido (lavaggio 1). Aggiungere 200 μL di tampone di lavaggio post-selezione, mescolare accuratamente e ripetere la centrifuga e rimuovere il surnatante lasciando 25 μL di liquido (lavaggio 2).
- Estrazione dell'RNA: per l'isolamento dell'RNA dalle cellule microgliali, utilizzare un kit di isolamento dell'RNA a basso input per rese di RNA elevate e coerenti e punteggi RIN superiori a 9 (vedere di seguito e la tabella dei materiali per le raccomandazioni sui prodotti). Al pellet cellulare, aggiungere 350 μL del tampone di lisi del kit consigliato + β-mercaptoetanolo (1:100) e mescolare bene.
NOTA: Se necessario, il protocollo può essere sospeso a questo punto. I campioni possono essere conservati nel tampone di lisi a -80 °C fino all'estrazione dell'RNA. Se si estrae l'RNA dopo la conservazione, scongelare il lisato su ghiaccio e procedere con le istruzioni specifiche del kit per l'isolamento. - Trasferire il lisato nel trituratore cellulare a colonna (vedere la tabella dei materiali per le raccomandazioni sui prodotti) e centrifugare alla massima velocità a 4 °C per 2 minuti. Eluire in un minimo di 14 μL di acqua priva di RNasi e determinare la concentrazione in base alle esigenze. L'RNA può essere utilizzato per qualsiasi applicazione a valle dopo questo punto.
- Inibitori della trascrizione e della traduzione: se si pianifica l'estrazione dell'RNA, per eliminare il rischio di isolamento delle firme trascrittomiche associate, includere inibitori della traduzione e della trascrizione nelle fasi del tampone. Preparare il cocktail di inibitori come descritto da Marsh et al. che includa actinomicina D, anisomicina e triptolide25.

Figura 3: Strategia di gating per l'ordinamento del flusso. Gli eventi sono controllati per le dimensioni delle celle su SSC-A rispetto a FSC-H (S1). Quindi, le cellule vengono controllate per essere singoletti su FSC-H rispetto a FSC-W (S2). Le cellule singoletto sono ordinate come vive se negative su Comp-FL8-A::405-526-52 (colorazione viola 525 viva morta) e come P2RY12+ se positive su Comp-FL32-A::640-671_30 (P2RY12-APC) utilizzando il controllo dell'isotipo P2RY12. Le cellule sono etichettate come MG e ordinate sia se vive che P2RY12+. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
| POPOLAZIONE RECINTATA | Frequenza del genitore | Frequenza del totale | Contare |
| Visualizzazione del materiale S1 | 68.10% | 68.10% | 162186 |
| S2>S1 | 93.59% | 63.70% | 151707 |
| P2Ry12+ (670+) > S2 > S1 | 83.05% | 52.90% | 125986 |
| Live (525-) > S2 > S1 | 92.78% | 59.10% | 140752 |
| MG (P2RY12+ Live) >S2>S1 | 78.96% | 50.30% | 119794 |
Tabella 1: Esempio di tabella di derivazione con percentuali di gating e numeri di eventi previsti.
2. Colorazione a flusso intranucleare per l'analisi dell'espressione proteica
NOTA: A questo punto è possibile iniziare altri tipi di cellule, questo protocollo viene testato con cellule in coltura, comprese le cellule HEK293, le cellule simili alla microglia BV2 e la microglia umana derivata da IPSC.
- Fissazione e colorazione delle cellule
NOTA: Per il seguente protocollo, utilizzare un kit di colorazione intracellulare ottimizzato per la colorazione nucleare. Vedere Tabella dei materiali per consigli sui prodotti.- Aliquote di cellule colorate extracellulari della sezione 1.5.2 in una piastra a 96 pozzetti (5 x 104- 1 x 106 cellule). Centrifugare le celle per 5 minuti a 500 x g a 4 °C e sfiorare per rimuovere il tampone FACS.
NOTA: Per ottenere dati con un'elevata confidenza dei livelli mediani, è necessario utilizzare un minimo di 10.000 cellule per pozzetto. Sebbene non esista un massimo raccomandato, è meglio mantenere costante il numero di cellule durante l'esperimento per garantire che non vi sia alcun effetto significativo di diversi coefficienti di variazione (CV). - Fissazione e permeabilizzazione: aggiungere 200 μL di concentrato fisso 1x e miscelare delicatamente con la pipetta P200 per risospendere le cellule. Incubare al buio per 45-60 min. Centrifugare la piastra per 5 minuti a 500 x g a temperatura ambiente (RT) e agitare per eliminare il surnatante.
NOTA: Se necessario, il protocollo può essere sospeso a questo punto. Dopo aver scartato il surnatante, sospendere nuovamente le cellule in un tampone di conservazione a lungo termine per le cellule immunitarie (vedere la tabella dei materiali per le raccomandazioni sui prodotti). I campioni possono essere conservati a 4 °C per 12-18 ore, protetti dalla luce e ricoperti da una pellicola trasparente per proteggere l'evaporazione del tampone. - Aggiungere 200 μL di tampone di permeabilizzazione 1x a ciascun pozzetto e pipettare con un P200 da miscelare. Centrifugare la piastra per 5 minuti a 500 x g a RT e agitare per eliminare il surnatante. Ripetere il lavaggio del tampone di permeabilizzazione per un totale di 3 volte.
- Preparazione dei controlli di flusso: suddividere il volume di cellule da ciascun campione per i controlli di flusso richiesti (sono sufficienti 10.000-30.000 cellule per pozzetto di controllo).
- Per preparare il controllo antimacchia, fissare le cellule antimacchia del tipo o dell'aliquota delle cellule non colorate in un pozzetto separato che non riceverà alcun anticorpo.
- Per preparare il controllo della fluorescenza meno uno (FMO), aliquote di cellule per ciascuno degli anticorpi sul pannello tranne quello in quel canale.
- Per i canali pertinenti, includere l'anticorpo di controllo dell'isotipo nell'FMO per il gating. Ad esempio, in un pannello contenente P2RY12-APC e H3K27Ac-AlexaFluor568, dovrebbero essere presenti due FMOS: (1) l'APC-FMO che contiene solo H3K27Ac-AlexaFluor568 e l'anticorpo di controllo dell'isotipo P2RY12 e (2) il 568-FMO che contiene solo P2RY12-APC e il controllo dell'isotipo primario e 568 secondario.
NOTA: Questo protocollo viene presentato per testare un singolo HPTM, tuttavia è possibile stabilire pannelli che contengono molti HPTM coniugati a diversi fluorofori.
- Colorazione con anticorpi primari: aggiungere 50 μL di tampone di permeabilizzazione 1x con la concentrazione appropriata di anticorpo primario in ciascun pozzetto. Incubare per 30 minuti a RT al buio. Lavare 2 volte con 200 μL di 1x tampone di permeabilizzazione.
NOTA: La concentrazione di anticorpi utilizzati per ciascun HPTM è inclusa nella Tabella dei materiali. La concentrazione viene determinata testando diverse concentrazioni degli anticorpi su cellule in coltura trattate con uno stimolante che causerebbe un aumento drammatico, ad esempio un inibitore HDAC per i segni di acetilazione e assicurandosi che sia le cellule non trattate che quelle trattate fossero ben all'interno dell'intervallo di rilevamento (al di sopra del controllo dell'isotipo e al di sotto dell'intervallo di rilevamento massimo del citometro). La concentrazione ottimale di anticorpi per gli HPTM dovrebbe avere un'intensità fluorescente mediana media nel canale del fluoroforo compresa tra 5 x 104 e 1 x 105. - Colorazione secondaria degli anticorpi: bloccare con 200 μL di tampone di permeabilizzazione 1x con siero d'asina normale (NDS) al 2% per 10 minuti a RT. Girare per 5 minuti a 500 x g a RT e agitare per rimuovere il surnatante.
- Aggiungere 50 μL di tampone di permeabilizzazione 1x con NDS al 2% e la concentrazione appropriata di anticorpo secondario e incubare per 30 minuti a RT al buio. Aggiungere 200 μL di tampone di permeabilizzazione 1x ai pozzetti da diluire, centrifugare la piastra per 5 minuti a 500 x g a RT e sbattere per scartare il surnatante. Celle di lavaggio 2x con 200 μL di 1x tampone di permeabilizzazione.
NOTA: Se necessario, sospendere il protocollo a questo punto. Risospendere le cellule in 200 μL di tampone di conservazione a lungo termine per cellule immunitarie con pipetta P200 (vedere la tabella dei materiali per le raccomandazioni) e conservare a 4 °C per 12-24 ore al riparo dalla luce. - Preparazione per la citometria a flusso: centrifugare la piastra per 5 minuti a 500 x g a RT e sbattere per eliminare il surnatante. Risospendere le cellule in 200 μL di tampone FACS utilizzando una pipetta P200 per la citometria a flusso. Sigillatura con pellicola trasparente per il trasporto al citometro.
- Aliquote di cellule colorate extracellulari della sezione 1.5.2 in una piastra a 96 pozzetti (5 x 104- 1 x 106 cellule). Centrifugare le celle per 5 minuti a 500 x g a 4 °C e sfiorare per rimuovere il tampone FACS.
- Citometria a flusso
- Per analizzare il pannello anticorpale proposto, assicurarsi che il citometro sia dotato di almeno quattro laser, tra cui viola (405 nm), blu (488 nm), giallo (561 nm) e rosso (633 nm). Il citometro necessita di filtri per rilevare FITC (blu-525 nm), KRO (viola-525 nm), PE (giallo-585 nm) e APC (rosso-660 nm). Aggiungere altri anticorpi a seconda del citometro scelto.
- Calibrazione e standardizzazione: all'inizio di ogni esperimento, eseguire le perle fluorescenti arcobaleno e regolare la tensione del tubo fotomoltiplicatore (PMT) fino a quando i picchi delle microsfere non sono paragonabili ai valori target eseguiti per gli esperimenti precedenti. Questo metodo di standardizzazione consente di adattarsi alla deriva delle apparecchiature nel tempo.
- Compensazione: dopo che la tensione e il guadagno PMT sono stati impostati per l'esperimento, utilizzare le perle di compensazione catturate dall'anticorpo per stabilire la matrice di compensazione per il pannello di anticorpi. Questo calcolo assicurerà che i fluorofori non contribuiscano alle variazioni di segnale in altri canali. Ciò è sempre più necessario quando si esegue il multiplexing di più anticorpi.
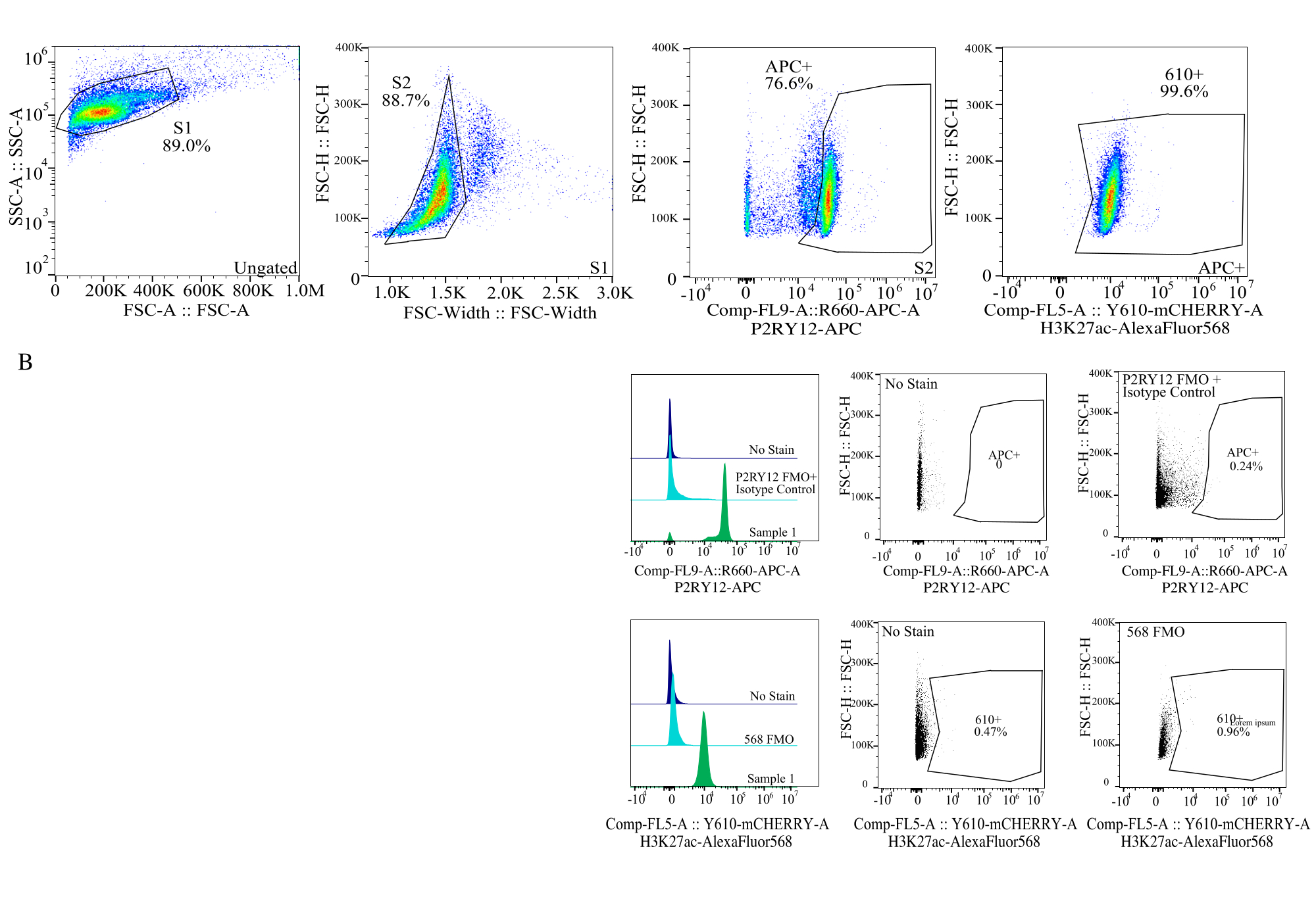
- Dimensione gating: in un dot plot, tracciare SSC-A su log rispetto a FSC-H su lineare. Escludere i detriti e selezionare la dimensione della cella utilizzando il gate S1. Selezionare per le celle singoletto in un grafico a punti di FSC-W rispetto a FSC-H e gate come S2. (Figura 4).
- Definizione delle porte del fluoroforo: utilizzando l'FMO pertinente per ciascun canale del fluoroforo, stabilire le porte per determinare qual è un segnale positivo in ciascun canale utilizzando istogrammi a parametro singolo (Figura 4).
- Misurazione dei campioni: registrare attentamente i campioni utilizzando la strategia di gating stabilita. Identificare la microglia utilizzando il segnale P2RY12+, determinare l'espressione della proteina nei rispettivi canali solo per la microglia.
- Analisi dei dati di citometria a flusso
- Stabilire le porte di analisi: utilizzando i passaggi precedenti per il citometro sull'interfaccia utente del software di analisi, utilizzare le stesse porte utilizzate per la registrazione per l'analisi.
- Ottenere i valori MFI utilizzando il software di analisi della citometria a flusso (vedere la tabella dei materiali per le raccomandazioni): ricapitolare la strategia di gating del citometro per l'analisi del flusso. Utilizzando la funzione di aggiunta di statistiche, selezionare la mediana per la popolazione di interesse (ad esempio, 568+) sull'altezza del canale compensata. Utilizzando l'editor di tabelle, esportare i valori mediani di intensità fluorescente (MFI) per i rispettivi canali in un foglio di calcolo per procedere con l'analisi statistica (Tabella 2).
NOTA: Il file supplementare S1 include dati di esempio da topi iniettati con lipopolisaccaridi (LPS) e soluzione salina tamponata con fosfato (PBS) e un file di analisi di esempio con la strategia di gating e i valori MFI. - Analisi dei valori di MFI per il cambiamento di ripiegamento della proteina: dopo aver ottenuto i valori di MFI, calcolare il cambiamento di ripiegamento dell'MFI rispetto alla popolazione di controllo o non trattata (Equazione 1). Il cambiamento di piega dell'MFI riflette il cambiamento di piega nei livelli di proteine. Utilizzando i valori di variazione della piega, valutare la variazione di espressione e calcolare la significatività statistica utilizzando un test t o ANOVA.
 Equazione 1
Equazione 1

Figura 4: Strategia di gating per la valutazione delle IFM proteiche. Gli eventi vengono controllati prima per le dimensioni delle celle su SSC-A rispetto a FSC-H (S1). Le cellule vengono quindi controllate per singoletti su FSC-H vs FSC-W (S2). Le cellule singoletto vengono quindi identificate come microglia dal segnale P2RY12-APC (APC+) con il gate stabilito in base alla fluorescenza in un controllo APC-FMO che contiene un anticorpo di controllo dell'isotipo. Le celle vengono quindi controllate per il segnale H3K27Ac-AlexaFluor568 su Comp-FL5-A::Y610-mCherry. L'intensità fluorescente delle cellule 610+ è determinata come proxy per l'espressione proteica. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Risultati
I topi adulti sono stati perfusi per via transcardiaca e sacrificati per l'isolamento della microglia. Le microglia sono state isolate su ghiaccio e colorate con anticorpi P2RY12-APC e viola 525 morti vivi. Le cellule che sono risultate positive per P2RY12 e negative per la colorazione morta viva viola 525 sono state classificate come microglia vive. La resa media di microglia da un cervello di topo sezionato è stata di 1,28 x 105 ± 0,05 (media ± errore standard della media (SEM), N = 100). Non c'è differenza nella resa della microglia tra topi femmine (1,25 x 105 ± 0,09 [media ± SEM, N=46]) e maschi (1,32 x 105 ± 0,07 [media ± SEM, N=54]) (t(98)=0,6365, p=0,526). Quando si isola da specifiche regioni cerebrali, la resa media della microglia dalla corteccia di topo è di 8,3 x 104 ± 0,08 (media ± SEM, N = 15) e dall'ippocampo del topo è 4,1 x 104 ± 0,02 (media ± SEM, N = 16). Come previsto, c'è una differenza significativa nella resa della microglia da ciascuna regione del cervello (F(2, 128)=25,25, P<0,0001). Dopo l'isolamento della microglia, l'RNA è stato estratto dalle cellule isolate utilizzando un kit di isolamento dell'RNA a basso input. Coerentemente il punteggio di integrità dell'RNA (RIN) è stato superiore a 9,0 (9,62 ± 0,05) e la resa media di RNA per cellula è stata di 0,25 ± 0,01 pg (media ± SEM, N=32; Fascicolo supplementare S2).
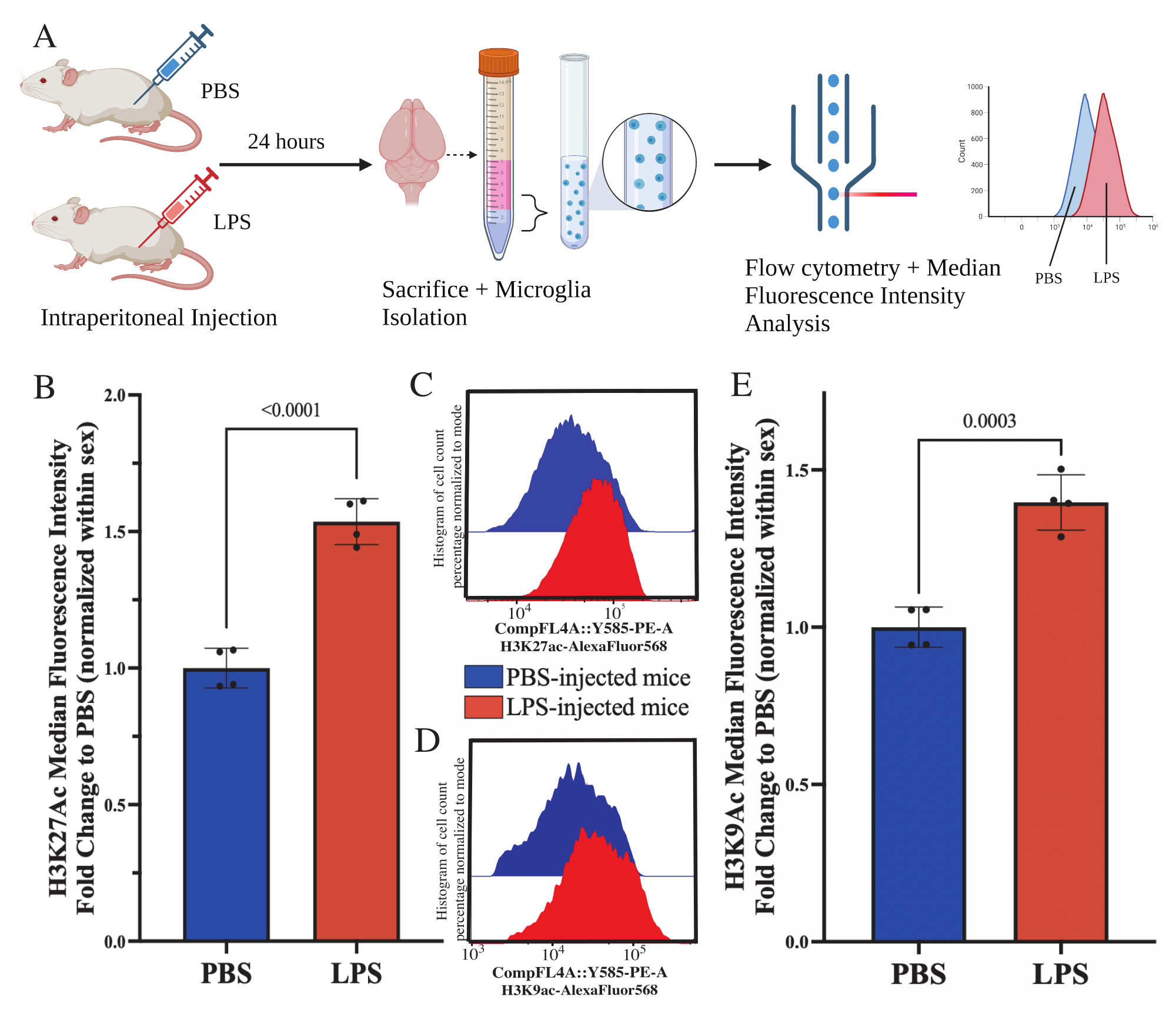
I topi adulti sono stati iniettati per via intraperitoneale con 1 mg/kg di lipopolisaccaride (LPS) 24 ore prima del sacrificio. I topi sono stati perfusi per via transcardiaca con HBSS e la microglia è stata isolata dall'intero cervello secondo il protocollo descritto (Figura 5A). Per ogni colorazione, 20.000-30.000 cellule sono state assegnate a ciascun pannello di anticorpi. I livelli globali di acetilazione dell'istone 3 lisina 27 (H3K27Ac) sono stati valutati in microglia isolate tramite citometria a flusso. Per topi maschi e femmine, il trattamento con LPS ha indotto un aumento di H3K27Ac quando l'MFI è normalizzato all'interno del sesso (t(6)=9.676, p<0.0001; Figura 5B). Quando si esaminano gli istogrammi per le cellule colorate, le popolazioni rimangono normalmente distribuite con variazioni simili; tuttavia, le cellule sono passate a una maggiore fluorescenza con conseguente aumento dell'MFI (Figura 5C). Quando si esamina H3K9Ac nello stesso trattamento, si osserva un aumento simile di H3K9Ac (t(6)=7,299, p=0,0003; Figura 5D,E) tuttavia la variazione di piega dell'LPS rispetto al PBS del segnale H3K9Ac è inferiore al segnale H3K27Ac.

Figura 5: Cambiamenti globali nell'acetilazione degli istoni nella microglia isolata. (A) I topi vengono iniettati per via intraperitoneale con soluzione salina tamponata con fosfato (PBS) o 1 mg/kg di lipopolisaccaride (LPS) 24 ore prima del sacrificio. Le microglia vengono raccolte dalla frazione arricchita di immunità e fissate per la citometria a flusso e la valutazione della modificazione post-traduzionale degli istoni globali. L'intensità fluorescente mediana è valutata come proxy per l'espressione proteica. Creato con BioRender.com. (B) I livelli globali di H3K27Ac sono aumentati in risposta al trattamento con LPS. Cambio di piega in PBS normalizzato all'interno dell'esperimento e del sesso. T-test spaiato a due code, t(6)=9.676, p<0.0001. Il grafico a barre rappresenta la media ± SEM. N=8 animali; 2 per condizione in 2 esperimenti indipendenti. (C) Istogrammi di esempio che raffigurano lo spostamento dell'intensità della fluorescenza H3K27Ac. Modal raffigura gli istogrammi dei topi iniettati con PBS rispetto a quelli iniettati con LPS. (D) Istogrammi di esempio che raffigurano lo spostamento dell'intensità della fluorescenza H3K9Ac. Modal raffigura gli istogrammi dei topi iniettati con PBS rispetto a quelli iniettati con LPS. (E) I livelli globali di H3K9Ac sono aumentati in risposta al trattamento con LPS. Cambio di piega in PBS normalizzato all'interno dell'esperimento e del sesso. T-test a due code spaiato, t(6)=7.299, p=0.0003. Il grafico a barre rappresenta la media ± SEM. N=8 animali; 2 per condizione in 2 esperimenti indipendenti. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Per confermare che il metodo descritto era paragonabile ad altri metodi precedentemente utilizzati per la quantificazione della modificazione istonica globale, abbiamo mirato a utilizzare l'immunoblot come strumento comparativo. Tuttavia, la resa della microglia isolata è semplicemente troppo bassa per consentire una valutazione ragionevole. Pertanto, abbiamo utilizzato cellule BV2 in coltura per confrontare il metodo di citometria a flusso intracellulare con un Western blot (WB). Le cellule BV2 sono state coltivate in terreni completi (DMEMF12, 10% FBS, 1x penicillina/streptamicina e 1x L-glutammina) a 37 °C, 5% CO2. Le cellule sono state passate con tripsina-EDTA allo 0,25% e piastrate a una densità di 250.000 cellule/pozzetto e trattate in terreni sierici ridotti (DMEM F12, 2% FBS, 1x penicillina/streptamicina e 1x L-glutammina) e lasciate recuperare per 12 ore a 37 °C, 5% CO2. Le cellule sono state trattate con 25 ng/mL di LPS per 24 ore prima della fissazione come descritto sopra o della lisi con un tampone di lisi WB. Il segnale di H3K27Ac è stato eseguito con entrambi i metodi con GAPDH utilizzato come controllo di carico per WB. L'analisi dell'intensità fluorescente normalizzata rispetto al controllo PBS è stata determinata per ciascun gruppo (Figura 6A). Quando si esamina la variazione del segnale H3K27Ac normalizzato da parte di WB, c'è stato un aumento di 1,527 volte nella condizione trattata con LPS rispetto al controllo H2O che è stato determinato come significativo dal test t spaiato (t=3,024, df=5; p=0,0293). Quando si esamina il cambiamento utilizzando la citometria a flusso, c'è stato un aumento di 1,482 volte nella condizione trattata con LPS che è stata determinata come significativa (t = 7,843, df = 10; p<0,0001). Utilizzando un'ANOVA a 2 vie per confrontare i metodi, è stato determinato che c'è un effetto significativo del trattamento (F(1,15)=45,21,p<0,0001), ma non il metodo (F(1,15)=0,05545, p=0,8697) o l'interazione (F(1,15)=0,02785, p=0,8697). Inoltre, verifichiamo qui che non vi è alcun cambiamento nei livelli di istone H3 sia mediante Western blot che con citometria a flusso, poiché l'ANOVA a 2 vie non ha rivelato alcun effetto significativo del trattamento LPS (F(1,7)=0,02170,p=0,8870), del metodo (F(1,7)=0,01191, p=0,9162) o dell'interazione (F(1,7=0,01191, p=0,9162; Figura 6B). Vengono mostrati anche esempi di blot e spostamenti dell'istogramma per questi dati (Figura 6C,D).

Figura 6: Confronto dei metodi per la quantificazione della variazione globale della modificazione istonica tra citometria a flusso e western blot. (A) Le cellule BV2 vengono trattate con lipopolisaccaride (LPS) o H2O da 25 ng/mL per 24 ore prima dell'analisi. L'intensità fluorescente di H3K27Ac è rappresentata come variazione di piega al controllo del veicolo, soluzione salina tamponata con fosfato (PBS), sia per la citometria a flusso che per il western blot. L'ANOVA a 2 vie ha rivelato un effetto significativo del trattamento LPS (F(1,15)=45,21, p<0,0001), ma non del metodo (F(1,15)=0,05545, p=0,8697) o dell'interazione (F(1,15)=0,02785, p=0,8697). La correzione di Tukey per la verifica di ipotesi multiple è stata applicata per i residui. * presenta 0,0332, ** presenta 0,0021. (B) L'intensità fluorescente per l'istone H3 è rappresentata come variazione di piega in PBS sia per la citometria a flusso che per il western blot. L'ANOVA a 2 vie non ha rivelato alcun effetto significativo del trattamento LPS (F(1,7)=0,02170, p=0,8870) o del metodo (F(1,7)=0,01191, p=0,9162) o dell'interazione (F(1,7=0,01191, p=0,9162). (C) Vengono rappresentati blot esemplificativi (D) spostamenti della citometria a flusso. La dimensione dell'istogramma viene normalizzata in percentuale in base al numero di cellule presenti all'intensità fluorescente modale. Il grafico a barre mostra il SEM medio. n=2 esperimenti indipendenti, 2 per condizione per esperimento. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Tutti insieme, questi risultati mostrano che questa tecnica può essere utilizzata per valutare quantitativamente i livelli globali di HPTM nella microglia isolata. Inoltre, il metodo ha dimostrato di essere paragonabile alle tecniche precedenti, ma che richiede input cellulari molto più bassi. Inoltre, anche se non mostrata, con un'adeguata compensazione, la presente tecnica può essere utilizzata con più anticorpi sullo stesso pannello che valutano diversi HPTM.
File supplementare S1: File di analisi di esempio. Questo file contiene un file di analisi wsp e 7 file fcs tra cui nessun colorante, P2RY12FMO, 568FMO, due animali trattati con PBS e due animali trattati con LPS colorati con H3K27Ac. Lo scopo di questo file è quello di dimostrare l'analisi e il gating di un esperimento in grado di rappresentare l'aspetto di un esperimento riuscito. Fare clic qui per scaricare il file.
File supplementare S2: Dati di isolamento. Il file incluso contiene i dati rilevanti dopo l'ordinamento della microglia che contiene la microglia e la resa dell'RNA dal protocollo descritto. Fare clic qui per scaricare il file.
| POPOLAZIONE RECINTATA | Frequenza del genitore | Frequenza del totale | Contare |
| Visualizzazione del materiale S1 | 89.00% | 89.00% | 25672 |
| S2>S1 | 88.73% | 78.97% | 22779 |
| APC+ > S2 > S1 | 76.61% | 60.50% | 17452 |
| 610+ > APC+ > S2 > S1 | 99.56% | 60.24% | 17376 |
Tabella 2: Il grafico di derivazione del campione di esempio illustra la percentuale e i numeri di eventi necessari per un rilevamento accurato delle proteine.
Discussione
Il protocollo presentato consente la valutazione quantitativa dei livelli globali di HPTM attraverso la citometria a flusso. Sebbene questo protocollo presenti un metodo innovativo, studi precedenti hanno effettuato una valutazione quantitativa delle proteine utilizzando un approccio simile26. I metodi precedentemente utilizzati per valutare i livelli globali di HPTM includono l'immunoistochimica e il western blot 16,17,19,20. Il metodo basato sulla citometria a flusso presentato è un metodo facilmente quantificabile, mentre il western blot e l'immunoistochimica sono semiquantitativi e hanno una produttività inferiore. Il Western blot si basa sulla lisi cellulare e quindi richiede sia la normalizzazione della proteina che una proteina di controllo del carico che si presume sia invariata dalla condizione sperimentale27. L'immunoistochimica è semiquantitativa e a bassissimo rendimento, in quanto è difficile valutare quantitativamente la quantità di proteine senza esaminarla a livello di singola cellula16. Allo stesso modo, per la microglia isolata, c'è un vantaggio nell'utilizzare il metodo della citometria a flusso a causa della resa limitata, poiché il western blot richiede un apporto proteico molto più grande19. I bassi requisiti di numero di celle consentono di eseguire più pannelli di colorazione dallo stesso animale.
Tuttavia, come con qualsiasi altro metodo, ci sono limitazioni a questa tecnica, tra cui il costo e la disponibilità degli anticorpi, poiché non tutti gli anticorpi funzionano bene in un contesto di citometria a flusso. Inoltre, rispetto all'immunoblot, la concentrazione di anticorpi richiesta è molto più alta. Mentre il multiplexing consente di utilizzare più anticorpi sullo stesso pannello di cellule, le cellule non possono essere private dell'anticorpo dopo l'analisi, limitando così l'uso delle cellule a uno per specie di anticorpi. Questo è diverso dall'immunoblot in cui lo stesso blot può essere utilizzato ripetutamente. Tuttavia, a seconda della disponibilità di anticorpi e del numero di canali di rilevamento su un citometro, sarebbe possibile esaminare fino a una dozzina di segni contemporaneamente.
Il metodo attuale cattura solo i livelli globali di espressione proteica e non la posizione genomica specifica, e i cambiamenti nei livelli globali potrebbero non riflettere i cambiamenti nei singoli loci genomici. Allo stesso modo, la mancanza di cambiamenti nei livelli globali potrebbe non significare che nessun loci genomico stia subendo cambiamenti, ma semplicemente che i cambiamenti globali non comportano differenze tra i trattamenti. In quanto tale, questa tecnica è pensata per essere utilizzata come schermo per identificare HPTM di interesse per l'analisi genomica. Inoltre, questo metodo non consente il confronto tra diversi marcatori proteici, tranne quando viene valutato come un cambiamento di piega per il controllo. Pertanto, questo è limitato rispetto a un metodo standard basato su curve come l'ELISA per la determinazione delle proteine.
Il protocollo presentato offre una strategia per isolare la microglia cerebrale viva. Questo protocollo si basa sull'espressione della proteina P2RY12 per l'isolamento della microglia. Tuttavia, P2RY12 è un marcatore omeostatico nella microglia e può essere sottoregolato in modelli di malattia, come 5XFAD22. Pertanto, quando si utilizza un animale modello di malattia, assicurarsi di scegliere altre proteine marcatrici come TMEM119, CD11b o CD45 per aiutare nell'isolamento della microglia23. Allo stesso modo, presentiamo questo protocollo come isolamento dall'ippocampo e/o dalla corteccia. Questo protocollo funzionerebbe per isolare la microglia da altre regioni del cervello, comprese le regioni della sostanza bianca, tuttavia, potrebbero essere necessari più animali per ottenere una quantità sufficiente di microglia a seconda delle dimensioni delle regioni di interesse.
Il protocollo presentato è in grado di isolare in modo robusto la microglia cerebrale viva, ma ci sono diversi passaggi, descritti di seguito, nella fase di isolamento che possono ridurre la resa cellulare se eseguiti in modo errato.
Le perfusioni per questo protocollo si traducono in una percentuale maggiore di microglia nel frammento arricchito di immuni, che ridurrà la quantità di tempo al selezionatore. Tuttavia, la perfusione non è richiesta e, se necessario, possono essere utilizzati altri metodi di eutanasia.
Durante l'isolamento della microglia, la mielina deve essere completamente rimossa. I citometri a flusso si basano sul fatto che le cellule siano in grado di viaggiare attraverso tubi stretti a un ritmo rapido. A causa della sua viscosità e della tendenza ad aggregarsi, la mielina causa problemi con i citometri, spesso causando intasamenti che possono danneggiare l'apparecchiatura e distruggere il campione, riducendo drasticamente la resa. Prestare attenzione a rimuovere tutta la mielina durante la raccolta di frammenti immuno-arricchiti per evitare di avere problemi a valle.
Colorazione su piastra e colorazione in provetta: in questo protocollo, abbiamo descritto due opzioni per la colorazione delle cellule in provette da 1,5 mL o in una piastra a 96 pozzetti. Il caso d'uso per ciascuno dipende dall'esperimento; Tuttavia, in generale, la colorazione delle provette comporta un rischio inferiore di influire sulla resa rispetto alla colorazione delle piastre, poiché il flick rischia la perdita di cellule se eseguita in modo errato. La colorazione delle piastre è molto più veloce poiché l'aspirazione del surnatante per ogni provetta richiede molto tempo. Prima della fissazione (per lo smistamento, ecc.), utilizzare la colorazione delle provette per massimizzare la resa e ridurre il rischio di perdita. Tuttavia, per l'analisi HPTM, una volta che le cellule sono fissate per la colorazione intranucleare, il pellet è più stabile e c'è un rischio ridotto di perdita con il flicking.
Stabilire il gradiente di densità discontinuo: Quando si stabilisce la stratificazione, è essenziale impostare correttamente gli strati per ottenere la frazione arricchita immunitaria. Se gli strati sono disturbati o mescolati e appaiono torbidi, le cellule non si ordineranno nella posizione desiderata e ci sarà difficoltà nell'ottenere la frazione cellulare arricchita con immunità. In tal caso, centrifugare con il mezzo di densità per rimuovere la mielina e quindi raccogliere tutte le frazioni rimanenti, diluire con 3 mL di tampone FACS in 1 mL di mezzo di densità e mescolare bene (ciò richiederà più provette). Girare a 500 x g per 10 min con il freno su 0. Scartare il surnatante, lasciando solo ~300 μL di soluzione. Raccogliere l'intero campione e la colorazione. Ciò produrrà percentuali di ordinamento ridotte e una maggiore quantità di tempo trascorso al citometro, ma la resa può comunque essere comparabile.
Quando si utilizza il metodo di isolamento, è utile essere in grado di raccogliere cellule per l'RNA e per la valutazione HPTM dallo stesso cervello di topo. In questa situazione, dopo aver selezionato la microglia viva, le cellule possono essere divise per allocare una parte alla valutazione dell'RNA (il numero minimo di cellule in ingresso per ottenere una resa decente di RNA è di 75.000 cellule) e una parte per un'ulteriore analisi citometrica a flusso (minimo 10.000 cellule per pozzetto per una buona determinazione dell'IFM). In questo caso, è necessario lo smistamento con citometro a flusso. Tuttavia, quando si prevede di utilizzare le cellule solo per l'analisi HPTM, non è necessario lo smistamento e la frazione immunitaria può essere colorata con l'anticorpo P2RY12 e l'anticorpo HPTM. Il gating sul citometro può quindi essere impostato per la microglia P2RY12+, come si farebbe per lo smistamento del flusso, per analizzare solo il segnale HPTM all'interno della microglia. L'eliminazione dello smistamento consente al protocollo di essere più veloce ed economico. Inoltre, se si valutano HPTM da cellule in coltura, è sufficiente iniziare con il protocollo di colorazione e non sono necessari anticorpi marcatori cellulari, come dimostrato nella Figura 6. Il protocollo di valutazione HPTM può essere utilizzato per molti tipi di cellule, tra cui cellule in coltura, primarie e derivate da IPSC.
Infine, mentre abbiamo presentato solo due potenziali usi della microglia a valle dell'isolamento, ce ne sono molti altri che includono tecniche epigenetiche come ChIP, CUT&Tag e CUT&RUN. Nel caso di tecniche epigenetiche genomiche, in cui è interessante caratterizzare i cambiamenti in loci specifici, scegliere inibitori specifici per scrittori e cancellatori di segni di cromatina11 su misura per gli esperimenti per garantire che eventuali modificazioni epigenetiche microgliali profilate non siano artefatti tecnici derivanti da qualsiasi fase della procedura di isolamento come la digestione enzimatica. Quando si valutano i cambiamenti nei livelli globali di marcatori epigenetici, ad esempio utilizzando la citometria a flusso quantitativa, non ci si aspetta che i cambiamenti indotti dalla procedura siano così grandi da essere rilevati a livello globale.
Nel complesso, i metodi discussi forniscono un nuovo metodo a singola cellula per quantificare i livelli globali di modificazioni istoniche e altri cambiamenti epigenetici mediante citometria a flusso. Abbiamo dimostrato che questo metodo è sufficientemente sensibile per rilevare cambiamenti globali nel marcatore enhancer H3K27ac nella microglia in risposta a LPS in vivo. Ciò è coerente con il precedente sequenziamento ChIP di H3K27ac a seguito di stimolazione LPS che mostrava un drammatico rimodellamento degli enhancer responsivi a LPS28. Le applicazioni di questo metodo consentiranno l'esame dei cambiamenti epigenetici globali in diversi tipi di cellule cerebrali nello sviluppo e nella malattia.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Grazie a Yanyang Bai per l'aiuto con l'immunoblot nella Figura 5. Questo lavoro è stato sostenuto dagli Istituti canadesi per la ricerca sulla salute [CRC-RS 950-232402 to AC]; Consiglio canadese per la ricerca in scienze naturali e ingegneria [RGPIN-2019-04450, DGECR-2019-00069 a AC]; Fondazione di beneficenza del rito scozzese [da 21103 ad AC] e la Fondazione Brain Canada [AWD-023132 ad AC]; Borsa di studio per laureati aborigeni dell'Università della British Columbia (da 6481 a MT); Borsa di studio per laureati della Columbia Britannica (da 6768 a MT); Premio canadese per studenti della piattaforma Open Neuroscience (da 10901 a JK); Borsa di dottorato quadriennale dell'Università della British Columbia (da 6569 a JK). I finanziatori non hanno avuto alcun ruolo nella progettazione dello studio, nella raccolta e nell'analisi dei dati, nella decisione di pubblicare o nella preparazione del manoscritto.
Materiali
| Name | Company | Catalog Number | Comments |
| 0.5M EDTA | Invitrogen | AM9260G | |
| 15 mL Falcon Centrifuge Tubes, Polypropylene, Sterile | Falcon | 352196 | |
| 24-well Clear Not Treated Plates | Costar | 3738 | |
| 2-Mercaptoethanol | Gibco | 21985023 | |
| 96 Well Clear Polystyrene Microplate, clear round bottom, non treated surface | Corning | 3788 | |
| Acetyl Histone 3 K9 (C5B11) | Cell Signalling Technology | 9649S | Dilution: 1:100 |
| Acetyl Histone H4 K8 (2594) | Cell Signalling Technology | 2594S | Dilution: 1:100 |
| Acetyl-Histone H3 K27 (D5E4) | Cell Signalling Technology | 8173S | Dilution: 1:100 |
| Acetyl-Histone H3 Lys27 (MA523516) | Invitrogen | MA523516 | Dilution: 1:100 |
| Actinomycin D | New England Biolabs | 15021S | |
| Anisomycin | New England Biolabs | 2222S | |
| Anti-Histone H3 (tri methyl K4) | Abcam | ab213224 | Dilution: 1:100 |
| Anti-Lactyl-Histone H4 (Lys 12) Rabbit mAb | PTM Biolabs | PTM-1411RM | Dilution: 1:250 |
| Anti-L-Lactyllysine Rabbit pAb | PRM Biolabs | PTM-1401RM | Dilution: 1:250 |
| Apc anti-P2RY12 Antibody, Clone: S16007D | BioLegend | 848006 | |
| BSA | Tocris | 5217 | |
| Cyto-Last Buffer | BioLegend | 422501 | |
| dimethylsulfoxide, sterile | Cell Signalling Technology | 12611S | |
| DNAse I | STEMCELL Technologies | 07900 | |
| Donkey Anti Mouse AlexaFluor488 | Jackson ImmunoResearch | 715-546-150 | Dilution: 1:500 |
| Donkey Anti Rabbit AlexaFluor488 | ABclonal | AS035 | Dilution: 1:500 |
| Donkey Anti Rabbit AlexaFluor568 | Invitrogen | A10042 | Dilution: 1:500 |
| Donkey Anti Rabbit Brilliant Violet 421 | BioLegend | 406410 | Dilution: 1:500 |
| Fisherbrand Disposable Graduated Transfer Pipettes | Fisherbrand | 13-711-9AM | |
| Fisherbrand Disposable PES Filter Unit, 250mL | Fisherbrand | FB12566502 | |
| H3K18ac Polyclonal Antibody | Invitrogen | 720095 | Dilution: 1:100 |
| HBSS (10X), no calcium, no magnesium, no phenol red | Gibco | 14185052 | |
| HBSS, no calcium, no magnesium, no phenol red | Gibco | 14175103 | |
| Histone 3 Trimethyl K27 (ab6002) | Abcam | ab6002 | Dilution: 1:100 |
| KONTES Dounce Tissue Grinders 125mm 7mL | VWR | 885300-0007 | |
| Lactyl-Histone H3 (Lys 18) Rabbit mAb | PTM BIolabs | PTM-1406RM | Dilution: 1:250 |
| Lipopolysacharide | Sigma-Aldrich | L5418 | |
| Normal Donkey Serum | Jackson ImmunoResearch | 017-000-121 | |
| OneComp eBeads Compensation Beads | Invitrogen | 01-1111-41 | |
| PDS Kit, Papain Vial - Worthington Biochemical | Cedarlane | LK003178 | |
| Percoll | Sigma-Aldrich | GE17-0891-02 | |
| Phenol Red | VWR | RC57004 | |
| QIAshredder | Qiagen | 79656 | |
| Rainbow Fluorescent Particles, 1 peak (3.0-3.4 uM - Mid Range Intensity | BioLegend | 422905 | |
| RNase-free Microfuge Tubes, 1.5 mL | Invitrogen | AM12400 | |
| Rneasy Plus Micro Kit | Qiagen | 74034 | |
| Round Bottom Polypropylene Tubes with Caps, 5 mL | Corning | 352063 | |
| Triptolide | New England Biolabs | 97539 | |
| True Nuclear Transcription Factor Buffer Set | BioLegend | 424401 | |
| TruStain FcX PLUS (anti-mouse CD16/32) Antibody | BioLegend | 156604 | |
| Trypan Blue | VWR | 97063-702 | |
| Zombie Aqua Fixable Viability Kit | BioLegend | 423102 |
Riferimenti
- Miller, J. L., Grant, P. A. The Role of DNA Methylation and Histone Modifications in Transcriptional Regulation in Humans. Epigenetics: Development and Disease. 61, 289-317 (2013).
- Kouzarides, T. Chromatin Modifications and Their Function. Cell. 128 (4), 693-705 (2007).
- Bannister, A. J., Kouzarides, T. Regulation of chromatin by histone modifications. Cell Research. 21 (3), 381-395 (2011).
- Barski, A., et al. High-resolution profiling of histone methylations in the human genome. Cell. 129 (4), 823-837 (2007).
- Vogel Ciernia, A., LaSalle, J. The landscape of DNA methylation amid a perfect storm of autism aetiologies. Nature Reviews. Neuroscience. 17 (7), 411-423 (2016).
- Keiser, A. A., et al. Systemic HDAC3 inhibition ameliorates impairments in synaptic plasticity caused by simulated galactic cosmic radiation exposure in male mice. Neurobiology of Learning and Memory. 178, 107367 (2021).
- McQuown, S. C., et al. HDAC3 is a critical negative regulator of long-term memory formation. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 31 (2), 764-774 (2011).
- Barrett, R. M., et al. Hippocampal Focal Knockout of CBP Affects Specific Histone Modifications, Long-Term Potentiation, and Long-Term Memory. Neuropsychopharmacology. 36 (8), 1545-1556 (2011).
- Datta, M., et al. Histone Deacetylases 1 and 2 Regulate Microglia Function during Development, Homeostasis, and Neurodegeneration in a Context-Dependent Manner. Immunity. 48 (3), 514.e6-529.e6 (2018).
- Belhocine, S., et al. Context-dependent transcriptional regulation of microglial proliferation. Glia. 70 (3), 572-589 (2022).
- Gosselin, D., et al. An environment-dependent transcriptional network specifies human microglia identity. Science (New York, N.Y.). 356 (6344), eaal3222 (2017).
- Kettenmann, H., Hanisch, U. -. K., Noda, M., Verkhratsky, A. Physiology of Microglia. Physiological Reviews. 91 (2), 461-553 (2011).
- Sullivan, O., Ciernia, A. V. Work hard, play hard: how sexually differentiated microglia work to shape social play and reproductive behavior. Frontiers in Behavioral Neuroscience. 16, 989011 (2022).
- Das, P. M., Ramachandran, K., vanWert, J., Singal, R. Chromatin immunoprecipitation assay. BioTechniques. 37 (6), 961-969 (2004).
- Mahmood, T., Yang, P. C. Western blot: technique, theory, and trouble shooting. North American Journal of Medical Sciences. 4 (9), 429-434 (2012).
- Crowe, A., Yue, W. Semi-quantitative Determination of Protein Expression Using Immunohistochemistry Staining and Analysis: An Integrated Protocol. BIO-PROTOCOL. 9 (24), (2019).
- Seligson, D. B., et al. Global histone modification patterns predict risk of prostate cancer recurrence. Nature. 435 (7046), 1262-1266 (2005).
- Liu, B., et al. Global Histone Modification Patterns as Prognostic Markers to Classify Glioma Patients. Cancer Epidemiology, Biomarkers & Prevention. 19 (11), 2888-2896 (2010).
- Pan, R. Y., et al. Positive feedback regulation of microglial glucose metabolism by histone H4 lysine 12 lactylation in Alzheimer's disease. Cell Metabolism. 34 (4), 634.e6-648.e6 (2022).
- Zhang, D., et al. Metabolic regulation of gene expression by histone lactylation. Nature. 574 (7779), 575-580 (2019).
- Pösel, C., Möller, K., Boltze, J., Wagner, D. C., Weise, G. Isolation and Flow Cytometric Analysis of Immune Cells from the Ischemic Mouse Brain. Journal of Visualized Experiments. (108), 53658 (2016).
- Oblak, A. L., et al. Comprehensive Evaluation of the 5XFAD Mouse Model for Preclinical Testing Applications: A MODEL-AD Study. Frontiers in Aging Neuroscience. 13, 713726 (2021).
- Bohlen, C. J., Bennett, F. C., Bennett, M. L. Isolation and Culture of Microglia. Current Protocols in Immunology. 125 (1), e70 (2019).
- McKinnon, K. M. Multiparameter Conventional Flow Cytometry. Flow Cytometry Protocols. 1678, 139-150 (2018).
- Marsh, S. E., et al. Dissection of artifactual and confounding glial signatures by single-cell sequencing of mouse and human brain. Nature Neuroscience. 25 (3), 306-316 (2022).
- Wang, L., Gaigalas, A. K., Marti, G., Abbasi, F., Hoffman, R. A. Toward quantitative fluorescence measurements with multicolor flow cytometry. Cytometry Part A. 73A (4), 279-288 (2008).
- Rumbaugh, G., Miller, C. A. Epigenetic changes in the brain: measuring global histone modifications. Methods in Molecular Biology (Clifton, N.J). 670, 263-274 (2011).
- Xavier, A. M., et al. Systematic delineation of signaling and epigenomic mechanisms underlying microglia inflammatory activity in acute and chronic brain pathologies. BioRvix. , (2022).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati