
Method Article
Agroinoculação direta de Mudas de Milho por Injeção com Vírus mosaico de rabo-de-cavalo recombinante e clones infecciosos do vírus do mosaico de cana-de-açúcar
* Estes autores contribuíram igualmente
Neste Artigo
Resumo
Um protocolo de injeção à base de agrobacterium (agroinjeção) é apresentado para a inoculação do vírus do mosaico de rabo-de-raposa e do vírus do mosaico de cana-de-açúcar em mudas de milho. A inoculação dessa forma leva à infecção viral, silenciamento genético induzido por vírus de genes marcadores e superexpressão viral de GFP.
Resumo
As abordagens de inoculação baseadas em agrobacterium são amplamente utilizadas para introduzir vetores virais em tecidos vegetais. Este estudo detalha um protocolo para a injeção de mudas de milho perto do tecido meristemático com Agrobacterium carregando um vetor viral. Clones de mosaico de cauda de raposa recombinante (FoMV) projetados para silenciamento genético e expressão genética foram usados para otimizar este método, e seu uso foi expandido para incluir um vírus de mosaico de cana recombinante (SCMV) projetado para expressão genética. Fragmentos genéticos ou sequências de codificação de interesse são inseridos em um genoma viral modificado e infeccioso que foi clonado no vetor binário T-DNA plasmid pCAMBIA1380. As construções plasmídeos resultantes são transformadas em tumefaciens agrobacterium tumefaciens strain GV3101. Mudas de milho com até 4 dias de idade podem ser injetadas perto do nó coleoptilar com bactérias resuspended na solução MgSO4. Durante a infecção pelo Agrobacterium, o T-DNA que carrega o genoma viral é transferido para células de milho, permitindo a transcrição do genoma rna viral. À medida que o vírus recombinante se replica e se espalha sistematicamente por toda a planta, sintomas virais e alterações fenotípicas resultantes do silenciamento da lesão dos genes-alvo imitam 22 (les22) ou fitoene desaturase (pds) podem ser observados nas folhas, ou expressão de proteína fluorescente verde (GFP) pode ser detectada após a iluminação com luz UV ou microscopia de fluorescência. Para detectar o vírus e avaliar a integridade da inserção simultaneamente, o RNA é extraído das folhas da planta injetada e o RT-PCR é conduzido usando primers flanqueando o local de clonagem múltipla (MCS) carregando a sequência inserida. Este protocolo tem sido usado efetivamente em vários genótipos de milho e pode ser facilmente expandido para outros vetores virais, oferecendo assim uma ferramenta acessível para introdução de vetores virais no milho.
Introdução
Clones infecciosos de muitos vírus vegetais foram projetados para silenciamento genético induzido por vírus (VIGS), superexpressão genética (VOX) e, mais recentemente, edição de genes habilitado para vírus (VEdGE)1,2,3,4,5,6,7,8,9,10,11 . À medida que novas construções virais são desenvolvidas, métodos para infectar com sucesso tecidos vegetais com esses vírus modificados também devem ser considerados. Os métodos atuais para lançar infecções por vírus em plantas incluem bombardeio de partículas, rub-inoculação de transcrições in vitro de RNA ou clones de DNA, inoculação de punção vascular ou inoculação de tumefaciens agrobacterium (agroinoculação)5,12,13,14,15,16,17 . Cada um desses métodos de inoculação tem vantagens e desvantagens inerentes, que incluem custo, necessidade de equipamentos especializados e viabilidade dentro de um determinado sistema de vírus vegetal. Métodos que utilizam infiltração ou injeção de cepas de Agrobacterium contendo construções binárias de T-DNA projetadas para fornecer vírus recombinantes são preferidos, pois são simples e baratos. No entanto, faltam métodos detalhados de agroindústria para espécies monocotíces, como zea mays (milho).
Um dos primeiros relatos de agroinoculação para entrega de vírus foi publicado em 1986, quando o genoma do vírus do mosaico de couve-flor (CaMV) foi inserido em uma construção T-DNA, e a Agrobacterium resultante que carregava essa construção foi esfregada em plantas de nabo18. Desde então, foram desenvolvidos métodos adicionais de agroindulação. Por exemplo, no caso do vírus do mosaico de cauda-raposa (FoMV), Nicotiana benthamiana pode ser usada como um hospedeiro intermediário para gerar partículas de vírus em folhas que fornecem uma fonte inóculo6. A inoculação de milho por meio de folhas infectadas de N. benthamiana é eficiente, rápida e simples, mas o uso de um hospedeiro intermediário não funciona para todos os vírus que infectam milho. O vírus do mosaico de cana-de-açúcar (SCMV), por exemplo, não pode infectar n. benthamiana, exigindo o uso de fontes alternativas de inóculo para vetores derivados desse vírus. Em 1988, a Agrobacterium contendo o vírus da raia do milho (MSV), um vírus de DNA, foi introduzida em mudas de milho por injeção (agroinjeção), demonstrando que métodos de inoculação à base de Agrobacterium também são úteis para monocotas19. Apesar desse sucesso precoce com a agroinjeção, poucos estudos utilizando essa técnica em milho foram publicados, deixando em aberto questões sobre a aplicabilidade deste método para vírus RNA e vetores VIGS, VOX e VEdGE20,21,22. No entanto, o amplo uso da agroinjeção em espécies monocotas é promissor, pois essa abordagem geral tem sido utilizada em orquídeas, arroz e trigo23,24,25,26,27,28.
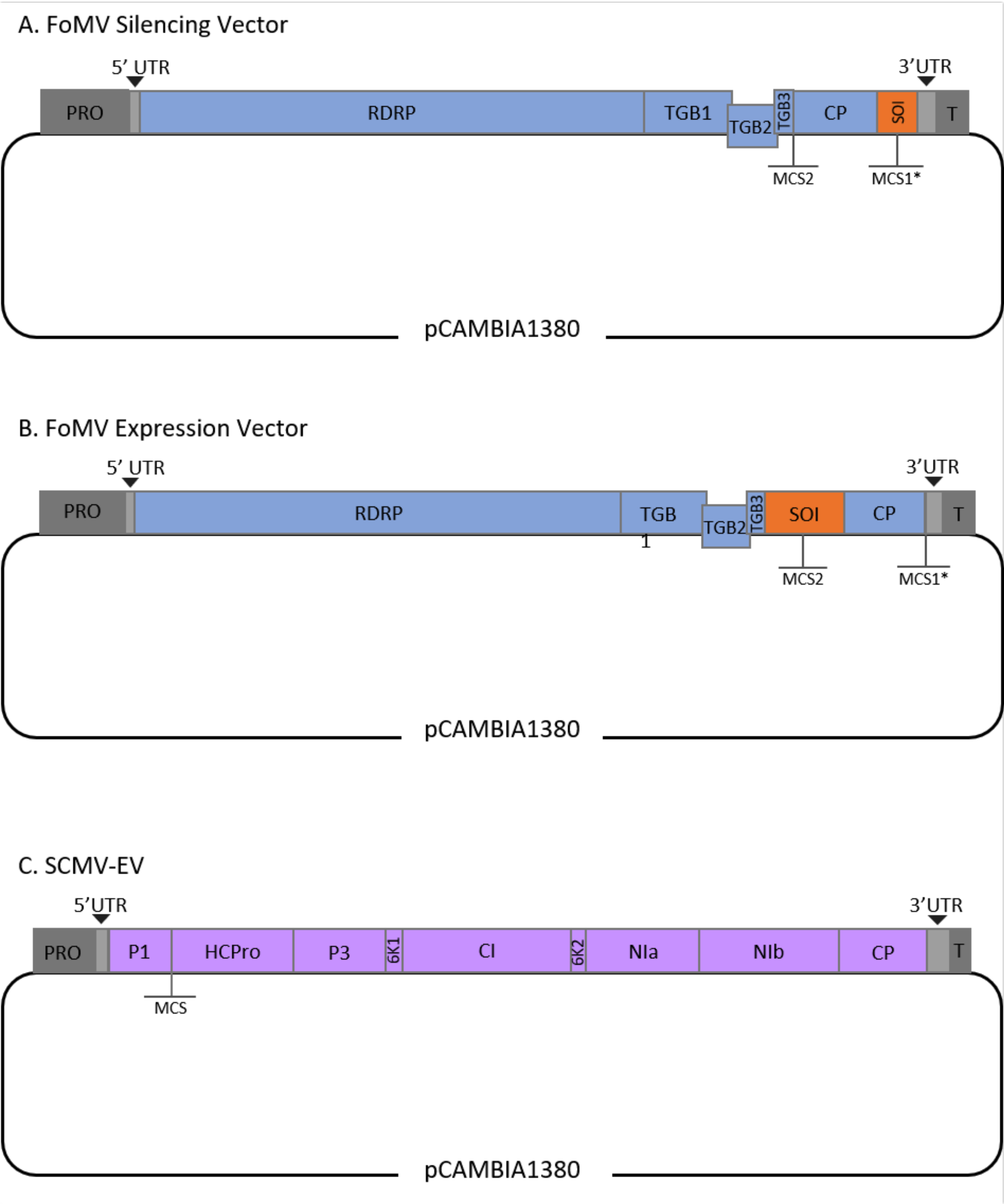
Este protocolo foi otimizado para fomv e agrobacterium strain GV3101 e também foi aplicado a um vetor SCMV. FoMV é um potexvírus com uma ampla gama de hospedeiros que inclui 56 espécies de monocot e dicot29. O FoMV tem um sentido positivo de 6,2 quilobase (kb), genoma de RNA de fio único que codifica cinco proteínas diferentes de cinco quadros de leitura aberto (ORFs)30,31,32,33,34,35. O FoMV foi previamente desenvolvido em um vetor VIGS e VOX para milho, incorporando um clone infeccioso em uma espinha dorsal plasmida de DNA T6,36,37. O genoma viral foi modificado para aplicações VIGS adicionando um local de clonagem (MCS1*) imediatamente a jusante da proteína do casaco (CP) (Figura 1A)36. Para aplicações VOX e VEdGE, o promotor cp foi duplicado e um segundo site de clonagem (MCS2) foi adicionado para permitir a inserção de sequências de interesse entre ORF 4 e o CP (Figura 1B)6. O vetor FoMV contendo MCS1 e MCS2 sem pastilhas é o vetor vazio FoMV (FoMV-EV) (Figura 1).
O SCMV é um vírus não relacionado que foi desenvolvido para VOX no milho38. É membro da família Potyviridae, da qual vários membros foram projetados para expressar proteínas estrangeiras em planta39,40,41,42,43,44. A gama hospedeira de SCMV inclui milho, sorgo e cana-de-açúcar45,46, tornando-o valioso para estudos funcionais genéticos nessas principais plantas agrícolas36,38. O SCMV tem um sentido positivo, genoma de RNA de uma única cadeia de aproximadamente 10 kb de comprimento47,48. Para criar o vetor VOX SCMV, a bem estabelecida junção P1/HCPro foi utilizada como um local de inserção para sequências heterólogas38. Este local de clonagem é seguido por uma sequência codificando um local de decote protease NIa-Pro, levando à produção de proteínas independentes da poliproteína SCMV (Figura 1C).
Plasmídeos de T-DNA que transportam cDNA infeccioso desses vírus recombinantes foram transformados em cepas de Agrobacterium GV3101. GV3101 é uma cepa do tipo nopaline, que é bem conhecida por ser capaz de transferir T-DNA para espécies monocotíces, incluindo milho26,28,49. Além disso, estudos anteriores de agroinjeção utilizaram as cepas C58 ou seu derivado GV3101, bem como 19,20,22,27.
Três genes marcadores foram usados no desenvolvimento deste protocolo: dois para silenciamento genético e um para expressão genética. Um fragmento de 329 pares de base (bp) da lesão do gene do milho imita 22 (les22, GRMZM2G044074) foi usado para construir o vetor de silenciamento FoMV-LES22. Quando les22 é silenciado em milho, pequenas manchas redondas de células necróticas aparecem ao longo da vasculatura de folhas que se expandem e se fundem em grandes áreas de tecido necrosado da folha50. FoMV-PDS, contendo um fragmento de 313 bp do gene de sorgo fitoene desaturase (pds, LOC110436156, 96% de sequência de identidade para pds de milho, GRMZM2G410515), induz o silenciamento de pds em milho, resultando em pequenas faixas de células fotobleachadas ao longo da vasculatura das folhas que alongam ao longo do tempo51. A sequência de codificação intacta para proteína fluorescente verde (GFP) foi usada para demonstrar expressão proteica tanto para FoMV (FoMV-GFP) quanto para SCMV (SCMV-GFP). A expressão GFP nas folhas é tipicamente mais detectável aos 14 dias após a inoculação (DPI)6. Embora tenha havido estudos anteriores utilizando a agroinjeção de vetores virais no milho, esses experimentos só mostraram que a agroinjeção pode facilitar a infecção viral de um clone infeccioso em mudas de milho e não se expandir para vírus recombinantes projetados para aplicações VIGS ou VOX19,20,21,22. O protocolo aqui apresentado baseia-se em métodos anteriores de agroinjeção, particularmente Grismley et al.19. No geral, este método de agroinjeção é compatível com vetores VIGS e VOX, não requer equipamentos especializados ou hosts alternativos como fontes inóculos, e diminui o tempo e o custo total necessários para configurar e realizar inoculações em relação a outros métodos comuns que requerem biolística ou transcrição in vitro. Este protocolo facilitará estudos de genômica funcional em milho com aplicações envolvendo VIGS, VOX e VEdGE.
Protocolo
1. Construção plasmid
NOTA: Este protocolo pode ser aplicado a outros vetores virais ou cepas de Agrobacterium , mas isso pode afetar o sucesso geral da inoculação por agroinjeção. Sempre realize etapas de inoculação bacteriana e revestimento em uma coifa de fluxo laminar.
- FoMV silenciando construção
NOTA: A mídia Luria-Bertani (LB) (Miller) é usada para todos os meios de comunicação, a menos que seja especificado de outra forma. O LÍQUIDO LB é feito suspendendo 25 g de grânulos em 1.000 mL de água destilada e autoclaving por 15 min a 121 °C. A mídia LB sólida é igualmente feita com a adição de 1,5% de ágar antes da autoclavagem. Os antibióticos são adicionados depois que a LB é resfriada a ~60 °C, e a solução é derramada em placas de Petri de 95 x 15 mm. As concentrações de antibióticos a serem utilizados são as seguintes: rifampicina (rif) a 25 μg/mL, gentamicina (gent) a 50 μg/mL e kanamycin (kan) a 50 μg/mL.- PCR amplifica fragmentos do gene do milho a serem silenciados (por exemplo, les22 ou pds) usando um primer avançado com um site de restrição PacI e um primer reverso com um site de restrição XbaI. Isso permitirá a ligadura dos fragmentos genéticos no MCS1* do vetor binário FoMV-pCAMBIA1380 na orientação antissamparada.
NOTA: Configure o PCR usando um polimerase de DNA de alta fidelidade, primers dianteiros e reversos a 10 μM cada, modelo de DNA plasmid e água, seguindo as especificações de polimerase de DNA. Amplie por 35 ciclos, usando uma temperatura de ressarificação de acordo com a polimerase de DNA e a temperatura de fusão do primer (Tm), e uma extensão de 30 s por quilobase a ser amplificada. - Realize a purificação do PCR usando um kit de purificação pcr de acordo com as especificações do kit.
- Digerir o produto PCR purificado e o FoMV-EV com as enzimas de restrição XbaI e PacI. Use 1 μg de plasmídeo ou todo o produto PCR purificado, 2 μL de tampão de 10x, 1 μL de enzima de restrição e adicione água para fazer um volume final de reação de 20 μL. Incubar de acordo com a especificação enzimápica.
- Ligate o produto PCR digerido e FoMV-EV juntamente com liga ligase de DNA T4 de acordo com o protocolo do fabricante.
- Transforme o plasmídeo ligado em células E. coli quimicamente competentes usando o método de choque térmico.
- Descongele as células no gelo e adicione 3 μL de plasmid ao tubo. Incubar no gelo por 30 minutos, depois choque térmico para 30 s a 42 °C.
- Coloque no gelo por 5 min, adicione 200 μL de caldo super ótimo com repressão catabólica (SOC) e permita que as células E. coli se recuperem em mídia SOC por 1 h a 37 °C com agitação a 225 rpm.
- Placa na mídia lb seletiva de kanamicina e incubar a 37 °C durante a noite.
- Verifique as colônias quanto a clones precisos por sequenciamento Sanger usando os primers FM-5840F e FM-6138R (Tabela Suplementar 1). Envie 250 ng de DNA plasmídeo para uma instalação que realizará sequenciamento Sanger. Para este experimento, amostras foram enviadas para o Centro de DNA da Universidade Estadual de Iowa.
- Inocular 2 mL de LÍQUIDO LB com a colônia escolhida e incubar a 37 °C durante a noite com agitação a 225 rpm. Extrair DNA plasmídeo da cultura da noite para o dia através de uma preparação de DNA plasmídeo alcalino lysmid 52.
- Transforme o DNA plasmídeo em células de cepa de agrobacterium GV3101 usando o método degelo congelante. Permita que 100 μL de células quimicamente competentes descongelem no gelo, adicionem 1-5 μL de plasmídeo e incubam no gelo por 30 minutos. Coloque em nitrogênio líquido por 1 min e incubar a 37 °C por 3 min. Adicione 1 mL de SOC, deixe se recuperar por 2-3 h a 28 °C com agitação, placa em rif, gent e kan selective LB media e incubar a 28 °C por 2 dias.
- Colônias de tela para a presença de inserção com a colônia PCR. Escolha uma única colônia bacteriana e misture-a em 30 μL de água. Configure uma reação pcr adicionando 12,5 μL de mistura mestre de polimerase, 1,25 μL de cada primer de 10 μM, FM-5840F e FM-6138R, 3 μL da suspensão da colônia bacteriana e água para um volume final de 25 μL. Ciclo 35 vezes com uma temperatura de renasamento de 64 °C e um tempo de extensão de 1 min (1 min para cada kb amplificado).
- Inocular 2-5 mL de LÍQUIDO LB (rif, gent, kan) com a colônia de Agrobacterium correta. Deixe crescer durante a noite a 28 °C com tremor a 225 rpm.
- Misture a cultura da noite para o dia com uma solução de 50% de glicerol 1:1. Armazene a -80 °C para armazenamento a longo prazo.
- PCR amplifica fragmentos do gene do milho a serem silenciados (por exemplo, les22 ou pds) usando um primer avançado com um site de restrição PacI e um primer reverso com um site de restrição XbaI. Isso permitirá a ligadura dos fragmentos genéticos no MCS1* do vetor binário FoMV-pCAMBIA1380 na orientação antissamparada.
- Construção de expressão FoMV
- O PCR amplifica a sequência de codificação de interesse, incluindo códons de início e parada (por exemplo, GFP) conforme descrito no 1.1.1, adicionando um site de restrição Bsu36I no primer dianteiro e um site de restrição PspOMI no primer reverso para permitir a clonagem direcional na orientação de sentido no MCS2.
- Realize a purificação do PCR usando um kit de purificação pcr de acordo com as especificações do kit.
- Digerir o produto PCR e o FoMV-EV com as enzimas de restrição Bsu36I e PspOMI, conforme descrito em 1.1.3.
- Ligate o produto PCR digerido e FoMV-EV juntamente com liga ligase de DNA T4 de acordo com o protocolo do fabricante.
- Transforme-se em células E. coli quimicamente competentes usando o método de choque térmico como descrito em 1.1.5. Placa na mídia lb seletiva de kanamicina e incubar a 37 °C durante a noite.
- Verifique as colônias para obter clones precisos por sequenciamento Sanger, conforme descrito em 1.1.6 usando os primers 5AmuS2 e 5AmuA2 (Tabela Suplementar 1).
- Inocular 2 mL de LÍQUIDO LB com a colônia escolhida e incubar a 37 °C durante a noite com agitação a 225 RPM. Extrair DNA plasmídeo da cultura da noite para o dia através de uma preparação de DNA plasmídeo alcalino lysmid 52.
- Transforme o DNA plasmídeo em células quimicamente competentes GV3101 usando o método de congelamento, conforme descrito em 1.1.8. Placa em mídia rif, gent e kan seletiva LB e incubar a 28 °C por 2 dias.
- Colônias de tela para a presença de inserção com a colônia PCR usando os primers 5AmuS2 e 5AmuA2.
- Inocular 2-5 mL líquido LB (rif, gent, kan) com a colônia de Agrobacterium correta. Agite durante a noite a 225 rpm a 28 °C.
- Misture a cultura da noite para o dia com uma solução de 50% de glicerol 1:1. Armazene a -80 °C para armazenamento a longo prazo.
- Construção de expressão SCMV
- O PCR amplifica o gene de interesse (por exemplo, GFP) excluindo o codon stop como descrito em 1.1.1, incluindo um local de digestão PspOMI no primer dianteiro e um local de digestão SbfI no primer reverso para permitir a clonagem direcional no vetor binário SCMV-pCAMBIA1380.
NOTA: A pastilha deve ser clonada na moldura com a poliproteína viral. - Realize a purificação do PCR usando um kit de purificação pcr de acordo com as especificações do kit.
- Digerir o produto PCR e o SCMV-EV com as enzimas de restrição PspOMI e SbfI, conforme descrito em 1.1.3.
- Ligate o produto PCR digerido e SCMV-EV juntamente com liga ligase de DNA T4 de acordo com o protocolo do fabricante.
- Transforme o produto em células E. coli quimicamente competentes usando o método de choque térmico descrito em 1.1.5. Placa na mídia kan seletiva LB e incubar a 37 °C durante a noite.
- Colônias de tela para clones precisos por sequenciamento Sanger como descrito em 1.1.6 usando os primers SC-745F e HCProR1 (Tabela Suplementar 1).
- Inocular 2 mL de LÍQUIDO LB com a colônia escolhida e incubar a 37 °C durante a noite com agitação a 225 rpm. Extrair o DNA plasmídeo da cultura da noite para o dia através de uma preparação de DNA lino lysmid plasmídeo52.
- Transforme o DNA plasmídeo em células quimicamente competentes GV3101 usando o método de congelamento, conforme descrito em 1.1.8. Placa em mídia rif, gent e kan seletiva LB e incubar a 28 °C por 2 dias.
- Colônias de tela para presença de inserção com pcr colônia com os primers SC-745F e HCProR1 como descrito em 1.1.9.
- Inocular 2-5 mL de LÍQUIDO LB (rif, gent, kan) com a colônia de Agrobacterium correta. Agite durante a noite a 225 rpm a 28 °C.
- Misture a cultura da noite para o dia com uma solução de 50% de glicerol 1:1. Armazene a -80 °C para armazenamento a longo prazo.
- O PCR amplifica o gene de interesse (por exemplo, GFP) excluindo o codon stop como descrito em 1.1.1, incluindo um local de digestão PspOMI no primer dianteiro e um local de digestão SbfI no primer reverso para permitir a clonagem direcional no vetor binário SCMV-pCAMBIA1380.
2. Preparação de mudas
- Plante 1-2 sementes de milho (milho doce 'Golden Bantam', FR1064, B73, etc.) em meio de cultivo à base de turfa em pequenas pastilhas colocadas dentro de bandejas 4-7 dias antes da injeção. Coloque em uma câmara de crescimento abaixo de 16 h dias a 25 °C e 8h noturnas a 22 °C (~185 radiação fotossintética (PAR)) ou em uma estufa abaixo de 16 h dias a 22-25 °C e 8h noites a 22-25 °C (350-400 PAR).
NOTA: A suscetibilidade ao Agrobacterium varia entre os genótipos de milho, afetando as taxas de sucesso. Além disso, alguns vetores virais podem ser incompatíveis com certos genótipos de milho. - Água regularmente e fertilizar uma vez por semana com 15-5-15 fertilizantes líquidos a 330 partes por milhão (PPM).
3. Preparação de Agrobacterium
- Um dia antes da injeção, prepare a mídia líquida LB com o antibiótico apropriado (rif, gent, kan) e inoculada com a cepa Agrobacterium carregando a construção viral desejada. Recomenda-se adicionar 20 μL de estoque de glicerol em 50 mL de LB, o que deve produzir cultura bacteriana suficiente para inocular >100 plantas e pode ser dimensionado para cima ou para baixo conforme necessário.
NOTA: Prepare o inóculo suficiente para ter uma quantidade final de suspensão bacteriana de pelo menos 1 mL a uma densidade óptica de 600 nm (OD600) de 1,0 para cada 4-5 plantas. - Agite a 225 rpm a 28 °C por 24 h.
- Bactérias de pelotas por 10 min a 4.000 x g em temperatura ambiente. Descarte o supernatante.
- Lave bem a pelota com 1 mL de água desionizada (DI) por tubulação ou vórtice suave.
- Repita o passo 3.3 para a bactéria pellet.
- Resuspenda a pelota em 1 mL de solução MgSO4 de 10 mM por pipetação ou vórtice suave.
- Opcionalmente, adicione acetossyringone de 200 μM à solução. Embora comumente utilizado, acetosyringone só aumenta a capacidade de transformação de algumas cepas de Agrobacterium . Os autores não constataram que a adição de acetosiingona afeta a eficiência neste protocolo (Tabela Suplementar 2).
NOTA: A solução MgSO4 de 10 mM pode ser feita a partir de uma solução de estoque de 1 M com um pH de 6,3 armazenados à temperatura ambiente. A solução provavelmente não exigirá ajuste de pH.
- Opcionalmente, adicione acetossyringone de 200 μM à solução. Embora comumente utilizado, acetosyringone só aumenta a capacidade de transformação de algumas cepas de Agrobacterium . Os autores não constataram que a adição de acetosiingona afeta a eficiência neste protocolo (Tabela Suplementar 2).
- Meça OD600 da amostra com um espectrofotômetro e dilua para 1,0 OD600 com solução mgso4 de 10 mM.
NOTA: Este é um ponto de parada seguro. A suspensão bacteriana pode ser mantida em temperatura ambiente por até 5 h antes da injeção.
4. Injeção
NOTA: Podem ser usadas mudas de milho de 4 a 7 dias de idade para injeção. A taxa de crescimento das mudas é muito afetada pelas condições de crescimento, quantidade de PAR (ou seja, par maior em estufa do que na câmara de crescimento) e genótipo, entre outras coisas que podem ser difíceis de controlar em condições de estufa. As plantas podem ser injetadas com até 4 dias de idade quando têm 2-3 cm de altura sem folhas expandidas e tão velhas quanto 7 dias quando a folha de ponta arredondada mais baixa é expandida. A taxa de sucesso desses métodos de inoculação cai rapidamente à medida que as plantas envelhecem além de 7 dias após a semeadura. O local da injeção é o mesmo, não importa a idade das mudas.
- Usando óculos de segurança, injete a suspensão bacteriana nas mudas 2-3 mm acima do nó coleoptilar usando uma agulha de 25G x 5/8" presa a uma seringa descartável de 1 mL.
NOTA: O nó coleoptilar é onde as raízes da coroa eventualmente se formarão. Este é o nó mais baixo da planta. Normalmente, haverá uma mudança de cor de verde para branco dentro e abaixo do nó. O local da injeção é logo acima do meristem. Dissecar algumas mudas nesta fase pode ajudar a visualizar a localização do meristem e, consequentemente, o local de injeção adequado. - Aplique pressão suave na seringa até que a suspensão encha o coleoptile ou seja visível na prostituta, dependendo do estágio de crescimento das plantas. Isto é aproximadamente 100-200 μL de suspensão.
NOTA: Se for difícil injetar a suspensão na muda, o local da injeção pode ser muito baixo. Pressão moderada é tudo o que deve ser necessário para injetar a suspensão. - Injete todas as mudas, trocando seringas e agulhas para cada construção.
5. Cuidados contínuos com plantas
- Transplante as mudas injetadas para 13 x 13 x 15 cm ou potes maiores quando tiverem 7-8 dias de idade.
- Manter as condições de crescimento (fotoperíodo 16h e fertilização uma vez por semana).
6. Confirmação da infecção (Fenotipicamente e RT-PCR)
- Plantas fenotipicamente pontuadas entre 14-21 DPI. Lesões provenientes do silenciamento da lesão dos genes de controle imitam 22 ou fitoene desaturase podem ser facilmente vistas nas folhas e são distintas dos sintomas do FoMV. A expressão GFP pode ser detectada através de imagens de microscópio fluorescente ou outras imagens de luz UV.
NOTA: Alguns construtos/vetores virais podem demorar mais tempo para apresentar sintomas ou podem não apresentar nenhum sintoma. As altas condições de luz aumentam muito os fenótipos causados pelo silenciamento da lesão imitar 22 e desaturar fitoeno. As lesões podem ser menos visíveis ou ausentes se as plantas forem mantidas em condições de luz mais baixas, como uma câmara de crescimento, porém a taxa real de infecção determinada pelo RT-PCR não deve ser afetada (Tabela 1). - Para confirmar a infecção molecularmente, a folha de amostra 6 entre 14-21 DPI e extrair RNA total usando uma extração fenol-clorofórmio de acordo com a instrução do fabricante.
- Usando o RNA extraído como um modelo para gerar cDNA de primeira linha.
- Configure a reação cDNA com até 5 μg de RNA total, 1 μL de primers hexamer aleatórios, 1 μL de oligo (dT)18 primers, 1 μL de dNTPs, 1 μL de transcriptase reversa e água para um volume final de 14,5 μL.
- Usando primers projetados para a construção viral e o cDNA como modelo, execute PCR em cada amostra para confirmar a infecção viral e determine a integridade do gene ou fragmento genético de interesse conforme descrito em 1.1.1, exceto reduzir os ciclos para 25 para FoMV e 30 para SCMV para evitar falsos positivos.
- Para silenciar construções de silenciamento do FoMV, use primers FM-5840F e FM-6138R para amplificar através do MCS1*, que contém o fragmento genético de milho. Para construções de expressão FoMV, use primers 5AmuS2 e 5AmuA2 para amplificar através do MCS2, que contém o gene inserido.
- Para construções de expressão SCMV, utilize primers SC745-F e HCProR1 para amplificar em todo o MCS, que contém o gene inserido (Figura Suplementar 3).
- Para um gene de controle endógeno, use os primers ZmActS e ZmActA, que amplificam um fragmento de mRNA de ato de milho (GRMZM2G126010) ou primers ZmUbiF e ZmUbiR, que amplificam um fragmento mRNA de poliugenquitina de milho (GRMZM2G409726_T01).
- Visualize o produto PCR em um gel de 1% de agarose contendo uma mancha de ácido nucleico para determinar a presença ou ausência de vírus e gene ou fragmento genético.
Resultados
O objetivo deste estudo foi desenvolver um protocolo simples para introduzir diretamente vírus recombinantes projetados para silenciamento genético ou expressão genética em mudas de milho (Figura 2). Os vetores do vírus que carregam pastilhas são projetados e clonados usando técnicas padrão de biologia molecular. Fragmentos genéticos para silenciamento são inseridos no MCS1* no FoMV-EV e sequências de codificação para expressão são inseridas no FoMV-EV no MCS2 ou SCMV-EV no MCS. Os plasmídeos resultantes são transferidos para a cepa Agrobacterium GV3101. Posteriormente, as mudas de milho são injetadas dentro de uma semana ou menos após o plantio. Duas semanas após a injeção, as plantas podem ser avaliadas fenotipicamente e molecularmente para infecção viral, silenciamento genético e expressão genética.
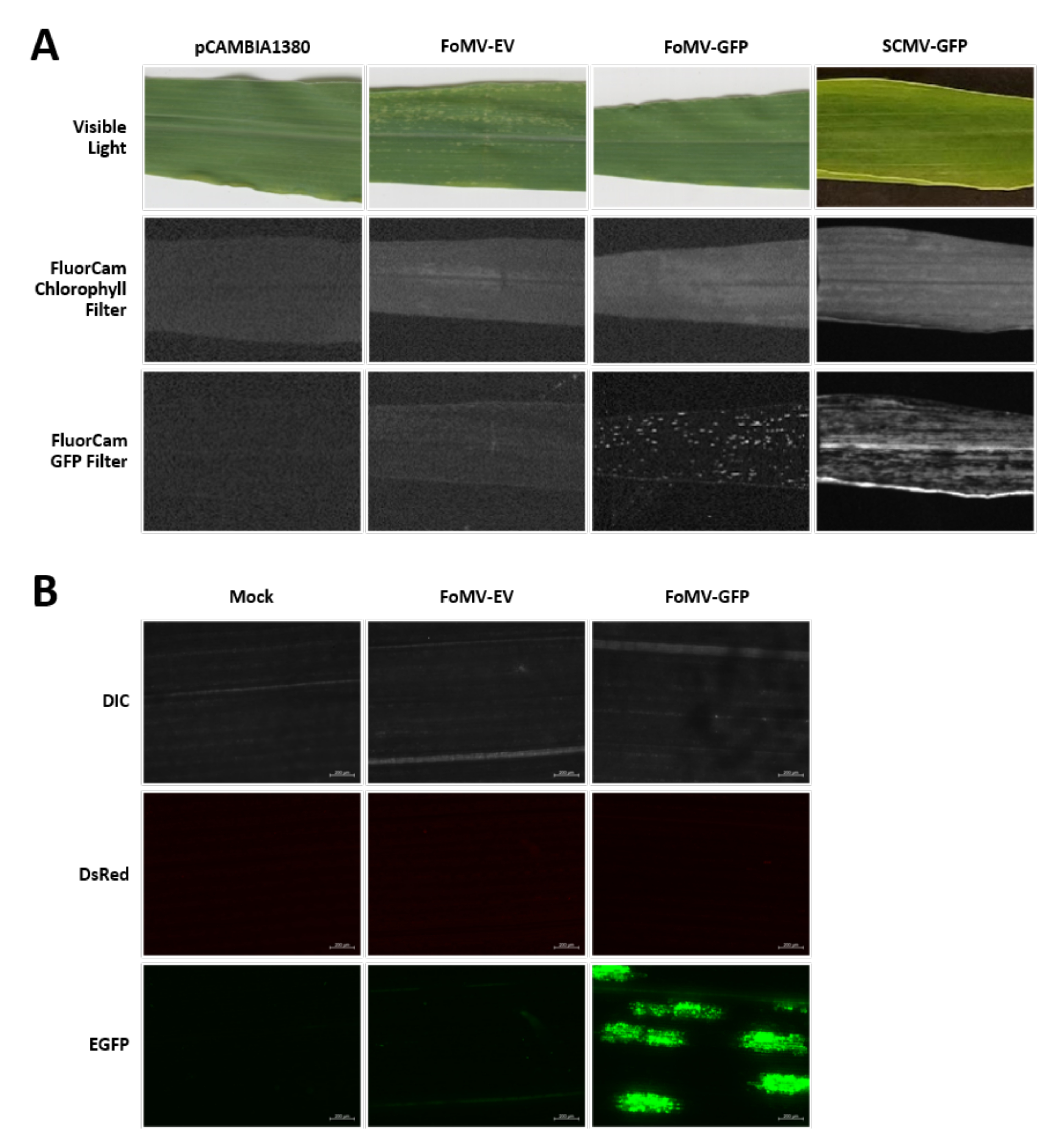
As plantas de milho são cultivadas em um meio à base de turfa por 4-7 dias. Nesta fase, o meristem apical de tiro é logo acima do nó coleoptilar (Figura 3A). Após o coleoptile ter estendido de 2-3 centímetros ou até 7 dias após a semeadura, as plantas são injetadas 2-3 mm acima do nó coleoptilar (Figura 3B-F). Aproximadamente 12 dias após a injeção, as plantas começarão a exibir fenótipos silenciados em suas folhas, comumente observados perto do tecido vascular, e essas lesões são visualmente distintas dos sintomas do mosaico viral do FoMV (Figura 4). Tanto a presença do FoMV quanto o silenciamento de genes-alvo são detectáveis em plantas injetadas (Figura 5). A expressão GFP pode ser detectada até 2 semanas após a injeção sob um microscópio fluorescente e é mais forte nas folhas 5-7 (Figura 6). Quando observada sob um sistema de imagem de fluorescência, a expressão GFP do FoMV pode ser visualizada como muitas pequenas áreas pontuais de fluorescência distribuídas através de folhas próximas ao tecido vascular, enquanto a expressão GFP do SCMV consiste em manchas maiores (Figura 6, Figura Suplementar 1). Embora os sintomas de mosaico viral sejam frequentemente visíveis em plantas infectadas com construções de silenciamento fomv, plantas injetadas com construtos de expressão GFP que expressam com sucesso GFP muitas vezes não têm esses sintomas. Como resultado, uma planta sem sintomas visíveis ainda pode ser positiva para a expressão do vírus e da GFP. Além disso, a puncção do meristem durante o procedimento de agroinjeção deve ser evitada, pois isso pode causar defeitos morfológicos, mas as plantas resultantes sobrevivem e são muitas vezes sintomáticas (Figura 7).
Embora este protocolo tenha sido originalmente desenvolvido usando milho doce, várias linhas de milho podem ser inoculadas com sucesso com construtos de silenciamento genético FoMV usando a agroinjeção. Por exemplo, FR1064 e B73 normalmente têm altas taxas de infecção viral (Tabela 2). Notavelmente, mo17, uma linha com resistência genética conhecida ao FoMV, teve uma eficiência de infecção de 0% como esperado36,53. Além disso, o construto utilizado influencia a eficiência da infecção (Tabela 3). No caso do FoMV, FoMV-EV e FoMV-LES22 normalmente têm as maiores eficiências de infecção em 53% e 54%, respectivamente. O FoMV-PDS tem uma eficiência ligeiramente menor, de 38%, e o FoMV-GFP é o menor em 17%. O SCMV-GFP tem uma eficiência de infecção de 8%. Esses percentuais são médias ao longo de vários experimentos; experimentos individuais podem ter maior ou menor eficiência de infecção.

Figura 1: Representações esquemáticas dos clones FoMV e SCMV T-DNA utilizados para a agroinjeção no milho. O vetor FoMV contém dois vários locais de clonagem (MCS1* e MCS2). O vetor vazio, FoMV-EV, tem 7.269 bp e não contém nenhuma inserção em nenhum dos MCS. (A) O silenciamento genético usando o vetor FoMV pode ser alcançado inserindo fragmentos genéticos no MCS1*, designado como sequência de interesse (SOI), tipicamente na orientação anti-sentido. (B) A expressão genética usando o vetor FoMV pode ser realizada inserindo ORFs genéticos no MCS2 na orientação de sentido, designado como SOI. (C) O vetor SCMV foi projetado para ter um MCS entre P1 e HCPro. O vetor vazio, SCMV-EV, tem 11.015 bp e não contém nenhuma inserção no MCS. OrFs genéticos inseridos no MCS que estão em quadro com a poliproteína SCMV serão expressos como proteínas. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Resumo esquemático do protocolo de agroinjeção. (A) Clone SOI, seja um CDS ou fragmento genético, no vetor viral e transformado em cepa de Agrobacterium GV3101. (B) Plantar milho e crescer por 4-7 dias. (C) Cresça GV3101 na cultura líquida durante a noite a 28 °C. (D) Prepare a suspensão bacteriana para injeção. (E) Injetar mudas 2-3 mm acima do nó coleoptilar com 100-200 μL de suspensão. (F) As mudas de transplante quando têm 7 dias de idade para potes maiores e crescem por 2-3 semanas até que a 5ª folha seja visível. Fenótipo, se desejado. (G) Folha de amostra 5 e extrato de RNA. (H) Faça cDNA e realize o PCR para amplificar o vírus/SOI. (I) Executar em gel para análise qualitativa para determinar presença/ausência de vírus e um SOI truncado ou intacto. Este número foi criado com BioRender.com. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Método de agroinjeção de inoculação de mudas logo acima do nó coleoptile. (A) plantas de 4-5 dias de idade. O coleoptile é totalmente expandido, e a primeira folha verdadeira pode ser parcialmente visível, mas não é desenrolada. (B) plantas de 6 a 7 dias. A primeira folha pode ser expandida, mas nenhuma coleira será visível. A segunda folha também será visível e pode estar começando a se desenrolar nesta fase. (C) Dissecção de plantas de 6 a 7 dias de idade mostrando a localização do meristem apical de tiro em relação ao nó coleoptile. (D) Injeção de plantas de 4 a 5 dias de idade. (E) Injeção de plantas de 6 a 7 dias de idade. (F) Injeção de plantas de 6 a 7 dias de idade usando uma solução de corante, mostrando inóculo tingido saindo da prostituta da muda. (G) Close-up do local de injeção de plantas de 6-7 dias de idade em relação ao nódulo coleoptile. (H) Close-up de uma planta de 6-7 dias de idade pós-injeção, mostrando inóculo tingido na prostituta da planta. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Sintomas dos genes de controle de silenciamento utilizados nos experimentos de agroinjeção. (A) Uma folha fotografada a 17 DPI após a planta ser injetada com FoMV-LES22. O FoMV-LES22 carrega uma inserção de 329 bps do CDS de 3' da lesão imitar 22 gene de milho na orientação antissense. O silenciamento resulta no acúmulo de um metabólito tóxico que, por sua vez, causa as lesões necrosas que aparecem primeiro como raias ao longo da vasculatura e crescem em manchas maiores, como mostrado aqui. (B) Uma folha fotografada em 17 DPI após a planta ser injetada com FoMV-PDS. O FoMV-PDS carrega uma inserção de par de 313 bases do CDS de 3' do gene desaturase de fitoeno de sorgo na orientação antissense. O silenciamento de pds no milho causa um fenótipo de fotobleaching sistêmico que começa como pequenas e finas listras ao longo da vasculatura que crescem em raias mais longas ao longo do comprimento da folha, como mostrado aqui. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: qRT-PCR de plantas injetadas com construtos de silenciamento genético FoMV. Confirmação da infecção sistêmica de FoMV e silenciamento genético induzido pelas construções FoMV-LES22 e FoMV-PDS entregues via agroinjeção em plantas de milho doce (Golden x Bantam). (A) As imagens em gel mostram análises rt-PCR confirmando a presença de vetor vazio FoMV-MCS1* (315 bp amplicon) e FoMV-PDS (amplicon de 625 bp) na folha 6 de cinco plantas individuais. Os primers PCR usados produzem uma amplicon que abrange o MCS1*. O amplicon de gene de milho (Zm-Actin) serve como gene de referência. O gráfico da barra representa os valores de expressão relativa qRT-PCR para expressão de pds na folha 6 aos 37 dias pós-inoculação (DPI) por agroinjeção com FoMV-MCS1* ou FoMV-PDS. A supressão de pds é detectável em cada uma das cinco réplicas biológicas (p=0,003; teste pós hoc Dunnett; barras de erro indicam desvio padrão (DS) de três réplicas técnicas). (B) As imagens em gel mostram análises rt-PCR confirmando a presença de FoMV-MCS1* (315 bp amplicon) na folha 6 de cinco plantas individuais. FoMV-LES22 (625 bp amplicon) foi detectado na folha 6 (amostras FoMV-LES22 1-5, 38 DPI) e folha 4 (amostras FoMV-LES22 6-10, 20 DPI) para dez plantas individuais. O amplicon Zm-Actin serviu como gene de referência. O gráfico da barra representa os valores de expressão relativa qRT-PCR para a expressão les22 em tecidos de milho por agroinjeção de construções virais FoMV-MCS1* ou FoMV-LES22. A supressão de Les22 ocorre em 9 de 10 réplicas biológicas (p=<0,0001; teste pós hoc Dunnett; barras de erro indicam SD para três réplicas técnicas). Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6. Fenótipos de várias construções utilizadas nos experimentos de agroinjeção. Todas as plantas imagens foram injetadas quando tinham de 6 a 7 dias de idade com a cepa Agrobacterium GV3101 carregando as construções indicadas. As imagens foram tiradas em 16 DPI. (A) Sintomas da folha de pCAMBIA1380 (espinha dorsal plasmida vazia), FoMV-EV, FoMV-GFP e SCMV-GFP em luz visível, sob o filtro de clorofila FluorCam a 250 μs de exposição, e sob o filtro FluorCam GFP em exposição de 10 ms. (B) Imagens de microscopia fluorescente das folhas de plantas tratadas com simulação (injetadas apenas apenas com solução MgSO4), FoMV-EV e plantas injetadas FoMV-GFP. Os canais DIC, DsRed e EGFP são mostrados e foram cada um tomados em exposição de 1500 ms. A barra de escala é de 200 μm. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 7. Efeitos morfológicos da injeção. Um exemplo dos efeitos morfológicos mais graves que podem ocorrer a partir da injeção direta no tecido meristeático. Esta lesão pode resultar em "trituração" das folhas e divisão da haste. Clique aqui para ver uma versão maior desta figura.
{kind=link}
| Vírus | Condições de crescimento | Genótipo | # Plantas Infectadas | Total # de Plantas | % Infecção | Avg % de Infecção |
| FoMV-EV | Câmara de Crescimento | Milho | 22 | 23 | 96% | 97% |
| B73 | 18 | 18 | 100% | |||
| B104 | 20 | 21 | 95% | |||
| Estufa | Milho | 20 | 23 | 87% | 89% | |
| B73 | 17 | 18 | 94% | |||
| B104 | 16 | 19 | 84% | |||
| SCMV-EV | Câmara de Crescimento | Milho | 14 | 21 | 67% | 47% |
| B73 | 5 | 18 | 28% | |||
| B104 | 10 | 21 | 48% | |||
| Estufa | Milho | 14 | 23 | 61% | 49% | |
| B73 | 0 | 19 | 0% | |||
| B104 | 19 | 22 | 86% |
Tabela 1: Efeito das condições da câmara de estufa e crescimento na eficiência da inoculação de agroinjeção. As sementes foram germinadas em condições de crescimento idênticas. As mudas germinadas foram agroinjetadas e metade delas foi movida para uma câmara de crescimento (25 °C 16 h luz do dia/ 22C 8h à noite; 185 PAR) e a outra metade foi movida para uma estufa (22-25 °C 16 h luz do dia/22-25 °C 8 h noite; 350-400 PAR). Esta tabela relata a taxa de infecção em percentagem, calculada a partir do número de plantas confirmadas pelo RT-PCR para serem infectadas pelo respectivo vírus dividido pelo número total de plantas agroinjetadas. Não há diferença estatística na eficiência da infecção entre a câmara de crescimento e as condições do efeito estufa (FoMV dois t-test tailed p=0,08; SCMV dois t-test de cauda p=0,96).
| Genótipo de Milho | FoMV-EV | FoMV-LES22 | Total Combinado | ||||
| Infectados | Total | % Infectado | Infectados | Total | % Infectado | % Infectado | |
| Milho | 18 | 23 | 78% | 15 | 23 | 65% | 72% |
| MO47 | 7 | 22 | 32% | 1 | 21 | 5% | 19% |
| K55 | 1 | 15 | 7% | 3 | 17 | 18% | 13% |
| W64A | 10 | 22 | 45% | 8 | 20 | 40% | 43% |
| MO17 | 0 | 16 | 0% | 0 | 13 | 0% | 0% |
| B73 | 10 | 18 | 56% | 7 | 17 | 41% | 49% |
| B101 | 12 | 21 | 57% | 8 | 24 | 33% | 44% |
| FR1064 | 4 | 4 | 100% | 4 | 4 | 100% | 100% |
| B104 | 10 | 22 | 45% | 5 | 21 | 24% | 35% |
| WCC22 | 2 | 7 | 29% | 4 | 6 | 67% | 46% |
| A188 | 0 | 3 | 0% | 4 | 6 | 67% | 44% |
Tabela 2: Eficiência de infecção do FoMV se constrói em todos os genótipos de milho. FoMV-EV e FoMV-LES22 foram agroinjetados em 11 genótipos de milho. Após a injeção, as mudas foram movidas para a estufa. Esta tabela detalha a taxa de infecção como um percentual, calculada a partir do número de plantas infectadas pelo FoMV, conforme confirmado pela RT-PCR dividido pelo número total de plantas agroinjetadas. A taxa total combinada de infecção mostra as taxas médias de infecção de cada genótipo para ambos os construtos fomv testados.
| Fase da Planta | Plantas velhas de 4 a 5 dias | Plantas velhas de 6 a 7 dias | Total Combinado | ||||
| Sintomático | Plantas Totais | % Infectado | Sintomático | Plantas Totais | % Infectado | % Infectado | |
| FoMV-EV | 42 | 72 | 58% | 80 | 170 | 47% | 53% (A) |
| FoMV-PDS | 65 | 157 | 41% | 66 | 184 | 36% | 39% (B C) |
| FoMV-LES22 | 115 | 195 | 59% | 144 | 292 | 49% | 54% (A B) |
| FoMV-GFP | 16 | 103 | 16% | 37 | 217 | 17% | 16% (C) |
| SCMV-GFP | 10 | 95 | 11% | 5 | 82 | 6% | 8% (C) |
Tabela 3: Resumo dos experimentos de injeção. Esta tabela representa um resumo dos experimentos de injeção realizados de agosto de 2017 a agosto de 2018 em mudas de milho doce Golden Bantam. As plantas foram avaliadas para sintomas virais (FoMV-EV), silenciamento de sintomas (pds e les22) ou fluorescência GFP (GFP) por meio da triagem visual (FoMV-EV, FoMV-PDS e FoMV-LES22) ou FluorCam (FoMV-GFP e SCMV-GFP). Os resultados são mostrados individualmente para plantas de 4 a 5 dias de idade e plantas de 6 a 7 dias de idade, bem como um resumo em todas as idades das plantas. Não há diferença significativa encontrada entre plantas de 4 a 5 dias de idade e plantas de 6 a 7 dias de idade (ANOVA unidirecional, F=0,6513). Há uma diferença encontrada entre a construção viral (Onaway ANOVA, F=<0.0001), com as letras representando o relatório de letras de conexão Tukey-Kramer HSD.
Tabela suplementar 1: Tabela listando todos os nomes e sequências de primer usados neste protocolo. Clique aqui para baixar esta Tabela.
Tabela suplementar 2: Teste de acetosyringona. (A) Teste inicial de acetossiingona, comparando as taxas de sintomas de simulação, FoMV-EV e Plantas injetadas foMV-LES22 entre suspensões de inoculação com acetossyringona de 200 μM (+) ou sem acetosyringona (-). (B) Comparando as taxas de infecção do FoMV-LES22 determinadas por RT-PCR entre suspensões de inoculação sem acetossiringona (-), com acetossyringona de 200 μM (+), e adição de 20 μM de acetosyringona à cultura bacteriana 4 horas antes da ressuspensão no buffer, juntamente com a adição de acetossyringona uM à suspensão final (++). No geral, não foi encontrada diferença significativa entre os tratamentos de aceotysyringone (Oneway ANOVA, f=0,5452). Clique aqui para baixar esta Tabela.
Figura suplementar 1: Imagem de fluorescência e validação molecular do SCMV agroinjetado e expressão de proteínas heterólogas no milho. O milho foi agroinjetado com um construto modificado de SCMV contendo tanto CDSs de GFP quanto nano luciferase (NLuc). (A) A imagem fluorcam foi utilizada para triagem e detecção de GFP. A esquerda é uma planta simulada injetada e a direita é a planta injetada SCMV-NLucGFP. (B) Os extratos de proteína de folha foram separados por SDS-PAGE e avaliados para a presença de proteína de casaco NLuc, GFP e SCMV (CP) por ensaio de luciferase em gel ou imunoblot conforme indicado. Clique aqui para baixar este Arquivo.
Discussão
A agrobacterium é uma ferramenta essencial que facilita inúmeras técnicas de biologia molecular em pesquisas relacionadas às plantas. Este estudo fornece um protocolo de agroinjeção para inoculação de vetores virais FoMV e SCMV diretamente em tecidos de milho para aplicações VIGS e VOX. O objetivo principal é aumentar a facilidade e a utilidade das tecnologias baseadas em vírus para pesquisas em plantas de monocot. Embora a agroinoculação direta do milho tenha sido relatada para alguns vírus, os autores não estão cientes de um protocolo detalhado, e não há exemplos de aplicações VIGS e VOX nesses estudos19,22.
Foi relatado, e foi confirmado durante o desenvolvimento deste protocolo, que o local da injeção é um fator-chave para o lançamento com sucesso de uma infecção viral sistêmica através da agroinjeção19. A injeção consistente do local recomendado na planta é considerada a maior variável, pois a posição exata do meristem em mudas de milho é virtualmente indetectável pelo olho. Para minimizar a variação interpessoal, é recomendado dissecar algumas mudas de milho até o meristem para melhor visualizar sua localização (Figura 3C). A posição do meristem em relação ao nódulo coleoxtilar deve ser aproximadamente a mesma para plantas de 4 a 7 dias de idade. Além disso, praticar a injeção com um líquido tingido fornece uma demonstração facilmente visível de como o inóculo preenche a da folha, e como o local da injeção é marcado com corante, a precisão do local de injeção pode ser corroborada (Figura 3G,H). Os tecidos meristemáticos são os mais suscetíveis à agroinjeção, mas a injeção de suspensões de Agrobacterium diretamente neste tecido resulta em efeitos morfológicos indesejáveis (Figura 6)19. Plantas com meristems danificados sobrevivem, mas os defeitos resultantes são indesejáveis e, portanto, a injeção direta desse tecido deve ser evitada.
Existem várias variáveis que podem impactar o lançamento bem-sucedido de uma infecção viral sistêmica por meio da agroinjeção, pois três sistemas biológicos complexos (estirpe vegetal, vírus e Agrobacterium ) devem interagir na coordenação. Essa interação complexa pode ser auxiliada pelas células que se dividem rapidamente da região meristemática, tornando-a um local ideal para a agroinoculação19. A cepa agrobacterium deve ser capaz de infectar células dos tecidos vegetais para entregar o T-DNA carregando o genoma viral, e a planta deve ser suscetível ao vírus para iniciar a replicação viral e infecção sistêmica. Os genótipos de milho diferem em sua suscetibilidade aos vírus (por exemplo, o Mo17 é resistente às cepas de FoMV) ou Agrobacterium , mas a maioria que foi testada parece ser suscetível tanto ao FoMV quanto ao SCMV (Tabela 1 e Tabela 2)53. Por exemplo, a linha de raça FR1064 e a variedade de milho doce Golden Bantam podem ser particularmente suscetíveis tanto aos vetores GV3101 Agrobacterium quanto à base de FoMV.
O número de folhas amostrados e o tempo de amostragem para RT-PCR são fundamentais para uma avaliação precisa da infecção viral. Nos exemplos aqui apresentados, o número da folha foi determinado começando na primeira folha arredondada (comumente conhecida como "folha do polegar") e contando para cima. As folhas foram amostradas uma vez que foram expandidas e a folha seguinte começou a emergir. No entanto, as folhas são ótimas para a amostragem podem variar de acordo com as espécies de vírus utilizadas, condições de crescimento e genótipo de milho. Portanto, recomenda-se um experimento de curso de tempo inicial ao aplicar este protocolo a um novo sistema de vírus para otimizar a estratégia de amostragem em relação às folhas e ao tempo.
O construto específico utilizado afeta significativamente a eficiência deste protocolo. Por exemplo, os vetores vazios, FoMV-EV e SCMV-EV, e FoMV-PDS e FoMV-LES22, que contêm pequenas pastilhas (313 bp e 329 bp, respectivamente), normalmente produzem os maiores percentuais de plantas com sintomas virais nesses experimentos (Tabelas 1 e Tabela 2). No entanto, os vírus recombinantes que carregam inserções maiores do GFP ORF (720 bp) em FoMV-GFP e SCMV-GFP, apresentaram taxas de infecção muito menores quando comparadas às plantas injetadas com os construtos de vetor vazio ou silenciamento genético. Essa tendência pode ser devido aos impactos negativos sobre a aptidão viral causados pelo aumento de quantidades de material genético exógeno no genoma viral. Vários estudos têm demonstrado que a estabilidade da inserção de vetores virais vegetais depende em grande parte do tamanho da inserção e sequenciar 36,54,55,56,57. Além disso, houve uma diferença notável no percentual de plantas que se infectam após a inoculação com o vetor vazio FoMV ou SCMV, sugerindo que é necessário um trabalho adicional para otimizar este protocolo para SCMV (Tabela 1). Esses resultados indicam que alguns problemas podem ser necessários ao desenvolver uma construção, pois a sequência e o comprimento do fragmento podem afetar a eficiência.
No geral, este estudo mostrou que a agroinjeção de mudas de milho é um método de inoculação eficaz para dois vírus diferentes da planta de RNA, configurações de múltiplos vetores e 11 genótipos de milho. Este trabalho com FoMV e SCMV, emparelhado com trabalhos anteriores utilizando injeção com vírus de mottle clorotic de milho (MCMV) ou MSV, indica que a agroinjeção é adequada para inoculação de mudas de milho com clones infecciosos de vírus de RNA e DNA19,20,21,22. Além disso, este trabalho mostra ainda que a agroinjeção é um método viável para vetores VIGS e VOX e pode ser aplicada a plantas com até quatro dias de idade (Tabela 3). Espera-se que o protocolo aqui apresentado seja prontamente adaptado por biólogos de milho para facilitar a pesquisa em estudos de genômica funcional envolvendo silenciamento genético transitório (VIGS) e superexpressão (VOX). A agroinjeção também tem a capacidade de facilitar abordagens de edição de genes baseadas em vírus (VEdGE) que de outra forma seriam limitadas pela dependência da transformação da planta, potencialmente melhorando a eficiência de edição, bem como a acessibilidade58,59,60. Dada a variedade agrobacterium adequada, genótipos de milho e vetores virais são cuidadosamente combinados, espera-se que a inoculação por agroinjeção se torne uma ferramenta valiosa para análises transitórias da função genética no milho.
Divulgações
Os pesquisadores não têm conflitos de interesse para divulgar.
Agradecimentos
A Universidade Estadual de Iowa faz parte de uma equipe que apoia o programa Desetos Aliados de Insetos da DARPA HR0011-17-2-0053. Este trabalho também foi apoiado pelo Iowa State University Plant Sciences Institute, Iowa State University Crop Bioengineering Center, USDA NIFA Hatch project number 3808, e State of Iowa Funds. K.L.H. também foi parcialmente apoiado pelo programa de pós-graduação de phenomics da Universidade Estadual de Iowa financiado pela National Science Foundation (DGE #1545453) e pela Agricultural and Food Research Initiative no 2019-07318 do Instituto Nacional de Alimentos e Agricultura do USDA. Os financiadores não tiveram papel na concepção do estudo e coleta, análise e interpretação dos dados e na redação do manuscrito. Quaisquer opiniões, achados e conclusões ou recomendações expressas neste material são dos autores e não refletem necessariamente as opiniões dos financiadores.
Agradecemos a Nick Lauter (USDA-ARS, Ames, IA) pela semente de linhas de milho, Christian F. Montes-Serey (Iowa State University) por fazer o clone FoMV-GFP, e Tyler Austin (Iowa State University) para assistência técnica.
Materiais
| Name | Company | Catalog Number | Comments |
| 1 mL syringes | Fisher Scientific | 14955450 | alternatively, BD 309659 |
| 15 mL Falcon Tubes | Corning Science | 352059 | |
| 1kb+ Ladder | ThermoFisher Scientific | 10787018 | For assessing sizes of PCR products |
| 25G x 5/8" PrecisionGlide Needles | Becton, Dickinson and Company (BD) | 305122 | |
| 28°C Incubator | For Agrobacterium | ||
| 37°C Incubator | For E. coli | ||
| Acetosyringone | MilliporeSigma | D134406 | Optional |
| Agar | MilliporeSigma | A4800 | |
| Agarose | GeneMate | E-3120 | For making gels to check for virus/insert stability |
| Agrobacterium tumefaciens Strain GV3101 | Carries vir plasmid encoding T-DNA transfer machinery, RifR, GmR, from lab stock | ||
| Bsu36I | New England Biolabs | R0524 | |
| cDNA Kit | ThermoFisher Scientific | K1672 | Maxima First Strand cDNA Synthesis Kit with Dnase |
| Chloroform | Fisher Scientific | C298 | For RNA extraction |
| Cuvettes | Fisher Scientific | 14955127 | 1.5 mL |
| D-(+)-Glucose | MilliporeSigma | G7528 | Alkaline Lysis |
| DH5alpha Competent E. coli Cells | New England Biolabs | C2987 | |
| DNA Ligase | ThermoFisher Scientific | K1422 | Rapid DNA Ligation Kit |
| EDTA (Ethylenediamine Tetraacetic Acid, Disodium Salt Dihydrate) | Fisher Scientific | S311 | Alkaline Lysis |
| Ethanol | For RNA extraction | ||
| Fertilizer | Peters Fertilizer 15-15-15 Concentrate | ||
| Flat Inserts | T.O. Plastics | 715357C | For germinating seeds in trays |
| Flats | T.O. Plastics | 710245C | For germinating seeds in trays |
| FluorCam | Photon Systems Instruments | To assess maize plants for GFP expression before microscope | |
| Fluorescence Microscope | |||
| Gel Electrophoresis Box | |||
| Gentamycin Sulfate | Fisher Scientific | BP918 | |
| Glacial Acetate | Fisher Scientific | A38 | Alkaline Lysis |
| Glycerol | Fisher Scientific | G33-500 | For saving frozen stocks of bacteria |
| Go-Taq, 2X | Promega | M7123 | |
| Hydrochloric Acid | Fisher Scientific | A144 | for pHing solutions |
| Isopropanol | Sigma-Aldrich | 109827 | For RNA extraction |
| Kanamycin, Monosulfate | Fisher Scientific | BP906 | |
| Large Pots | Kordlok | SQL0550 | 5x5x4" or bigger. For transplanting seedlings. |
| Luria Bertani (LB) Broth, Miller | Himedia | M1245 | |
| Magnesium Sulfate Heptahydrate | Amresco | 662 | |
| Maize Golden Bantam Sweet Corn Seed | American Meadows, West Coast Seeds | ||
| Maize Inbred Seed | Our seed comes from our institution, but we are not able to provide this for other researchers. | ||
| Maxima H Minus Reverse Transcriptase | ThermoFisher Scientific | EP0753 | |
| MilliQ | Elga | Purelab Ultra | |
| Monarch PCR & DNA Cleanup Kit | New England Biolabs | T1030 | |
| PacI | New England Biolabs | R0547 | |
| Peters Excel 15-5-15 Fertilizer | ICL Specialty Fertilizers | G99140 | |
| Petri Dish, 95 mm x 15 mm | Fisher Scientific | FB0875714G | |
| pH Meter | |||
| Potassium Acetate | Fisher Scientific | P171 | Alkaline Lysis |
| Primers | Our primers were synthesized through our institutional DNA facility or through IDT | ||
| PspOMI | New England Biolabs | R0653 | |
| Q5 High-Fidelity DNA Polymerase | New England Biolabs | M0491 | |
| Rifampicin | EMD Millipore Corp | 557303 | |
| Rnase A | ThermoFisher Scientific | 12091021 | Alkaline Lysis |
| SbfI | New England Biolabs | R0642 | |
| Scale | For weighing chemicals for media or buffers | ||
| SDS (Sodium Dodecyl Sulfate) | Fisher Scientific | BP166 | Alkaline Lysis |
| Sodium Hydroxide | Fisher Scientific | S318 | Alkaline Lysis |
| Soil Substrate | SunGro Horticulture | SS#1-F1P | Sunshine Mix #1/Fafard-1P, any soil mix that maize grows well in is sufficient |
| Spectrophotometer | For measuring OD600 | ||
| Sybr Safe, 10,000X | Invitrogen | S33102 | For making gels to check for virus/insert stability |
| Thermocycler | For PCR | ||
| Tris Base | Fisher Scientific | BP154 | Alkaline Lysis |
| Trizol | Ambion | 15596018 | For RNA extraction |
| Weigh Paper | For weighing chemicals for media or buffers | ||
| XbaI | New England Biolabs | R0145 |
Referências
- Zaidi, S. E. A., Mansoor, S. Viral vectors for plant genome engineering. Frontiers in Plant Science. 8, 539 (2017).
- Kant, R., Dasgupta, I. Gene silencing approaches through virus-based vectors: speeding up functional genomics in monocots. Plant Molecular Biology. 100, 3-18 (2019).
- Hu, J., et al. A barley stripe mosaic virus-based guide RNA delivery system for targeted mutagenesis in wheat and maize. Molecular Plant Pathology. 20 (10), 1463-1474 (2019).
- Pasin, F., Menzel, W., Daròs, J. A. Harnessed viruses in the age of metagenomics and synthetic biology: an update on infectious clone assembly and biotechnologies of plant viruses. Plant Biotechnology Journal. 17 (6), 1010-1026 (2019).
- Cody, W. B., Scholthof, H. B. Plant virus vectors 3.0: Transitioning into synthetic genomics. Annual Review of Phytopathology. 57 (1), 211-230 (2019).
- Mei, Y., et al. Protein expression and gene editing in monocots using foxtail mosaic virus vectors. Plant Direct. 3 (11), 00181 (2019).
- Ruiz, M. T., Voinnet, O., Baulcombe, D. C. Initiation and maintenance of virus-induced gene silencing. Plant Cell. 10 (6), 937-946 (1998).
- Bekele, D., Tesfaye, K., Fikre, A. Applications of virus induced gene silencing (VIGS) in plant functional genomics studies. Journal of Plant Biochemistry & Physiology. 07 (01), 1000229 (2019).
- Scholthof, H. B., Scholthof, K. B. G., Jackson, A. O. Plant virus gene vectors for transient expression of foreign proteins in plants. Annual Review of Phytopathology. 34 (1), 299-323 (1996).
- Holzberg, S., Brosio, P., Gross, C., Pogue, G. P. Barley stripe mosaic virus-induced gene silencing in a monocot plant. Plant Journal. 30 (3), 315-327 (2002).
- Wang, R., et al. An efficient virus-induced gene silencing vector for maize functional genomics research. Plant Journal. 86 (1), 102-115 (2016).
- Redinbaugh, M. G., et al. Transmission of viral RNA and DNA to maize kernels by vascular puncture inoculation. Journal of Virological Methods. 98 (2), 135-143 (2001).
- Scholthof, H. B. The capsid protein gene of tomato bushy stunt virus is dispensable for systemic movement and can be replaced for localized expression of foreign genes. Molecular Plant-Microbe Interactions. 6 (3), 309 (1993).
- Scholthof, H. B., Scholthof, K. B. G., Kikkert, M., Jackson, A. O. Tomato bushy stunt virus spread is regulated by two nested genes that function in cell-to-cell movement and host-dependent systemic invasion. Virology. 213 (2), 425-438 (1995).
- Scholthof, H. B. Rapid delivery of foreign genes into plants by direct rub-inoculation with intact plasmid dna of a tomato bushy stunt virus gene vector. Journal of Virology. 73 (9), 7823-7829 (1999).
- Zhang, J., et al. Vacuum and co-cultivation agroinfiltration of (germinated) seeds results in tobacco rattle virus (TRV) mediated whole-plant virus-induced gene silencing (VIGS) in wheat and maize. Frontiers in Plant Science. 8, 393 (2017).
- Vaghchhipawala, Z., Rojas, C. M., Senthil-Kumar, M., Mysore, K. S. Agroinoculation and agroinfiltration: simple tools for complex gene function analyses. Methods in Molecular Biology. 678, 65-76 (2011).
- Grimsley, N., Hohn, B., Hohn, T., Walden, R. "Agroinfection," an alternative route for viral infection of plants by using the Ti plasmid. Proceedings of the National Academy of Sciences. 83 (10), 3282-3286 (1986).
- Grimsley, N. H., Ramos, C., Hein, T., Hohn, B. Merisfematic tissues of maize plants are most suscepnsle to agroinfection with maize streak virus. Bio/Technology. 6 (2), 185-189 (1988).
- Martin, D. P., Rybicki, E. P. Improved efficiency of Zea mays agroinoculation with Maize streak virus. Plant Disease. 84 (10), 1096 (2000).
- Martin, D. P., Willment, J. A., Rybicki, E. P. Evaluation of maize streak virus pathogenicity in differentially resistant Zea mays genotypes. Phytopathology. 89 (8), 695-700 (1999).
- Wang, Q., et al. Further characterization of Maize chlorotic mottle virus and its synergistic interaction with Sugarcane mosaic virus in maize. Scientific Reports. 7, 39960 (2017).
- Hsieh, M. H., et al. Optimizing virus-induced gene silencing efficiency with Cymbidium mosaic virus in Phalaenopsis flower. Plant Science. 201-202 (1), 25-41 (2013).
- Hsieh, M. H., et al. Virus-induced gene silencing unravels multiple transcription factors involved in floral growth and development in Phalaenopsis orchids. Journal of Experimental Botany. 64 (12), 3869-3884 (2013).
- Zenna, N. S., et al. Genetic analysis of tolerance to rice tungro bacilliform virus in rice (Oryza sativa L.) through agroinoculation. Journal of Phytopathology. 154 (4), 197-203 (2006).
- Marks, M. S., Kemp, J. M., Woolston, C. J., Dale, P. J. Agroinfection of wheat: A comparison of Agrobacterium strains. Plant Science. 63 (2), 247-256 (1989).
- Dasgupta, I., et al. Rice tungro bacilliform virus DNA independently infects rice after Agrobacterium-mediated transfer. Journal of General Virology. 72 (6), 1215-1221 (1991).
- Boulton, M. I., Buchholz, W. G., Marks, M. S., Markham, P. G., Davies, J. W. Specificity of Agrobacterium-mediated delivery of maize streak virus DNA to members of the Gramineae. Plant Molecular Biology. 12 (1), 31-40 (1989).
- Paulsen, A. Q. Purification and properties of foxtail mosaic virus. Phytopathology. 77 (11), 1346 (1977).
- Bancroft, J. B., Rouleau, M., Johnston, R., Prins, L., Mackie, G. A. The entire nucleotide sequence of foxtail mosaic virus RNA. Journal of General Virology. 72 (9), 2173-2181 (1991).
- Bruun-Rasmussen, M., Madsen, C. T., Johansen, E., Albrechtsen, M. Revised sequence of foxtail mosaic virus reveals a triple gene block structure similar to potato virus X. Archives of Virology. 153 (1), 223-226 (2008).
- Rouleau, M., Bancroft, J. B., Mackie, G. A. Partial purification and characterization of foxtail mosaic potexvirus RNA-dependent RNA polymerase. Virology. 197 (2), 695-703 (1993).
- Rouleau, M., Smith, R. J., Bancroft, J. B., Mackie, G. A. Purification, properties, and subcellular localization of foxtail mosaic potexvirus 26-kDa protein. Virology. 204 (1), 254-265 (1994).
- Samuels, T. D., et al. Subcellular targeting and interactions among the potato virus X TGB proteins. Virology. 367 (2), 375-389 (2007).
- Cho, S. Y., Kim, K. H. Identification of the capsid protein-binding region of the SL1(+) RNA located at the 5' region of the potato virus X genome. Plant Pathology Journal. 28 (1), 75-80 (2012).
- Mei, Y., Zhang, C., Kernodle, B. M., Hill, J. H., Whitham, S. A. A foxtail mosaic virus vector for virus-induced gene silencing in maize. Plant Physiology. 171 (2), 760-772 (2016).
- Bouton, C., et al. Foxtail mosaic virus: A viral vector for protein expression in cereals. Plant Physiology. 177 (4), 1352-1367 (2018).
- Mei, Y., Liu, G., Zhang, C., Hill, J. H., Whitham, S. A. A sugarcane mosaic virus vector for gene expression in maize. Plant Direct. 3 (8), 00158 (2019).
- Gal-On, A., Meiri, E., Huet, H., Hua, W. J., Raccah, B., Gaba, V. Particle bombardment drastically increases the infectivity of cloned DNA of zucchini yellow mosaic potyvirus. Journal of General Virology. 76 (12), (1995).
- Gao, R., et al. Construction of an infectious cDNA clone and gene expression vector of Tobacco vein banding mosaic virus (genus Potyvirus). Virus Research. 169 (1), 276-281 (2012).
- López-Moya, J. J., García, J. A. Construction of a stable and highly infectious intron-containing cDNA clone of plum pox potyvirus and its use to infect plants by particle bombardment. Virus Research. 68 (2), (2000).
- Choi, I. R., French, R., Hein, G. L., Stenger, D. C. Fully biologically active in vitro transcripts of the eriophyid mite-transmitted wheat streak mosaic tritimovirus. Phytopathology. 89 (12), (1999).
- Kim, K. S., et al. Infectivity of in vitro transcripts of Johnsongrass mosaic potyvirus full-length cDNA clones in maize and sorghum. Archives of Virology. 148 (3), 563-574 (2003).
- Stewart, L. R., Bouchard, R., Redinbaugh, M. G., Meulia, T. Complete sequence and development of a full-length infectious clone of an Ohio isolate of Maize dwarf mosaic virus (MDMV). Virus Research. 165 (2), 219-224 (2012).
- Wylie, S. J., et al. ICTV virus taxonomy profile: Potyviridae. Journal of General Virology. 98 (3), 352-354 (2017).
- Shukla, D. D. taxonomy of potyviruses infecting maize, sorghum, and sugarcane in Australia and the United States as determined by reactivities of polyclonal antibodies directed towards virus-specific N-termini of coat proteins. Phytopathology. 79 (2), 223 (1989).
- Shukla, D. D., Ward, C. W. Amino Acid sequence homology of coat proteins as a basis for identification and classification of the potyvirus group. Journal of General Virology. 69 (11), 2703-2710 (1988).
- Chung, B. Y. W., Miller, W. A., Atkins, J. F., Firth, A. E. An overlapping essential gene in the Potyviridae. Proceedings of the National Academy of Sciences of the United States of America. 105 (15), 5897-5902 (2008).
- Jarchow, E., Grimsley, N. H., Hohn, B. virF, the host-range-determining virulence gene of Agrobacterium tumefaciens, affects T-DNA transfer to Zea mays. Proceedings of the National Academy of Sciences of the United States of America. 88 (23), 10426-10430 (1991).
- Hu, G., Yalpani, N., Briggs, S. P., Johal, G. S. A porphyrin pathway impairment is responsible for the phenotype of a dominant disease lesion mimic mutant of maize. The Plant Cell. 10 (7), 1095 (2007).
- Qin, G., et al. Disruption of phytoene desaturase gene results in albino and dwarf phenotypes in Arabidopsis by impairing chlorophyll, carotenoid, and gibberellin biosynthesis. Cell Research. 17 (5), 471-482 (2007).
- Jones, P. Isolation of plasmid DNA from E. coli. Encyclopedia of Life Sciences. , (2003).
- Ji, Q., Yang, B., Lee, M., Chen, Y., Lübberstedt, T. Mapping of quantitative trait loci/locus conferring resistance to foxtail mosaic virus in maize using the intermated B73-×-Mo17 population. Plant Breeding. 129 (6), 721-723 (2010).
- Pacak, A., et al. The brome mosaic virus-based recombination vector triggers a limited gene silencing response depending on the orientation of the inserted sequence. Archives of Virology. 155 (2), 169-179 (2010).
- Miché, L., Battistoni, F., Gemmer, S., Belghazi, M., Reinhold-Hurek, B. Host-dependent expression of Rhizobium leguminosarum bv. viciae hydrogenase is controlled at transcriptional and post-transcriptional levels in legume nodules. Molecular Plant-Microbe Interactions. 19 (5), 1323-1331 (2018).
- Yamagishi, M., Masuta, C., Suzuki, M., Netsu, O. Peanut stunt virus-induced gene silencing in white lupin (lupinus albus). Plant Biotechnology. 32 (3), 181-191 (2015).
- Avesani, L., et al. Stability of Potato virus X expression vectors is related to insert size: Implications for replication models and risk assessment. Transgenic Research. 16 (5), 587-597 (2007).
- Ali, Z., et al. Efficient virus-mediated genome editing in plants using the CRISPR/Cas9 system. Molecular Plant. 8 (8), 1288-1291 (2015).
- Cody, W. B., Scholthof, H. B., Mirkov, T. E. Multiplexed gene editing and protein overexpression using a tobacco mosaic virus viral vector. Plant Physiology. 175 (1), 23-35 (2017).
- Ali, Z., Eid, A., Ali, S., Mahfouz, M. M. Pea early-browning virus-mediated genome editing via the CRISPR/Cas9 system in Nicotiana benthamiana and Arabidopsis. Virus Research. 244, 333-337 (2018).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados