
Method Article
Yalıtım ve floresan görüntüleme Chlamydomonas Centrioles tek-parçacık yeniden inşası için
* Bu yazarlar eşit katkıda bulunmuştur
Bu Makalede
Özet
Arındırmak ve centrioles farklı yönelimleri Süper çözünürlük mikroskobu ve tek-parçacık ortalama için mükellef içinde çok sayıda resim için bir strateji geliştirdik.
Özet
Centrioles büyük makromoleküllerin grupları temel hücre hücre bölünmesi, hücre hareketliliği veya hücre sinyallemesi biyolojik işlemler düzgün yürütülmesi için önemli olan. Yeşil algler Chlamydomonas reinhardtii çalışmaya centriole mimari, işlev ve protein kompozisyon anlayışlı bir model olduğu ispatlanmıştır. Anlayış centriolar mimari doğru büyük gelişmeler rağmen mevcut zorluklardan biri içindeki rolü daha iyi anlamak için centriolar bileşenleri yapısal centriole bölgelerinde içinde kesin lokalizasyonu tespit etmektir Centriole Biyogenez. Büyük bir sınırlama bu organel kırınım sınıra yakın boyutları ile protein yerelleşme yorumlanması için sadelik floresans mikroskobu ile çözünürlükte yatıyor. Bu soruyu çözmek için biz bir yöntem arındırmak ve C. reinhardtii centrioles, çok sayıda farklı yönelimleri Süper çözünürlük mikroskobu kullanarak görüntü sağlıyor. Bu tekniği daha da floresan tek-parçacık (Fluo-SPA) satın centrioles çok sayıda nedeniyle ortalama ile veri işleme sağlar. Fluo-SPA oluşturur ortalamalar, C. reinhardtii centrioles farklı yönleri, böylece centriolar alt bölgelerinde farklı proteinler lokalizasyonu kolaylaştırmak içinde lekeli. Önemlisi, bu yöntem diğer türler veya diğer büyük makromoleküllerin grupları görüntü centrioles için uygulanabilir.
Giriş
Centriole centrosome hayvan hücrelerinde çekirdek yer alır ve bir bazal vücut (centrioles bundan sonra adlandırılır) şablonu kirpikler veya kamçı birçok Ökaryotlar1,2olarak hareket edebilir bir evrimsel korunmuş organel olduğunu. Bu nedenle, centrioles sinyal hücre için iğ derlemesinden değişen temel hücre biyolojik işlemler için kritik öneme sahiptir. Bu nedenle, centriole derleme ya da işlev3ciliopathies ve kanser de dahil olmak üzere birçok insan patolojileri ile ilişkili bulunmuştur.
Centrioles nine-fold, simetrik, Mikrotubul üçlüsü tabanlı silindirik yapılar vardır, genellikle, ~ 450 nm uzun ve ~ 250 nm geniş4,5,6,7. Geleneksel elektron mikroskobu ve cryo-elektron tomografi, centrioles farklı türlerden olduğunu açıkladın centrioles üç farklı bölgeleri ile onların uzun ekseni boyunca polarize: proksimal bölge, merkezde bir çekirdek ve distal bölge5 , 7 , 8 , 9 , 10 , 11. önemlisi, her bu bölgede belirli yapısal özelliklerini görüntüler. İlk olarak, 100 mil uzunluğundaki proksimal bölge lümen den iğne başı öğe12Mikrotubul üçlüsü bağlı çember hareketi yapısını içerir. İkincisi, 300-400 nm-uzun iç bölgesi fibröz yoğunlukları mikrotübüller iç yüzü boyunca lümen ve yapısal özellikler içerir: Y-şekilli bağlayıcı, C-tübül kuyruğu ve A-tübül9saplama. Son olarak, 50-100 nm distal bölge distal centriole5,13bölümünü surround alt distal ve distal ekleri sergiler.
Son yirmi yılda yaklaşık 100 farklı proteinler centriole14,15,16, bir parçası olmak bir geçerli tahmini için önde gelen centriolar proteinler giderek artan sayıda keşfi tarafından işaretlenmiş 17. bu gelişmeler rağmen bu proteinler centriole içinde kesin lokalizasyonu zor, özellikle yapısal alt bölge içinde kalır. Önemlisi, hassas bir yerelleştirme centriole yapısal bölgelerine atama işlevlerine daha iyi anlamak için çok önemlidir. Bu bağlamda, C. reinhardtii centrioles ilk sonra verilen silindir9,18,19, boyunca farklı yapısal özellikler delimitating tarafından her iki yönden vesile olmuştur Araştırmacılar bir alt alt yapısal bir bölgesine floresan mikroskopisi kullanılarak proteinlerin lokalizasyonu aralarındaki ilişkileri belirlemektir. Bu, örneğin, proteinler Bld12p ve Bld10p, proksimal bölge ve çember hareketi yapısı özellikle yerelleştirmek20,21,22,23içerir. Yerelleştirilmiş altyapı proteinler listesi de POB15 ve POC16, C. reinhardtii centrioles17iç Merkezi çekirdek bölgenin süslemeleri kütle spektrometresi tarafından tanımlanan iki roman protein içerir.
Bu kağıt yalıtmak ve C. reinhardtii centrioles sonraki süper çözünürlük mikroskobu ve tek-parçacık ortalama için görüntü için geliştirilen Yöntem tam bir açıklamasını sağlar. Bu hedefe ulaşmak için aşılması gereken teknik sınırlamalar betimlemek önemlidir. İlk olarak, centriole arıtma kez yalıtım9çeşitli adımlar sırasında kaybetmiş olmanın çember hareketi yapısı ile genel mimarisi etkileyebilir. İkinci olarak, centriole boyutlarının çok optik mikroskobu kırınım sınıra yakın olduğundan. Gerçekten de, confocal mikroskobu elde edilebilir yanal çözünürlük yaklaşık 200 nm24, centriole ve zçözünürlüğünde çapı benzer olduğunu-eksendir hakkında 2-3 x daha düşük, anisotropic bir birime lider. Üçüncü olarak, antikor etiketleme ve centriole yönlendirme heterojen bir protein belirli bir bölgesinde centriolar alt yerelleştirmek için gerekli yorumu sınırı olabilir. Son olarak, çok sayıda resim elde etmek ve bir benzersiz centriole yön bulmak üzere hücre başına sadece iki kopya centrioles bulunmaktadır. Bu teknik sorunları aşmak için Süper çözünürlük mikroskobu çeşitli yönleri kabul izole centrioles çok sayıda uygulama üzerinde dayanır bir yöntem geliştirdi. Önce yapısal olarak sağlam centrioles ve çember hareketi içeren procentrioles arınma sağlayan C. reinhardtii centrioles arındırmak için bir protokol anlatacağım. Sonra centrioles coverslips tarafından geleneksel görüntüleme veya Süper çözünürlük floresan mikroskopi için konsantre için adım adım bir protokol anlatacağız. Bu önemli bir adım centrioles içinde birden fazla yönelimleri yansıma sayısını artırmak için sağlar. Son olarak, tek-parçacık farklı yönelimleri centrioles algılama kolaylaştırır floresan mikroskoplar alınan veriler üzerinde ortalama gerçekleştirmek için bir prosedür anlatacağım. Özet olarak, bu yöntem görüntü centrioles için çeşitli türler veya diğer büyük makromoleküllerin grupları uygulanabilir.
Protokol
1. medya hazırlık C. reinhardtii kültür ve Centriole yalıtım için
-
Medya kültür C. reinhardtii hücrelere hazırlanması
Not: 1 x dokunun (Tris-asetat fosfat) orta hisse senedi çözümleri hazırlanması aşağıdaki adımları açıklanmaktadır.- Bir fosfat tampon (pH 7), ~ 170 mL / 1 M KH2PO4 (KH2PO4 ' te 1 L 136.09 g) ile 250 mL 1 M K2HPO4 (K2HPO4 için 1 L distile su ile tamamlanmaktadır 174.2 g) karıştırarak hazırlayın. PH 7 ulaşmak için karışım ayarlayın.
- 96.8 g Tris, fosfat tampon (pH 7) 40 mL ve 40 mL asetik asit karışımı ile çözüm (40 x) hazırlamak ve 1 L için çözüm distile su ile ayarlayın.
- Çözüm B hazırlamak (40 x) kullanarak 16 g NH4Cl, CaCl22 g ve 4 g MgSO4. Distile su içinde CaCl2 ayrı olarak diğer bileşenleri eklemeden önce dağıtılması için dikkatli olun. 1 L için çözüm distile su ile ayarlayın.
- Hutner'ın eser elementler arabellek25 aşağıdaki gibi hazırlayın.
- Arabellek 1 litre su Belirtilen birimdeki her bileşik dağıtılması: EDTA disodyum tuz 250 ml (50 g), ZnSO4.7H2O (100 ml 22 g), H3BO3 (200 mL 11,4 g), MnCl2.4H2O (50 mL 5.06 g) , CoCl2.6H2O (50 mL 1.61 g), CuSO4.5h2O (50 mL 1.57 g), (NH4) 6Mo7O24.4H2O (1,10 g 50 mL) ve FeSO4.7H2O (50 mL 4,99 g).
Not: EDTA kaynar suda çözünmüş ve FeSO4 son oksidasyonu önlemek için hazır olmalıdır. - EDTA hariç tüm çözümler karıştırın ve kaynatın getirmek. EDTA ekleyin ve çözüm yeşil açmak. Her şey sonra eriterek, serin çözüm ile 70 ° c Bu sıcaklıkta 85 mL sıcak % 20 KOH çözeltisi (100 ml 20 g) ekleyin. 1 L için çözüm, oda sıcaklığında (RT) distile su ile ayarlayın.
- Pamuk tak balon için ekleyin ve 1-2 hafta için sallıyorum ayakta çözüm izin 1 x bir gün. Çözüm başlangıçta yeşil ve mor, pas-kahverengi küçülmesinihızlandırmalı bırakarak açmak; çökelti kadar çözüm açıktır filtre kağıdı kullanarak kaldırın. Aliquots donma ve -20 ° C'de depolayın
- Arabellek 1 litre su Belirtilen birimdeki her bileşik dağıtılması: EDTA disodyum tuz 250 ml (50 g), ZnSO4.7H2O (100 ml 22 g), H3BO3 (200 mL 11,4 g), MnCl2.4H2O (50 mL 5.06 g) , CoCl2.6H2O (50 mL 1.61 g), CuSO4.5h2O (50 mL 1.57 g), (NH4) 6Mo7O24.4H2O (1,10 g 50 mL) ve FeSO4.7H2O (50 mL 4,99 g).
- 1 x dokunun orta aşağıdaki bileşenleri karıştırılarak C. reinhardtii hücreleri büyümek için hazırlamak: 25 mL çözeltisi (40 x) ve 25 mL çözeltisi B (40 x) ile Hutner'ın iz elementler 1 mL25arabellek ve 1 L karışıma distile su ile ayarlayın. 0.4 µm filtre kullanarak karışımı sterilize.
-
Centriole arıtma medyası hazırlama
- %5 sukroz 500 mL distile su ile düzeltilmiş son hacmi 10 mM HEPES (pH 7) kullanarak bir deflagellation arabellek hazırlayın.
- 250 mL 0.5 M asetik asit hazırlayın.
- 250 mL bir 1 M K-borular stok çözeltisi (pH 7.2) borular toz çözülmeye H2O ilk ekleyerek 50 mL tarafından hazırlamak, sonra 10 ekleme çözüm kadar N KOH açık olmaya başlar. PH 7.2 10 N ve 1 N KOH ile titre ve 250 mL H2O. son hacmi için çözüm ayarlayın
- 5 sükroz çözümleri (w/w) aşağıdaki gibi hazırlayın.
Not: Tüm sükroz çözümler 0.4 µm filtre kullanarak solubilization bir şırınga takılı sonra filtre gerekir. %60 ve % 70 sükroz çözümleri solubilize zordur ve 60 ° C solubilization kolaylaştırmak için önceden sıcak su banyosu yerleştirilmelidir unutmayın. Her 10 min sükroz tamamen eriyene kadar karıştırın.- % 25 sükroz hazırlamak, sukroz 25 g tartmak ve 10 mM K-borular (pH 7.2) ekleyerek 100 gr ağırlık ayarlamak için.
- % 60 sükroz hazırlamak, sukroz 60 g tartmak ve 10 mM K-borular (pH 7.2) ile 100 gr ağırlık ayarlamak için.
- Sükroz çözümleri sükroz degrade için hazırlayın. Sükroz 40 g ağırlığında ve 100 g 10 mM K-borular (pH 7.2) ile çözüm ayarlayarak % 40 sükroz hazırlayın. Benzer şekilde, (w/w) % 50 ve % 70 (w/w) sükroz hazırlayın.
- Çözümler-20 ° C'de depolayın Sükroz çözümleri düzgün çözdürme sonra resuspend için dikkatli olun.
- 100 mL lizis arabellek karıştırma 1 mM HEPES (pH 7) tarafından 0,5 mM MgCl2ve %1 NP-40 ve tutmak hazırlamak Bu 4 ° C'de Her zaman bu arabellek taze deneme günü hazır olun. Anti-proteaz tablet centriole yalıtım gününde ekleyin.
- NaCl 8 g, 2 g KCl, Na2HPO41,44 g ve KH2PO4 800 ml distile H2O. ayarlama pH 7.4 için 0,24 g 1 karışıma hacmi HCl. Bring ile karıştırılarak 1 fosfat tampon serum (PBS) (pH 7,4) x hazırlamak L distile H2O.
2. C. reinhardtii Centrioles yalıtım
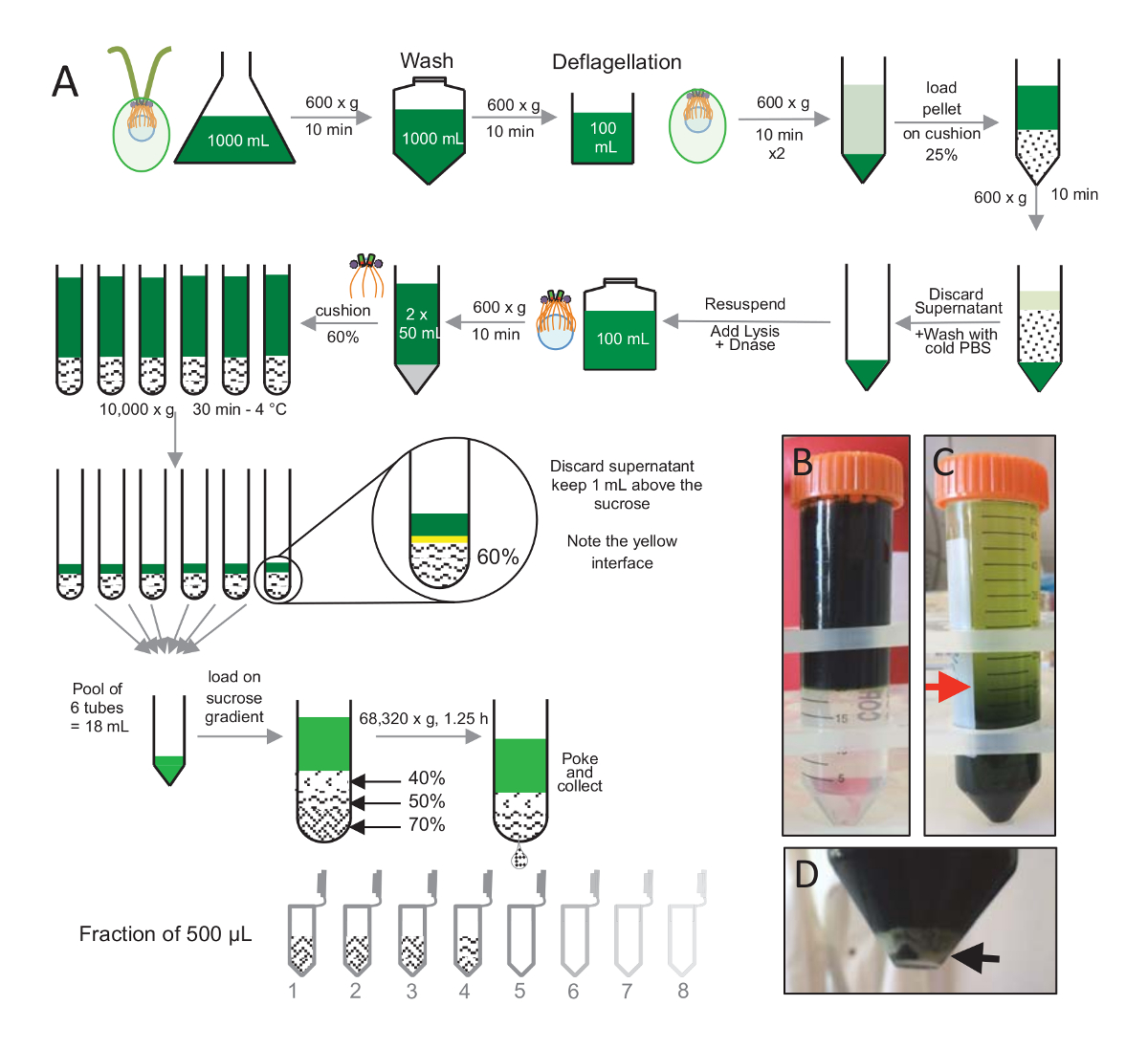
Not: Bkz. Şekil 1.
-
Kültür ve C. reinhardtii hücre genişletme
- 1 gün akşamı, bir cw15 - yük bir katı plaka bir kültür Erlenmeyer şişesi 1 10 mL içeren aşılamak x dokunun. 23 ° C'de 2-3 gün boyunca beyaz floresan ışıklar (60 µE/m2s) altında hücrelerin büyümesine
- 3 günde 10 kültür seyreltik (100 mL için) x 1 x dokunun ve 23 ° C'de 2-3 gün boyunca ışık altında hücrelerin büyümesine
- 6 günde 10 kültür seyreltik x 1 x 1 L kültür elde etmek için dokunun. Kültür ~ 1 x 107 hücre/mL26 (gün 9 – 10) yaklaşık hücre yoğunluğu gösterir koyu yeşil renk ulaşıncaya kadar ışık 23 ° c altında hücrelerin büyümesine.
-
C. reinhardtii centrioles arıtma
- Hücreleri cw15 - 600 x g 10 dk 50 mL konik tüpler santrifüj kapasitesi. Pelet, yıkama 1 x 50 mL 1 x PBS ile hücreleri ve Pelet oda sıcaklığında deflagellation arabelleği bir pipet ile 100 ml 600 x g 10 dk. Resuspend için de sıralayacağız.
- Yavaş yavaş son pH 4,5-4,7 bir manyetik karıştırıcı tarih değeri 0,5 M asetik asit damla ekleyerek bir pH şok hücrelerle deflagellate ve yavaş yavaş 2 dk. eklemek için pH 7,0 için geri yüklemek için 1 N KOH damlaları hücreleri kuluçkaya.
- Hücreleri, 600 x g müstakil herhangi bir kamçı kaldırmak 10 dk santrifüj kapasitesi. Süpernatant çıkarın ve Pelet buza saklayın. Pelet 2 yıkama x 50 mL 1 x PBS 4 ° C'de precooled ile O zaman, Pelet 600 x g 4 ° C'de 10 dakika için de spin
- Pelet 30 mL 1 x PBS resuspend ve üzerine % 25-sükroz minder 20 ml Süspansiyon (Şekil 1B) karıştırma olmadan yük yavaş.
- 600 x g 15 dakika kalan kamçı süpernatant içinde kaldırmak için 4 ° c de spin; hücreleri % 25 sukroz (Şekil 1C) yayılır. Sadece en alt 20 mL (kırmızı ok, Şekil 1C) bir aspiratör kullanarak süpernatant aspirating tarafından dikkatle tutun.
- Kalan 20 mL 20 mL soğuk 1 x PBS ekleyerek yıkayın. 600 x g 4 ° C'de 10 dk da örneğine spin Pelet soğuk 1 x PBS (4 ° C'de) 10 ml resuspend. Böylece aşağıdaki lizis tüm hücreleri aynı anda hits hiçbir kümeleri vardır emin olun.
- Resuspended Pelet yeni bir 250 mL şişe aktarın. DNaz 5.000 adet hücreleri ile takıma lizis arabellek 100 mL ekleyin. Hücreleri ve başka bir yol lizis arabellek eklemek önemlidir. Karışımı 4 ° C'de 1 h için kuluçkaya ve şişe her 15dk herhangi bir baloncuklar kurma olmadan ters çevirme tarafından dikkatlice karıştırın.
- Lysed hücreleri, 600 x g herhangi bir hücre artıkları kaldırmak için bir 50 mL konik tüp 4 ° C'de 10 dakika santrifüj kapasitesi. Lizis doğru sahne aldı, hücre Pelet beyaz olmalıdır (Şekil 1D). Bir pipet ile süpernatant toplamak ve buz üzerinde yerleştirilen bir % 60-sükroz yastık içeren bir 30 mL yuvarlak alt tüp üzerine yükleyin. O zaman, tüp 10.000 x g 4 ° C'de 30 dk için de spin
Not: Birkaç tur-alt tüpler (Tablo malzemeler), süpernatant hacmine bağlı olarak gerekli olabilir. - 1 mL sukroz yastık yukarıda süpernatant ilâ Aspire edin. Arasında kalan süpernatant 1 mL ve sukroz yastık 2 mL sarı arabirim unutmayın. Sukroz ve kalan süpernatant ile kesme P1000 bahşiş karışımı yavaşça. Girdap bu adımda değil mi; Aksi takdirde, procentrioles bu aşamada kayıp olabilir. Tüm sükroz yastıkları havuz ve buza depolayabilirsiniz.
- %40 - %70 sükroz Gradyanda 38,5 mL ince duvarlı polipropilen boru yavaşça ardından 3 mL % 50 ve son olarak, 3 mL % 40 sükroz % 70 sukroz (4 ° C'de), 3 mL ekleyerek hazırlayın. Havuza alınan arabirimleri %40 - %70 sükroz degrade üzerine yük; soğuk sükroz çok viskoz olduğu için yavaş yavaş bunu. 10 mM K-borular arabellek (pH 7.2) tüplerini denge ve onları 68,320 x g (Örneğin, bir SW32Ti rotor uçağı ile) için 1 saat 15 dk 4 ° C'de, santrifüj kapasitesi
- 12 x 500 µL kesirler 4 ° C'de bir delik tüpün dibine 0.8 mm iğne ile farklı sükroz katmanları bozmadan yaparak toplamak. Bir kesim P200 pipet ucu ile aşağıdaki ayirt için kullanılacak olan her kesir üzerinden ek 10 µL aliquots hazırlayın. Ek-kesirler sıvı azot kıpırdama.
Not: İzole centrioles elektron mikroskobu tarafından centrioles genel ultrastructure korunur emin olmak için analiz edilebilir.
3. Coverslips Tarih izole Centrioles miktar: Santrifüjü ve ayirt
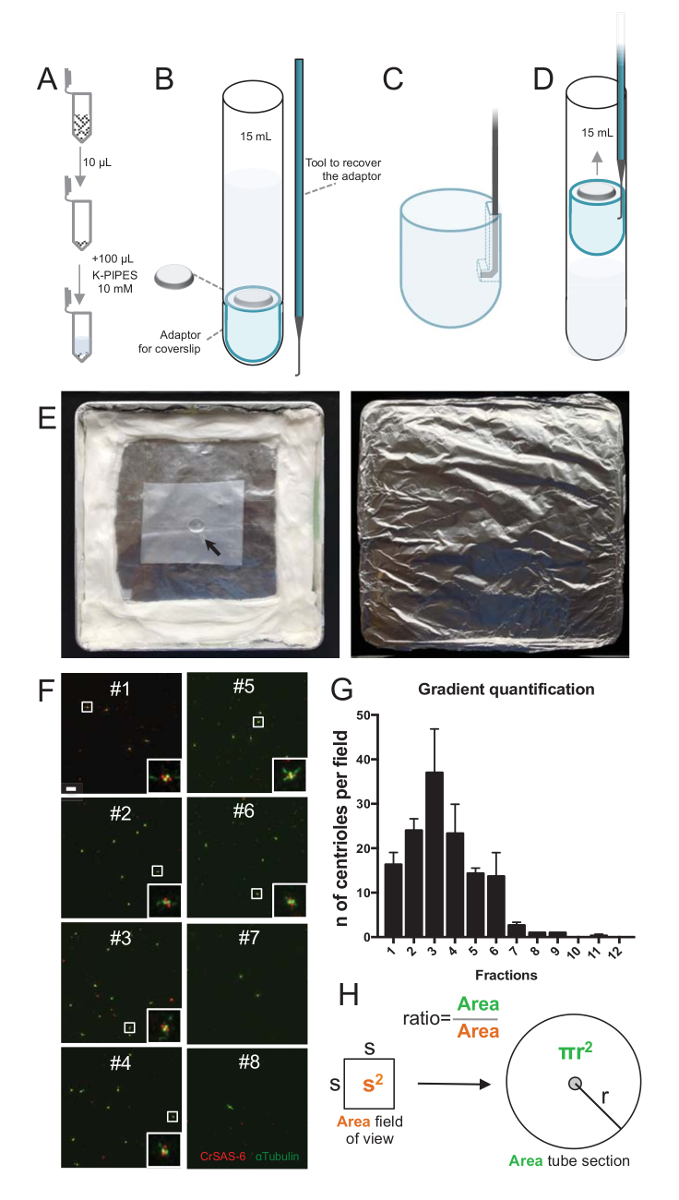
Not: Şekil 2bakın.
-
Tüpler ve coverslips hazırlanması
- 1 cam yuvarlak alt tüp (15 mL) kesir centriolar kesirler (Toplam 12 tüpler) çözümlemek için kullanın.
- (Bundan sonra adaptör olarak adlandırılır) bir özel coverslip destek adaptör yuvarlak alt tüp içine koymak. Steril 12 mm coverslip yuvarlak alt tüp içine yerleştirin.
Not: Burada 12 mm coverslips kullanılan, ancak protokol 30 ml Yuvarlak alt borular kullanarak 18 mm coverslips için adapte edilebilir. - Önceden soğutulmuş 10 mM K-borular (pH 7.2) 5 ml 4 ° C'de ekleyin Coverslip değil yüzüyor ve aşağı bağdaştırıcısında kalır emin olun. Tüpler buza koyun.
-
Santrifüjü centrioles
- Her 10 µL kesir ile soğuk 10 mm K-borular (pH 7.2) 100 µL sulandırmak. Tam ortadan kaybolması kadar seyreltme sukroz (Şekil 2A), resuspend. Her seyreltilmiş kesir bir yuvarlak alt tüp içine yükleyin.
- Tüp 10.000 x g 10 dk (Örneğin, bir JS-13,1 sallanan kova rotor ile) için de 4 ° C'de spin
- Coverslip adaptör oluklu kenarına mevcut deliğe el yapımı bir çengel cihaz ekleyerek yeniden elde etmek ve yavaşça yukarı kaldırın.
Not: El yapımı çengel aygıtı el ile çengel ve ev yapımı bir sopa (rakamlar 2B-2D) kalıp şırınga iğne ile yapılabilir. - Yuvarlak alt Tüp üst ulaşan, üzerine eldivenli bir parmak ile adaptör kenarına tuzak ve coverslip cımbızla kaldırın. Centrioles coverslip hangi tarafında içerir unutmayın için dikkatli olun. Ayirt coverslips tedavi etmek için devam edin.
-
Boyama ve, Imaging ayirt C. reinhardtii centrioles izole
Not: Şekil 2bakın.- Malzeme aşağıdaki gibi boyama ayirt için hazırlayın.
- Bir kristal polistiren iletim laboratuvar kutusunda (60 mm uzunluğunda 50 mm genişliğinde 43 mm içinde yükseklik tarafından tarafından)-cam boyama rafa hazırlamak. % 100 metanol ile doldurun ve -20 ° C'de saklayın
- Nemli bir odası hazırlamak. İçin nemli odası ile birlikte iç su nemlendirilmelidir doku yerleştirerek monte bir kare Petri dish (Şekil 2E) kenarlarını. Şal mühürleme laboratuvar bir parça eklemek ( Tablo malzemelerigörmek) ortasına kadar Petri kabına üzerine antikor karışımları ayirt işlem sırasında (adımları 3.3.2–3.3.3) yerleştirilmiş. Kapak kapağı ve ışıktan korumak için alüminyum folyo ile nemli odası.
- Immunostain izole centrioles olarak izler.
- Santrifüjü (adım 3.2.4) onları kutusunda 5 min için kuluçka tarafından-20 ° C metanol (adım 3.3.1.1) ile dolu sonra coverslips centrioles ile doğrudan tamir et.
- Cımbız ve yer onları şeffaf laboratuvar kutusunda (bkz. Tablo reçetesi) 50 mL 1 x PBS ile dolu ve oda sıcaklığında 5 min için yıkayın coverslips kaldırın.
- Birincil antikor Mix 60 µL pipet [birincil antikorlar % 1 sığır serum albumin (BSA) ve % 0.05 seyreltilmiş ara-20 PBS] sarma nemli odasında mühürleme laboratuvar parçası üzerinde. Dikkatle coverslips antikor karışımı üzerine doğrudan açılan karşı karşıya centrioles ile yatıyordu. Coverslips birincil antikorlar ile 45 dk için kuluçkaya.
Not: temsilcisi sonuçları oluşturmak için kullanılan birincil antikorlar tavşan poliklonal Bld12 (1:300) ve fare α-tübülin (DM1A) (1:300). - Coverslips kaldırmak ve 3.3.2.2. adımda açıklandığı gibi 1 x PBS içinde 5 dakika yıkayın. Coverslips % 1 BSA ve % 0.05 içeren PBS ikincil antikor ile 45 dk için kuluçkaya ara-20.
Not: temsilcisi sonuçları oluşturmak için kullanılan ikincil antikorlar keçi Anti-fare Alexa 488 (1:1, 000) birleştiğinde ve keçi Anti-tavşan Alexa 568 (1:1, 000) birleştiğinde vardır. - Coverslips kaldırmak ve 3.3.2.2. adımda açıklandığı gibi 1 x PBS içinde 5 dakika yıkayın.
- Coverslips bir cam slayt üzerinde orta slayt üzerinde montaj ve dikkatle coverslips (centrioles montaj orta karşı karşıya) üstüne yerleştirerek 3 µL ekleyerek bağlayın. Coverslip kenarına oje ile kapatın.
- Deconvolution27 uygulanırken izole centrioles petrol amacı ile bir N.A. 1.4 X 63, confocal mikroskop üzerinde görüntü ( Tablo malzemelerigörmek).
Not: Burada, aşağıdaki ayarlar kullanıldı: 500-545 nm Alexa 488 ve 580-635 nm Alexa 568 için için.
- Malzeme aşağıdaki gibi boyama ayirt için hazırlayın.
4. Centrioles Coverslips Merkezi üzerine konsantrasyonu
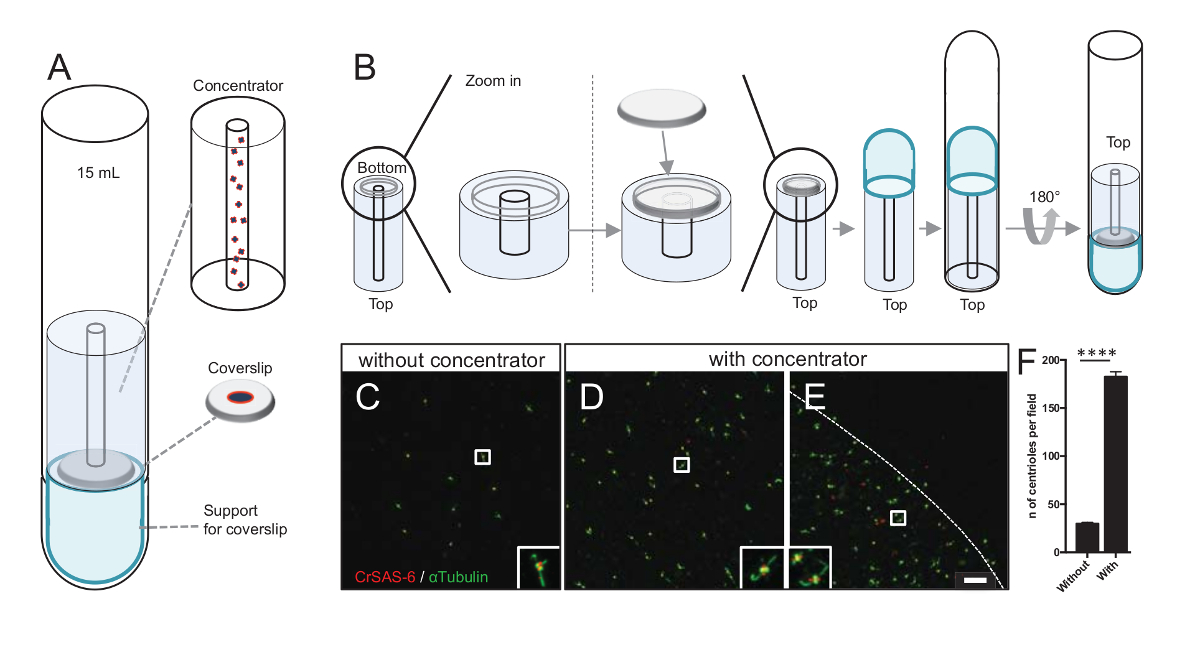
Not: Bkz. Şekil 3.
-
Malzeme hazırlama
- 15 mL Cam yuvarlak alt tüp buz, bir özel coverslip destek adaptör (adaptör bundan sonra .stl dosyasını ek dosya 1sağlanan olarak adlandırılır), bir özel yoğunlaştırıcı ( ek dosya 2olarak sağlanan .stl dosyası) ve 10 mM K-borular (pH hazırlamak 7.2) 4 ° C'de
- Poli-D-lizin (PDL) hazırlamak-kaplı coverslips. 10 x 1 mg/mL PDL hisse senedi çözüm H2O. sulandırmak İlk olarak, coverslips %70 alkol, kaldır etanol ve coverslips kuru izin ile yıkayın. PDL ile coverslips kat ve oda sıcaklığında 30 dk için kuluçkaya. 3 coverslips yıkama su ile x ve koyup kuruyuncaya kadar bırakın.
Not: Kat izole centrioles sayısını artırmak için PDL ile coverslips için coverslips bağlı.
-
Santrifüjü
- Steril 12 mm coverslip gömme üzerine yerleştirin, alt-son yoğunlaştırıcının PDL tutmak ceket yüzü koyun. Coverslip doğrudan üstüne adaptör yerleştirerek kap. Yuvarlak alt tüp ters çevir ve yoğunlaştırıcı, coverslip ve adaptör yerleştirin.
- Nazikçe alt kısmında yuvarlak alt tüp ulaşana kadar ensemble cımbız ile itmek ve tüp tersine çevirin. Dar kurutma başlığını tepesine gelene 10 mM K-borular arabellek (pH 7.2) yuvarlak alt tüp ekleyin. Değil herhangi bir baloncuklar yoğunlaştırıcının Merkez silindir içinde kaldığından emin olun.
- Yavaşça 10 mM K-borular arabelleği (pH 7.2) 100 µL zenginleştirilmiş centriole kesir içeren bir aliquot ve birimin iyice karıştırın.
- 10 mM K-borular arabelleği (pH 7.2) 100 µL yoğunlaştırıcıyı içi boş Merkezi'nden kaldırın ve zenginleştirilmiş centriolar fraksiyonunun 100 µL içeriği boş ortasında kalır dikkat çekici yoğunlaştırıcının içi boş merkezine 10 mM K-borular arabellekte (pH 7.2) ekleyin.
- 10.000 x g önceden Soğutmalı Santrifüj 4 ° C'de 10 min (Örneğin, bir JS-13,1 sallanan kova rotor ile) için de santrifüj kapasitesi
- Dar kurutma başlığını cımbızla kaldırın.
- Coverslip adaptör oluklu kenarına mevcut deliğe el yapımı bir çengel cihaz ekleyerek yeniden elde etmek ve yavaşça yukarı kaldırın. Yuvarlak alt Tüp üst ulaşan, üzerine eldivenli bir parmak ile adaptör kenarına tuzak ve coverslip cımbızla kaldırın. Coverslip hangi tarafında centrioles içeren hatırlamak dikkat et. Ayirt coverslips tedavi etmek için devam edin.
Not: El yapımı çengel aygıtı el ile çengel ve ev yapımı bir sopa (Rakamlar 2B-D) kalıp bir şırınga iğnesi ile yapılabilir. - Fiksasyon ve adım 3.3.2–3.3.4 içinde bitmiş gibi konsantre centrioles ayirt boyama gerçekleştirmek.
5. tek-parçacık ortalama
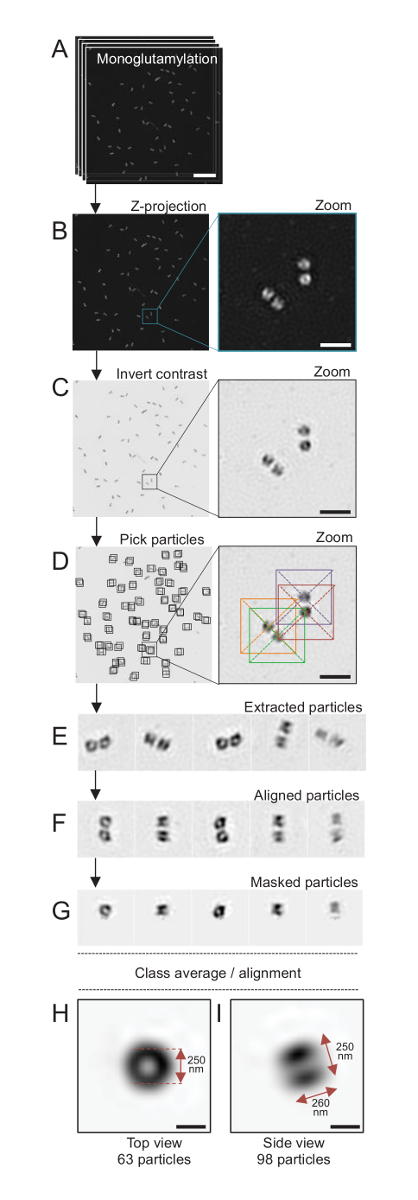
Not: Bkz. Şekil 4.
- Tek-parçacık ortalama için görüntüleme
- Coverslip bir düzenli Anti-solma montaj orta kullanarak bir slaytta bağlayın. CFI Apochromat TIRF amaç kullanarak yapılandırılmış aydınlatma mikroskobu (2D SIM) görüntüleme gerçekleştirmek (100 X, NA 1.49, WD 0,12 mm) ve bir arka ışıklı EM CCD kamera.
Not: 100 ms 3 MHz dışında okumak bir kamera satın alma zaman ayarlandı. 2.5 X objektif kullanılan SIM görüntüleme için.
Not: burada sunulan veri kümesi bir 3-b SIM mikroskobunun elde edildi ( Tablo malzemelerigörmek). - Görüntülerin toplam centriole oluşan büyük bir yığın edinme tarafından centrioles sinyal, sırasıyla üst ve alt pozisyon Z-yığın yukarıda ve aşağıda centriole sinyal ayarlayarak görüntü. Yığın proje ve tek-parçacık adım başı olarak 5.2 ve 5.3 Ortalama gerçekleştirmek için devam edin.
- Coverslip bir düzenli Anti-solma montaj orta kullanarak bir slaytta bağlayın. CFI Apochromat TIRF amaç kullanarak yapılandırılmış aydınlatma mikroskobu (2D SIM) görüntüleme gerçekleştirmek (100 X, NA 1.49, WD 0,12 mm) ve bir arka ışıklı EM CCD kamera.
- Bir yığın projeksiyon
- Görüntü yığını ile ImageJ açın. O zaman tıkırtı üstünde 'görüntü → yığınları → Z proje'. Koymak belgili tanımlık projeksiyon tip olarakMax yoğunluk'.
- Görüntü yanında tıkırtı üstünde ters çevir'i 'düzenleme → ters çevir'i'. Oluşturulan projeksiyon kaydetmek (.tif biçimi).
- Scipion'u hizalamayla tek-parçacık
- Kırmızı basarak durum içinde yeni bir proje oluşturun ' sayfanın üstündekiProje oluşturmak' düğmesi. Sol panelde, çift-tıkırtı üstünde 'Imports → alma filmler'.
- Doldurmak 'dosyaları dizin' ve 'desen ' alanları veri dizini ve adları göre. Varsayılan parametreler tutmak. 'Çalıştır'seçeneğini tıklatın.
- 'Parçacıklar → malzeme çekme → xmipp3 - (adım 1) toplama el kitabı'üzerinde çift tıklatın. 'Giriş Filmler' alan yakın büyüteç simgesini tıklatın ve alınan Filmler 5.3.1 adımından seçin. 'Çalıştır'seçeneğini tıklatın.
- Yeni açılan pencerede--dan farklı filmler yanında tıkırtı üstünde onları seçen. Her test ile bittiğinde, kırmızı düğmeye '+ koordinatları'üzerinde'ı tıklatın.
- 'Parçacıklar → özü → xmipp3 - özü parçacıklar'üzerinde çift tıklatın. 'Giriş koordinatları' alan yakın büyüteç simgesini tıklatın ve çekilen koordinatları 5.3.4 adımından seçin. 'Parçacık kutusu boyutu (piksel)' parçacıklar boyutları göre doldurun. 'Preprocess' sekmesinde toz kaldırma ayarlayın: yok (Bu eserler üretebilir); Kontrast tersine çevirme: yok (siyah parçacıklar, beyaz renkli); Faz flipping: Hayır (CTF düzeltme bağlı); Normalize: Evet.
- Sonra 'Execute'üzerinde'ı tıklatın. İş bittiğinde, ayıklanan parçacıklar iş (daha kalın hale sınırları) ve tıkırtı üstünde kutusunu seçerek kontrol 'analiz sonuçları (ana menünün sol alt).
- '2D → → xmipp3 hizalama - cl2d ile Hizala'üzerinde çift tıklatın. 'Giriş parçacıklar' alan yakın büyüteç simgesini tıklatın ve hulâsa--dan adım 5.3.5 seçen. Bir başvuru yansıması kullanma. 'Çalıştır'seçeneğini tıklatın.
- Çift-týkýrtý üstünde ' 2D → → daha fazla → xmipp3 - hizalama hizalama uygulamak 2d'. Hizalanmış 5.3.7 adımından seçen ve 'Giriş parçacıklar' alan yakın büyüteç simgesine tıklayın.
- 'Execute' üzerinde adım 5.3.6 olduğu gibi'ı tıklatın. Sonuçları iş kutusu seçimden sonra 'Analiz sonuçları' ' ı tıklatarak kontrol edilebilir.
- 'Parçacıklar → maske → xmipp3 - 2d maskesi uygulama'üzerinde çift tıklatın.
- Hizalanmış adımından 5.3.9 seçen ve 'Giriş parçacıklar' alan yakın büyüteç simgesine tıklayın. 'Maskesi kaynağı' 'Geometri'ayarlanır. Sonra maskeyi parametrelerinin maske türü olarak ayarlayın: dairesel; RADIUS (piksel): bir parçacık (bir centriole) etrafında bir şey olmadan arıyorsunuz, tıkırtı üstünde 'sihirli değnek ' mükemmel değer; bulmanıza yardımcı olmak için bir pencere açılır sol üstünde kutsal kişilerin resmi Shift Merkezi: Eğer (parçacık mükemmel ortalanır) Hayır veya Evet (parçacık değiştirdi Eğer); X-merkezi uzaklık: göre parçacık pozisyon; Y merkezine uzaklığı: parçacık konumuna göre. 'Çalıştır'seçeneğini tıklatın.
- Maskeyi doğru bir şekilde uygulanmış olup olmadığını kontrol etmek için ' analiz sonuçları ' düğmesini kullanın. Eğer değilse, '2d maske Uygula' işi sağ tıklatın ve 'Düzenle'yi seçin. Tekrar ile 'Execute' düğme maskesi (boyutu ve/veya vardiya) ve çalışma parametrelerini değiştirin.
- '2D → sınıflandırmak → xmipp3 - cl2d'üzerinde çift tıklatın.
- Maskeli 5.3.11 adımından seçen ve 'Giriş parçacıklar' alan yakın büyüteç simgesine tıklayın. Sınıflar sayısı yaklaşık 50 parçacıklar her sınıfta elde etmek olmalıdır. 'Çalıştır'seçeneğini tıklatın.
- Sonuçları ' analiz sonuçları ' butonuna basarak denetleyebilirsiniz. Açılan pencerede, hemen yanındaki "Ne göstermek" 'göz' simgesini tıklatın.
- Yeni pencere denetim bir oluşturulmuş sınıfı 'Classes2D' 'Blok' menüsünden seçerek sağlar. Her sınıfın içerik aynı menüde 'Class00N_Particles' seçerek kontrol edin. Hangilerinin tek kötü parçacıkları içeren tanımlamak için her sınıf bakarak kontrol edin. 'Classes2D' görünümüne geri dönün ve iyi parçacıklar sınıflarıyla her birini tıklatarak seçin. 'Ctrl' tuşunu seçim sırasında basılı tutarak çeşitli sınıflar seçmek mümkündür.
- İyi parçacıkları ile tüm sınıfları seçtikten sonra bir alt bu parçacıklar ile oluşturmak için '+ parçacıklar' tıklayın.
- Aynı pencerede birkaç farklı yönelimleri parçacıkların temsil eden sınıfları kümesi seçin. Yeni bir alt '+ ortalamalar'üzerinde tıklatarak oluşturun.
- Her yönlendirme/seçili ortalama için aşağıdaki gibi yapın.
- '2D → → xmipp3 hizalama - cl2d ile Hizala'üzerinde çift tıklatın.
- 'Giriş parçacıklar' alan yakın büyüteç simgesini tıklatın ve iyi 5.3.17 adımından seçen. 'Kullanım bir başvuru yansıması' 'Evet'olarak ayarlayın. 'Başvuru yansıması' alan yakın büyüteç simgesine tıklayın ve beyaz ok sol tarafta 'alt oluştur' iş adımından 5.3.18 tıklatın ve bir referans olarak kullanmak üzere resim seçin. Ayrı bir pencerede açmak ve hangi nesnenin kontrol için bir nesne üzerinde çift tıklatın. 'Çalıştır'seçeneğini tıklatın.
- Çift-týkýrtý üstünde ' 2D → → daha fazla → xmipp3 - hizalama hizalama uygulamak 2d'. Hizalanmış adımından 5.3.19.2 seçen ve 'Giriş parçacıklar' alan yakın büyüteç simgesine tıklayın.
- 'Çalıştır'seçeneğini tıklatın.
- Hizalama sonucu kontrol edin ('Analiz sonuçları); Bu hizalanmış parçacıklar ve bu parçacıklar ortalamasını gösterir.
- Ortalama iyi ise ve parçacıkları tüm aynı şekilde odaklı, ortalama yanında tıkırtı üstünde kurtarmak ' → Gelişmiş ImageJ' ve ImageJ ile kaydetmek.
- Ortalama geliştirilebilir, iyi yönlendirmeli görüntüler'i seçin ve yeni bir alt ile '+ parçacıklar' düğme oluşturmak. Alt küme küme (kaldırıldı tüm kötü partikülleri) temizlendikten kadar adımları 5.3.19.1–5.3.19.4 yinelemek. Her zaman hizalama (adım 5.3.19.2) gerçekleştirilen, başvuru için son oluşturulan ortalama (üzerinden yineleme yinelemeye, ortalama kalite artar) ayarlanır.
Sonuçlar
C. reinhardtii Centriole yalıtım:
Centrioles, cw15- yalıtmak için C. reinhardtii hücreleri birkaç gün ışığı altında sıvı kültüründe yetiştirilen ve 10 dk. Pelleted hücreleri 1 x PBS ile yıkanmış için daha sonra Santrifüjü vasıl 600 x g tarafından pelleted ve içinde resuspended bir arabellek öncesinde son pH 4,5-4,7 değeri 0,5 M asetik asit 2 dk(Şekil 1)kullanarak bir pH şok gerçekleştirerek deflagellation deflagellation. 1 N KOH ekleme pH 7,0 için geri yüklemek için kullanıldı. Müstakil kamçı hücre vücutlarından ayırmak için deflagellated hücreleri ilk kamçı toplu kaldırmak için centrifuged. Pelet yapıldı sonra PBS ile 2 x yıkanmış ve yavaş yavaş bir % 25-sükroz yastık (Şekil 1B) önce yükleme 30 mL PBS resuspended. Tüp 600 x g 4 ° C'de müstakil kamçı çoğunu kaldırmak için 15 dk, bükülmüş. Santrifüjü sonra hücre cesetleri yayılır sukroz yastık (Şekil 1C) ve kaldırarak kurtarılan yaklaşık 30 mL süpernatant (kadar kırmızı ok Şekil 1C). Yıkanmış hücre elde edilen 20 mL sonra 20 mL soğuk PBS resuspended ve 600 x g 10 min için de centrifuged. Daha sonra süpernatant atıldı ve hücreleri tamamen 10 mL PBS resuspended. Hücreleri 250 mL şişe transfer edildi ve lizis arabellek resuspended hücrelere hemen eklendi. DNaz lizis için eklendi ve 1 h 4 ° C'de inkübe Hücre artıkları kaldırmak için bir Santrifüjü adım sonra (bkz. Şekil 1Dbeyaz Pelet) süpernatant ve dikkatle üzerine 2 mL % 60-sükroz yastık Santrifüjü 10.000 x g 4 ° C'de 30 dk için de önce yüklü toplanır Lizis için 100 mL unutmayın, 15 mL 8 Tüpler 12,5 mL sukroz tüp başına 2 mL yüklendi lizis arabellek karşılık gelen bu adımı gerçekleştirmek için kullanıldı. Santrifüjü sonra süpernatant çoğunu kaldırıldı ve minder üstünde ilâ 1 mL. Kalan süpernatant 1 mL sonra 2 mL yastık ile toplanan ve daha sonra karışık ve 24 mL son hacmi elde etmek için havuza alınmış. Havuz sonra bir 40 dolu olduğunu % - %50, % 70-sükroz degrade ve bükülmüş 68,320 x g 1 saat 15 dk 4 ° C'de için de Son olarak, izole centrioles bir iğne kullanarak Santrifüjü tüp alt kısmında bir delik yaparak toplanmıştır ve damla 12 x 500 µL kesirler içinde toplanmıştır. Farklı sükroz yüksek yoğunluk nedeniyle, çok yavaş başlangıç (% 70 sükroz) ve sonra daha fazla damla kurulan hızla (% 40 sükroz).
İzole Centrioles ayirt
Yalıtım yordamı kalitesini değerlendirmek için toplanan her degrade kesir 10 µL sonra bir coverslip destek adaptör (Şekil 2A-2B) kullanarak bir coverslip centrifuged. Önemlisi, coverslip güvenle kaldırmak için özel bir çengel aygıt dizayn edilmiştir (Şekil 2B). Daha sonra coverslips ayirt tarafından analiz edildi. CrSAS-6(Bld12p) karşı antikor Bu çalışmada çember hareketi yapısı ve α-tübülin (DMA1) centriolar duvar vurgulamak için varlığı belirtmek için kullanılmıştır. CrSAS-6 ve α-tübülin görüş alanı başına olumlu centrioles sayılması ve centrioles kesirler başına toplam sayısının hesaplanmasında hangi kesirler izole centrioles ( zenginleştirilmiş belirlemek mümkün Rakam 2F-2 H). Son kesirler arıtma işe yaradığını gösteren değil, iken ilginçtir, 6 kesirler (Şekil 2F ve 2 G, kesirler #1-6), centrioles için kesir #3, bir tepe ile zenginleştirilmiş. Belirli bu deneyde, toplam centrioles % 95'i CrSAS-6 ve α-tübülin kesir #3 için olumlu olduğunu unutmayın. Bu en izole centrioles onların saldırı korunur belirtir. Hiçbir zenginlik centrioles ilk kesirler gözlem yapılırsa, yalıtım yordam işe yaramadı ve yinelenmelidir. Not Bazı kamçı parçalar çoğunlukla kesirler centrioles yoksun olarak görülebilir.
Ardından, centrioles µL başına toplam sayısını hesaplamak için sayısının centriole görüş alanı başına Şekil 2' deHsunulan oranla çarpımı. Ortaya çıkan sayı sonra için ayirt centrioles µL başına sayısı elde etmek için kullanılan kesir hacmen bölünmüş olmalıdır. Bu özel yalıtım yordamda en zenginleştirilmiş kesir (ile görüş alanı 92 x 92 µm2) 0.00846 mm2 bir bölgenin yaklaşık 37 centrioles içerdiği. Yüzey tüp bölümünün 7.5 mm çapındaki 176 mm2toplam yüzey ile oldu. Karşılık gelen oranı sonra 176/0.00846 oldu 20,803.8, yani = toplam 10 µL içinde 769,740 centrioles (37 x 20, 803.8). Bu nedenle, 1 µL centrioles sayısı 76,974 oldu.
İzole Centrioles konsantrasyonu Coverslips üzerinde:
Centrioles başına sayısını artırarak daha fazla parçacık ortalama yordamlar için kullanılan benzer yönler tespit şansımızın artması yanı sıra kesin centriole yönünü tespit olasılığını artırır. Centrioles konsantre kesirler üzerinden coverslip üzerinde hala seyrek olduğu için centrioles bir yoğunlaştırıcı adlı coverslip (Şekil 3A) ortasında konsantre bir Santrifüjü aksesuar geliştirdik. 3-b yazdırma için kesin önlemler ile bir .stl dosyası bu el yazması ile temin edilebilir.
İlk olarak, 12 mm bir coverslip yoğunlaştırıcı (Şekil 3B) monte edilmiş. Adaptör coverslip üzerine yerleştirildi ve yuvarlak alt tüp ters ve birleştirilmiş adaptör ve yoğunlaştırıcı yerleştirilir. Yuvarlak alt tüp sonra yavaşça, böylece örnek (iletişim kuralı adım 4.2) yüklenmesini sağlayan ters. Centrioles sonra 10.000 x g 4 ° C'de 10 dakika için de centrifuged Bundan sonra centrioles ayirt için tabi ve CrSAS-6/Bld12p ve α-tübülin (Şekil 3 c-3E) için lekeli. Önemlisi, dar kurutma başlığını kullanılan (şekil 3D-3F) yaşındayken 183 centrioles görüş alanı görüldü ise yoğunlaştırıcı, görüş alanı (Şekil 3C), yaklaşık 30 centrioles görüldü. Centrioles 4 mm çapında coverslip ortasında tek bir diski kaplı unutmayın. Bu sonuç konsantrasyon adım çalışır ve böylece algılama hareket hızı ve görüntüleme coverslips tanımlanmış bir bölgede centrioles 6-fold bir zenginlik sağlar gösterir.
Floresan tek-parçacık izole, ortalama C. reinhardtii Centrioles:
Burada, yaklaşık 120 çözünürlüğe ulaşabilirsiniz SIM mikroskobu kullanarak görüntülü24nm, monoglutamylated tübülin (GT335, Şekil 4), centriolar mikrotübüller içinde mevcut bir tübülin değişiklik için lekeli centrioles vardı. C. reinhardtii centrioles olan yaklaşık 500 nm her zaman çiftler halinde uzun ve sık sık bulunan ile ilişkili olan, yeni probasal organları (procentrioles bundan sonra adlandırılır) çoğaltılamaz ve çizgili Mikrotubul ilişkili lifleri19. Bu nedenle, bu son montaj yaklaşık 1 µm büyük oldu. Bu nedenle ve tamamen centrioles görüntü için Z yığında izole centrioles satın öneririz.
Burada, satın alma sonra ImageJ28 (resim/yığınları/Z proje/en fazla yoğunluk projeksiyon, Şekil 4B) kullanarak bir maksimum yoğunluk projeksiyon yaparak son görüntü oluşturuldu. Böyle görüntüleri, bir tek-parçacık analiz cryo-elektron mikroskobu yazılımını kullanarak sınıfların centrioles benzer yönler ile yapmak gerçekleştirildi ve sonra ortalama gerçekleştirildi. Bunu yapmak için resmin rengini ilk nesneleri (Şekil 4C) daha iyi görmek için ters. Centrioles aldı el ile Xmipp3 gibi birkaç elektron mikroskobu yazılım programları entegre ve serbestçe kullanılabilir durum yazılım29 kullanarak her parçacık üzerinde ortalanmış bir kutusunda (Şekil 4D). Kutusunun boyutunu Kullanıcı tarafından tanımlanmış olması gerektiğini unutmayın. Burada, 31.84 bir piksel boyutu için 50 x 50 piksel kutularca nm kullanılmıştır. Ardından, tüm parçacıklar vardı (Şekil 4E) çıkarılan ve Xmipp3 kullanarak uyumlu (Şekil 4F). Ardından, dairesel bir maske 12 piksel yarıçapı her centriole centriolar çifti (Şekil 4G) üzerinden yalıtmak için uygulandı. Parçacıklar sonra sınıflandırılmış Xmipp3, birkaç ortalamalar üretmek için kullanıyorum. Sadece sınıf ortalamalar sapmak parçacıklar el ile dışlandı anlam homojen sınıf ortalamaları tutuldu. Bu adım, seçilen her yönü için kusursuza yakın ortalama oluşturmak için tekrarlandı. Beş yineleme sonra iki sınıf ortalama örneği oluşturulan: 63 nesneleri (Şekil 4H) en iyi bir görünüm ve bir yan görüntülemek 98 parçacıkların monoglutamylated centrioles (Şekil 4ben). Nesne boyutları yoğunluğu Arsa profil monoglutamylation sinyal boyunca doruklarına arasındaki uzaklığı ölçülerek belirlenmiştir.
Önemlisi, yan görünümü sınıf ortalama uzunluğu 260 ise nm çapında 250 nm, çekirdek içinde uzunluğu centriole17286 ± 33 nm bir bölgesine yerelleştirmek için gösterildi ölçülen monoglutamylated tübülin sinyal karşılaştırılabilir.

Resim 1 : Arıtma C. reinhardtii centrioles. (A)bu C. reinhardtii centrioles yalıtım için önde gelen her adım şematik gösterimi. Santrifüjü % 25-sükroz degrade üzerine sonra önce Protokolü (B) ve (C) temsilcisi adımları içerir. C panelinde kırmızı ok Santrifüjü sonra tutmak için en küçük birim gösterir. (D) Bu panel lysed hücre beyaz Pelet Santrifüjü sonra gösterir. Siyah oku Pelet gösterir. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Resim 2 : Ayirt gerçekleştirileceği Santrifüjü set-up izole centrioles. (A) 10 µL toplanan her kesir, 100 µL (pH 7.2) 10 mM K-borular içinde seyreltilmiş ilk. (B) bu kapsayan bir 15 mL yuvarlak alt Tüp, 12 mm coverslip, coverslip için bir adaptör Santrifüjü cihazlar ve coverslip sonra Santrifüjü kurtarmak için ısmarlama bir çengel cihaz, şematik bir gösterimidir. Panelleri C ve D coverslip aralıklarla sonra kurtarmak açıklayan çizimler göster. (D) ve (C) yer çengel araç adaptörü oluklu kenarına mevcut deliğe yavaşça çekin. (E) bu resimleri ayirt görüntüleme gerçekleştirmek için gereken nemli odası vardır. Ok 12 mm coverslip gösterir. (F) bu (2 zoom) 63 X degrade kesirler #1-8, arıtma işlemi sırasında toplanan temsilcisi confocal görüntülere ve CrSAS-6 (kırmızı) ve α-tübülin (yeşil) için lekeli. İnsets beyaz kutu daha düşük büyütme sayısı olarak belirtilen bölge karşılık gelir. Ölçek çubuğu 10 µm. (G) = CrSAS-6 ve α-tübülin her kesir görüş alanı başına centrioles olumlu sayısını gösteren bir grafiktir. Zenginleştirme kesirler #1-6 Not. Centrioles her kesir alanında başına ortalama sayısı: #1 = 16,3 ± 4.7, #2 = 24.0 ± 4.6, #3 = 37,0 ± 17,0, #4 = 23.3 ± 11,4, #5 = 14,3 ± 2.1, #6 = 13,7 ± 9.3, #7 = 2.7 ± 1.2, #8 = 1.0 ± 0.0, #9 = 1.0 ± 0.0, #10 = 0.0 ± 0.0, #11 = ± 0,3 0,6, #12 = 0.0 ± 0.0. Her bölümü için 3 rasgele alanı görüntüsü. En zenginleştirilmiş kesir #3 sayarken centrioles % 95'i CrSAS-6 ve α-tübülin için olumlu bulduğumuzu, Not (n = 205 centrioles). Centrioles filmler ölçülen alanın mevcut sayısı oranını (H) A centrioles tüp alanında mevcut toplam sayısını hesaplamak için kullanılır. Centriole numarası µL başına 10 µL aslında centrifuged kesirler hesaplanır. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Şekil 3 : İzole centrioles centrifuged bağlantı, dağıtma kutuları kullanarak. (A) Bu panel gösterir centrioles coverslips ayirt 15 mL yuvarlak alt Tüp, yoğunlaştırıcı, 12 mm coverslip ve bir destek adaptör aparatı dahil olmak üzere önce üzerine konsantre için gerekli Santrifüjü cihazlar. (B) Bu panel Santrifüjü cihaz montajı için gereken adımları gösterir. Panelleri C-E CrSAS-6 (kırmızı) ve α-tübülin (yeşil) (C) olmadan veya (D-E) için dar kurutma başlığını ile lekeli centrioles confocal görüntülerini göster. İnsets beyaz kutu daha düşük büyütme sayısı olarak belirtilen bölge karşılık gelir. Ölçek çubuğu 10 µm. centrioles (konsantre alan kenarlığını noktalı çizgi gösterir) coverslip ortasında zenginleştirilmiş Not olduğunu. (F) Bu grafik centrioles görüş alanı olmadan ve dar kurutma başlığını başına sayısını temsil eder. Beş rastgele alanlar bakış analiz edildi. Centrioles ortalama sayısı, yoğunlaştırıcı, 29.8 ± 2.9, olmadan ve yoğunlaştırıcı, olduğunu 182.6 ± 11.5, P < 0,0001. İstatistiksel anlamlılık bir unpaired ttarafından değerlendirildi-test. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Şekil 4 : Tek-parçacık izole üzerinde ortalama C. reinhardtii centrioles. (A) Bu panel gösterir Z yığın görüntüleri ile GT335 lekeli ve SIM mikroskop kullanarak elde centrioles. (B) bu paneller bir maksimal yoğunluğu Z projeksiyon yığılmış görüntülerin göster. Ölçek çubuğu 1 µm. (C) bu panelleri göstermek bir temsili resim ters bir kontrast =. İlave bir zum centrioles daha iyi görselleştirmek için bileşenini temsil eder. (D) bu paneller Haritayı parçacık malzeme çekme. İlave nasıl parçacıklar tutuklandılar gösterir. (E) çıkarılan 5 örnekleri bunlar parçacıklar. (F) Bu panel parçacıklar hizalama sonra gösterir. (G) Bu panel parçacıklar bir maske uygulandıktan sonra gösterir. Panelleri H ve ı göstermek iki sınıf ortalama: (H) bir üst görünüm (63 parçacıklar) ve (ı) bir yan görünüm (98 parçacıklar). Çift Kişilik oklar GT335 sinyal boyutları gösterir. Ölçek çubuğu = 250 nm. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}
Ek dosya 1. Bu dosyayı indirmek için buraya tıklayınız.
Ek dosya 2. Bu dosyayı indirmek için buraya tıklayınız.
Tartışmalar
Biyolojide zorluklardan biri bir mimari bağlamda proteinlerin kesin yerelleştirme deşifre etmektir. Onun Mimarlık cryo-elektron tomografi, ilginç ultrastructural özellikleri, uzunluğu boyunca açığa kullanarak okudu gibi centriole bu yöntemleri uygulamak için ideal bir yapıdır. Ancak, çözünürlük limit optik mikroskobu içinde yakın boyutları nedeniyle, tam olarak ayirt geleneksel mikroskoplar30kullanarak centriole bir yapısal alt bölgeye göre bir protein yerelleştirmek zordur.
Optik mikroskobu çözünürlük verir, kabaca, 200 yanal maksimum çözünürlük ışığın kırınım tarafından sınırlı optik mikroskobu24nm. Ancak, bu sınır içinde optik mikroskobu son 20 yılda büyük devrimler biri tarafından olmuştur olmuştur: Süper çözünürlük yöntemleri icadı. Bu yaklaşımlardan farklı çözünürlüklerde kırınım sınırları aşan görüntü: 120 nm SIM için yaklaşık 50 nm için uyarılmış emisyonu tükenmesi (STED) ve 20-40 nm tek molekül yerelleştirme mikroskobu (SMLM)24. Süper çözünürlük mikroskobu ile bu yeni gelişmeler centriole yapısal alt bölgelerinde ulaşılabilir. Ancak, uygulamada, doğru bir protein yapısal bir öğeye lokalizasyonu olgun centrioles hücre başına sadece 2 kopya bulunmaktadır ve yorumlanması kılan rasgele yönleri, var temel nedeni belirlemek hala zor Yerelleştirme zor. Bu nedenle, araştırmacılar tarafından Süper çözünürlük centrioles belirsiz sigara yönelimleri gözlemlemek için şansımızın artması, çok sayıda görüntü için izin veren bir protokol kurulmuştur. Bu yöntem izole centrioles kullanımına dayanıyor gibi önemlisi, bozulmamış C. reinhardtii arındırmak için bir yöntem sağlıyoruz olgun centrioles ve procentrioles içeren centrioles.
Son olarak, bu iletişim kuralıyla görüntüsü centriole yönelimleri aralığı sayesinde, tek-parçacık analiz elektron mikroskobu yazılım kullanılarak uygulanabilir. Belirli bir yönde centrioles ortalama sınıfları nesil sonuçlanır. Önemlisi, elde edilen bu 2B görüntüleri daha sonra belirli bir protein centriole boyunca lokalizasyonu değerlendirmek için kullanılabilir. Nitekim, Çift renkli süper çözünürlük fotoğraf için bu yöntem uygulanabilir ve diğer renk iken centriole iskelet (Örneğin, tübülin), belirli bir centriolar protein bağlanabilir ortaya çıkarmak için bir renk kullanılır. Bir renk veya iki renk ile elde edilen ortalamalar çıkararak, bir protein centriole (proksimal, merkezi veya distal) boyunca kayıt daha kolay olur. Not iki kanal doğru bir şekilde herhangi bir yanıltıcı Yorumları önlemek için uyumlaştırılması gerekmektedir. Ayrıca, en iyi sayısı ortalamaları içinde centriolar lümen, Mikrotubul duvar boyunca veya centriole dışında bir protein yerelleştirir Eğer deşifre yardımcı olacaktır.
Bu yöntem Aksi takdirde türdeş olmayan etiketleme nedeniyle yerelleştirmek zor olabilir belirli proteinlerin yerelleştirme tespit etmek için bir avantajdır. Proteinler centrioles içinde eşlemek için diğer yöntemleri bağdaşık 3-b SIM/SMLM ile içinde örnek olarak, centrioles belirli yönelimleri simit çevresinde oluşturan bir işaretleyici eliptik profil belirlenerek değerlendirilmesi tarif edilmistir olduğunu unutmayın Centriole yanında SIM düşsel. Bu parametreyi kullanarak, 4 – 5 nm30bir hassasiyetle protein yerelleştirmek mümkündür. Ayrıca açıklanan yöntemi burada izole centrioles olduğu gibi procentrioles, centriole mimarisi büyük ölçüde korunmuş en yüksek olduğu bir işaret kullandığına dikkat edin. Ancak, bazı mimari özellikleri gibi insan centrosome5yalıtım ile güçlendirilmiş divalent katyonlar konsantrasyon ile değişen centriole çapı gibi arıtma sırasında rahatsız dışlayamazsınız.
Burada sunulan Protokolü'nün önemli adımlardan biri yeterince konsantre izole centrioles farklı yönelimleri Fluo-SPA için mükellef içinde elde etmektir. Bunu yapmak için önce saflık ve centriole yalıtım yordamı verimliliğini sağlamak. İzole centrioles düşük konsantrasyon uygun görüntüleme ve görüntü işleme daha fazla engeller. Bu amaçla, biz centrioles görüş alanı başına sayısı zenginleştirmek için bir yöntem sağlıyor. Centrioles kullanılan kesir sayısına bağlı olarak, Yoğunlaştırıcı yüklenen birim ayarlanmalıdır, maksimum hacmi 250 µL ile.
Önemlisi, bu yöntem bir hücre duvarı cw15-C. reinhardtii hücreleri eksi için geliştirilmiştir. Bu zorlanma, uygun lizis hücrelerin ve içeriği kurtuluş için böylece, hücre duvarı kırılganlık sağlar. Bu iletişim kuralı için vahşi-türü C. reinhardtii verimli değildir hücreleri, hücre duvarı uygun lizis engeller gibi. Alternatif stratejileri sonication veya autolysin, hücre duvarı31, düşürebilir bir enzim içeren hücrelerin ön kuluçka gibi burada sunulan yalıtım iletişim kuralı uygulamadan önce hücre duvarı alter yerde koymak zorunda kalacaktı.
Bu kurulum, mikroskoplar, geleneksel confocal mikroskoplar yüksek üretilen iş adanmış Süper çözünürlük mikroskoplar için arasında değişen farklı türleriyle kullanılabilir. SMLM yaparken, özel bir arabellek uygun görüntüleme için gerekli olduğunu unutmayın ve bu nedenle, arabellek coverslip üstüne ile 12 mm coverslip için uyarlanmış bir odası kullanılmalıdır. Sonraki görüntüleme ile ters mikroskoplar gerçekleştirilecek. Mikroskop set-up bir 12 mm coverslip izin vermiyorsa, burada sunulan protokolü kullanarak bir 30 mL yuvarlak alt tüp ve değiştirilen adaptör ve yoğunlaştırıcı 18 mm coverslips için uygulanabilir. Ayrıca Not SMLM son yeniden kalitesini boyama ve kullanılan birincil antikor kalitesine bağlı olacaktır, hem de fiksasyon yöntemi için önemlidir.
Özetle, çok sayıda centrioles ardından görüntüye Fluo-ortalamalar centrioles farklı yönleri, oluşturacak böylece hassasiyetle centriolar protein yerelleştirilmesine yardımcı SPA tarafından uygulanan bir yöntem sağlıyoruz. Önemlisi, bu yöntem diğer türlerden izole centrioles, diğer organelleri veya büyük makromoleküllerin grupları daha genel olarak uygulanabilir. Son olarak, örnek hazırlık yaklaşım burada sunulan, son algoritma geliştirme floresan SMLM veri32ile tek-parçacık analiz için kombine, daha da iyileştirme moleküler haritacılık büyük içinde makromoleküllerin açar mısınız derlemeler.
Açıklamalar
Yazarlar ifşa gerek yok.
Teşekkürler
École Polytechnique Fédérale de Lausanne adlı (EPFL), Lozan, İsviçre, nerede centrioles SIM görüntüleri elde Pierre Gönczy ve BioImaging & Optik platformu (BIOP) teşekkür ederim. Nikolai Klena ve Davide Gambarotto Avrupa Araştırma Konseyi (ERC) başlangıç Grant (StG) 715289 (vurgu) ve Maeva Le Guennec, Paul Guichard ve Virginie Hamel İsviçre Ulusal Bilim Vakfı (SNSF) PP00P3_157517 tarafından desteklenir. Susanne Borgers Cenevre Üniversitesi tarafından desteklenir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Mouse monoclonal anti-apha tubulin (clone DM1A) | Abcam | ab7291 | dilution 1:300 |
| DNaseI | Roche | 10104159001 | |
| 12 mm coverslips | Roth | YX03.1 | |
| 18 mm coverslips | Roth | LH23.1 | |
| K2HPO4 | Fluka | 60355 | |
| KH2PO4 | Fluka | 60230 | |
| Tris base | Biosolve Chimie SARL | 0020092391BS | |
| acetic acid | Carlo Erba Reagents | 524520 | |
| NH4Cl | Sigma | A-4514 | |
| CaCl2 | Sigma | C-7902 | |
| MgSO4 | Sigma | 63140-500G-F | |
| steritop filter | Millipore | SCGPT05RE | |
| sucrose | Sigma | S7903-1KG | |
| HEPES | AppliChem PanReac | A3724,0250 | |
| PIPES | Sigma | P6757-500G | |
| MgCl2 | ACROS ORGANICS | 197530010 | |
| NP-40 | AppliChem PanReac | A1694,0250 | |
| Round-bottom (Kimble) tubes 15 mL | Fisherscientific | 09-500-34 | |
| Round-bottom (Kimble) tubes 30 mL | Fisherscientific | 09-500-37 | |
| cover glass staining rack | Thomas scientific | 8542E40 | |
| crystal polystyrene transmission lab box | FISHERS | 11712944 | |
| Methanol | VWR | 20864.32 | |
| BSA | Roche | 10735086001 | |
| Triton X100 | Roth | 3051.3 | |
| goat anti-mouse coupled to Alexa 488 | invitrogen | A11029 | dilution 1:1,000 |
| goat anti-rabbit coupled to Alexa 568 | invitrogen | A11036 | dilution 1:1,000 |
| mounting medium | abcam | ab188804 | |
| Tube, thinwall polypropylene | Beckman Coulter | 326823 | |
| Poly-D-Lysine 1 mg/mL | SIGMA | A-003-E | |
| Mouse monoclonal anti-Polyglutamylation modification mAb (GT335) | Adipogen | AG-20B-0020 | dilution 1:1,000 |
| glycerol mounting medium with DAPI and DABCO | Abcam | ab188804 | |
| 50 mL conical tubes | Falcon | 14-432-22 | |
| Eppendorf 5810R centrifuge | Eppendorf | 5811000622 | |
| Beckman JS-13.1 swinging bucket rotor | Beckman Coulter | 346963 | |
| Beckman SW 32Ti rotor | Beckman Coulter | 369694 | |
| parafilm | Bemis | 13-374-10 | |
| Leica TCS SP8 with Hyvolution mode | Leica | ||
| OSRAM L18W/954 LUMILUX | Luxe Daylight/ OSRAM | ||
| Whatman filter paper | Sigma | WHA1001325 | |
| CrSAS-6/Bld12 antibody | dilution 1:300 (Hamel el al., 2014) | ||
| Scipion | http://scipion.i2pc.es/ | ||
| EM CCD camera (Andor iXON DU897) | Andor |
Referanslar
- Bornens, M. The centrosome in cells and organisms. Science. 335 (6067), 422-426 (2012).
- Nigg, E. A., Holland, A. J. Once and only once: mechanisms of centriole duplication and their deregulation in disease. Nature Reviews Molecular Cell Biology. 19 (5), 297-312 (2018).
- Nigg, E. A., Raff, J. W. Centrioles, centrosomes, and cilia in health and disease. Cell. 139 (4), 663-678 (2009).
- Gönczy, P. Towards a molecular architecture of centriole assembly. Nature Reviews Molecular Cell Biology. 13 (7), 425-435 (2012).
- Paintrand, M., Moudjou, M., Delacroix, H., Bornens, M. Centrosome organization and centriole architecture: their sensitivity to divalent cations. Journal of Structural Biology. 108 (2), 107-128 (1992).
- Winey, M., O'Toole, E. Centriole structure. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 369 (1650), (2014).
- Dippell, R. V. The development of basal bodies in paramecium. Proceedings of the National Academy of Sciences of the United States of America. 61 (2), 461 (1968).
- Allen, R. D. The morphogenesis of basal bodies and accessory structures of the cortex of the ciliated protozoan Tetrahymena pyriformis. The Journal of Cell Biology. 40 (3), 716-733 (1969).
- Li, S., Fernandez, J. -. J., Marshall, W. F., Agard, D. A. Three-dimensional structure of basal body triplet revealed by electron cryo-tomography. The EMBO Journal. 31 (3), 552-562 (2012).
- Guichard, P., Chrétien, D., Marco, S., Tassin, A. -. M. Procentriole assembly revealed by cryo-electron tomography. The EMBO Journal. 29 (9), 1565-1572 (2010).
- Guichard, P., et al. Native architecture of the centriole proximal region reveals features underlying its 9-fold radial symmetry. Current Biology. 23 (17), 1620-1628 (2013).
- Hirono, M. Cartwheel assembly. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 369 (1650), (2014).
- Bornens, M., Paintrand, M., Berges, J., Marty, M. C., Karsenti, E. Structural and chemical characterization of isolated centrosomes. Cell Motility and the Cytoskeleton. 8 (3), 238-249 (1987).
- Keller, L. C., Romijn, E. P., Zamora, I., Yates, J. R., Marshall, W. F. Proteomic analysis of isolated chlamydomonas centrioles reveals orthologs of ciliary-disease genes. Current Biology. 15 (12), 1090-1098 (2005).
- Kilburn, C. L., et al. New Tetrahymena basal body protein components identify basal body domain structure. The Journal of Cell Biology. 178 (6), 905-912 (2007).
- Bauer, M., Cubizolles, F., Schmidt, A., Nigg, E. A Quantitative analysis of human centrosome architecture by targeted proteomics and fluorescence imaging. The EMBO Journal. 35 (19), 1-15 (2016).
- Hamel, V., et al. Identification of Chlamydomonas Central Core Centriolar Proteins Reveals a Role for Human WDR90 in Ciliogenesis. Current Biology. 27 (16), 2486-2498 (2017).
- Cavalier-Smith, T. basal body and flagellar development during the vegetative cell cycle and the sexual cycle of Chlamydomonas reinhardii. Journal of Cell Science. 16 (3), 529 (1974).
- Geimer, S., Melkonian, M. The ultrastructure of the Chlamydomonas reinhardtii basal apparatus: identification of an early marker of radial asymmetry inherent in the basal body. Journal of Cell Science. 117, 2663-2674 (2004).
- Nakazawa, Y., Hiraki, M., Kamiya, R., Hirono, M. SAS-6 is a cartwheel protein that establishes the 9-fold symmetry of the centriole. Current Biology. 17 (24), 2169-2174 (2007).
- Hamel, V., et al. Correlative multicolor 3D SIM and STORM microscopy. Biomedical Optics Express. 5 (10), 3326 (2014).
- Guichard, P., et al. Cell-free reconstitution reveals centriole cartwheel assembly mechanisms. Nature Communications. 8, 14813 (2017).
- Hiraki, M., Nakazawa, Y., Kamiya, R., Hirono, M. Bld10p constitutes the cartwheel-spoke tip and stabilizes the 9-fold symmetry of the centriole. Current Biology. 17 (20), 1778-1783 (2007).
- Sahl, S. J., Hell, S. W., Jakobs, S. Fluorescence nanoscopy in cell biology. Nature Reviews Molecular Cell Biology. 18 (11), 685-701 (2017).
- Hutner, S. H., Provasoli, L., Schatz, A., Haskins, C. P. Some Approaches to the Study of the Role of Metals in the Metabolism of Microorganisms. Proceedings of the American Philosophical Society. 94 (2), 152-170 (1950).
- Keller, L. C., Marshall, W. F. Isolation and proteomic analysis of chlamydomonas centrioles. Methods in Molecular Biology. 432 (1), 289-300 (2008).
- Borlinghaus, R. T., Kappel, C. HyVolution-the smart path to confocal super-resolution. Nature Methods. 13 (3), (2016).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- de la Rosa-Trevín, J. M., et al. Scipion: A software framework toward integration, reproducibility and validation in 3D electron microscopy. Journal of Structural Biology. 195 (1), 93-99 (2016).
- Gartenmann, L., et al. A combined 3D-SIM/SMLM approach allows centriole proteins to be localized with a precision of ∼4-5 nm. Current Biology. 27 (19), 1054-1055 (2017).
- Jaenicke, L., Kuhne, W., Spessert, R., Wahle, U., Waffenschmidt, S. Cell-wall lytic enzymes (autolysins) of Chlamydomonas reinhardtii are (hydroxy)proline-specific proteases. European Journal of Biochemistry. 170 (1-2), 485-491 (1987).
- Salas, D., et al. Angular reconstitution-based 3D reconstructions of nanomolecular structures from superresolution light-microscopy images. Proceedings of the National Academy of Sciences of the United States of America. 114 (35), 9273-9278 (2017).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır