Method Article
Enrichment of Bruch's Membrane from Human Donor Eyes
In This Article
Summary
Identifying proteins specifically associated with Bruch’s membrane in human eyes is an important step in understanding the biochemical mechanisms behind eye diseases such as age-related macular degeneration. This protocol describes how to enrich this sheet of extracellular matrix for down-stream biochemical analysis.
Abstract
Age-related macular degeneration (AMD) is a leading cause of visual impairment in the developed world. The disease manifests itself by the destruction of the center of the retina, called the macula, resulting in the loss of central vision. Early AMD is characterised by the presence of small, yellowish lesions called soft drusen that can progress onto late AMD such as geographic atrophy (dry AMD) or neovascularisation (wet AMD). Although the clinical changes are well described, and the understanding of genetic influences on conferring AMD risk are getting ever more detailed, one area lacking major progress is an understanding of the biochemical consequences of genetic risk. This is partly due to difficulties in understanding the biochemistry of Bruch’s membrane, a very thin extracellular matrix that acts as a biological filter of material from the blood supply and a scaffold on which the retinal pigment epithelial (RPE) cell monolayer resides. Drusen form within Bruch’s membrane and their presence disrupts nutrient flow to the RPE cells. Only by investigating the protein composition of Bruch’s membrane, and indeed how other proteins interact with it, can researchers hope to unravel the biochemical mechanisms underpinning drusen formation, development of AMD and subsequent vision loss. This paper details methodologies for enriching either whole Bruch’s membrane, or just from the macula region, so that it can be used for downstream biochemical analysis, and provide examples of how this is already changing the understanding of Bruch’s membrane biochemistry.
Introduction
The use of post mortem human eye tissue in ophthalmic research is an invaluable resource for the understanding of eye disease pathogenesis. Analysis of human donor eyes has made important contributions to discerning the mechanisms underpinning age-related macular degeneration (AMD), which is the leading cause of blindness in the western world1. Globally, AMD accounts for approximately 8.7% of all blindness and with an increasingly aging population, it is predicted to affect 196 million people by 20202. AMD results in the loss of central vision and has a profound impact on patient independence and quality of life3. Early AMD is characterised by the formation of drusen and it can progress to geographic atrophy (sometimes referred to as ‘dry’ AMD), or choroidal neovascularization (also referred to as neovascular or ‘wet’ AMD). Whilst anti-VEGF injections can stabilise or improve vision in neovascular AMD it is not curative, and at present, no treatment exists for geographic atrophy.
Whilst certain genetic variants have been shown to modify AMD risk 4–8, less is known about how these variants affect the macula at a biochemical level. An important site in AMD pathogenesis is Bruch’s membrane. Drusen form within this structure and various studies have provided evidence of complement activation in and around Bruch’s membrane in AMD9. Bruch’s membrane is a sheet of extracellular matrix that separates the retinal pigment epithelial (RPE) cells from the choroid, and is approximately 4 µm thick. Bruch’s membrane merges into the choriocapillaris, a layer of the choroid that contains fenestrated capillaries and external to this are layers containing larger blood vessels10 (Figure 1A).
Bruch’s membrane is a pentilaminar sheet of extracellular matrix (ECM), and this methodology details how to isolate Bruch’s membrane, along with the ECM of the underlying choriocapillaris (henceforth referred to enriched Bruch’s membrane), which is largely decellularized and free from red blood cells. Enriched Bruch’s membrane was then used for western blotting and histology experiments. Furthermore, methodology for enriching Bruch’s membrane solely from the macula region of the eye is described, which will be of particular interest to those investigating the biochemical aspects of AMD.
Protocol
Human eye tissue consented for research purposes was obtained from the Manchester Royal Eye Hospital Eye Bank following the removal of the cornea for transplantation. The use of human tissue for research was in accordance with the Human Tissue Act (2004) guidelines, and with University Research Ethics Committee approval (reference 11305).
1. Enrichment of Whole Bruch’s Membrane
- Clean the workspace around and underneath the dissecting microscope with 1% disinfectant (v/v) and then wipe down with 70% (v/v) ethanol.
- Ensure the following are to hand: one new disposable scalpel, two sterile cell scrapers (commonly used in tissue culture) and three pairs of tweezers (listed here as A, B, and C, see materials list).
- Place the eye globe into a disposable petri-dish, retaining the lid for later use. Using a new disposable scalpel, remove any residual optic nerve, which may interfere with creating a flat mount of the tissue. Next, make four incisions equally spaced apart to create four quadrants, if proceeding to part 3 (enrichment of macular Bruch’s membrane) do not cut through the macula region.
- Ensure that the incisions extend through the entire depth of the eye tissue – from the innermost neurosensory retina (NSR), through the choroid and sclera (white of the eye). Continue the incision all the way round the eye from the margin of the eyecup to the optic nerve. This will allow the eyecup to be flattened out (see Figure 1B).
- Holding one quadrant of flattened eyecup using tweezer A (to prevent choroid detachment), use the larger tweezer B to grip the vitreous and underlying NSR and pull this tissue away from the retinal pigment epithelium (RPE) starting at the periphery, moving towards the center of the opened eyecup and then to the opposing side. Usually, but not always both vitreous and NSR detach as one. Use the scalpel to excise any residual NSR anchored at the optic disc.
- Again, whilst holding one quadrant of tissue in place using tweezer A, use the sterile cell scraper to gently dislodge the RPE monolayer from the entire inside surface of the eyecup. These cells readily detach and can be seen as dark brown-pigmented clumps. Rinse with PBS to gently wash away dislodged RPE cells from the eyecup.
- Next, use tweezer A to gently peel off the overlying dark red tissue (comprising both Bruch’s membrane and the choroid) from all 4 eye quadrants. Place the excised tissue into a clean dish.
- Using the flattened edge of one cell scraper to hold the excised tissue in place, use the second scraper to gently scrape the tissue. Turn the tissue over and repeat, removing as much excess material as possible. Eventually, a translucent membrane will remain: this is the crudely enriched Bruch’s membrane. The use of the dissecting microscope at a minimum of 20X magnification is essential in aiding the removal of choroid and enrichment of Bruch’s membrane.
- Using a Pasteur pipette, briefly rinse the Bruch’s membrane with ultrapure water to remove residual choroid and blood before placing the membrane into a 1.5 ml microcentrifuge tube.
- Suspend the enriched Bruch’s membrane in approximately 1 ml ultrapure water and vortex briefly (approximately 15 sec) to help lyse any contaminating cellular matter (see Figure 2). Centrifuge the sample at 500 x g for 30 sec to pellet the membrane and aspirate off the supernatant.
Note: The remaining enriched Bruch’s membrane is now ready for solubilization (as described in section 2 below) or storage at -80 °C for use later.
2. Solubilization of Whole Enriched Bruch’s Membrane and Western Blot Analysis
- Extract protein from enriched Bruch’s membrane by immersing the tissue in 500 µl 8 M urea for 6 hr at RT in 1.5 ml microcentrifuge tubes ( Figure 3A).
- Centrifuge solubilized samples at 10,000 x g for 10 min to pellet any remaining extracellular matter. Remove the supernatant and transfer it to 3,500 Da MWCO dialysis membrane, then place the sample in 1 L of PBS to dialyse for 16 hr at 4 °C.
- For each sample, add 40 µl of protein isolation beads, and rotate sample/bead mix (20 rpm on a rotating mixer) at RT for 30 min.
- Centrifuge samples at 10,000 x g for 5 min to pellet the beads. Remove supernatant and solubilize protein bound to beads in 30 µl 2x sample loading buffer (65.8 mM Tris-HCl, pH 6.8, 2.1% SDS, 26.3% (w/v) glycerol, 0.01% bromophenol blue, 3% β-mercaptoethanol).
- Incubate the samples at 100 °C for 10 min, and then centrifuge at 10,000 x g for 5 min to pellet the remaining beads.
- Load the supernatant that contains the released proteins on to a pre-cast SDS-page gradient gel (see materials list) for protein separation.
- Visualize separated protein bands using a common staining protocol, e.g., Coomassie blue.
- In brief, add 20 ml of the brilliant blue stain (0.1% brilliant blue (w/v), 50% methanol, 10% glacial acetic acid, 40% water) to the gel, and incubate for 30 min at RT with gentle agitation.
- Discard the staining solution and wash the stained gel with deionized water until background staining is sufficiently reduced to allow stained protein band visualisation.
- Alternatively, transfer separated protein bands to nitrocellulose for western analysis, as described below.
- Transfer separated protein bands onto nitrocellulose membrane at 80 mA for 2.5 hr using semidry transfer apparatus in transfer buffer (25 mM Tris, 192 mM glycine, 10% [v/v] methanol).
- Block the membrane in 10 ml PBS, 10% (w/v) milk, and 0.02% (w/v) BSA for 16 hr at 4 °C before the addition of the desired primary antibodies for protein detection.
Note: As an example here, Figure 3C shows a representative immunoblot of solubilised Bruch’s membrane from three separate donors probed for a regulator of the complement cascade, factor H-like protein 1 (FHL-1).
3. Enrichment of Macular Bruch’s Membrane
- Follow steps 1.1 - 1.3 as previously described.
- Whilst the NSR is still in place, locate the yellow foveal colouration that represents the central macula region (see Figure 4). Using a sterile 6 mm biopsy punch, place it above the foveal region and firmly cut in to the macula.
- Keeping the biopsy punch in place, make a separate incision with a scalpel from the edge of the eyecup to the edge of biopsy punch blade, and then using tweezer B, peel away the surrounding eye material to achieve a clean biopsy.
- Using tweezer C, remove the 6 mm disc of tissue from the punch and place on the clean petri dish lid. Place the petri dish lid and macular punch under the dissecting microscope.
- Remove the thin NSR disc (check for yellow foveal staining to infer accuracy of biopsy). Use a sterile cell scraper to gently remove the RPE cells.
- Secure the macular disc via the sclera using the tips of tweezer A, and slowly peel off the choroid and Bruch’s membrane complex using tweezer C. Again, use the cell scraper to remove unwanted choroid and blood matter from Bruch’s membrane.
- Place the macular Bruch’s membrane into a 1.5 ml microcentrifuge tube and suspend in 1 ml ultrapure water. Vortex briefly (approximately 15 sec) to help lyse any contaminating cellular matter. Centrifuge the sample at 500 x g for 30 sec to pellet the membrane and aspirate off the supernatant.
Note: The enriched Bruch’s membrane isolated from the macula can now be used for proteomic analysis using the user’s preferred digestion protocols.
Results
The protocol described above to produce enriched Bruch’s membrane results in the isolation of Bruch’s membrane, the ECM of the choriocapillaris and removes most of the cells/cellular debris including all of the RPE (Figure 2). The washing of the enriched Bruch’s membrane in water is a key step in lysing any remaining choroidal cells. The pressure exerted during scrapping of the choroid appears to compact some of the few remaining cell nuclei into the region defined as Bruch’s membrane (Figure 2B).
The solubilisation of enriched Bruch’s membrane allows the downstream analysis of its protein content. The separation of protein bands from solubilised enriched Bruch’s membrane using a 4 - 12% gradient SDS-PAGE gel, while not useful in its own right to identify protein content, does demonstrate the ability of this technique to minimise contaminating blood proteins that are non-specifically bound (Figure 3B). In this regard, one notable absentee from the protein gel run is a sizeable band of human serum albumin (~66 kDa) that may otherwise make subsequent western blotting more difficult to interpret: the histological analysis of enriched Bruch’s membrane shown in Figure 2 also supports this. Indeed, as part of work described previously11, solubilised Bruch’s membrane from a series of individual donors were probed for a number of proteins using specific antibodies via western blotting (Figure 3C). In the case shown here, an antibody (termed OX23)12 directed against a 155 kDa regulator of innate immunity, called complement factor H (FH), was used and demonstrated the presence of a lower molecular weight band in Bruch’s membrane samples from three separate donors (Figure 3C). This subsequently turned out to be factor H-like protein 1 (FHL-1), a 49 kDa protein related to FH and arising from a splice variation of the same gene13. FHL-1 appears to be the main complement regulator present in Bruch’s membrane11.

Figure 1. Dissection of the Human Eye and Enrichment of Bruch’s Membrane. (A) Bruch’s membrane is an extracellular matrix barrier. The choroid contains a layer of fenestrated capillaries that continuously merge into Bruch’s membrane (called the choriocapillaris) and these separate the rest of the choroid (containing large blood vessels) from the retinal pigment epithelium (RPE) cell monolayer, which physiologically supports the photoreceptor cells of the neurosensory retina (NSR). (B) Four incisions into the human eyecup are made allowing it to be opened to create a flat mount. The vitreous can be removed with the use of tweezers. The NSR may come off along with the vitreous but can also be removed with tweezers. A cell scraper is used to remove the RPE cell layer from Bruch’s membrane while it is still anchored to the choroid and sclera. The Bruch’s membrane/choroid complex can be peeled away from the sclera and using a new cell scraper, the outer choroid can be removed. A final wash in ultrapure water lyses any remaining cellular matter. Please click here to view a larger version of this figure.
{kind=link}

Figure 2. Histology of Bruch’s Membrane and the Choroid. H&E stained sections before (A) and after (B) scraping away the outer choroidal structures from Bruch’s membrane and washing in ultrapure water to generate enriched Bruch’s membrane. Scale bars represent 20 µm. Please click here to view a larger version of this figure.
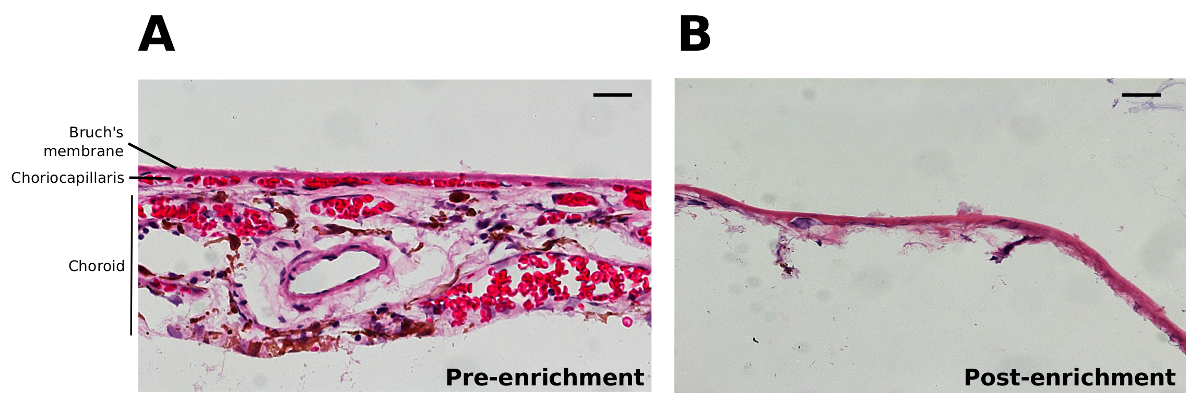{kind=link}

Figure 3. Solubilisation of Enriched Bruch’s Membrane for Protein Content Analysis. (A) Protein can be extracted from enriched Bruch’s membrane by incubating in 8 M urea for 6 hr at RT, before being dialysed against PBS. For Western analysis, proteins from the whole Bruch’s membrane dialysate can be isolated using protein isolation beads and eluted by boiling with laemmli reducing sample loading buffer. (B) A representative Coomassie blue stained SDS-PAGE gel, showing both neat and diluted solubilised Bruch’s membrane (BM) using the method described in (A). (C) A representative western blot of solubilised Bruch’s membrane from 3 individual donors (lanes 1 - 3) probed for factor H-like protein 1 (FHL-1). Please click here to view a larger version of this figure.
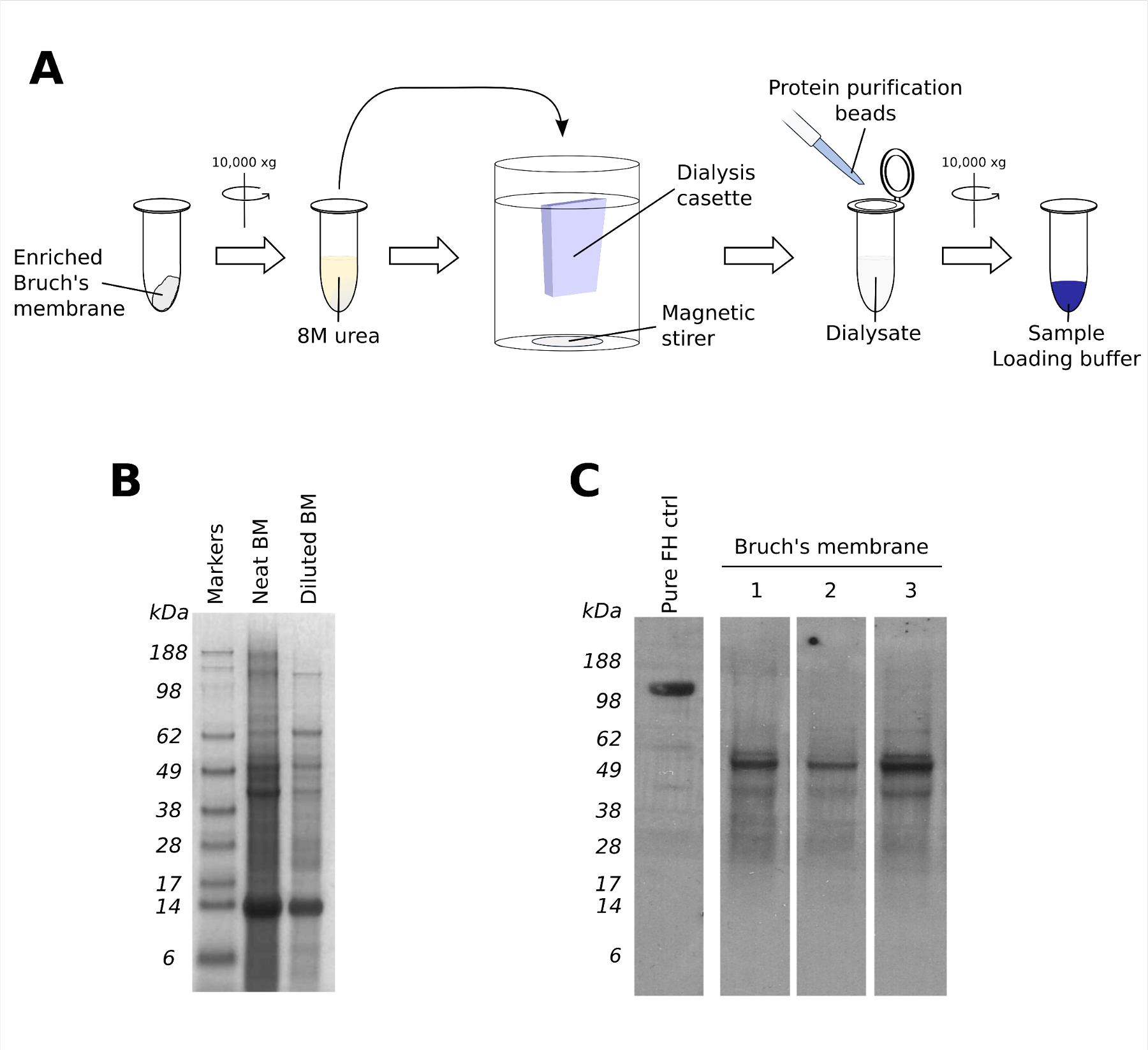{kind=link}

Figure 4. Isolation of Enriched Bruch’s Membrane from the Macula. The macula region of the retina is a 5 mm area that can be readily identified by the yellow colouration of the overlying fovea, and its proximity to the adjacent optic disc. Using the fovea as a guide, place a 6 mm biopsy punch on the flat mounted human eye to excise a 6 mm macula tissue block. Remove the overlying NSR and scrape off RPE cells from the excised macula. The remaining Bruch’s membrane/choroid complex can then be peeled away from the sclera and thoroughly scraped to remove choroid, lyse remaining cellular material by submersion in ultrapure water. Please click here to view a larger version of this figure.
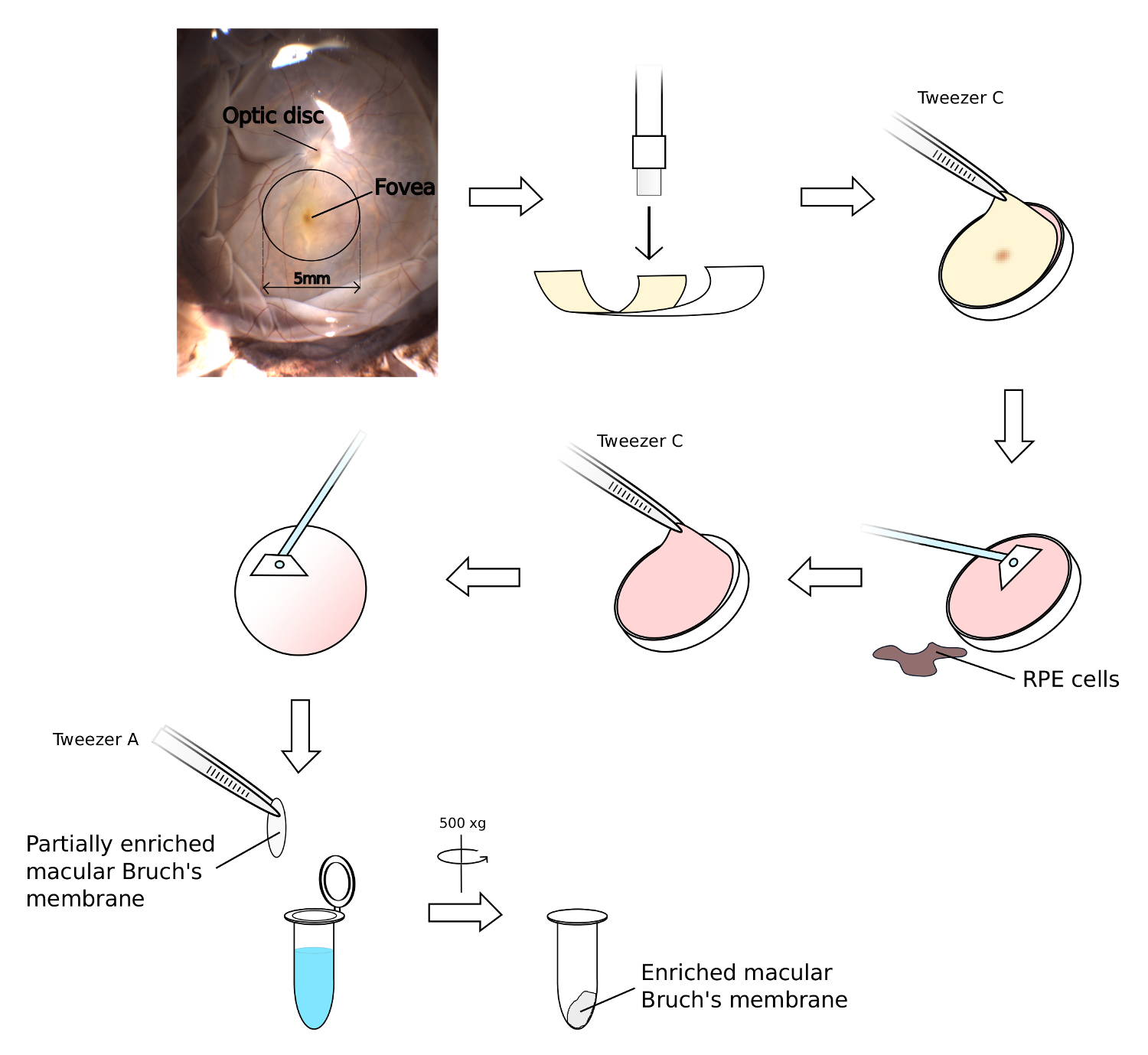{kind=link}
Discussion
Here we describe the enrichment of Bruch’s membrane for further analysis allowing subsequent analyses including western blotting11, mass spectrometry and the diffusion properties of Bruch’s membrane utilising modified Ussing chambers11,14. Critical steps include a) the thorough scraping and washing of the inner surface of the eye to remove all RPE cells b) the repeated and careful scraping of the underlying choroid without causing the Bruch’s to disintegrate and c) washing of the excised Bruch’s in ultrapure water to ensure lysis of residual cells. Furthermore, if the enriched Bruch’s membrane is to be utilized for proteomic analysis, protein extraction by urea treatment is the most effective in breaking down this resilient tissue as compared to other methods such as mechanical homogenisation. Histology of enriched Bruch’s membrane indicates the ECM of the choriocapillaris is not removed. This is to be expected as the outer layer of the pentilaminar Bruch’s membrane is formed by choriocapillaris endothelial cell basement membranes. Indeed it may be advantageous that the ECM of the choriocapillaris remains as changes occur in this in AMD including the deposition of terminal complement complex/membrane attack complex (MAC)15. It should be noted that for detailed histological analysis of Bruch’s membrane, its structure would be better preserved if sectioned in the presence of sclera, choroid, and Bruch’s membrane. Indeed, the histology presented in Figure 2 is solely intended to demonstrate the degree of enrichment resulting from the technique described in this manuscript.
Enrichment of macular Bruch’s requires careful dissection of the eyecup, ensuring that the NSR and particularly the fovea are not damaged; otherwise identification and thus isolation of the macula are not possible. The integrity of the tissue is also important to this process and it is therefore recommended that samples under 48 hr post-mortem time be used. The degree of enrichment required varies between donors and can be affected by donor age, post-mortem time, and the presence of any eye pathology. The use of the term enriched is deliberate in acknowledging a limitation of the technique, this being that ‘purification’ or ‘isolation’ of Bruch’s membrane using this method simply is not possible. Indeed, given that the ECM of the choriocapillaris merges into Bruch’s membrane there complete separation is not feasible.
Understanding the biochemical changes of the respective ocular tissues is essential in understanding eye disease progression. This unique enrichment of Bruch’s membrane facilitates this understanding; ongoing studies are investigating the proteome of enriched macular Bruch’s membrane using mass spectrometry at different stages of AMD and in samples originating from subjects with different genotypes to improve understanding of the molecular pathology of AMD. Already the technique described here has been used to successfully identify FHL-1 as the predominant complement regulator in Bruch’s membrane 11 and further studies on enriched Bruch’s membrane are likely to result in more new insights into the molecular pathology of AMD.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgements
The Authors would like to acknowledge Dr. Isaac Zambrano and the staff at the Manchester Eye Hospital Eye Bank for the supply of human donor eye tissue, and Mr. Pete Walker of the Faculty of Life Sciences Histology facility for the H&E stained images. Special thanks goes to Mr. Roger Meadows for his help with the microscopy. SJC is a recipient of a Medical Research Council (MRC) Career Development Fellowship (MR/K024418/1) and the authors also acknowledge other recent research funding from MRC (G0900538 and K004441), Fight for Sight (1866) and The Macular Society.
Materials
| Name | Company | Catalog Number | Comments |
| Auxillary objective 0.5x | GT-Vision Ltd | 3638 | Useful (but not essential) for lower magnification imaging of whole eye flat-mount |
| Biopsy Punch 6mm | Oncall Medical Supplies | SCH-33-36 | 6mm Biopsy Punches |
| Bovine serum albumin | Sigma - Aldrich | A3059 | |
| Bromophenol Blue | Sigma - Aldrich | B0126 | |
| Cell scrapers | Sarstedt | 83.183 | For membrane enrichment |
| CyroPure Cyrovials | Sarstedt | 72.38 | |
| Disposable Scalpels | Fisher Scientific | 12387999 | Sterile, individually wrapped Swann-Morton scalpels. |
| Dual Gooseneck LED spot lights on 30cm arms | GT-Vision Ltd | 0153 | Flexible light source essential for dissection and imaging |
| Dulbecco’s Phosphate Buffered Saline | Sigma - Aldrich | D8537 | Used to prevent macula drying out and during membrane enrichment. |
| (A) Forceps | Scientific Laboratory Supplies | INS4280 | (A) Fine (100mm) and straight |
| (B) Forceps | Scientific Laboratory Supplies | INS4272 | (B) Broad and blunt (150mm). Useful for lifting large portions of sample |
| (C) Forceps | Scientific Laboratory Supplies | INS4323 | (C) Straight fine points |
| Glycerol | Sigma - Aldrich | G1345 | |
| Glycine | Sigma - Aldrich | B0126 | |
| GXCAM 5 digital colour camera & GT-Vision software | GT-Vision Ltd | 1122 | |
| Methanol | Sigma - Aldrich | M/4000/17 | |
| Nitrocellulose membrane | Life Technologies | NP0007 | |
| NuPAGE Novex 4-12% Bis-Tris Protein Gels | Life Technologies | NP0322BOX | |
| Petri dish | Sterilin | 101VR20 | 90mm, used as a dish for samples |
| Protein isolation beads (Strataclean resin) | Agilent | 400714-61 | |
| SDS | Bio-Rad | 161-0301 | |
| Stereozoom microscope GXMXTL3T (magnification range 7x-45x) | GT-Vision Ltd | 5595 | Stereozoom microscope attached to boom stand for ease of use during dissection |
| Tris-HCl | Sigma - Aldrich | T3253 |
References
- Coleman, H. R., Chan, C. -. C., Ferris, F. L., Chew, E. Y. Age-related macular degeneration. Lancet. 372, 1835-1845 (2008).
- Wong, W. L., et al. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob Health. 2 (2), e106-e116 (2014).
- Coleman, A. L., et al. Impact of age-related macular degeneration on vision-specific quality of life: Follow-up from the 10-year and 15-year visits of the study of osteoporotic fractures. Am J Ophthalmol. 150 (5), 683-691 (2010).
- Hageman, G. S., et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc Natl Acad Sci U S A. 102 (20), 7227-7232 (2005).
- Klein, R. J., et al. Complement factor H polymorphism in age-related macular degeneration. Science. 308 (5720), 385-389 (2005).
- Haines, J. L., et al. Complement factor H variant increases the risk of age-related macular degeneration. Science. 308 (5720), 419-421 (2005).
- Edwards, A. O., Ritter, R., Abel, K. J., Manning, A., Panhuysen, C., Farrer, L. A. Complement factor H polymorphism and age-related macular degeneration. Science. 308 (5720), 421-424 (2005).
- Fritsche, L. G., et al. Seven new loci associated with age-related macular degeneration. Nat Genet. 45 (4), 433-439 (2013).
- Clark, S., Bishop, P. Role of Factor H and Related Proteins in Regulating Complement Activation in the Macula, and Relevance to Age-Related Macular Degeneration. J Clin Med. 4 (1), 18-31 (2014).
- Booij, J. C., Baas, D. C., Beisekeeva, J., Gorgels, T. G. M. F., Bergen, A. A. B. The dynamic nature of Bruch’s membrane. Prog Retin Eye Res. 29 (1), 1-18 (2010).
- Clark, S. J., et al. Identification of factor H-like protein 1 as the predominant complement regulator in Bruch’s membrane: implications for age-related macular degeneration. J Immunol. 193 (10), 4962-4970 (2014).
- Sim, E., Palmer, M. S., Puklavec, M., Sim, R. B. Monoclonal antibodies against the complement control protein factor H (beta 1 H). Biosci Rep. 3 (12), 1119-1131 (1983).
- Ripoche, J., Day, A. J., Harris, T. J., Sim, R. B. The complete amino acid sequence of human complement factor H. Biochem J. 249 (2), 593-602 (1988).
- Moore, D. J., Clover, G. M. The effect of age on the macromolecular permeability of human Bruch’s membrane. Invest Ophthalmol Vis Sci. I. 42 (12), 2970-2975 (2001).
- McHarg, S., Clark, S. J., Day, A. J., Bishop, P. N. Age-related macular degeneration and the role of the complement system. Mol Immunol. 67 (1), 43-50 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved