
Method Article
Agroinoculazione diretta di piantine di mais per iniezione con cloni infettivi del virus del mosaico della coda di volpe ricombinante e virus del mosaico della canna da zucchero
* Questi autori hanno contribuito in egual misura
In questo articolo
Riepilogo
Viene presentato un protocollo di iniezione (agroiniezione) a base di Agrobacterium per l'inoculazione del virus del mosaico a coda di volpe e dei virus del mosaico della canna da zucchero in piantine di mais. L'inoculazione in questo modo porta a infezioni virali, silenziamento genico indotto da virus di geni marcatori e sovraespressione virale di GFP.
Abstract
Gli approcci di inoculazione basati su Agrobacterium sono ampiamente utilizzati per introdurre vettori virali nei tessuti vegetali. Questo studio descrive un protocollo per l'iniezione di piantine di mais vicino al tessuto meristematico con Agrobacterium che trasporta un vettore virale. Per ottimizzare questo metodo sono stati utilizzati cloni del virus del mosaico a coda di volpe ricombinante (FoMV) progettati per il silenziamento genico e l'espressione genica è stato utilizzato per includere un virus del mosaico della canna da zucchero ricombinante (SCMV) progettato per l'espressione genica. Frammenti di geni o sequenze codificanti di interesse sono inseriti in un genoma virale infettivo modificato che è stato clonato nel vettore binario del plasmide T-DNA pCAMBIA1380. I costrutti plasmidici risultanti vengono trasformati in Agrobacterium tumefaciens ceppo GV3101. Piantine di mais di 4 giorni possono essere iniettate vicino al nodo coleoptilare con batteri risospesi in soluzione di MgSO4. Durante l'infezione da Agrobacterium, il T-DNA che trasporta il genoma virale viene trasferito alle cellule di mais, consentendo la trascrizione del genoma dell'RNA virale. Mentre il virus ricombinante si replica e si diffonde sistemicamente in tutta la pianta, si possono osservare sintomi virali e cambiamenti fenotipici derivanti dal silenziamento dei geni bersaglio lesione mimica 22 (les22) o fitoene desaturasi (pds) sulle foglie, o l'espressione della proteina fluorescente verde (GFP) può essere rilevata all'illuminazione con luce UV o microscopia a fluorescenza. Per rilevare il virus e valutare contemporaneamente l'integrità dell'inserto, l'RNA viene estratto dalle foglie della pianta iniettata e la RT-PCR viene condotta utilizzando primer che fiancheggiano il sito di clonazione multipla (MCS) che trasporta la sequenza inserita. Questo protocollo è stato utilizzato efficacemente in diversi genotipi di mais e può essere facilmente esteso ad altri vettori virali, offrendo così uno strumento accessibile per l'introduzione di vettori virali nel mais.
Introduzione
Cloni infettivi di molti virus vegetali sono stati progettati per il silenziamento genico indotto da virus (VIGS), la sovraespressione genica (VOX) e, più recentemente, l'editing genetico abilitato dal virus (VEdGE)1,2,3,4,5,6,7,8,9,10,11 . Man mano che vengono sviluppati nuovi costrutti virali, devono essere considerati anche metodi per infettare con successo i tessuti vegetali con questi virus modificati. I metodi attuali per lanciare infezioni virali nelle piante includono il bombardamento di particelle, l'inoculazione di trascritti di RNA in vitro o cloni di DNA, l'inoculazione della puntura vascolare o l'inoculazione di Agrobacterium tumefaciens (agroinoculazione)5,12,13,14,15,16,17 . Ognuno di questi metodi di inoculazione presenta vantaggi e svantaggi intrinseci, che includono costi, necessità di attrezzature specializzate e fattibilità all'interno di un determinato sistema di virus vegetale. I metodi che utilizzano l'infiltrazione o l'iniezione di ceppi di Agrobacterium contenenti costrutti binari di T-DNA progettati per fornire virus ricombinanti sono preferiti, perché sono semplici e poco costosi. Tuttavia, mancano metodi di agroinoculazione dettagliati per specie monocotiledoni come Zea mays (mais).
Uno dei primi rapporti sull'agroinoculazione per la consegna del virus è stato pubblicato nel 1986, quando il genoma del virus del mosaico del cavolfiore (CaMV) è stato inserito in un costrutto T-DNA e l'Agrobacterium risultante che trasportava questo costrutto è stato inoculato sulle piante di rapa18. Da allora sono stati sviluppati ulteriori metodi per l'agroinoculazione. Ad esempio, nel caso del virus del mosaico a coda di volpe (FoMV), Nicotiana benthamiana può essere utilizzato come ospite intermedio per generare particelle virali nelle foglie che forniscono una fonte di inoculo6. L'inoculazione di mais utilizzando foglie di N. benthamiana infette è efficiente, rapida e semplice, ma l'uso di un ospite intermedio non funziona per tutti i virus che infettano il mais. Il virus del mosaico della canna da zucchero (SCMV), ad esempio, non può infettare N. benthamiana, richiedendo l'uso di fonti alternative di inoculo per i vettori derivati da questo virus. Nel 1988, Agrobacterium contenente il virus della striscia di mais (MSV), un virus del DNA, è stato introdotto nelle piantine di mais per iniezione (agroiniezione), dimostrando che i metodi di inoculazione a base di Agrobacterium sono utili anche per le monocotiledoni19. Nonostante questo successo iniziale con l'agroiniezione, sono stati pubblicati pochi studi che utilizzano questa tecnica nel mais, lasciando aperte domande sull'applicabilità di questo metodo per i virus a RNA e i vettori VIGS, VOX e VEdGE20,21,22. Tuttavia, l'ampio uso dell'agroiniezione nelle specie monocot è promettente, perché questo approccio generale è stato utilizzato in orchidea, riso e grano23,24,25,26,27,28.
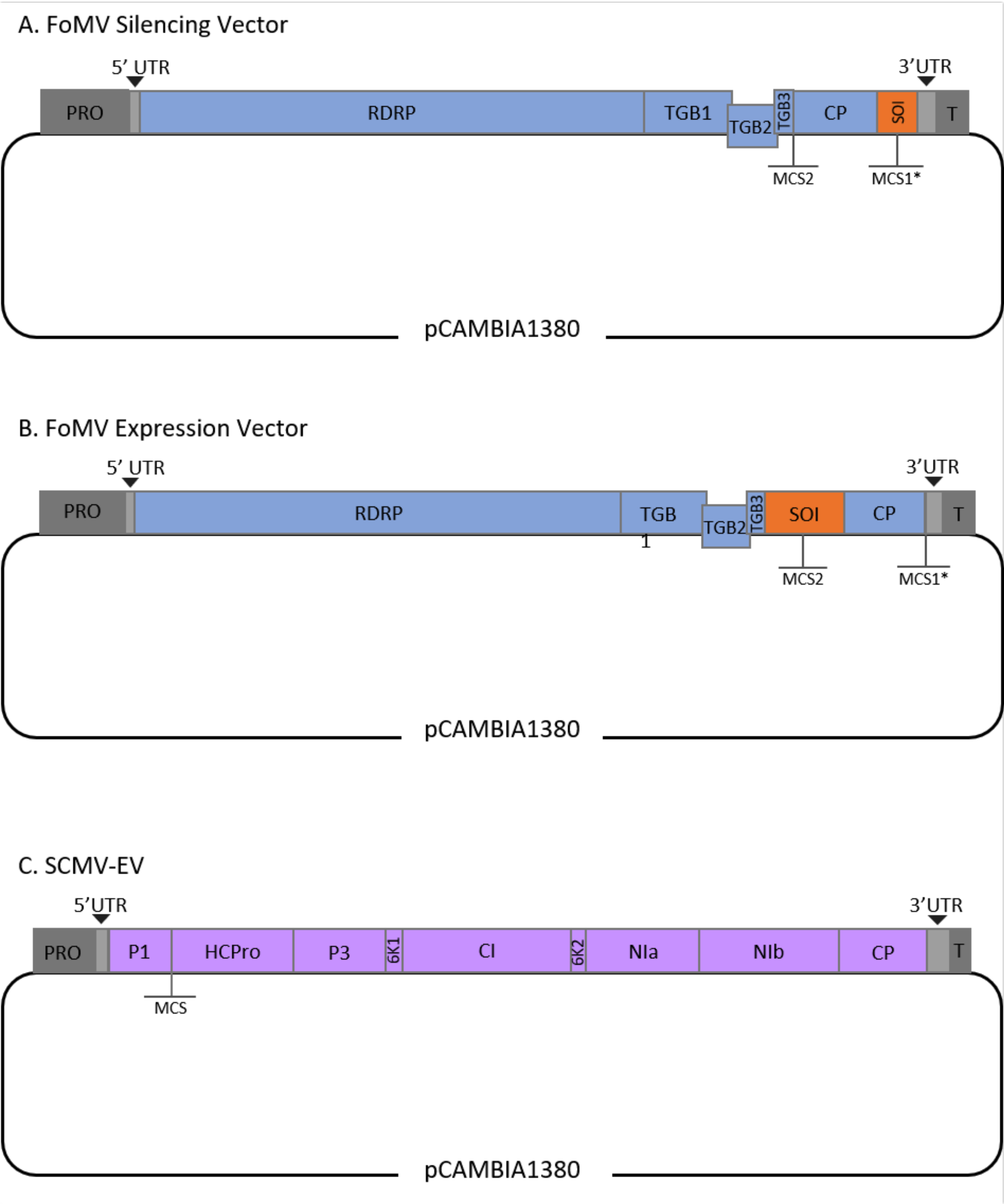
Questo protocollo è stato ottimizzato per FoMV e Agrobacterium ceppo GV3101 ed è stato applicato anche a un vettore SCMV. FoMV è un potexvirus con una vasta gamma di ospiti che comprende 56 specie monocot e dicot29. FoMV ha un senso positivo di 6,2 kilobase (kb), genoma di RNA a singolo filamento che codifica cinque diverse proteine da cinque frame di lettura aperti (ORF)30,31,32,33,34,35. FoMV è stato precedentemente sviluppato in un vettore VIGS e VOX per il mais incorporando un clone infettivo su una spina dorsale plasmidica T-DNA6,36,37. Il genoma virale è stato modificato per applicazioni VIGS aggiungendo un sito di clonazione (MCS1*) immediatamente a valle della proteina del mantello (CP) (Figura 1A)36. Per le applicazioni VOX e VEdGE, il promotore CP è stato duplicato ed è stato aggiunto un secondo sito di clonazione (MCS2) per consentire l'inserimento di sequenze di interesse tra ORF 4 e CP (Figura 1B)6. Il vettore FoMV contenente sia MCS1 che MCS2 senza inserti è il vettore vuoto FoMV (FoMV-EV) (Figura 1).
SCMV è un virus non correlato che è stato sviluppato per VOX nel mais38. È un membro della famiglia Potyviridae, di cui diversi membri sono stati progettati per esprimere proteine estranee in planta39,40,41,42,43,44. La gamma ospite di SCMV comprende mais, sorgo e canna da zucchero45,46, rendendolo prezioso per gli studi funzionali genici in queste principali piante coltivate36,38. SCMV ha un senso positivo, genoma di RNA a singolo filamento di circa 10 kb di lunghezza47,48. Per creare il vettore SCMV VOX, la ben consolidata giunzione P1/HCPro è stata utilizzata come sito di inserimento per sequenze eterologhe38. Questo sito di clonazione è seguito dalla sequenza che codifica un sito di scissione della proteasi NIa-Pro, che porta alla produzione di proteine indipendenti dalla poliproteina SCMV (Figura 1C).
I plasmidi T-DNA portatori di cDNA infettivo di questi virus ricombinanti sono stati trasformati in Agrobacterium ceppo GV3101. GV3101 è un ceppo di tipo nopalina, che è ben noto per essere in grado di trasferire T-DNA a specie monocotiledoni, tra cui il mais26,28,49. Inoltre, precedenti studi di agroiniezione hanno utilizzato i ceppi C58 o il suo derivato GV3101, così come 19,20,22,27.
Tre geni marcatori sono stati utilizzati nello sviluppo di questo protocollo: due per il silenziamento genico e uno per l'espressione genica. Un frammento di 329 coppie di basi (bp) dalla lesione del gene del mais mimic 22 (les22, GRMZM2G044074) è stato utilizzato per costruire il vettore silenziatore FoMV-LES22. Quando les22 viene silenziato nel mais, piccole macchie rotonde di cellule necrotiche appaiono lungo la vascolarizzazione delle foglie che si espandono e si fondono in vaste aree di tessuto fogliare necrotico50. FoMV-PDS, contenente un frammento di 313 bp dal gene del sorgo fitoene desaturasi (pds, LOC110436156, 96% di identità di sequenza al mais pds, GRMZM2G410515), induce il silenziamento di pds nel mais, con conseguente piccole striature di cellule fotosbiancate lungo la vascolarizzazione delle foglie che si allungano nel tempo51. La sequenza codificante intatta per la proteina fluorescente verde (GFP) è stata utilizzata per dimostrare l'espressione proteica sia per FoMV (FoMV-GFP) che per SCMV (SCMV-GFP). L'espressione della GFP nelle foglie è in genere più rilevabile a 14 giorni dopo l'inoculazione (DPI)6. Sebbene ci siano stati studi precedenti che utilizzano l'agroiniezione di vettori virali nel mais, questi esperimenti hanno solo dimostrato che l'agroiniezione può facilitare l'infezione virale da un clone infettivo nelle piantine di mais e non espandersi a virus ricombinanti progettati per applicazioni VIGS o VOX19,20,21,22. Il protocollo qui presentato si basa su precedenti metodi di agroiniezione, in particolare Grismley et al.19. Nel complesso, questo metodo di agroiniezione è compatibile con i vettori VIGS e VOX, non richiede attrezzature specializzate o ospiti alternativi come fonti di inoculo e riduce il tempo e il costo complessivi necessari per impostare ed eseguire inoculazioni rispetto ad altri metodi comuni che richiedono biolistica o trascrizione in vitro. Questo protocollo faciliterà gli studi di genomica funzionale nel mais con applicazioni che coinvolgono VIGS, VOX e VEdGE.
Protocollo
1. Costruzione del plasmide
NOTA: Questo protocollo può essere applicato ad altri vettori virali o ceppi di Agrobacterium , ma ciò può influire sul successo complessivo dell'inoculazione mediante agroiniezione. Eseguire sempre fasi di inoculazione e placcatura batterica in una cappa a flusso laminare.
- Costrutto di silenziamento FoMV
NOTA: Luria-Bertani (LB) media (Miller) viene utilizzato per tutti i media se non diversamente specificato. Liquid LB viene prodotto sospendendo 25 g di granuli in 1.000 ml di acqua distillata e autoclave per 15 minuti a 121 °C. I supporti LB solidi sono realizzati in modo simile con l'aggiunta dell'1,5% di agar prima dell'autoclave. Gli antibiotici vengono aggiunti dopo che LB è stato raffreddato a ~ 60 ° C e la soluzione viene versata in piastre di Petri da 95 x 15 mm. Le concentrazioni di antibiotici da utilizzare sono le seguenti: rifampicina (rif) a 25 μg/mL, gentamicina (gent) a 50 μg/mL e kanamicina (kan) a 50 μg/mL.- La PCR amplifica i frammenti del gene del mais da silenziare (ad esempio, les22 o pds) utilizzando un primer in avanti con un sito di restrizione PacI e un primer inverso con un sito di restrizione XbaI. Ciò consentirà la legatura dei frammenti genici nel MCS1* del vettore binario FoMV-pCAMBIA1380 nell'orientamento antisenso.
NOTA: Impostare la PCR utilizzando una DNA polimerasi ad alta fedeltà, primer avanti e indietro a 10 μM ciascuno, modello di DNA plasmidico e acqua, seguendo le specifiche della DNA polimerasi. Amplificare per 35 cicli, utilizzando una temperatura di ricottura secondo la temperatura di fusione della DNA polimerasi e del primer (Tm), e un'estensione di 30 s per kilobase da amplificare. - Eseguire la purificazione PCR utilizzando un kit di purificazione PCR in base alle specifiche del kit.
- Digerire il prodotto PCR purificato e il FoMV-EV con gli enzimi di restrizione XbaI e PacI. Utilizzare 1 μg di plasmide o tutto il prodotto PCR purificato, 2 μL di tampone 10x, 1 μL di enzima di restrizione e aggiungere acqua per ottenere un volume finale di 20 μL di reazione. Incubare secondo le specifiche enzimatiche.
- Ligate il prodotto PCR digerito e FoMV-EV insieme a T4 DNA ligasi secondo il protocollo del produttore.
- Trasformare il plasmide legato in cellule di E. coli chimicamente dh5α chimicamente competenti utilizzando il metodo dello shock termico.
- Scongelare le cellule sul ghiaccio e aggiungere 3 μL di plasmide al tubo. Incubare sul ghiaccio per 30 minuti, quindi shock termico per 30 s a 42 °C.
- Mettere sul ghiaccio per 5 min, aggiungere 200 μL di brodo super ottimale con repressione catabolica (SOC) e permettere alle cellule di E. coli di recuperare in mezzi SOC per 1 ora a 37 °C con agitazione a 225 rpm.
- Piastra su terreni LB selettivi di kanamicina e incubazione a 37 °C durante la notte.
- Controllare le colonie per cloni accurati mediante sequenziamento Sanger utilizzando i primer FM-5840F e FM-6138R (Tabella supplementare 1). Presentare 250 ng di DNA plasmidico a una struttura che eseguirà il sequenziamento Sanger. Per questo esperimento i campioni sono stati inviati alla Iowa State University DNA Core Facility.
- Inoculare 2 mL di LB liquido con la colonia scelta e incubare a 37 °C durante la notte con agitazione a 225 rpm. Estrarre il DNA plasmidico dalla coltura notturna attraverso una preparazione di DNA plasmidico di lisi alcalina52.
- Trasformare il DNA plasmidico in cellule Agrobacterium ceppo GV3101 utilizzando il metodo freeze-thaw. Lasciare scongelare 100 μL di cellule chimicamente competenti sul ghiaccio, aggiungere 1-5 μL di plasmide e incubare sul ghiaccio per 30 minuti. Mettere in azoto liquido per 1 min, quindi incubare a 37 °C per 3 min. Aggiungere 1 mL di SOC, lasciare recuperare per 2-3 ore a 28 °C con agitazione, piastra su terreni LB selettivi rif, gent e kan e incubare a 28 °C per 2 giorni.
- Colonie di schermo per la presenza di inserto con colonnina PCR. Scegli una singola colonia batterica e mescolala in 30 μL di acqua. Impostare una reazione PCR aggiungendo 12,5 μL di miscela master di polimerasi, 1,25 μL di ogni primer da 10 μM, FM-5840F e FM-6138R, 3 μL della sospensione della colonia batterica e acqua per un volume finale di 25 μL. Ciclo 35 volte con una temperatura di ricottura di 64 °C e un tempo di estensione di 1 minuto (1 min per ogni kb amplificato).
- Inoculare 2-5 ml di LB liquido (rif, gent, kan) con la colonia di Agrobacterium corretta. Lasciare crescere per una notte a 28 °C con agitazione a 225 giri/min.
- Mescolare la coltura notturna con una soluzione di glicerolo al 50% 1:1. Conservare a -80 °C per la conservazione a lungo termine.
- La PCR amplifica i frammenti del gene del mais da silenziare (ad esempio, les22 o pds) utilizzando un primer in avanti con un sito di restrizione PacI e un primer inverso con un sito di restrizione XbaI. Ciò consentirà la legatura dei frammenti genici nel MCS1* del vettore binario FoMV-pCAMBIA1380 nell'orientamento antisenso.
- Costrutto di espressione FoMV
- La PCR amplifica la sequenza di codifica di interesse, compresi i codoni di avvio e arresto (ad esempio, GFP) come descritto in 1.1.1, aggiungendo un sito di restrizione Bsu36I sul primer anteriore e un sito di restrizione PspOMI sul primer inverso per consentire la clonazione direzionale nell'orientamento del senso in MCS2.
- Eseguire la purificazione PCR utilizzando un kit di purificazione PCR in base alle specifiche del kit.
- Digerire il prodotto PCR e il FoMV-EV con gli enzimi di restrizione Bsu36I e PspOMI, come descritto al punto 1.1.3.
- Ligate il prodotto PCR digerito e FoMV-EV insieme a T4 DNA ligasi secondo il protocollo del produttore.
- Trasformare in DH5α cellule di E. coli chimicamente competenti utilizzando il metodo dello shock termico descritto al punto 1.1.5. Piastra su terreni LB selettivi di kanamicina e incubazione a 37 °C durante la notte.
- Controllare le colonie per cloni accurati mediante sequenziamento Sanger come descritto in 1.1.6 utilizzando i primer 5AmuS2 e 5AmuA2 (Tabella supplementare 1).
- Inoculare 2 mL di LB liquido con la colonia scelta e incubare a 37 °C durante la notte con agitazione a 225 RPM. Estrarre il DNA plasmidico dalla coltura notturna attraverso una preparazione di DNA plasmidico di lisi alcalina52.
- Trasformare il DNA plasmidico in cellule chimicamente competenti del ceppo Agrobacterium GV3101 utilizzando il metodo del congelamento-disgelo come descritto al punto 1.1.8. Piastra su terreni LB selettivi rif, gent e kan e incubare a 28 °C per 2 giorni.
- Schermare colonie per la presenza di inserto con colonia PCR utilizzando i primer 5AmuS2 e 5AmuA2.
- Inoculare 2-5 ml di LB liquido (rif, gent, kan) con la colonia di Agrobacterium corretta. Agitare per una notte a 225 giri/min a 28 °C.
- Mescolare la coltura notturna con una soluzione di glicerolo al 50% 1:1. Conservare a -80 °C per la conservazione a lungo termine.
- Costrutto di espressione SCMV
- La PCR amplifica il gene di interesse (ad esempio, GFP) escludendo il codone di arresto come descritto al punto 1.1.1, incluso un sito di digestione PspOMI sul primer anteriore e un sito di digestione SbfI sul primer inverso per consentire la clonazione direzionale nel vettore binario SCMV-pCAMBIA1380.
NOTA: l'inserto deve essere clonato in frame con la poliproteina virale. - Eseguire la purificazione PCR utilizzando un kit di purificazione PCR in base alle specifiche del kit.
- Digerire il prodotto PCR e l'SCMV-EV con gli enzimi di restrizione PspOMI e SbfI, come descritto al punto 1.1.3.
- Ligate il prodotto PCR digerito e SCMV-EV insieme a T4 DNA ligasi secondo il protocollo del produttore.
- Trasformare il prodotto in celle E. coli chimicamente competenti DH5α utilizzando il metodo dello shock termico descritto al punto 1.1.5. Piastra su terreno LB selettivo kan e incubazione a 37 °C durante la notte.
- Selezionare le colonie per cloni accurati mediante sequenziamento Sanger come descritto in 1.1.6 utilizzando i primer SC-745F e HCProR1 (Tabella supplementare 1).
- Inoculare 2 mL di LB liquido con la colonia scelta e incubare a 37 °C durante la notte con agitazione a 225 rpm. Estrarre il DNA plasmidico dalla coltura notturna attraverso una preparazione di DNA plasmidico di lisi alcalina52.
- Trasformare il DNA plasmidico in cellule chimicamente competenti del ceppo Agrobacterium GV3101 utilizzando il metodo del gelo-disgelo come descritto al punto 1.1.8. Piastra su terreni LB selettivi rif, gent e kan e incubare a 28 °C per 2 giorni.
- Screening colonie per presenza di inserto con colonia PCR con i primer SC-745F e HCProR1 come descritto in 1.1.9.
- Inoculare 2-5 ml di LB liquido (rif, gent, kan) con la colonia di Agrobacterium corretta. Agitare per una notte a 225 giri/min a 28 °C.
- Mescolare la coltura notturna con una soluzione di glicerolo al 50% 1:1. Conservare a -80 °C per la conservazione a lungo termine.
- La PCR amplifica il gene di interesse (ad esempio, GFP) escludendo il codone di arresto come descritto al punto 1.1.1, incluso un sito di digestione PspOMI sul primer anteriore e un sito di digestione SbfI sul primer inverso per consentire la clonazione direzionale nel vettore binario SCMV-pCAMBIA1380.
2. Preparazione della piantina
- Piantare 1-2 semi di mais (mais dolce 'Golden Bantam', FR1064, B73, ecc.) in substrato di coltivazione a base di torba in piccoli inserti posti all'interno di vassoi 4-7 giorni prima dell'iniezione. Posto in una camera di crescita sotto 16 giorni h a 25 °C e 8 ore notti a 22 °C (~ 185 radiazione fotosinteticamente attiva (PAR)) o in una serra sotto 16 giorni a 22-25 °C e 8 ore notti a 22-25 °C (350-400 PAR).
NOTA: La suscettibilità all'Agrobacterium varia tra i genotipi di mais, influenzando i tassi di successo. Inoltre, alcuni vettori virali possono essere incompatibili con alcuni genotipi di mais. - Annaffiare regolarmente e concimare una volta alla settimana con 15-5-15 fertilizzanti liquidi a 330 parti per milione (PPM).
3. Preparazione di Agrobacterium
- Un giorno prima dell'iniezione, preparare il mezzo liquido LB con l'antibiotico appropriato (rif, gent, kan) e inoculare con il ceppo Agrobacterium che trasporta il costrutto virale desiderato. Si raccomanda di aggiungere 20 μL di brodo di glicerolo in 50 ml di LB, che dovrebbe produrre abbastanza coltura batterica per inoculare >100 piante e può essere scalato verso l'alto o verso il basso secondo necessità.
NOTA: Preparare abbastanza inoculo per avere una quantità finale di sospensione batterica di almeno 1 mL ad una densità ottica di 600 nm (OD600) di 1,0 per ogni 4-5 piante. - Agitare a 225 giri/min a 28 °C per 24 ore.
- Batteri del pellet per 10 minuti a 4.000 x g a temperatura ambiente. Scartare il surnatante.
- Lavare accuratamente il pellet con 1 mL di acqua deionizzata (DI) mediante pipettaggio o vortice delicato.
- Ripetere il passaggio 3.3 ai batteri del pellet.
- Risospesso il pellet in 1 mL di soluzione MgSO4 da 10 mM mediante pipettaggio o vortice delicato.
- Facoltativamente, aggiungere 200 μM di acetosiringone alla soluzione. Sebbene comunemente usato, l'acetosiringone migliora solo la capacità di trasformazione di alcuni ceppi di Agrobacterium . Gli autori non hanno trovato che l'aggiunta di acetosiringone influisce sull'efficienza in questo protocollo (Tabella supplementare 2).
NOTA: la soluzione di MgSO4 da 10 mM può essere ottenuta da una soluzione madre da 1 M con un pH di 6,3 conservato a temperatura ambiente. La soluzione probabilmente non richiederà la regolazione del pH.
- Facoltativamente, aggiungere 200 μM di acetosiringone alla soluzione. Sebbene comunemente usato, l'acetosiringone migliora solo la capacità di trasformazione di alcuni ceppi di Agrobacterium . Gli autori non hanno trovato che l'aggiunta di acetosiringone influisce sull'efficienza in questo protocollo (Tabella supplementare 2).
- Misurare OD600 del campione con uno spettrofotometro e diluire a 1.0 OD600 con 10 mM di soluzione MgSO4 .
NOTA: questo è un punto di arresto sicuro. La sospensione batterica può essere mantenuta a temperatura ambiente fino a 5 ore prima dell'iniezione.
4. Iniezione
NOTA: le piantine di mais di 4-7 giorni possono essere utilizzate per l'iniezione. Il tasso di crescita della piantina è fortemente influenzato dalle condizioni di crescita, dalla quantità di PAR (cioè par più alto in serra che in camera di crescita) e dal genotipo, tra le altre cose che può essere difficile da controllare in condizioni di serra. Le piante possono essere iniettate a partire da 4 giorni quando sono alte 2-3 cm senza foglie espanse e vecchie come 7 giorni quando viene espansa la foglia più bassa a punta arrotondata. Il tasso di successo di questi metodi di inoculazione diminuisce rapidamente man mano che le piante invecchiano oltre i 7 giorni dopo la semina. Il sito di iniezione è lo stesso indipendentemente dall'età delle piantine.
- Indossando occhiali di sicurezza, iniettare la sospensione batterica nelle piantine 2-3 mm sopra il nodo coleoptilare utilizzando un ago da 25G x 5/8" attaccato a una siringa monouso da 1 mL.
NOTA: il nodo coleoptilar è dove alla fine si formeranno le radici della corona. Questo è il nodo più basso della pianta. In genere, ci sarà un cambiamento di colore da verde a bianco in corrispondenza e sotto il nodo. Il luogo di iniezione è appena sopra il meristema. Sezionare alcune piantine in questa fase può aiutare a visualizzare la posizione del meristema e di conseguenza il sito di iniezione corretto. - Applicare una leggera pressione sulla siringa fino a quando la sospensione riempie il coleoptile o è visibile nella spirale, a seconda della fase di crescita delle piante. Si tratta di circa 100-200 μL di sospensione.
NOTA: Se è difficile iniettare la sospensione nella piantina, il sito di iniezione potrebbe essere troppo basso. Una pressione moderata è tutto ciò che dovrebbe essere necessario per iniettare la sospensione. - Iniettare tutte le piantine, cambiando siringhe e aghi per ogni costrutto.
5. Cura continua delle piante
- Trapiantare le piantine iniettate in vasi 13 x 13 x 15 cm o più grandi quando hanno 7-8 giorni.
- Mantenere le condizioni di crescita (fotoperiodo di 16 ore e concimazione una volta alla settimana).
6. Conferma dell'infezione (fenotipicamente e RT-PCR)
- Punteggio fenotipico delle piante tra 14-21 DPI. Le lesioni da silenziamento della lesione dei geni di controllo mimica 22 o fitoene desaturasi possono essere facilmente viste sulle foglie e sono distinte dai sintomi foMV. L'espressione GFP può essere rilevata tramite imaging al microscopio fluorescente o altre immagini a luce UV.
NOTA: Alcuni costrutti / vettori virali possono richiedere più tempo per mostrare i sintomi o potrebbero non mostrare alcun sintomo. Le condizioni di luce elevata aumentano notevolmente i fenotipi causati dal silenziamento della lesione mimica 22 e della fitoene desaturasi. Le lesioni possono essere meno visibili o assenti se le piante vengono mantenute in condizioni di scarsa illuminazione come una camera di crescita, tuttavia il tasso di infezione effettivo determinato dalla RT-PCR non deve essere influenzato (Tabella 1). - Per confermare l'infezione molecolarmente, campionare la foglia 6 tra 14-21 DPI ed estrarre l'RNA totale utilizzando un'estrazione fenolo-cloroformio secondo le istruzioni del produttore.
- Utilizzo dell'RNA estratto come modello per generare cDNA di primo filamento.
- Impostare la reazione del cDNA con un massimo di 5 μg di RNA totale, 1 μL di primer esametrici casuali, 1 μL di primer oligo (dT)18 , 1 μL di dNTPs, 1 μL di trascrittasi inversa e acqua per un volume finale di 14,5 μL.
- Utilizzando primer progettati per il costrutto virale e il cDNA come modello, eseguire la PCR su ciascun campione per confermare l'infezione virale e determinare l'integrità del gene o del frammento genico di interesse come descritto in 1.1.1, tranne ridurre i cicli a 25 per FoMV e 30 per SCMV per evitare falsi positivi.
- Per i costrutti di silenziamento FoMV, utilizzare i primer FM-5840F e FM-6138R per amplificare attraverso l'MCS1 *, che contiene il frammento del gene del mais. Per i costrutti di espressione FoMV, utilizzare i primer 5AmuS2 e 5AmuA2 per amplificare attraverso l'MCS2, che contiene il gene inserito.
- Per i costrutti di espressione SCMV, utilizzare i primer SC745-F e HCProR1 per amplificare attraverso l'MCS, che contiene il gene inserito (Figura supplementare 3).
- Per un gene di controllo endogeno, utilizzare i primer ZmActS e ZmActA, che amplificano un frammento di mRNA di actina di mais (GRMZM2G126010) o primer ZmUbiF e ZmUbiR, che amplificano un frammento di mRNA di poliubiquitina di mais (GRMZM2G409726_T01).
- Visualizza il prodotto PCR su un gel di agarosio all'1% contenente una macchia di acido nucleico per determinare la presenza o l'assenza di virus e frammento di gene o gene.
Risultati
L'obiettivo di questo studio era quello di sviluppare un protocollo semplice per introdurre direttamente virus ricombinanti progettati per il silenziamento genico o l'espressione genica nelle piantine di mais (Figura 2). I vettori virali che trasportano inserti sono progettati e clonati utilizzando tecniche standard di biologia molecolare. I frammenti genici per il silenziamento vengono inseriti in MCS1* in FoMV-EV e le sequenze codificanti per l'espressione vengono inserite in FoMV-EV a MCS2 o SCMV-EV a MCS. I plasmidi risultanti vengono trasferiti al ceppo Agrobacterium GV3101. Successivamente, le piantine di mais vengono iniettate entro una settimana o meno dalla semina. Due settimane dopo l'iniezione, le piante possono essere valutate sia fenotipicamente che molecolarmente per l'infezione virale, il silenziamento genico e l'espressione genica.
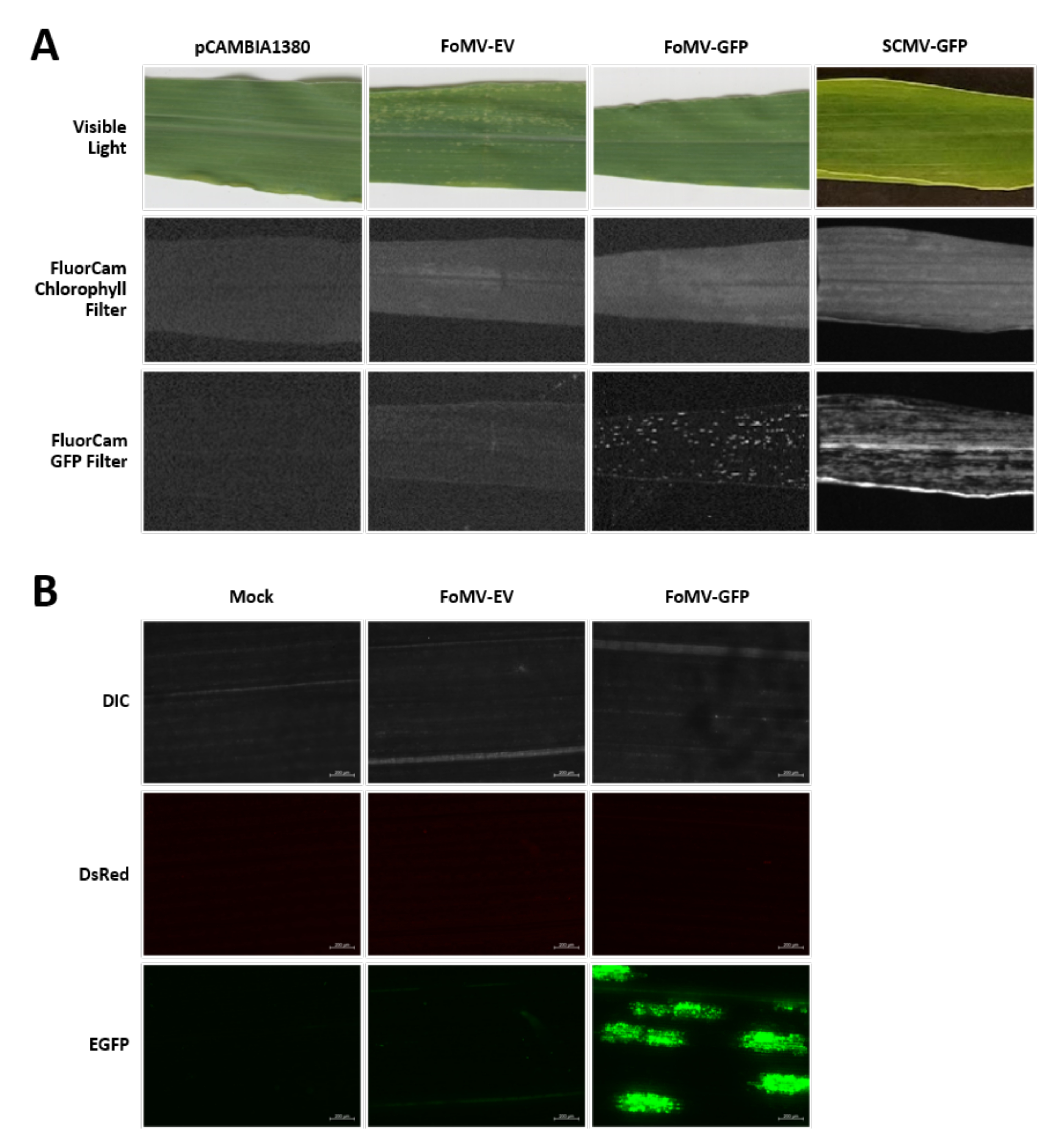
Le piante di mais vengono coltivate in un mezzo a base di torba per 4-7 giorni. In questa fase, il meristema apicale di germoglio si trova appena sopra il nodo coleoptilare (Figura 3A). Dopo che il coleoptile si è esteso di 2-3 centimetri o fino a 7 giorni dopo la semina, le piante vengono iniettate 2-3 mm sopra il nodo coleoptilare (Figura 3B-F). A circa 12 giorni dall'iniezione, le piante inizieranno a mostrare fenotipi di silenziamento sulle foglie, comunemente osservati vicino al tessuto vascolare, e queste lesioni sono visivamente distinte dai sintomi del mosaico virale FoMV (Figura 4). Sia la presenza di FoMV che il silenziamento dei geni bersaglio sono rilevabili nelle piante iniettate (Figura 5). L'espressione della GFP può essere rilevata entro 2 settimane dall'iniezione al microscopio fluorescente ed è più forte sulle foglie 5-7 (Figura 6). Se osservata sotto un sistema di imaging a fluorescenza, l'espressione GFP da FoMV può essere visualizzata come molte piccole aree puntiche di fluorescenza distribuite tra le foglie vicino al tessuto vascolare mentre l'espressione GFP da SCMV è costituita da patch più grandi (Figura 6, Figura supplementare 1). Sebbene i sintomi del mosaico virale siano spesso visibili sulle piante infette da costrutti di silenziamento FoMV, le piante iniettate con costrutti di espressione GFP che esprimono con successo GFP spesso non hanno questi sintomi. Di conseguenza, una pianta senza sintomi visibili può ancora essere positiva per l'espressione di virus e GFP. Inoltre, la perforazione del meristema durante la procedura di agroiniezione dovrebbe essere evitata in quanto ciò può causare difetti morfologici, ma le piante risultanti sopravvivono e sono spesso sintomatiche (Figura 7).
Sebbene questo protocollo sia stato originariamente sviluppato utilizzando mais dolce, diverse linee inbred di mais possono essere inoculate con successo con costrutti di silenziamento genico FoMV utilizzando l'agroiniezione. Ad esempio, FR1064 e B73 hanno in genere alti tassi di infezione virale (Tabella 2). In particolare, Mo17, una linea con nota resistenza genetica a FoMV, ha avuto un'efficienza di infezione dello 0% come previsto36,53. Inoltre, il costrutto utilizzato influenza l'efficienza dell'infezione (Tabella 3). Nel caso di FoMV, FoMV-EV e FoMV-LES22 hanno in genere le più alte efficienze di infezione rispettivamente al 53% e al 54%. FoMV-PDS ha un'efficienza leggermente inferiore al 38% e FoMV-GFP è il più basso al 17%. SCMV-GFP ha un'efficienza di infezione dell'8%. Queste percentuali sono medie su diversi esperimenti; i singoli esperimenti possono avere efficienze di infezione più o meno elevate.

Figura 1: Rappresentazioni schematiche dei cloni di FoMV e SCMV T-DNA utilizzati per l'agroiniezione nel mais. Il vettore FoMV contiene due siti di clonazione multipli (MCS1* e MCS2). Il vettore vuoto, FoMV-EV, è di 7.269 bp e non contiene alcun inserto in nessuno dei due MCS. (A) Il silenziamento genico utilizzando il vettore FoMV può essere ottenuto inserendo frammenti di geni in MCS1*, designato come sequenza di interesse (SOI), tipicamente nell'orientamento anti-senso. (B) L'espressione genica utilizzando il vettore FoMV può essere realizzata inserendo ORF del gene nell'MCS2 nell'orientamento del senso, designato come SOI. (C) Il vettore SCMV è stato progettato per avere un MCS tra P1 e HCPro. Il vettore vuoto, SCMV-EV, è di 11.015 bp e non contiene alcun inserto nell'MCS. Gli ORF genici inseriti nella MCS che sono in frame con la poliproteina SCMV saranno espressi come proteine. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Riepilogo schematico del protocollo agroiniezione. (A) Clonare SOI, un CDS o un frammento genico, nel vettore virale e trasformarsi in Agrobacterium ceppo GV3101. (B) Piantare mais e crescere per 4-7 giorni. (C) Coltivare GV3101 in coltura liquida durante la notte a 28 °C. (D) Preparare la sospensione batterica per iniezione. (E) Iniettare piantine 2-3 mm sopra il nodo coleoptilare con 100-200 μL di sospensione. (F) Trapiantare piantine quando hanno 7 giorni in vasi più grandi e crescere per 2-3 settimane fino a quando la 5a foglia è visibile. Fenotipo se lo si desidera. (G) Foglia campione 5 ed estratto di RNA. (H) Produrre cDNA e condurre PCR per amplificare virus / SOI. (I) Eseguire su gel per l'analisi qualitativa per determinare la presenza / assenza di virus e un SOI troncato o intatto. Questa figura è stata creata con BioRender.com. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Metodo di agroiniezione per inoculare piantine appena sopra il nodo coleoptile. (A) Piante vecchie di 4-5 giorni. Il coleoptile è completamente espanso e la prima vera foglia può essere parzialmente visibile, ma non è spiegata. (B) piante di 6-7 giorni. La prima foglia può essere espansa ma non saranno visibili collari. Anche la seconda foglia sarà visibile e potrebbe iniziare a dispiegarsi in questa fase. (C) Dissezione di piante di 6-7 giorni che mostrano la posizione del meristema apicale del germoglio in relazione al nodo coleoptile. (D) Iniezione di piante di 4-5 giorni. (E) Iniezione di piante di 6-7 giorni. (F) Iniezione di piante di 6-7 giorni utilizzando una soluzione colorante, che mostra l'inoculo tinto che esce dalla spirale della piantina. (G) Primo piano del sito di iniezione di piante di 6-7 giorni in relazione al nodo coleoptile. (H) Primo piano di una pianta di 6-7 giorni dopo l'iniezione, che mostra l'inoculo tinto nella spirale della pianta. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Sintomi dei geni di controllo del silenziamento utilizzati negli esperimenti di agroiniezione. (A) Una foglia fotografata a 17 DPI dopo che la pianta è stata iniettata con FoMV-LES22. FoMV-LES22 trasporta un inserto di 329 bp del CDS 3' del gene della lesione mimica del mais 22 nell'orientamento antisenso. Il silenziamento provoca l'accumulo di un metabolita tossico che a sua volta causa le lesioni necrotiche che appaiono per la prima volta come striature lungo la vascolarizzazione e crescono in chiazze più grandi come mostrato qui. (B) Una foglia fotografata a 17 DPI dopo che la pianta è stata iniettata con FoMV-PDS. FoMV-PDS trasporta un inserto di coppia di basi 313 del CDS 3' del gene del fitoene desaturasi del sorgo nell'orientamento antisenso. Il silenziamento dei pds nel mais provoca un fenotipo di fotosbiancamento sistemico che inizia come piccole e sottili striature lungo la vascolarizzazione che crescono in strisce più lunghe lungo la lunghezza della foglia, come mostrato qui. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: qRT-PCR di piante iniettate con costrutti di silenziamento genico FoMV. Conferma dell'infezione sistemica da FoMV e del silenziamento genico indotto dai costrutti FoMV-LES22 e FoMV-PDS erogati tramite agroiniezione in piante di mais dolce (Golden x Bantam). (A) Le immagini in gel mostrano analisi RT-PCR che confermano la presenza di vettore vuoto FoMV-MCS1* (amplicon 315 bp) e FoMV-PDS (amplicon 625 bp) nella foglia 6 di cinque singole piante. I primer PCR utilizzati producono un amplicon che si estende su MCS1*. L'actina del gene del mais (Zm-Actin) amplicon funge da gene di riferimento. Il grafico a barre rappresenta i valori di espressione relativa qRT-PCR per l'espressione pds nella foglia 6 a 37 giorni dopo l'inoculazione (DPI) mediante agroiniezione con FoMV-MCS1* o FoMV-PDS. La soppressione di pds è rilevabile in ciascuna delle cinque repliche biologiche (p = 0,003; test di Dunnett post hoc; le barre di errore indicano la deviazione standard (SD) di tre repliche tecniche). (B) Le immagini in gel mostrano analisi RT-PCR che confermano la presenza di FoMV-MCS1* (amplicon 315 bp) nella foglia 6 di cinque singole piante. FoMV-LES22 (625 bp amplicon) è stato rilevato nel tessuto fogliare 6 (campioni FoMV-LES22 1-5, 38 DPI) e foglia 4 (campioni FoMV-LES22 6-10, 20 DPI) per dieci singole piante. L'amplicon Zm-Actin fungeva da gene di riferimento. Il grafico a barre rappresenta i valori di espressione relativa qRT-PCR per l'espressione di les22 nei tessuti di mais mediante agroiniezione di costrutti virali FoMV-MCS1* o FoMV-LES22. La soppressione di Les22 avviene in 9 delle 10 repliche biologiche (p=<0.0001; test di Dunnett post hoc; le barre di errore indicano SD per tre repliche tecniche). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 6. Fenotipi di vari costrutti utilizzati negli esperimenti di agroiniezione. Tutte le piante fotografate sono state iniettate quando avevano 6-7 giorni con agrobatterio ceppo GV3101 che trasportava i costrutti indicati. Le immagini sono state scattate a 16 DPI. (A) Sintomi fogliari di pCAMBIA1380 (spina dorsale plasmidica vuota), FoMV-EV, FoMV-GFP e SCMV-GFP in luce visibile, sotto il filtro clorofilla FluorCam a 250 μs di esposizione e sotto il filtro FluorCam GFP a 10 ms di esposizione. (B) Immagini al microscopio fluorescente delle foglie di impianti iniettati con trattamento simulato (iniettati solo con soluzione di MgSO4), FoMV-EV e FoMV-GFP. Vengono mostrati i canali DIC, DsRed ed EGFP e sono stati presi ciascuno con un'esposizione di 1500 ms. La barra della scala è di 200 μm. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 7. Effetti morfologici dell'iniezione. Un esempio degli effetti morfologici più gravi che possono verificarsi dall'iniezione diretta nel tessuto meristematico. Questa lesione può causare la "triturazione" delle foglie e la scissione del gambo. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Virus | Condizioni di crescita | Genotipo | # Piante infette | Numero totale di piante | % Infezione | Percentuale media di infezione |
| FoMV-EV | Camera di crescita | Mais | 22 | 23 | 96% | 97% |
| B73 · | 18 | 18 | 100% | |||
| B104 · | 20 | 21 | 95% | |||
| Serra | Mais | 20 | 23 | 87% | 89% | |
| B73 · | 17 | 18 | 94% | |||
| B104 · | 16 | 19 | 84% | |||
| SCMV-EV | Camera di crescita | Mais | 14 | 21 | 67% | 47% |
| B73 · | 5 | 18 | 28% | |||
| B104 · | 10 | 21 | 48% | |||
| Serra | Mais | 14 | 23 | 61% | 49% | |
| B73 · | 0 | 19 | 0% | |||
| B104 · | 19 | 22 | 86% |
Tabella 1: Effetto delle condizioni di serra e camera di crescita sull'efficienza dell'inoculazione agroiniezione. I semi sono stati germinati in condizioni di crescita identiche. Le piantine germinate sono state agroiniettate e metà di esse sono state spostate in una camera di crescita (25 °C 16 h luce diurna / 22C 8 h notte; 185 PAR) e l'altra metà è stata spostata in una serra (22-25 °C 16 h luce diurna / 22-25 °C 8 h notte; 350-400 PAR). Questa tabella riporta il tasso di infezione in percentuale, calcolato dal numero di piante confermate dalla RT-PCR per essere infettate dal rispettivo virus diviso per il numero totale di piante agroiniettate. Non vi è alcuna differenza statistica nell'efficienza dell'infezione tra camera di crescita e condizioni di serra (FoMV due code t-test p = 0,08; SCMV due coda t-test p=0,96).
| Genotipo del mais | FoMV-EV | FoMV-LES22 · | Totale combinato | ||||
| Infetto | Totale | % infetti | Infetto | Totale | % infetti | % infetti | |
| Mais | 18 | 23 | 78% | 15 | 23 | 65% | 72% |
| MO47 · | 7 | 22 | 32% | 1 | 21 | 5% | 19% |
| K55 · | 1 | 15 | 7% | 3 | 17 | 18% | 13% |
| W64A · | 10 | 22 | 45% | 8 | 20 | 40% | 43% |
| MO17 · | 0 | 16 | 0% | 0 | 13 | 0% | 0% |
| B73 · | 10 | 18 | 56% | 7 | 17 | 41% | 49% |
| B101 · | 12 | 21 | 57% | 8 | 24 | 33% | 44% |
| FR1064 · | 4 | 4 | 100% | 4 | 4 | 100% | 100% |
| B104 · | 10 | 22 | 45% | 5 | 21 | 24% | 35% |
| WCC22 · | 2 | 7 | 29% | 4 | 6 | 67% | 46% |
| A188 · | 0 | 3 | 0% | 4 | 6 | 67% | 44% |
Tabella 2: Efficienza di infezione dei costrutti FoMV tra i genotipi di mais. FoMV-EV e FoMV-LES22 sono stati agroiniettati in 11 genotipi di mais. Dopo l'iniezione, le piantine sono state spostate nella serra. Questa tabella descrive il tasso di infezione in percentuale, calcolato dal numero di piante infette da FoMV come confermato dalla RT-PCR diviso per il numero totale di piante agroiniettate. Il tasso totale combinato di infezione mostra i tassi medi di infezione di ciascun genotipo per entrambi i costrutti FoMV testati.
| Fase dell'impianto | Piante vecchie di 4-5 giorni | Piante vecchie di 6-7 giorni | Totale combinato | ||||
| Sintomatico | Totale Impianti | % infetti | Sintomatico | Totale Impianti | % infetti | % infetti | |
| FoMV-EV | 42 | 72 | 58% | 80 | 170 | 47% | 53% (A) |
| FoMV-PDS | 65 | 157 | 41% | 66 | 184 | 36% | 39% (B C) |
| FoMV-LES22 · | 115 | 195 | 59% | 144 | 292 | 49% | 54% (A B) |
| FoMV-GFP | 16 | 103 | 16% | 37 | 217 | 17% | 16% (C) |
| SCMV-GFP | 10 | 95 | 11% | 5 | 82 | 6% | 8% (C) |
Tabella 3: Riepilogo degli esperimenti di iniezione. Questa tabella rappresenta un riassunto degli esperimenti di iniezione condotti da agosto 2017 ad agosto 2018 su piantine di mais dolce Golden Bantam. Le piante sono state valutate per i sintomi virali (FoMV-EV), i sintomi di silenziamento (pds e les22) o la fluorescenza GFP (GFP) attraverso lo screening visivo (FoMV-EV, FoMV-PDS e FoMV-LES22) o FluorCam (FoMV-GFP e SCMV-GFP). I risultati sono mostrati individualmente per piante di 4-5 giorni e piante di 6-7 giorni, nonché un riepilogo di tutte le età delle piante. Non vi è alcuna differenza significativa riscontrata tra piante di 4-5 giorni e piante di 6-7 giorni (ANOVA unidirezionale, F = 0,6513). C'è una differenza trovata tra il costrutto virale (Onaway ANOVA, F = <0.0001), con le lettere che rappresentano il rapporto sulle lettere di collegamento Tukey-Kramer HSD.
Tabella supplementare 1: Tabella che elenca tutti i nomi e le sequenze di primer utilizzati in questo protocollo. Fare clic qui per scaricare questa tabella.
Tabella supplementare 2: Test dell'acetosiringone. (A) Test iniziale dell'acetosiringone, confrontando i tassi dei sintomi di piante iniettate finte, FoMV-EV e FoMV-LES22 tra sospensioni di inoculazione con acetosiringone (+) da 200 μM o senza acetosiringone (-). (B) Confronto dei tassi di infezione da FoMV-LES22 determinati dalla RT-PCR tra sospensioni di inoculazione senza acetosiringone (-), con 200 μM di acetosiringone (+), e aggiunta di 20 μM di acetosiringone alla coltura batterica 4 ore prima della risospensione in tampone insieme all'aggiunta di 200 uM di acetosiringone alla sospensione finale (++). Nel complesso, non è stata riscontrata alcuna differenza significativa tra i trattamenti con aceotiringone (Oneway ANOVA, f = 0,5452). Fare clic qui per scaricare questa tabella.
Figura supplementare 1: Imaging a fluorescenza e validazione molecolare di SCMV agroiniettato ed espressione di proteine eterologhe nel mais. Il mais è stato agroiniettato con un costrutto SCMV modificato contenente sia CDS di GFP che nano luciferasi (NLuc). (A) L'imaging Fluorcam è stato utilizzato per lo screening e il rilevamento della GFP. La sinistra è un impianto iniettato simulato e il destro è un impianto iniettato SCMV-NLucGFP. (B) Gli estratti di proteine fogliari sono stati separati da SDS-PAGE e valutati per la presenza di NLuc, GFP e scMV coat protein (CP) mediante test in-gel luciferasi o immunoblot come indicato. Fare clic qui per scaricare questo file.
Discussione
Agrobacterium è uno strumento essenziale che facilita numerose tecniche di biologia molecolare nella ricerca legata alle piante. Questo studio fornisce un protocollo di agroiniezione per inoculare vettori virali FoMV e SCMV direttamente nei tessuti di mais per applicazioni VIGS e VOX. L'obiettivo principale è quello di aumentare la facilità e l'utilità delle tecnologie basate su virus per la ricerca nelle piante monocotiledoni. Sebbene l'agroinoculazione diretta del mais sia stata segnalata per alcuni virus, gli autori non sono a conoscenza di un protocollo dettagliato e non ci sono esempi di applicazioni VIGS e VOX in questi studi19,22.
È stato riportato, ed è stato confermato durante lo sviluppo di questo protocollo, che la posizione di iniezione è un fattore chiave per lanciare con successo un'infezione virale sistemica tramite agroiniezione19. Si presume che l'iniezione coerente della posizione raccomandata sulla pianta sia la variabile più grande, perché la posizione esatta del meristema nelle piantine di mais è praticamente non rilevabile a occhio. Per ridurre al minimo la variazione interpersonale, si consiglia di sezionare alcune piantine di mais fino al meristema per visualizzarne meglio la posizione (Figura 3C). La posizione del meristema in relazione al nodo coleoptilare dovrebbe essere all'incirca la stessa per le piante di età compresa tra 4 e 7 giorni. Inoltre, praticare l'iniezione con un liquido tinto fornisce una dimostrazione facilmente visibile di come l'"inoculo" riempie la spirale fogliare e, poiché il sito di iniezione è contrassegnato con colorante, l'accuratezza del sito di iniezione può essere confermata (Figura 3G, H). I tessuti meristematici sono i più suscettibili all'agroiniezione, ma l'iniezione di sospensioni di Agrobacterium direttamente in questo tessuto provoca effetti morfologici indesiderati (Figura 6)19. Le piante con meristemi danneggiati sopravvivono, ma i difetti risultanti sono indesiderabili e, quindi, l'iniezione diretta di questo tessuto dovrebbe essere evitata.
Ci sono diverse variabili che possono influire sul successo del lancio di un'infezione virale sistemica tramite agroiniezione perché tre sistemi biologici complessi (pianta, virus e ceppo Agrobacterium) devono interagire in coordinamento. Questa complessa interazione può essere aiutata dalle cellule in rapida divisione della regione meristematica, rendendola un luogo ideale per l'agroinoculazione19. Il ceppo Agrobacterium deve essere in grado di infettare le cellule dei tessuti vegetali per fornire il T-DNA che trasporta il genoma virale e la pianta deve essere suscettibile al virus per avviare la replicazione virale e l'infezione sistemica. I genotipi del mais differiscono nella loro suscettibilità ai virus (ad esempio, Mo17 è resistente ai ceppi di FoMV) o Agrobacterium, ma la maggior parte di quelli testati sembra essere suscettibile sia a FoMV che a SCMV (Tabella 1 e Tabella 2)53. Ad esempio, la linea inbred FR1064 e la varietà di mais dolce Golden Bantam possono essere particolarmente sensibili sia a GV3101 Agrobacterium che a vettori basati su FoMV.
Il numero di foglie campionate e la tempistica del campionamento per la RT-PCR sono fondamentali per una valutazione accurata dell'infezione virale. Negli esempi mostrati qui, il numero di foglie è stato determinato partendo dalla prima foglia arrotondata (comunemente nota come "foglia del pollice") e contando verso l'alto. Le foglie sono state campionate una volta che sono state espanse e la foglia successiva ha iniziato a emergere. Tuttavia, quali foglie sono ottimali per il campionamento potrebbero variare in base alle specie di virus utilizzate, alle condizioni di crescita e al genotipo del mais. Pertanto, si consiglia un esperimento iniziale di corso temporale quando si applica questo protocollo a un nuovo sistema di virus per ottimizzare la strategia di campionamento rispetto alle foglie e ai tempi.
Il costrutto specifico utilizzato influisce in modo significativo sull'efficienza di questo protocollo. Ad esempio, i vettori vuoti, FoMV-EV e SCMV-EV, e FoMV-PDS e FoMV-LES22, che contengono entrambi piccoli inserti (rispettivamente 313 bp e 329 bp), in genere producono le percentuali più alte di piante con sintomi virali in questi esperimenti (Tabelle 1 e Tabella 2). Tuttavia, i virus ricombinanti che trasportavano inserti più grandi del GFP ORF (720 bp) in FoMV-GFP e SCMV-GFP, avevano tassi di infezione molto più bassi rispetto alle piante iniettate con il vettore vuoto o i costrutti di silenziamento genico. Questa tendenza può essere dovuta agli impatti negativi sulla forma virale causati da quantità crescenti di materiale genetico esogeno nel genoma virale. Diversi studi hanno dimostrato che la stabilità dell'inserto dei vettori virali delle piante dipende in gran parte dalle dimensioni e dalla sequenza dell'inserto36,54,55,56,57. Inoltre, c'è stata una notevole differenza nella percentuale di piante che si infettano dopo l'inoculazione con il vettore vuoto FoMV o SCMV, suggerendo che è necessario un ulteriore lavoro per ottimizzare questo protocollo per SCMV (Tabella 1). Questi risultati indicano che potrebbe essere necessaria una certa risoluzione dei problemi durante lo sviluppo di un costrutto, perché la sequenza e la lunghezza del frammento possono entrambi influire sull'efficienza.
Nel complesso, questo studio ha dimostrato che l'agroiniezione di piantine di mais è un metodo di inoculazione efficace per due diversi virus vegetali a RNA, configurazioni vettoriali multiple e 11 genotipi di mais. Questo lavoro con FoMV e SCMV, abbinato a lavori precedenti che utilizzano l'iniezione con virus clorotico della chiazza di mais (MCMV) o MSV, indica che l'agroiniezione è adatta per inoculare piantine di mais con cloni infettivi di virus sia a RNA che a DNA19,20,21,22. Inoltre, questo lavoro mostra ulteriormente che l'agroiniezione è un metodo praticabile per i vettori VIGS e VOX e può essere applicato a piante di quattro giorni (Tabella 3). Il protocollo qui presentato dovrebbe essere prontamente adattato dai biologi del mais per facilitare la ricerca negli studi di genomica funzionale che coinvolgono il silenziamento genico transitorio (VIGS) e la sovraespressione (VOX). L'agroiniezione ha anche la capacità di facilitare gli approcci di modifica genetica basati su virus (VEdGE) che altrimenti sarebbero limitati dalla dipendenza dalla trasformazione delle piante, migliorando potenzialmente l'efficienza di editing e l'accessibilità58,59,60. Dato che il ceppo di Agrobacterium appropriato, i genotipi di mais e i vettori virali sono accuratamente combinati, l'inoculazione mediante agroiniezione dovrebbe diventare uno strumento prezioso per le analisi transitorie della funzione genica nel mais.
Divulgazioni
I ricercatori non hanno conflitti di interesse da rivelare.
Riconoscimenti
La Iowa State University fa parte di un team che supporta il programma Insect Allies della DARPA HR0011-17-2-0053. Questo lavoro è stato supportato anche dall'Iowa State University Plant Sciences Institute, dall'Iowa State University Crop Bioengineering Center, dal progetto USDA NIFA Hatch numero 3808 e dai fondi dello Stato dell'Iowa. K.L.H. è stato anche parzialmente supportato dal programma di formazione universitaria Iowa State University Predictive Plant Phenomics finanziato dalla National Science Foundation (DGE # 1545453) e dalla sovvenzione Agricultural and Food Research Initiative n. 2019-07318 dell'USDA National Institute of Food and Agriculture. I finanziatori non hanno avuto alcun ruolo nella progettazione dello studio e della raccolta, analisi e interpretazione dei dati e nella scrittura del manoscritto. Eventuali opinioni, risultati e conclusioni o raccomandazioni espresse in questo materiale sono quelle degli autori e non riflettono necessariamente le opinioni dei finanziatori.
Ringraziamo Nick Lauter (USDA-ARS, Ames, IA) per le sementi di linee inbred di mais, Christian F. Montes-Serey (Iowa State University) per aver realizzato il clone FoMV-GFP e Tyler Austin (Iowa State University) per l'assistenza tecnica.
Materiali
| Name | Company | Catalog Number | Comments |
| 1 mL syringes | Fisher Scientific | 14955450 | alternatively, BD 309659 |
| 15 mL Falcon Tubes | Corning Science | 352059 | |
| 1kb+ Ladder | ThermoFisher Scientific | 10787018 | For assessing sizes of PCR products |
| 25G x 5/8" PrecisionGlide Needles | Becton, Dickinson and Company (BD) | 305122 | |
| 28°C Incubator | For Agrobacterium | ||
| 37°C Incubator | For E. coli | ||
| Acetosyringone | MilliporeSigma | D134406 | Optional |
| Agar | MilliporeSigma | A4800 | |
| Agarose | GeneMate | E-3120 | For making gels to check for virus/insert stability |
| Agrobacterium tumefaciens Strain GV3101 | Carries vir plasmid encoding T-DNA transfer machinery, RifR, GmR, from lab stock | ||
| Bsu36I | New England Biolabs | R0524 | |
| cDNA Kit | ThermoFisher Scientific | K1672 | Maxima First Strand cDNA Synthesis Kit with Dnase |
| Chloroform | Fisher Scientific | C298 | For RNA extraction |
| Cuvettes | Fisher Scientific | 14955127 | 1.5 mL |
| D-(+)-Glucose | MilliporeSigma | G7528 | Alkaline Lysis |
| DH5alpha Competent E. coli Cells | New England Biolabs | C2987 | |
| DNA Ligase | ThermoFisher Scientific | K1422 | Rapid DNA Ligation Kit |
| EDTA (Ethylenediamine Tetraacetic Acid, Disodium Salt Dihydrate) | Fisher Scientific | S311 | Alkaline Lysis |
| Ethanol | For RNA extraction | ||
| Fertilizer | Peters Fertilizer 15-15-15 Concentrate | ||
| Flat Inserts | T.O. Plastics | 715357C | For germinating seeds in trays |
| Flats | T.O. Plastics | 710245C | For germinating seeds in trays |
| FluorCam | Photon Systems Instruments | To assess maize plants for GFP expression before microscope | |
| Fluorescence Microscope | |||
| Gel Electrophoresis Box | |||
| Gentamycin Sulfate | Fisher Scientific | BP918 | |
| Glacial Acetate | Fisher Scientific | A38 | Alkaline Lysis |
| Glycerol | Fisher Scientific | G33-500 | For saving frozen stocks of bacteria |
| Go-Taq, 2X | Promega | M7123 | |
| Hydrochloric Acid | Fisher Scientific | A144 | for pHing solutions |
| Isopropanol | Sigma-Aldrich | 109827 | For RNA extraction |
| Kanamycin, Monosulfate | Fisher Scientific | BP906 | |
| Large Pots | Kordlok | SQL0550 | 5x5x4" or bigger. For transplanting seedlings. |
| Luria Bertani (LB) Broth, Miller | Himedia | M1245 | |
| Magnesium Sulfate Heptahydrate | Amresco | 662 | |
| Maize Golden Bantam Sweet Corn Seed | American Meadows, West Coast Seeds | ||
| Maize Inbred Seed | Our seed comes from our institution, but we are not able to provide this for other researchers. | ||
| Maxima H Minus Reverse Transcriptase | ThermoFisher Scientific | EP0753 | |
| MilliQ | Elga | Purelab Ultra | |
| Monarch PCR & DNA Cleanup Kit | New England Biolabs | T1030 | |
| PacI | New England Biolabs | R0547 | |
| Peters Excel 15-5-15 Fertilizer | ICL Specialty Fertilizers | G99140 | |
| Petri Dish, 95 mm x 15 mm | Fisher Scientific | FB0875714G | |
| pH Meter | |||
| Potassium Acetate | Fisher Scientific | P171 | Alkaline Lysis |
| Primers | Our primers were synthesized through our institutional DNA facility or through IDT | ||
| PspOMI | New England Biolabs | R0653 | |
| Q5 High-Fidelity DNA Polymerase | New England Biolabs | M0491 | |
| Rifampicin | EMD Millipore Corp | 557303 | |
| Rnase A | ThermoFisher Scientific | 12091021 | Alkaline Lysis |
| SbfI | New England Biolabs | R0642 | |
| Scale | For weighing chemicals for media or buffers | ||
| SDS (Sodium Dodecyl Sulfate) | Fisher Scientific | BP166 | Alkaline Lysis |
| Sodium Hydroxide | Fisher Scientific | S318 | Alkaline Lysis |
| Soil Substrate | SunGro Horticulture | SS#1-F1P | Sunshine Mix #1/Fafard-1P, any soil mix that maize grows well in is sufficient |
| Spectrophotometer | For measuring OD600 | ||
| Sybr Safe, 10,000X | Invitrogen | S33102 | For making gels to check for virus/insert stability |
| Thermocycler | For PCR | ||
| Tris Base | Fisher Scientific | BP154 | Alkaline Lysis |
| Trizol | Ambion | 15596018 | For RNA extraction |
| Weigh Paper | For weighing chemicals for media or buffers | ||
| XbaI | New England Biolabs | R0145 |
Riferimenti
- Zaidi, S. E. A., Mansoor, S. Viral vectors for plant genome engineering. Frontiers in Plant Science. 8, 539(2017).
- Kant, R., Dasgupta, I. Gene silencing approaches through virus-based vectors: speeding up functional genomics in monocots. Plant Molecular Biology. 100, 3-18 (2019).
- Hu, J., et al. A barley stripe mosaic virus-based guide RNA delivery system for targeted mutagenesis in wheat and maize. Molecular Plant Pathology. 20 (10), 1463-1474 (2019).
- Pasin, F., Menzel, W., Daròs, J. A. Harnessed viruses in the age of metagenomics and synthetic biology: an update on infectious clone assembly and biotechnologies of plant viruses. Plant Biotechnology Journal. 17 (6), 1010-1026 (2019).
- Cody, W. B., Scholthof, H. B. Plant virus vectors 3.0: Transitioning into synthetic genomics. Annual Review of Phytopathology. 57 (1), 211-230 (2019).
- Mei, Y., et al. Protein expression and gene editing in monocots using foxtail mosaic virus vectors. Plant Direct. 3 (11), 00181(2019).
- Ruiz, M. T., Voinnet, O., Baulcombe, D. C. Initiation and maintenance of virus-induced gene silencing. Plant Cell. 10 (6), 937-946 (1998).
- Bekele, D., Tesfaye, K., Fikre, A. Applications of virus induced gene silencing (VIGS) in plant functional genomics studies. Journal of Plant Biochemistry & Physiology. 07 (01), 1000229(2019).
- Scholthof, H. B., Scholthof, K. B. G., Jackson, A. O. Plant virus gene vectors for transient expression of foreign proteins in plants. Annual Review of Phytopathology. 34 (1), 299-323 (1996).
- Holzberg, S., Brosio, P., Gross, C., Pogue, G. P. Barley stripe mosaic virus-induced gene silencing in a monocot plant. Plant Journal. 30 (3), 315-327 (2002).
- Wang, R., et al. An efficient virus-induced gene silencing vector for maize functional genomics research. Plant Journal. 86 (1), 102-115 (2016).
- Redinbaugh, M. G., et al. Transmission of viral RNA and DNA to maize kernels by vascular puncture inoculation. Journal of Virological Methods. 98 (2), 135-143 (2001).
- Scholthof, H. B. The capsid protein gene of tomato bushy stunt virus is dispensable for systemic movement and can be replaced for localized expression of foreign genes. Molecular Plant-Microbe Interactions. 6 (3), 309(1993).
- Scholthof, H. B., Scholthof, K. B. G., Kikkert, M., Jackson, A. O. Tomato bushy stunt virus spread is regulated by two nested genes that function in cell-to-cell movement and host-dependent systemic invasion. Virology. 213 (2), 425-438 (1995).
- Scholthof, H. B. Rapid delivery of foreign genes into plants by direct rub-inoculation with intact plasmid dna of a tomato bushy stunt virus gene vector. Journal of Virology. 73 (9), 7823-7829 (1999).
- Zhang, J., et al. Vacuum and co-cultivation agroinfiltration of (germinated) seeds results in tobacco rattle virus (TRV) mediated whole-plant virus-induced gene silencing (VIGS) in wheat and maize. Frontiers in Plant Science. 8, 393(2017).
- Vaghchhipawala, Z., Rojas, C. M., Senthil-Kumar, M., Mysore, K. S. Agroinoculation and agroinfiltration: simple tools for complex gene function analyses. Methods in Molecular Biology. 678, Clifton, N.J. 65-76 (2011).
- Grimsley, N., Hohn, B., Hohn, T., Walden, R. "Agroinfection," an alternative route for viral infection of plants by using the Ti plasmid. Proceedings of the National Academy of Sciences. 83 (10), 3282-3286 (1986).
- Grimsley, N. H., Ramos, C., Hein, T., Hohn, B. Merisfematic tissues of maize plants are most suscepnsle to agroinfection with maize streak virus. Bio/Technology. 6 (2), 185-189 (1988).
- Martin, D. P., Rybicki, E. P. Improved efficiency of Zea mays agroinoculation with Maize streak virus. Plant Disease. 84 (10), 1096(2000).
- Martin, D. P., Willment, J. A., Rybicki, E. P. Evaluation of maize streak virus pathogenicity in differentially resistant Zea mays genotypes. Phytopathology. 89 (8), 695-700 (1999).
- Wang, Q., et al. Further characterization of Maize chlorotic mottle virus and its synergistic interaction with Sugarcane mosaic virus in maize. Scientific Reports. 7, 39960(2017).
- Hsieh, M. H., et al. Optimizing virus-induced gene silencing efficiency with Cymbidium mosaic virus in Phalaenopsis flower. Plant Science. 201-202 (1), 25-41 (2013).
- Hsieh, M. H., et al. Virus-induced gene silencing unravels multiple transcription factors involved in floral growth and development in Phalaenopsis orchids. Journal of Experimental Botany. 64 (12), 3869-3884 (2013).
- Zenna, N. S., et al. Genetic analysis of tolerance to rice tungro bacilliform virus in rice (Oryza sativa L.) through agroinoculation. Journal of Phytopathology. 154 (4), 197-203 (2006).
- Marks, M. S., Kemp, J. M., Woolston, C. J., Dale, P. J. Agroinfection of wheat: A comparison of Agrobacterium strains. Plant Science. 63 (2), 247-256 (1989).
- Dasgupta, I., et al. Rice tungro bacilliform virus DNA independently infects rice after Agrobacterium-mediated transfer. Journal of General Virology. 72 (6), 1215-1221 (1991).
- Boulton, M. I., Buchholz, W. G., Marks, M. S., Markham, P. G., Davies, J. W. Specificity of Agrobacterium-mediated delivery of maize streak virus DNA to members of the Gramineae. Plant Molecular Biology. 12 (1), 31-40 (1989).
- Paulsen, A. Q. Purification and properties of foxtail mosaic virus. Phytopathology. 77 (11), 1346(1977).
- Bancroft, J. B., Rouleau, M., Johnston, R., Prins, L., Mackie, G. A. The entire nucleotide sequence of foxtail mosaic virus RNA. Journal of General Virology. 72 (9), 2173-2181 (1991).
- Bruun-Rasmussen, M., Madsen, C. T., Johansen, E., Albrechtsen, M. Revised sequence of foxtail mosaic virus reveals a triple gene block structure similar to potato virus X. Archives of Virology. 153 (1), 223-226 (2008).
- Rouleau, M., Bancroft, J. B., Mackie, G. A. Partial purification and characterization of foxtail mosaic potexvirus RNA-dependent RNA polymerase. Virology. 197 (2), 695-703 (1993).
- Rouleau, M., Smith, R. J., Bancroft, J. B., Mackie, G. A. Purification, properties, and subcellular localization of foxtail mosaic potexvirus 26-kDa protein. Virology. 204 (1), 254-265 (1994).
- Samuels, T. D., et al. Subcellular targeting and interactions among the potato virus X TGB proteins. Virology. 367 (2), 375-389 (2007).
- Cho, S. Y., Kim, K. H. Identification of the capsid protein-binding region of the SL1(+) RNA located at the 5' region of the potato virus X genome. Plant Pathology Journal. 28 (1), 75-80 (2012).
- Mei, Y., Zhang, C., Kernodle, B. M., Hill, J. H., Whitham, S. A. A foxtail mosaic virus vector for virus-induced gene silencing in maize. Plant Physiology. 171 (2), 760-772 (2016).
- Bouton, C., et al. Foxtail mosaic virus: A viral vector for protein expression in cereals. Plant Physiology. 177 (4), 1352-1367 (2018).
- Mei, Y., Liu, G., Zhang, C., Hill, J. H., Whitham, S. A. A sugarcane mosaic virus vector for gene expression in maize. Plant Direct. 3 (8), 00158(2019).
- Gal-On, A., Meiri, E., Huet, H., Hua, W. J., Raccah, B., Gaba, V. Particle bombardment drastically increases the infectivity of cloned DNA of zucchini yellow mosaic potyvirus. Journal of General Virology. 76 (12), (1995).
- Gao, R., et al. Construction of an infectious cDNA clone and gene expression vector of Tobacco vein banding mosaic virus (genus Potyvirus). Virus Research. 169 (1), 276-281 (2012).
- López-Moya, J. J., García, J. A. Construction of a stable and highly infectious intron-containing cDNA clone of plum pox potyvirus and its use to infect plants by particle bombardment. Virus Research. 68 (2), (2000).
- Choi, I. R., French, R., Hein, G. L., Stenger, D. C. Fully biologically active in vitro transcripts of the eriophyid mite-transmitted wheat streak mosaic tritimovirus. Phytopathology. 89 (12), (1999).
- Kim, K. S., et al. Infectivity of in vitro transcripts of Johnsongrass mosaic potyvirus full-length cDNA clones in maize and sorghum. Archives of Virology. 148 (3), 563-574 (2003).
- Stewart, L. R., Bouchard, R., Redinbaugh, M. G., Meulia, T. Complete sequence and development of a full-length infectious clone of an Ohio isolate of Maize dwarf mosaic virus (MDMV). Virus Research. 165 (2), 219-224 (2012).
- Wylie, S. J., et al. ICTV virus taxonomy profile: Potyviridae. Journal of General Virology. 98 (3), 352-354 (2017).
- Shukla, D. D. taxonomy of potyviruses infecting maize, sorghum, and sugarcane in Australia and the United States as determined by reactivities of polyclonal antibodies directed towards virus-specific N-termini of coat proteins. Phytopathology. 79 (2), 223(1989).
- Shukla, D. D., Ward, C. W. Amino Acid sequence homology of coat proteins as a basis for identification and classification of the potyvirus group. Journal of General Virology. 69 (11), 2703-2710 (1988).
- Chung, B. Y. W., Miller, W. A., Atkins, J. F., Firth, A. E. An overlapping essential gene in the Potyviridae. Proceedings of the National Academy of Sciences of the United States of America. 105 (15), 5897-5902 (2008).
- Jarchow, E., Grimsley, N. H., Hohn, B. virF, the host-range-determining virulence gene of Agrobacterium tumefaciens, affects T-DNA transfer to Zea mays. Proceedings of the National Academy of Sciences of the United States of America. 88 (23), 10426-10430 (1991).
- Hu, G., Yalpani, N., Briggs, S. P., Johal, G. S. A porphyrin pathway impairment is responsible for the phenotype of a dominant disease lesion mimic mutant of maize. The Plant Cell. 10 (7), 1095(2007).
- Qin, G., et al. Disruption of phytoene desaturase gene results in albino and dwarf phenotypes in Arabidopsis by impairing chlorophyll, carotenoid, and gibberellin biosynthesis. Cell Research. 17 (5), 471-482 (2007).
- Jones, P. Isolation of plasmid DNA from E. coli. Encyclopedia of Life Sciences. , (2003).
- Ji, Q., Yang, B., Lee, M., Chen, Y., Lübberstedt, T. Mapping of quantitative trait loci/locus conferring resistance to foxtail mosaic virus in maize using the intermated B73-×-Mo17 population. Plant Breeding. 129 (6), 721-723 (2010).
- Pacak, A., et al. The brome mosaic virus-based recombination vector triggers a limited gene silencing response depending on the orientation of the inserted sequence. Archives of Virology. 155 (2), 169-179 (2010).
- Miché, L., Battistoni, F., Gemmer, S., Belghazi, M., Reinhold-Hurek, B. Host-dependent expression of Rhizobium leguminosarum bv. viciae hydrogenase is controlled at transcriptional and post-transcriptional levels in legume nodules. Molecular Plant-Microbe Interactions. 19 (5), 1323-1331 (2018).
- Yamagishi, M., Masuta, C., Suzuki, M., Netsu, O. Peanut stunt virus-induced gene silencing in white lupin (lupinus albus). Plant Biotechnology. 32 (3), 181-191 (2015).
- Avesani, L., et al. Stability of Potato virus X expression vectors is related to insert size: Implications for replication models and risk assessment. Transgenic Research. 16 (5), 587-597 (2007).
- Ali, Z., et al. Efficient virus-mediated genome editing in plants using the CRISPR/Cas9 system. Molecular Plant. 8 (8), 1288-1291 (2015).
- Cody, W. B., Scholthof, H. B., Mirkov, T. E. Multiplexed gene editing and protein overexpression using a tobacco mosaic virus viral vector. Plant Physiology. 175 (1), 23-35 (2017).
- Ali, Z., Eid, A., Ali, S., Mahfouz, M. M. Pea early-browning virus-mediated genome editing via the CRISPR/Cas9 system in Nicotiana benthamiana and Arabidopsis. Virus Research. 244, 333-337 (2018).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati