
JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
심장 수술 후 혈청 샘플의 영향을 건강한 자원 봉사자와 그들의 철새 잠재력에서 내피 전구 세포의 분리
요약
내피 전구 세포 (내피 전구 세포는) 결정적으로 허혈성 조직의 혈관 신생에 관여한다. 이 방법은 말초 혈액에서 인간 내피 전구 세포의 분리뿐만 아니라, 심장 수술 환자의 혈청 샘플에 대한 그들의 이동성 전위의 식별을 설명한다.
초록
내피 전구 세포 (내피 전구 세포)은 저산소증과 같은 병리 적 조건에서 골수로부터 모집 결정적 허혈성 조직의 혈관 신생에 관여한다. 내피 전구 세포의 기원, 분류 및 특성은 복잡; 에도 불구하고, 내피 전구 세포의 두 눈에 띄는 하위 유형이 설립되었습니다 소위 "초"내피 전구 세포는 늦게-가지 내피 전구 세포 (후기 내피 전구 세포) (이후로-초기 내피 전구 세포 함). 이들은 생물학적 특성에 의해뿐만 아니라, 체외 배양시 그 모양에 따라 분류 할 수있다. "초기"내피 전구 세포는 EC-특정 미디어의 말초 혈액 유래 단핵구 세포의 배양 후보다 일주일에 표시되지만, 늦은 가지 내피 전구 세포 2 ~ 3 주 후에 찾을 수 있습니다. 늦게 가지 내피 전구 세포는 "초기"내피 전구 세포가 Endogen 사 등 다양한 혈관 신생 인자를 표현하는 반면, 직접 주로 성숙 내피 세포로 분화 할 수있는 능력을 통해, 혈관 신생에 관여하는 것으로 인정 받고 있습니다OU에화물이 분비 방식으로 혈관 신생을 촉진합니다. 심근 허혈 / 재관류 (I / R) 동안, 다양한 요소가 혈관 형성의 영역에 내피 전구 세포의 원점 복귀를 제어 할 수 있습니다.
대 식세포 이동 억제 인자 (MIF)은 케 모킨과 같은 염증성 사이토킨 및 보편적으로 발현하고 최근 생리적 농도 1 내피 전구 세포 이주 같은 주요 조절 기능하도록 설명 하였다. 흥미롭게 MIF는 세포 풀에 저장되고 신속하게 여러 자극 후의 혈류 내로 (예를 들면, 심근 경색)을 해제 할 수있다.
이 프로토콜은 혈청 샘플에 대한 시험 관내 마이그레이션 분석에 사용 피브로넥틴 - 코팅 된 플레이트에 내피 성장 인자가 포함 된 배지에서 이후의 문화와 CD34 양성 선택에 따라 성인의 말초 혈액에서 초기 내피 전구 세포의 신뢰성 분리 및 배양하는 방법을 설명합니다 심장 수술 환자. 또한,다른 잘 알려진 혈관 신생 촉진 사이토 카인에 비해 내피 전구 세포의 화학 주성에 MIF의 철새 영향이 설명된다.
서문
내피 전구 세포 (내피 전구 세포)는 인간의 혈액 순환 및이 내피 세포로 분화 할 수있는 능력을 가지고있다. 이들은 혈관 형성에 참여하고 다양한 방법으로 3,4- 염증 및 허혈 / 재관류 (I / R) 손상에 의한 피해를 최소화 할 수있다. 예를 들어, 내피 전구 세포는 카탈라아제, 글루타티온 과산화 망간 슈퍼 옥사이드 dismutases (MnSOD) 5와 같은 세포 내 항산화 효소의 높은 수준을 보여줍니다. 산화 적 스트레스에 대한 높은 저항은 내피 전구 세포가 허혈 손상 6 후 상승 된 활성 산소 종 (ROS)와 미세 환경에서 작동 할 수 있습니다. 이전의 연구는 또한 내피 전구 세포의 개수는 혈관 내피 전구 세포 수리하고 순환 수가 감소는 심혈 관계 사건 (7)의 발생을 예측하는 것이 상관 될 수 있음을 지적클래스 = "외부 참조"> 8. 그러나, EPC 명확한 정의은 아직 발견하지 못했다. 지금까지가 내피 전구 세포에 대한 특정한 세포 표면 마커 또는 일관성 표현형이 없으며 이들 세포는 말초 혈액 (9)은 매우 드물다. 인간 EPC는 손상된 내피 새로운 혈관 구조의 재구성에 기여하는 기능과 순환 셀로서 고려되어야한다.
단리 및 내피 전구 세포를 특징 짓는 한 가지 방법은, 피브로넥틴의 밀착성을 통해서이다. 따라서, 이들 전지의 용량은 실시 예 3, 10, 11, 1, 콜라겐을 입력 비해 코팅 요리 피브로넥틴 우수한 밀착성을 표시하는 데 사용된다. 그러나, 다른 임의의 이전 또는 추가의 정제 단계없이 피브로넥틴 코팅 접시에 단핵구 도금하는 골수 전구 세포, 단핵구 및 T 림프구를 포함 하나 콜로니 리드 것으로2, 13, 14. 또한,이 경우, 혈소판 단핵 세포 (MNC) 부분을 오염시킬 수 있으며 따라서 임의의 부착 세포 (15) 세포막 단백질을 옮긴다.
시험 관내 접착 분석을 통해 특성화 이외에 다른 세포 표면 마커의 조합은 EPC 간주 세포 유형을 서술하기 위해 사용된다. 이 때, 피브로넥틴 매개을 접착 한 후, 세포는 내피 세포와 같은 특성에 관하여 분석된다. 이 과정에서, 두 개의 내피 세포 - 관련 마커 아세틸 - 저밀도 지단백질 (acLDL) 및 혈관 내피 성장 인자 수용체 2 (VEGFR-2, KDR)에 역할을한다. 내피 세포와 대 식세포는 특히 "폐품 세포 경로"(16)라고하는 과정에서 acLDL을 차지하는 것으로 나타났다. 또 다른 마커 단백질은 내피 세포의 주 VEGF 수용체로 KDR입니다세포 17. 일반 내피 전구 세포는 혈관 내피 성장 인자 및 소 태아 혈청이 보충 된 배지에서 배양 그러나, 마크로파지 것은 또한 실수, 전시 내피 형 마커 프로필 격리되었을 수있는 것이 가능하다. 이전에 도시 된 바와 같이, 내피 에어컨 배지에서 배양하면, 대 식세포는 "내피 별"단백질 18 표현한다.
일반적으로, 혈액에서 발견 될 수 있거나 시험 관내에서 배양 이상의 아형 내의 내피 전구 세포의 두 가지 종류가있다. 늦게 가지 내피 전구 세포 (후기 내피 전구 세포)를 배양 2-3 주 후에 나타납니다. 이러한 세포는 인간 제대 정맥 내피 세포의 단층으로 빠른 통합되어 모세관 튜브 (19)를 형성 할 수있다. 또한, 혈관 내피 성장으로 제공하는 혈관 신생 분자를 통해 더 수동적 인 방법으로 일주일 정도 및 행위의 혈액 순환 "초기 내피 전구 세포 소위"인자 (VEGF), 또는 CXCL8 19. 관상 동맥 질환 (CAD) 환자들은 CAD (20)가없는 대조군에 비해 초기 내피 전구 세포의 매우 낮은 양을 보여 주었다. 흥미롭게도, 같은 그룹은 대조군에 비해 늦은 내피 전구 세포의보다 많은 양을 보였다. 또 다른 연구는 초기 내피 전구 세포가 분비 방식으로 6 산화 조건에서 세포 사멸에서 차별화 된 내피 전구 세포를 보호하는 것으로 나타났다. 따라서, 초기 내피 전구 세포는 말초 혈액 내에서 자동 또는 분비 방식으로 다른 세포의 이동을 통해 관련 보호 효과를 제공 할 수 있습니다.
이 프로토콜은 먼저 인간의 말초 혈액에서 PBMC-부분을 분리하고 이후에 원치 않는 세포에서이 세포 현탁액을 취소 할 PBMC-부분에서 CD34 + 세포를 분리하여 초기 내피 전구 세포를 정화하는 방법을 설명합니다. CD34 인간 조혈 줄기 세포 (9)의 분리에 사용되는 마커 인 . 이후, CD34 + 세포가 피브로넥틴 코팅 조직 배양 표면에 배양한다. 사흘, 배지함으로써 모든 비 부착 세포를 잃고, 변경된다. 마지막으로, 고립 된 내피 전구 세포는 acLDL의 흡수 및 형광 활성화 된 셀 정렬 (FACS)를 사용하여 내피 세포 마커로 KDR의 존재를 확인하기 위해 염색된다. 추가적인 마커로서, 우리는 또한 내피 세포에서 발생 혈소판 내피 세포 부착 분자 (PECAM-1, CD31)을 분석 하였다.
내피 전구 세포의 강화 된 채용에 의해 손상되거나으로 경색 심근 조직의 복원은 심혈관 질환에 집중적으로 조사 치료 전략에 속한다. 그러나, 임상으로 실험 결과의 번역은 다양한 병태 생리 학적 조건에서 인체의 복잡한 세포의 상호 작용 주어, 여전히 도전이다. 또한, 심근 난 / R 부상 다양한 사이토 카인, 호르몬과 성장 FAC의 과도한 분비를 유발 혈관 형성 (13)의 영역에 내피 전구 세포의 유도를 제어 TORS. 이미 도시 된 바와 같이, CXCL8, 간질 세포 유래 인자 1α (SDF-1α, CXCL12)은 VEGF 및 대 식세포 이동 억제 인자 (MIF)를 크게 심근 I / R 손상 한 다음 혈청 샘플에서 증가된다. 이들 요소 중에서, MIF 주로 친 염증 특성을 갖는다면 발현 케모카인과 같은 사이토 카인이다. 역사적 이름 달리 MIF는 다양한 세포 유형 1 (21, 22)에 대한 진정한 케모카인으로 작용 친 이동성 기능을 갖는다. MIF 매개 세포 채용 프로세스 MIF는 결합 및 비 - 동족 방법 (21)에 활성화 케모카인 수용체 CXCR4 및 CXCR2에 연결되어있다. 참고로, 내피 전구 세포는 또한 저산소 조건 하에서 상향 조정되고 그 표면에 수용체를 모두 표현엉덩이 = "외부 참조"> 23, 24. 또한, 축적 증거는 MIF 심장 22, 25, 26의 I / R 손상시 전체 심장 보호 효과가 있음을 시사한다. 이러한 맥락에서, 상기 MIF는 손상된 심근 (27)의 제한된 복구 메커니즘을 고려하면, 특히 관련이있다 저산소 스트레스 동안의 혈관 신생을 지원할 수 있음을 보여왔다. 전 임상 마우스 모델에서 이전 체외 연구와 실험은 내피 전구 세포 모집 4 MIF의 역할에 대해 첫 번째 증거를 제공했다. 참고로, MIF는 허혈성 사이트 (28) 내에서 내피 전구 세포 모집 중에 해제 될 수 내피 전구 세포의 저명한화물 단백질이다. 그러나, 다른 (혈관) 혈청 사이토 카인에 비해 특히 임상에서 연구하기 어려운 남아있다.
Access restricted. Please log in or start a trial to view this content.
프로토콜
내피 전구 세포의 분리를위한 혈액은 지역 윤리위원회에 따라 동의 후 건강한 자원자로부터 얻은 것입니다. 마이그레이션 분석에 사용 된 혈청 샘플 심폐 (CPB)를 이용하여 종래의 심장 수술을받은 환자로부터 얻었다. 제외 기준은 비상 운영, 알려진 또는 의심되는 임신했다 18 년 이상 이하 연령, 동의를 얻기 위해 실패를 patient`s. 혈청 샘플은 임상 측정 루틴 (즉시 수술 전 즉시 심근 재관류 / 대동맥의 개시 후) 후속 최종 분석까지 -80 ℃에서 보관 이외에 그려졌다. 제도적 검토위원회 (윤리위원회, RWTH 아헨 대학)이 연구를 승인했다. 환자는 68.6 년의 평균 연령 81.7 kg의 평균 중량을 나타내었다. 기존 질환 포함 고혈압 (65 %), 만성 폐 질환 (19 %), 추가 심장 arteriopathy (16 %), 뇌 기능 장애 (6 %), 불안 정형 협심증 (3 %), 최근 심근 경색 (90 (D) 내에 28 %), 만성 신장 질환 (14 %), 간 질환 (2 %), 당뇨병 (34 %).
T75 플라스크 1. 코팅 :
- 5 ㎖의 피브로넥틴 용액을 T75 플라스크 당 (15ml의 초순수로 희석하여 1 mg의 인간 피브로넥틴)을 준비한다.
- T75 플라스크에 솔루션을 추가하고 물이 증발 될 때까지 기다립니다. 증발에 의한 분리 공정 동안 불필요한 중단을 방지하기 위해, 사전에이 프로세스를 수행하는 (예를 들면 밤새) 실온에서. 이 솔루션은 4.44 μg의 피브로넥틴 / ㎠를 제공합니다.
- 제조자의 성장 인자로 보충 기저 MV2 매체를 보충하여 내피 세포 성장 배지 MV2를 준비한다.
60 mL의 혈액에서 내피 전구 세포 (내피 전구 세포) 2. 절연 :
참고 : CD34 양성 선택 칵테일 A와ND 자성 비드는 제조자가 제공되는 농도가없는, 선택지의 선택 세트에서 상업적으로 이용 가능하다. 그러나, 항체는 약 100 μL를 5 × 108 개의 셀을 처리하기에 충분하다. 자기 구슬이 물에 희석, 덱스 트란이 코팅 된 약 5,000 배 더 작은 다른 시중에서 판매하는 구슬에 비교했다. 자세한 내용은 제조자의 지침을 참조하십시오.
- 칼슘 -mg 2 + - 무료 PBS로 1 : (또는 항응고제 제외) 혈액 1 섞는다.
- 50 ㎖ 튜브 당 밀도 구배 용액 15 mL를 넣고. 자세한 내용은 제조자의 지침을 참조하십시오.
- 천천히 밀도 구배 용액의 상부에 희석 된 혈액 층을 포함한다.
- 원심 분리기 샘플 속도가 느린 가속 및 제동없이 30 분 동안 2,500 XG에.
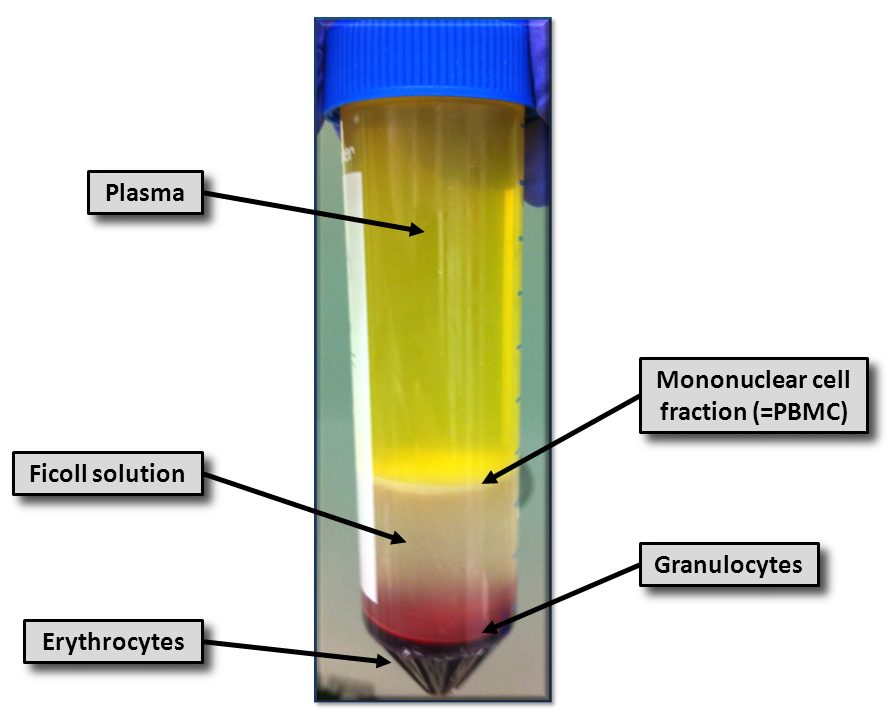
- 조심스럽게 멸균 플라스틱 피펫을 사용하여 모든 관 (그림 1)의 버피 코트 층을 수집하고 다른 튜브에 넣어.밀도 구배 솔루션의 컬렉션을 피하십시오. 예상 PBMC 수율은 약 3-4 × 10 6 세포 / mL의 혈액이다.

그림 1 : 피콜 솔루션에 Bbuffy 코트의 밀도 구배 원심 분리. 밀도 구배 용액에 30 분 동안 2,500 XG에서 밀도 구배 원심 분리의 결과가 도시되어있다. 적혈구 및 과립구 (적색), 단핵 세포 분획 (백색 층) 플라즈마 (옐로우), 및 밀도 구배 용액 (백탁)이다 묘사했다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- PBS 중 적어도 3 볼륨으로 말초 혈액 단핵 세포 분획을 희석하고 혼합한다. 200 x g에서 15 분 동안 실온에서 원심 분리한다.
- 두 번 단계를 반복 2.6.
- 5 ㎖ MV2 배지 상등액과 세포 펠렛 재현 탁 대기음.
- 사용 버피 코트 당 (5 × 10 8 셀을 처리하기위한 충분한) 인간 CD34 항체의 100 μL를 추가하고 5 % CO 2와 37 ° C에서 15 분 동안 회전합니다.
- 사용 버피 코트 당 덱스 트란 코팅 자기 구슬 50 μL를 추가하고 5 % CO 2와 37 ° C에서 10 분 동안 회전합니다.
- 부화 후, 각각의 튜브에 3 mL를 최대 FACS 튜브에 정지를 전송합니다. 더 많은 양의 자석으로 허용되지 않습니다.
- 자석 (들)에 FACS 튜브 (들)을 넣고 5 분 동안 기다립니다.
- 자석 밖으로 FACS 튜브를 당기없이 상층 액을 버린다.
- 자석의 외부에 3 ㎖ MV2 매체 각각 FACS 튜브에 세포를 재현 탁.
- 반복 두 번 2.12-2.14 단계를 반복합니다.
- 자석 밖으로 FACS 튜브를 당겨 3 ㎖ MV2 배지에서 세포를 재현 탁.
- 프레 코트 T75의 F에 세포 현탁액을 전송lasks 및 플라스크 당 17 mL의 MV2 매체를 추가 할 수 있습니다.
주 : 매체 3 일 후 변경해야합니다. 일주 후, 세포는 스핀들 같은 구조 (그림 2)를 표시하고 사용할 준비가 된 것입니다. 분리의 간과를 들어 그림 3을 참조하십시오.

그림 2 : 격리 된 내피 전구 세포의 현미경 이미지. 분리에 앞서 T75 플라스크에서 격리 된 초기 내피 전구 세포의 대표 이미지가 보여진다. 겉보기은 내피 전구 세포의 스핀들 형 구조이다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3 : 흐름도는 EPC 격리 절차를 표시합니다. 도시는 방식 쇼EPC를 분리 프로토콜의 단일 단계를 보내고. 자세한 프로토콜에 대한 프로토콜 섹션의 제 2 장을 참조하십시오.
3. 마이그레이션 분석
- T75 플라스크에 내피 전구 세포에서 매체를 제거합니다.
- 조심스럽게 흔들어 5 mL의 PBS로 세척.
- PBS를 제거하고 5 ㎖ 상업 세포 분리 솔루션을 추가 할 수 있습니다. 세포가 분리 될 때까지 (현미경으로 확인) 기다립니다. 조심스럽게 플라스크의 바닥을 탭하여 분리를 가속화합니다.
- 세포가 분리되는 경우, 신속 MV2 완전 배지 5 ㎖를 추가하여 다른 튜브에 세포 현탁액을 전송.
- 5 분 동안 2,000 XG에 원심 분리기.
- 5 분 동안 2000 × g으로 다시 50-10 ㎖의 PBS에서 세포를 재현 탁 및 원심 분리기.
- 단계를 반복 3.6.
- MV2 결핍 배지에서 세포 펠릿 (윤회 웰 당 75 μL가 필요한 50,000 세포)에 재현 탁.
주 : 필요한 이동 시스템은 세포 형태 및 분석에 따라 달라진다. 사랑하는 있습니다 erent 기공 직경 및 판 크기 (우물의 수). 내피 전구 96 웰 시스템에 5 ㎛의 공극 크기가 최적이다. - 하부 챔버 (그림 4)에 : 혈청 샘플의 235 μL를 추가하여 마이그레이션 판을 준비 (MV2 굶어 매체에 5 혈청 1 희석).

그림 4 : 수정 된 보이든 상공 회의소에서 마이그레이션 분석. 수정 된 보이든 챔버의 일반적인 설계가 도시. 세포 배양 인서트 (= 상부 챔버) 진한 청색으로 표시되고, 상기 하부 챔버 내에 삽입된다. 이 인서트의 바닥 (그러나 벽)은 공극을 포함하는 필터 시스템을 나타낸다. (a)는 시점 제로 설정을 표시합니다. (b) 선택된 시점 후에, 세포를 자극쪽으로 필터를 통해 이주된다.fig4large.jpg "대상 ="_ 빈 ">이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
- 얼마 전에 세포 용액을 첨가 삽입을 추가합니다.
- 상부 챔버 내로 세포 용액 75 μL (= 50,000 세포)를 추가한다.
- 내피 전구 세포 2 (이동 시간은 세포 유형 및 시스템에 따라 다름) 37 ° C에서 3 시간 5 % CO에 대한 마이그레이션 할 수 있습니다.
- 혈청 자동 형광 유물을 방지하기 위해, 반 자동화 된 소프트웨어 "ImageJ에"를 사용하여 세포를 현미경으로 잘 사진을 복용하여 마이그레이션 세포를 정량화 계산합니다.
- 상부 챔버 (모든 이주되지 않은 세포를 포함)를 제거합니다.
- (1,000 1 희석) 훽스트 염료를 포함, 70 μL 3.6 % 파라 포름 알데히드 용액을 추가합니다. 플레이트를 37 ℃ 및 5 % CO 2에서 밤새 저장 될 수있다.
- 1 ~ 2 분 동안 2,000 XG에서 같은 초점면에 모든 세포를 얻기 위해 곧 접시를 원심 분리기.
- w 당 5 사진을 촬영엘 (도 5 및 6) 100 배 배율.

그림 5 : 촬영 한 사진의 위치를 표시 제도. 계획은 잘 관련 마이그레이션 세포의 판정에 필요한 다섯 찍은 사진의 위치를 나타낸다. 사진은 세포를 계산하고 잘 모든에 대한 평균 값을 계산 가져옵니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 6 : 셀 정량화에 대한 현미경 이미지. 세포 정량 찍은 대표적인 이미지는 보여진다. 세포를 염색 USI을 고정시켰다3.6 % 파라 포름 알데히드에 NG 훽스트 (Hoechst) 염료. 점은 고정 내피 전구 세포를 염색 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- 건강의 국립 연구소에 의해 반 자동화 된 소프트웨어 "ImageJ에"를 사용하여 마이그레이션 된 세포를 계산합니다.
Access restricted. Please log in or start a trial to view this content.
결과
격리 된 내피 전구 세포의 특성
먼저 acLDL의 흡수가 확인되었다뿐만 아니라 단리 된 세포 집단의 표면에 KDR의 발현, 및 CD31. 도 7a에서 보듯이, 고립 된 내피 전구 세포의 85.1 %가 acLDL의 흡수를 보여 CD31를 표명했다. 두 마커에 대한 부정적인 작은 인구있을 것 같습니다 있지만,도 7B 및 7C는 상?...
Access restricted. Please log in or start a trial to view this content.
토론
본 연구의 첫 번째 부분은 심장 수술 환자의 혈액의 포괄적 인 평가를 가능하게 건강한 지원자의 말초 혈액에서 인간의 내피 전구 세포의 분리를 포함했다. 따라서, 밀도 구배 원심 분리 혈장, 적혈구 및 과립구로부터 PBMC의 분획을 분리 하였다. 오염 혈소판의 대부분을 제거하려면,이 세포 분획 짧은 하였다 느린 세척, (30)를 29 단계를 반복합니다. 다음...
Access restricted. Please log in or start a trial to view this content.
공개
저자는 공개 아무것도 없어.
감사의 말
저자는 어떤 승인이 없습니다.
Access restricted. Please log in or start a trial to view this content.
자료
| Name | Company | Catalog Number | Comments |
| Fibronectin | Biochrom AG | L7117 | Coating of T-75 flasks |
| Aqua ad iniectabilia | Fresenius Kabi | ||
| Endothelial cell growth medium MV2 | Promo Cell | C-22221 | |
| Endothelial cell growth medium MV2 SupplementMix | Promo Cell | C-39226 | |
| Ficoll-Paque plus | GE Healthcare | 17-1440-03 | Density centrifugation |
| EasySep human CD34 positive Selection Kit | Stemcell Technologies | 18056 | Isolation of CD34+ cells |
| EasySep magnet | Stemcell Technologies | 18000 | |
| Accutase | Sigma-Aldrich | A6964-100ML | Detachment of cells |
| Corning HTS transwell 96 well permeable supports | Sigma-Aldrich | CLS3387-8EA | Migration system |
| Hoechst solution | ThermoFisher | 33342 | Staining of migrated cells |
| ImageJ | National institutes of health | xxx | Counting of migrated cells |
참고문헌
- Emontzpohl, C., et al. Key role of MIF in the migration of endothelial progenitor cells in patients during cardiac surgery. Int J Cardiol. 181C, 284-287 (2014).
- Smadja, D. M., et al. Interleukin 8 is differently expressed and modulated by PAR-1 activation in early and late endothelial progenitor cells. J Cell Mol Med. 13 (8B), 2534-2546 (2009).
- Asahara, T., et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 275 (5302), 964-967 (1997).
- Simons, D., et al. Hypoxia-induced endothelial secretion of macrophage migration inhibitory factor and role in endothelial progenitor cell recruitment. J Cell Mol Med. 15 (3), 668-678 (2011).
- Dernbach, E., et al. Antioxidative stress-associated genes in circulating progenitor cells: evidence for enhanced resistance against oxidative stress. Blood. 104 (12), 3591-3597 (2004).
- Yang, Z., et al. Paracrine factors secreted by endothelial progenitor cells prevent oxidative stress-induced apoptosis of mature endothelial cells. Atherosclerosis. 211 (1), 103-109 (2010).
- Asahara, T., Kawamoto, A., Masuda, H. Concise review: Circulating endothelial progenitor cells for vascular medicine. Stem Cells. 29 (11), 1650-1655 (2011).
- Schmidt-Lucke, C., et al. Reduced number of circulating endothelial progenitor cells predicts future cardiovascular events: proof of concept for the clinical importance of endogenous vascular repair. Circulation. 111 (22), 2981-2987 (2005).
- Yoder, M. C. Human endothelial progenitor cells. Cold Spring Harb Perspect Med. 2 (7), a006692(2012).
- Ito, H., et al. Endothelial progenitor cells as putative targets for angiostatin. Cancer Res. 59 (23), 5875-5877 (1999).
- Hill, J. M., et al. Circulating endothelial progenitor cells, vascular function, and cardiovascular risk. N Engl J Med. 348 (7), 593-600 (2003).
- Yoder, M. C., et al. Redefining endothelial progenitor cells via clonal analysis and hematopoietic stem/progenitor cell principals. Blood. 109 (5), 1801-1809 (2007).
- Rohde, E., et al. Blood monocytes mimic endothelial progenitor cells. Stem Cells. 24 (2), 357-367 (2006).
- Rohde, E., et al. Immune cells mimic the morphology of endothelial progenitor colonies in vitro. Stem Cells. 25 (7), 1746-1752 (2007).
- Prokopi, M., et al. Proteomic analysis reveals presence of platelet microparticles in endothelial progenitor cell cultures. Blood. 114 (3), 723-732 (2009).
- Voyta, J. C., Via, D. P., Butterfield, C. E., Zetter, B. R. Identification and isolation of endothelial cells based on their increased uptake of acetylated-low density lipoprotein. J Cell Biol. 99 (6), 2034-2040 (1984).
- Koch, S., Claesson-Welsh, L. Signal transduction by vascular endothelial growth factor receptors. Cold Spring Harb Perspect Med. 2 (7), a006502(2012).
- Hirschi, K. K., Ingram, D. A., Yoder, M. C. Assessing identity, phenotype, and fate of endothelial progenitor cells. Arterioscler Thromb Vasc Biol. 28 (9), 1584-1595 (2008).
- Hur, J., et al. Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arterioscler Thromb Vasc Biol. 24 (2), 288-293 (2004).
- Tagawa, S., et al. Determination of Early and Late Endothelial Progenitor Cells in Peripheral Circulation and Their Clinical Association with Coronary Artery Disease. Int J Vasc Med. , 2015(2015).
- Bernhagen, J., et al. MIF is a noncognate ligand of CXC chemokine receptors in inflammatory and atherogenic cell recruitment. Nat Med. 13 (5), 587-596 (2007).
- Stoppe, C., et al. Interaction of MIF Family Proteins in Myocardial Ischemia/Reperfusion Damage and Their Influence on Clinical Outcome of Cardiac Surgery Patients. Antioxid Redox Signal. 23 (11), 865-879 (2015).
- Kanzler, I., et al. Differential roles of angiogenic chemokines in endothelial progenitor cell-induced angiogenesis. Basic Res Cardiol. 108 (1), 310(2013).
- Walenta, K. L., Bettink, S., Bohm, M., Friedrich, E. B. Differential chemokine receptor expression regulates functional specialization of endothelial progenitor cell subpopulations. Basic Res Cardiol. 106 (2), 299-305 (2011).
- Rassaf, T., Weber, C., Bernhagen, J. Macrophage migration inhibitory factor in myocardial ischaemia/reperfusion injury. Cardiovasc Res. 102 (2), 321-328 (2014).
- Stoppe, C., et al. High postoperative blood levels of macrophage migration inhibitory factor are associated with less organ dysfunction in patients after cardiac surgery. Mol Med. 18, 843-850 (2012).
- Amin, M. A., et al. Migration inhibitory factor mediates angiogenesis via mitogen-activated protein kinase and phosphatidylinositol kinase. Circ Res. 93 (4), 321-329 (2003).
- Kupatt, C., et al. Embryonic endothelial progenitor cells expressing a broad range of proangiogenic and remodeling factors enhance vascularization and tissue recovery in acute and chronic ischemia. FASEB J. 19 (11), 1576-1578 (2005).
- Colotta, F., et al. Expression of a monocyte chemotactic cytokine by human mononuclear phagocytes. J Immunol. 148 (3), 760-765 (1992).
- Casale, T. B., Kaliner, M. A rapid method for isolation of human mononuclear cells free of significant platelet contamination. J Immunol Methods. 55 (3), 347-353 (1982).
- Lewandowska, K., Kaplan, D., Husel, W. CD34 expression on platelets. Platelets. 14 (2), 83-87 (2003).
- Stellos, K., et al. Platelet-derived stromal cell-derived factor-1 regulates adhesion and promotes differentiation of human CD34+ cells to endothelial progenitor cells. Circulation. 117 (2), 206-215 (2008).
- Thornton, M. A., Poncz, M. In vitro expansion of megakaryocytes from peripheral blood hematopoietic progenitors. Methods Mol Med. 31, 337-345 (1999).
- Ivetic, N., et al. Producing megakaryocytes from a human peripheral blood source. Transfusion. 56 (5), 1066-1074 (2016).
- Friedrich, E. B., Walenta, K., Scharlau, J., Nickenig, G., Werner, N. CD34-/CD133+/VEGFR-2+ endothelial progenitor cell subpopulation with potent vasoregenerative capacities. Circ Res. 98 (3), e20-e25 (2006).
- Wijelath, E. S., et al. Novel vascular endothelial growth factor binding domains of fibronectin enhance vascular endothelial growth factor biological activity. Circ Res. 91 (1), 25-31 (2002).
- Hulkower, K. I., Herber, R. L. Cell migration and invasion assays as tools for drug discovery. Pharmaceutics. 3 (1), 107-124 (2011).
- Yao, E. H., et al. Effects of the antioxidative beta-blocker celiprolol on endothelial progenitor cells in hypertensive rats. Am J Hypertens. 21 (9), 1062-1068 (2008).
- Takahashi, T., et al. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med. 5 (4), 434-438 (1999).
- Kawamoto, A., et al. Therapeutic potential of ex vivo expanded endothelial progenitor cells for myocardial ischemia. Circulation. 103 (5), 634-637 (2001).
- Kim, B. S., et al. Myocardial Ischemia Induces SDF-1alpha Release in Cardiac Surgery Patients. J Cardiovasc Transl Res. 9 (3), 230-238 (2016).
- Frangogiannis, N. G., Smith, C. W., Entman, M. L. The inflammatory response in myocardial infarction. Cardiovasc Res. 53 (1), 31-47 (2002).
- Zernecke, A., Bernhagen, J., Weber, C. Macrophage migration inhibitory factor in cardiovascular disease. Circulation. 117 (12), 1594-1602 (2008).
- White, D. A., et al. Pro-inflammatory action of MIF in acute myocardial infarction via activation of peripheral blood mononuclear cells. PLoS One. 8 (10), e76206(2013).
Access restricted. Please log in or start a trial to view this content.
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유