
Method Article
常规 rt-pcr 和 sybr 绿色 rt-qpcr 检测罗非鱼
摘要
该协议使用 rt-pcr 方法诊断罗非鱼组织中的罗非鱼病毒 (tilv)。整个方法描述了从组织解剖到总 rna 提取, 然后是 cdna 合成和检测 tilv 的常规 pcr 或定量 pcr 使用 dsdna 结合染料。
摘要
该方法的目的是促进罗非鱼湖病毒 (tlev) 在罗非鱼组织中的快速、灵敏和特异性检测。该协议可作为监测方案、生物安保措施和 tlv 基础研究实验室的一部分使用。病毒诊断的黄金标准通常涉及病毒分离, 然后是补充技术, 如逆转录酶链反应 (rt-pcr), 以供进一步验证。这可能是繁琐的, 耗时的, 通常需要组织样本严重感染病毒。rt-定量 (q) pcr 在病毒检测中的应用具有定量、灵敏度高、特异性强、可扩展性快、可快速的特点, 具有较高的检测效果。本文介绍了基于 pcr 的 tilv 检测方法的全方法, 从罗非鱼器官切片、使用包那氰化物-苯酚-氯仿溶液提取总核糖核酸 (rna)、rna 定量, 然后是两步 pcr协议, 包括补充脱氧核糖核酸 (cdna) 合成和检测 tlv 的常规 pcr 或定量鉴定通过 qpcr 绿色 i 染料。传统的 pcr 需要 pcr 后的步骤, 只需告知病毒的存在。后一种方法将允许对 tilv 进行绝对定量, 使其减少到2个副本, 因此对亚临床病例中的 tilv 诊断特别有用。对两种 pcr 方法的详细说明、两个实验室的代表性结果以及对这两种方法的关键参数的深入讨论, 都包括在内, 以确保研究人员和诊断人员找到最适合和最适用的方法tilv 检测方法。
引言
2014年, 全球人均鱼类供应量达到20公斤的新纪录, 这是因为水产养殖强劲增长。水产养殖仍然是全世界增长最快的动物食品生产部门之一, 也是唯一增长快于人类的动物食品生产部门1。tilapiine cichilds 是全世界养殖的第二大淡水鱼, 全球总产量为640万吨, 2015年估计价值98亿美元 2.罗非鱼产量最大的十大生产国是中国 (1.78 吨)、印度尼西亚 (1.78 吨) 和埃及 (0.88 吨), 其次是孟加拉国、越南、菲律宾、巴西、泰国、哥伦比亚和乌干达.预计到 2030年, 全球罗非鱼产量将在7.3 吨左右.罗非鱼已成为如此重要的全球食物来源, 不仅因为它们是一种廉价的蛋白质来源 4, 而且还因为它们在广泛的水和气候条件下很容易繁殖 5,6。就在几十年前, 人们认为, 威胁罗非鱼养殖的具有商业意义的疾病很少, 但这已经不是事实了。一种新出现的病毒性疾病叫罗非鱼湖病毒病 (tilvd), 是罗非鱼有史以来发现的首例严重疾病流行, 整个行业都面临风险。这一疾病造成严重的社会经济后果, 对非洲7、亚洲和南美洲数百万人的粮食安全构成直接威胁。2018年初, 世界动物卫生组织 (动物卫生组织) 报告说, 在八大洲8个国家正式发现了这种疾病的病原药 tilv, 自这张病原体信息卡以来 ,更新有更多的报告, tilv 在坦桑尼亚, 乌干达9, 印度尼西亚10, 台湾11和秘鲁12.tilv 是一种新型的单链 rna 病毒, 被描述为类似正霉菌的病毒, 因为它包含各种特征, 让人想起其他正生瘤病毒, 如流感或传染性三文鱼-贫血病毒 (isav)13。它最早是在以色列加利利14湖野生和养殖罗非鱼大规模损失后确定的.此后, 埃及15例尼罗河罗非鱼和尼罗河和红色杂交罗非鱼报告了类似的疾病爆发, 称为夏季死亡和与 tilv 感染有关的一个月死亡率综合征(奥雷诺克利斯)分别在泰国16。水生动物病毒的检测历史上是通过细胞培养中病毒的生长和分离来进行的。对不同的细胞系进行了 tilv 的繁殖和分离测试, 其中包括来自奥雷奥科勒克利斯的17、18、omb 和 tmb 中提取的 e-11 细胞 。蚊子 18岁, 翁布和翁拉原产于尼罗河罗非鱼 (o. niloticus)19。虽然病毒培养的优点是它为进一步的实验提供了材料, 但它的缺点是, 它至少需要4-7 的时间来观察细胞病理效应 (cpe) 的形成, 关键是不同的鱼氨酸病毒, 这些病毒更适合复制可能会传播, 并产生类似的 cpe。
在过去的几十年里, 已经从传统的、往往耗时的诊断方法, 如细胞培养、血清学和抗原检测, 并替换为更快和更敏感的核酸检测测试20, 21岁这一点是显而易见的, 许多 qpcr 检测已开发成为重要的诊断方法, 在水生动物的多种病毒性疾病, 如 isav22,23, 病毒出血性败血症病毒 (vhsv)24 , 25, β-病毒26,27沙粒甲型六病毒 28, 鱼虹膜病毒29, 安吉利德疱疹病毒 1 (anghv1) 30, 和淋巴细胞疾病病毒 (lcdv)31.迫切需要可靠的诊断和病原体监测方法, 以减少 tilv 的传播。这种方法应允许在临床体征发展之前及早发现感染, 并检测低病毒负荷。迄今为止, 已经开发了不同的 pcr协议,包括 rt-pcr 14、32、嵌套 rt-pcr18、半嵌套的 rt-pcr 33 和 rt-qpcr32、34, 用于检测 tilv在鱼组织中。通过对用于 tilv 检测的易感细胞系中的 rt-qpcr 和病毒分离的比较, 发现 rt-qpcr 的敏感性是病毒分离32 的 1, 000倍。尽管每个已发布的 pcr 协议都报告了对 tilv 检测的不同敏感性, 但大多数检测方法都具有高度敏感性, 病毒拷贝的检测限值为 7.5份, 样本份为 33, 7 份为 18 或2份拷贝, 每份 32份反应。
本文的目的是详细解释如何进行 tilv 检测检测, 从罗非鱼组织采集开始, 到总 rna 提取、cdna 合成, 再到 tilv 特异性 pcr 检测。具体而言, 已经描述了常规 rt-pcr 和 sybr 绿色的 rt-qpcr 的综合协议, 以吸引广泛的科学家, 旨在检测 tilv。前者不太敏感, 但通常是一种更便宜的检测选项。后者需要更精细的基础设施, 如定量 pcr 机器和更昂贵的试剂, 但它的优点是定量、快速和高度敏感, 这意味着它可用于亚临床检测 tilv受感染的鱼。rt-pcr 和 rt-qpcr 协议是在两个不同的实验室进行的, 这些实验室具有不同的地理分离株 tilv, 其中的结果突出了这里描述的检测的敏感性和重现性。
研究方案
这项研究的动物使用协议是由卡塞萨特大学动物伦理委员会根据许可证号 acku 59-vet-016 批准的。
注: 有关本协议所建议的试剂和设备的详细信息, 请参阅材料表。
1. 组织样品采集
- 使用过量的丁香油对鱼类进行安乐死 (体积取决于鱼的大小和产品的浓度, 通常超过 3 mll)。将四分之一的钳子和蛋黄酱浸入 95% (vv) 乙醇中, 然后使用酒精燃烧器燃烧设备对设备进行消毒。
注: 可以使用三联安咪唑磺酸 (ms-222) 代替丁香油。 - 找到肝脏, 切断一小块 (约20-100 毫克) 或收集200μl 的粘液使用盖板玻璃或手术刀片, 以去除粘液从前至后罗非鱼, 并将样品放入 1.5 ml 微离心管。
- 立即处理样品, 储存在 rna 稳定溶液中, 或将其移动到-80°c, 直到进一步使用。
注: 使用 rna 的最大任务是制备完整的 rna 分子, 并在以后的任何处理过程中保持它们不受损害。rna 主干天生比 dna 对损伤更敏感。从组织细胞中提取和分离总 rna 需要仔细的实验室技术;采取所有规定, 以防止 rnase 污染, 戴手套, 使用 rnase 免费水, 试剂, 设备, 塑料制品, 玻璃器皿, 工作空间和使用过滤器提示进行移液。
2. 硫氰酸酯-苯酚-氯仿 rna 提取
- 在含有第1节组织样本的试管中加入1毫升含有苯酚和鸟嘌呤异硫氰酸酯的单相溶液。
注意: 此解决方案毒性很强, 必须小心地在层状流动罩中, 配备防护设备, 并佩戴适当的防护眼镜、衣物和安全手套。 - 使用组织孔均质机研磨组织样品, 直到均匀。
注: 样品也可以使用与陶瓷珠结合的功率均质机进行均质化。确保组织样品完全同质化, 然后再继续下一步或在这里停止协议, 并将完全均质样品保存在-80°c, 直到进一步使用。 - 加入200μl 氯仿进行相分离。
氯仿是一种潜在的麻醉剂, 极其危险。应小心地在带防护设备的层流罩中处理, 并佩戴适当的防护眼镜、衣服和安全手套。作为一种毒性较小的替代品, 也可以使用 1-溴-3-氯丙烷。
注: 在适当的情况下向上或向下扩展卷。例如, 如果只使用含有苯酚和鸟嘌呤异硫氰酸酯的500μl 单相溶液, 则在此步骤中只添加100μl 的氯仿。- 通过反转15秒很好地混合样品。
- 在室温 (rt) 下对样品进行3分钟的孵化。
- 在 12, 000 xg和4°c 下离心15分钟。
注: 应该有一个明确的分离到一个较低的有机相, 一个白色的间相和含有 rna 的上水相。这个顶部的阶段通常是无色的, 但根据同质组织的类型和数量, 它可以有一个浅粉色的外观。 - 将上水层 (约 500μl) 转移到新鲜的微离心管, 而不会干扰中间期。
注: 不要试图转移整个水相, 留下少量, 以防止任何潜在的污染 rna 含有水相与有机或中间相。 - 加入1体积100% 异丙醇沉淀 rna。
- 或者, 如果使用了极少量的组织, 则在每个样本中添加 1μl (5-10μg) 的无 rnase-糖原, 以促进有效的 rna 沉淀。这将有助于在步骤2.8 中识别 rna 颗粒。
注: 糖原起到 rna 载体的作用, 并将防止少量 rna 粘附在试管的一侧。 - 通过多次倒置将管道混合好。
- 将样品存放2小时至夜间-20°c。
- 或者, 如果使用了极少量的组织, 则在每个样本中添加 1μl (5-10μg) 的无 rnase-糖原, 以促进有效的 rna 沉淀。这将有助于在步骤2.8 中识别 rna 颗粒。
- 在 12, 000 x g和4°c 下离心样品15分钟。
- 丢弃上清液, 注意不要将微离心管底部的 rna 颗粒移出。
- 加入1% 毫升的75% 乙醇 (v/v), 并通过多次倒置试管混合 rna 样品。
- 在 10, 000 x g和4°c 下离心15分钟。
注: 该协议可以在此停止, 包含75% 乙醇中 rna 颗粒的样品可以储存在-20°c, 直到进一步使用。 - 丢弃上清液, 注意不要将微离心管底部的 rna 颗粒移出。
- (可选) 使用70% 乙醇 (v/v) 重复步骤 2.9-2.11。彻底清洗 rna 颗粒将最大限度地减少任何可能干扰敏感下游应用的盐或污染物结转。
- 用移液器取出剩余的乙醇, 然后在室温下风干 rna 颗粒, 时间不超过5至10分钟。
注: 过度干燥的颗粒将很难重新悬浮。 - 加入30-60μl 的无 rnase 水, 预热至 55-60°c, 使 rna 颗粒溶解。
- 将 rna 放在冰上立即使用, 或存放在-80°c, 供以后使用。
3. 使用微体积分光光度计对 rna 浓度进行定量
- 将分光光度计的设置切换到 rna。
- 使用1-2 微米的无 rnase 水作为空白。
- 使用每个 rna 样本的1-2 μl 来评估 rna 的数量。
- 记录每个样本的读数为230纳米、260纳米和280纳米。
- 使用无 rnase 的水将 rna 稀释至200ngμμl。
4. 使用总 rna 合成互补 dna (cdna)
- 将 2, 2μm 寡核苷酸 (dt)、0.5 mm dntp 混合物中的总 rna 1 微克与无核酸酶水混合, 使最终体积达到10μl。为此, 请根据要测试的样品和控件的数量准备 rt 母版组合。
注: 这些控制是一个微小的逆转录酶样本 (-rt), 其中 rt 酶被无核酸酶水 (见步骤 4.3) 和无模板控制 (ntc), 其中无核酸酶的水被添加到主混合而不是 rna 模板。- 通过移液将样品混合均匀, 然后进行短暂的离心。
- 在65°c 孵育样品 5分钟, 然后在冰上孵育2分钟。
- 对样品进行简单的离心, 以收集管内底部的所有液体。
- 加入1x 逆转录酶缓冲液, 100 u 逆转录酶, 并使用无核酸酶的水, 将每个样品的最终体积提高到20μl。
- 通过移液将样品混合均匀, 然后进行短暂的离心。
- 将样品在42°c 下培养 60分钟, 然后在85°c 下培养5分钟。
- 通过添加适当数量的无核酸水将合成的 cdna 稀释到所需的浓度, 并将 cdna 放在冰上立即使用, 或将其储存在-20°c, 供以后使用。
5. 常规 pcr
- 使用协议第4节中生成的 cdna、样本和控件作为 pcr 反应模板, 使用表 1中详述的任何既定引物对, 以及所选的 dna 聚合酶。
注: 在 pcr 反应中, 应通过用 cdna 代替无核酸水, 在这里加入额外的无模板控制 (ntc)。如果有的话, 也应包括一个阳性对照, 包括先前核实的 tilv 阳性样本或克隆到质粒中的适当的 tilv cdna 片段。 - 根据所使用的 dna 聚合酶系统的指南以及要测试的样品和控制的数量, 准备 pcr 主混音。这种组合应包括正向引物、反向底漆、dntp、mgcl2和选定的 dna 聚合酶及其缓冲液。
- 根据所选 dna 聚合酶的指导原则, 将指定的主混合物体积与建议的 cdna 样本和对照样本量结合起来。
注: 准备0.5倍反应过量往往是有益的, 因为一些主混料在移液过程中丢失。 - 根据所使用的 dna 聚合酶系统的指导原则, 对使用中的引物使用适当的退火温度, 执行 pcr 循环条件 (表 1)。通常情况下, 这样的程序将涉及在95°c 下2-5 的初始变性 2-5, 然后是在95°c 下的30-40 变性周期 30秒, 在建议温度下退火 30秒, 在72°c 下的伸长率为 30秒, 最后的伸长率为72°c 为5-10 最小值。
- 根据预期的 pcr 产品大小, 将每个 pcr 反应和适当的 dna 阶梯加载到1-2% 琼脂糖凝胶的井中。通过凝胶电泳分离扩增的 dna, 并通过溴化乙酯 (ethidium) 染色凝胶, 以便于使用紫外线在凝胶文档机中显示预期大小的 dna 带 (表 1)。
注意: etbr 是有毒的;应小心处理, 并戴上适当的防护服和安全手套。
| 目标 tilv 基因组段 | 正向底漆 5 '-3 ' | 反向底漆 5 '-3 ' | pcr 产品尺寸 (bp) | tm °c | 原始参考 | ||||
| 1 | ccaacttcttattataccac | gcaattcttctccct | 1641年 | 50 | surachetpong 等人, 2017年 | ||||
| 1 | cctcttttgttaagt | agagtgtgtggttag | 1000元 | 62 | mgimba 等人, 2018年 | ||||
| 2 | actcttattacacattattact | ttactattataggaggc | 1445 | 45 | surachetpong 等人, 2017年 | ||||
| 2 | gtccgggggtgttatg | cttactgcagctctaag | 834 | 62 | mgimba 等人, 2018年 | ||||
| 3个 | tgggcacagcatcta | taccagggtactctgtgt | 250人 | 56 | eyngor 等人, 2014年 | ||||
| 3个 | taggtatcccccccc | ttctctgagagtacacc | 491 | 57 | eyngor 等人, 2014年 | ||||
| 3个 | accccattactaacgtta | ccatatttattaggtgt | 1352年 | 50 | surachetpong 等人, 2017年 | ||||
| 3个 | gtggggcattaccagtaag | gagtaagggggcttg | 834 | 62 | mgimba 等人, 2018年 | ||||
| 4个 | agcagcagagagag | acccttttctgagg | 358 | 60 | nicolson 等人, 2017年 | ||||
| 4个 | ccaagtttattactaccaga | gcaatctccattcct | 1250 | 50 | surachetpong 等人, 2017年 | ||||
| 4个 | gccaatgtcccattct | gccaatgtcccattct | 524 | 62 | mgimba 等人, 2018年 | ||||
| 5 | ccaatttcttttctcacactc | cttttctttcattttag | 1087 | 57 | surachetpong 等人, 2017年 | ||||
| 5 | catcttcctggaatac | ctttcctcctggtaca | 696 | 62 | mgimba 等人, 2018年 | ||||
| 6 | ccaattttcccccat | tcafactaactacc | 1027 | 45 | surachetpong 等人, 2017年 | ||||
| 6 | ccacacacacaggacadag | gagtggttagggggtaaga | 948 | 62 | mgimba 等人, 2018年 | ||||
| 7。 | cttttcattatccit | gacatattccctctctca | 704 | 57 | surachetpong 等人, 2017年 | ||||
| 7。 | tccttagggattggcactaac | ttccatcactccctaga | 486 | 62 | mgimba 等人, 2018年 | ||||
| 8 | 阿卡卡卡塔卡茨卡 | tcatatacatacagggct | 637 | 50 | surachetpong 等人, 2017年 | ||||
| 8 | cttaaggcccctgtcatc | tgcacaatccacactaa | 476 | 62 | mgimba 等人, 2018年 | ||||
| 9 | ttggggatgacgagata | agtttattcccccatg | 351 | 60 | nicolson 等人, 2017年 | ||||
| 9 | acagtgattttccc | tttccccttaagtca | 530 | 50 | surachetpong 等人, 2017年 | ||||
| 9 | gattccacacacccccttc | gtac gcactgccattac | 261 | 62 | mgimba 等人, 2018年 | ||||
| 10 | aaccctacacacacacatagct | cttccccccccccct | 450元 | 50 | surachetpong 等人, 2017年 | ||||
| 10 | tcctctctctctcttt | cagagggggcafatt | 276 | 62 | mgimba 等人, 2018年 | ||||
表 1.已发布的引物对, 用于利用端点 pcr 扩增 tilv cdna。以粗体显示的引物集用于生成图 3 a 和 3 b 所示的代表性结果。
6. tilv 定量聚合酶链反应 (qpcr)
- 以含有适当的 tilv 基因组段 3 cdna 的质粒为标准, 如 ptilv32,准备一个复制或三联的10倍系列稀释。
- 为所有样品、标准和控制准备 qpcr 母版混合物, 同时考虑到反应应使用0.4μl 的无核酸酶水、0.3μl 的正向底漆、0.3μl 的反向底漆和5μl 的 2x sybr 绿色进行一式或三式三份的反应每个反应的 dna 聚合酶母模混合。
- 在浓度为10μm 的情况下使用引物, 引物信息和标准的 ptilv 如下:
正向底漆: tilv-12f (5 '-ctgagtaagtaagtaatgatggattt-3 ')
反向底漆: tilv-112r (5 '-cggggtactttagtagttctt-3 ')
标准 pllv:10 pg/μl
注: 如果样品和对照的总数为 10个, 并将以三元组合进行, 这相当于 qpcr 主混结构, 包括12μl 无核酸酶水、9μl 正向底漆、9μl 反向底漆和 150μl sybr 绿色 dna 聚合主混合。商业购买的2x 通用 sybr 绿色 dna 聚合酶母版混合包含 qpcr 反应所需的所有成分, 即 sybr 绿色 i 染料、热启动 taq dna 聚合酶、dntp、mgcl2和被动参考染料。保护 sybr 绿色主混音不受光线影响。
- 在浓度为10μm 的情况下使用引物, 引物信息和标准的 ptilv 如下:
- 将 qpcr 母管的6μl 混合分配到 qpcr 带管或与使用中的 qpcr 机器兼容的96孔板中。
- 在96井板的管或井中加入4μl 的 cdna 模板, 控制或连续稀释 tilv 标准。
- 关闭 qpcr 管或密封96井板与兼容的板盖的 qpcr 机器在使用中
- 轻轻轻弹 qpcr 管, 混合溶液, 然后用离心机将 qpcr 管或96井板旋转下来, 收集容器底部的所有液体。
- 将管材或板材放入实时热循环器中。
- 将 qpcr 热环器编程在95°c 下进行3分钟的初始变性, 然后在65°c 的温度下进行40次周期的 10秒, 60°c 的70°c 循环, 用于底漆的退火和伸长率, 最终以65°c 至95°c 的熔融曲线步长结束, 增量为0.5°c/5是的。
- 选择 sybr 作为荧光染料, 然后选择未知作为样本类型, 并在示例名称框中插入名称。
- 打开 rt-qpcr 机的盖子, 将 qpcr 条带放入指定的井中, 然后关闭盖子。
- 在选定的条件下进行 rt-qpcr 检测。盖子达到所需温度后, 机器将开始运转。在每个扩展步骤后收集每个样品的荧光, 以监测反应的进展。
注: qpcr 机器和相关软件将自动计算检测的所有参数, 并实时显示扩增曲线, 同时在 qpcr 周期结束时生成标准曲线和熔融曲线。 - 首先进行数据分析和采集, 确保每个样品和标准的熔融曲线在放大器的预期温度下具有一个均匀的峰值。
- 在所有样品中, cDNAs 的放大率都是相同的, 因此对样品和标准的放大曲线进行了评价, 并在一个区域内设定了阈值。这通常由软件自动执行, 但应仔细检查。
- 使用标准曲线计算 tilv 副本的数量。
结果
按照第1节所述的协议, 显示 tilv 感染临床症状的垂死的红色杂交罗非鱼 (图 1a) 被安乐死, 在高浓度的丁香油中沐浴, 作为麻醉剂。报告的临床症状是可变的, 但常见的症状似乎是嗜睡, 皮肤侵蚀和变色, 眼球突出, 分离鳞片, 开放的伤口/病变和异常行为15,16,33, 35,36, 其中一些可以清楚地看到在图 1a.腹腔壁被切除, 以收集肝脏、脾脏或头颅肾等内脏 (图 1b)。在此阶段, 还通过使用盖板玻璃或手术刀片37轻轻刮伤鱼的前后部皮肤, 采集粘液样本.

图 1.罗非鱼解剖和样品采集.a. 受 tiv 感染的红色杂交罗非鱼, 有皮肤, 口腔和小孔周围发红, 皮肤侵蚀和角膜混浊。b. 分离的红色杂交罗非鱼, 以允许从肝脏 (在蓝色箭头点), 脾脏或头肾脏器官的组织收集。请点击这里查看此图的较大版本.
{kind=link}
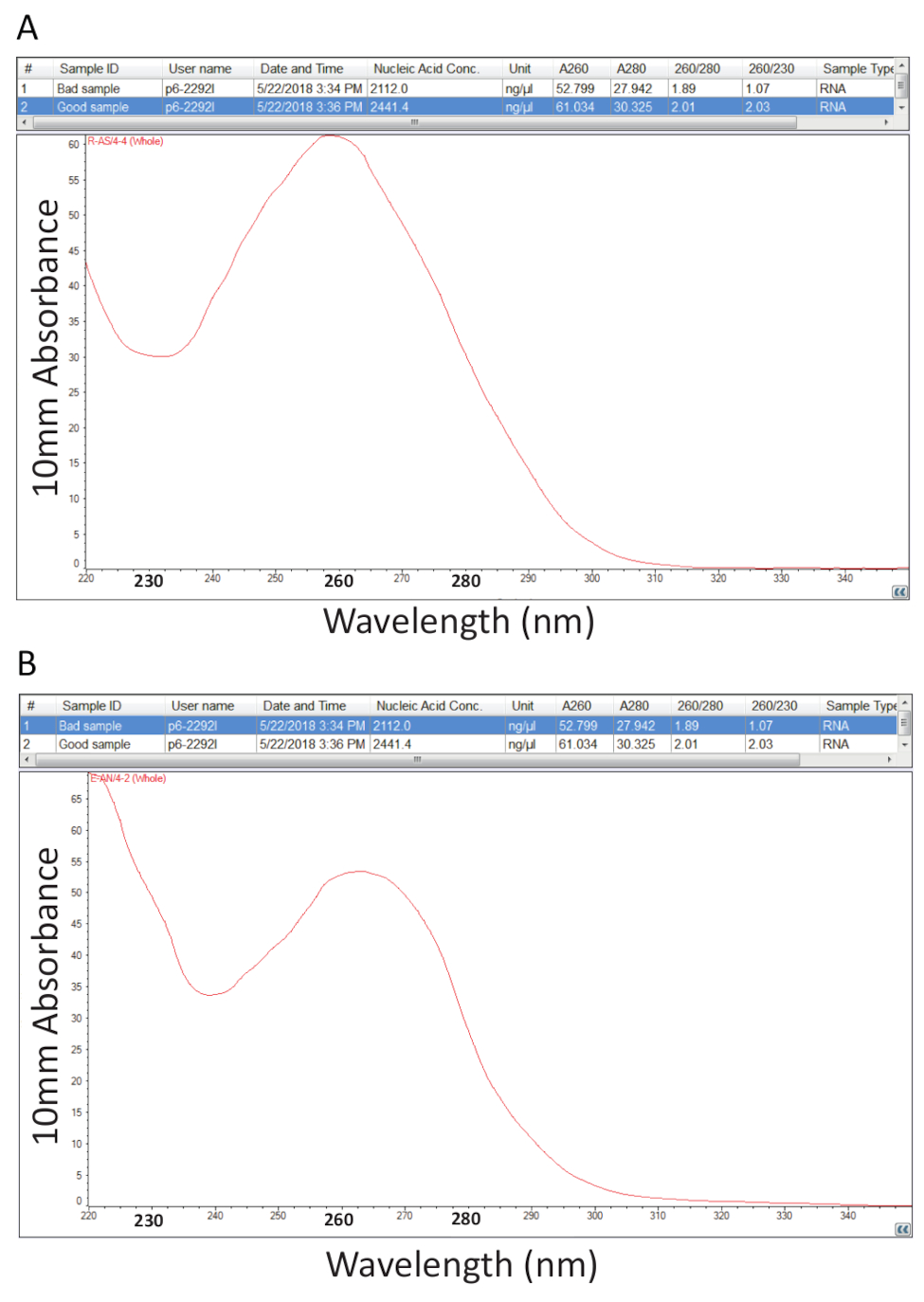
此后, 遵循了第2节详述的关于总 rna 的硫氰酸酯-氯仿提取的协议, 并通过计算纯度比和光谱剖面的检测 (图 2)。图 2a显示了成功的总 rna 提取过程的代表性结果, 而图 2A则表示 rna 制备不良。核酸在260时具有吸收最大值, 而蛋白质在280纳米时具有吸收最大值。测量值为260纳米和280纳米, 表明每个样品的纯度和1.9 至2.1 的比率表明纯 rna, 如图 2 a 中的样品。图 2b中观察到的较低 a260/280 比率表明 rna 提取过程中可能剩余的蛋白质或苯酚污染。在230纳米的吸收可以是样品污染的结果, 并计算了 a26/230 纳米比的原因。对于纯 rna 制剂, 这一比率应在2.0-2.2 的范围内,如图 2 a中的样本的2.03 所示,而图2A 的a260/230 比较低, 为 1.07, 光谱剖面显示低谷在230纳米向240的转变 纳米, 表明样品中残留的鸟嘌呤或苯酚。对于图2b 所示的样品, 重新沉淀 rna 以去除污染可能会提高样品的纯度。

图 2.从病变罗非鱼组织中提取的总 rna 的分光光度定量。a. 成功的 rna 制备的纯度比率和光谱分布。b. 作为 a, 除了代表一个糟糕的 rna 提取程序。请点击这里查看此图的较大版本.
{kind=link}
为了用 rt-pcr 检测 tilv,图 2 a中所述的纯样本被逆转录酶 (协议 4) 逆转录酶转化为 cdna, 并作为第5节详述的 pcr 检测模板, 如图 3 a 所示。表 1中的粗体引物被用来放大 tilv 基因组段的 491 bp 片段.用凝胶电泳分离 pcr 产物, 用 etbr 染色, 进行可视化。图 3a显示了使用 4个 cdna 样本 (s1-s4) 进行两步 rt-pcr 的结果, 这些样本来自泰国分离的病罗非鱼肝脏, 在每个样本中, 都可以观察到大约 500 bp 的清洁单波段, 因此, 样本1-4 是 tilv积极。相同的 pcr 产物是从阳性对照样品中获得的, 其中包括克隆到质粒32 中的 tilv 片段3的 cdna, 而无模板控制 (ntc) 不产生 pcr 产物。图 3b中的检测方法使用与图 3 a相同的引物, 但在不同的实验室中, 使用一步 rt-pcr 方法, 并从源自埃及水产养殖的罗非鱼头肾组织中提取 5个 rna 样本15. 利用该检测方法确定, 样品1、3和5的 tilv 呈阳性, 而样本2和4为 tilv 阴性, 因为没有发现大小正确的 pcr 产物。阴性对照, 包括两个负逆转录酶对照和两个 ntc 没有产生任何 pcr 产物。针对罗非鱼放线菌基因, 还进行了一步 rt-pcr 检测。每个样本 (s1-s5) 中的放大器大小为 217 bp, 如预期的 38。该检测方法可作为 rna 样品完整性的控制, 并允许对 tilv 阳性样品进行半定量检测。鉴于生成的 Tilapia actb 产品相对相等, 则生成的 tilv 特异性 pcr 产品数量的差异可以解释为在给定组织样本中真正反映了 tilv 的含量。

图 3.tilv rt-pcr。从泰国采集的病罗非鱼肝脏组织产生的 cdna 样本, 用2步 rt-pcr 法, 用 tilv 第3段 (粗体显示在表 1中) 的特异性引物进行 tilv 感染筛查。m = 标记以基对显示;s1-s4 = 样品 1-4;c1 = 使用 ptlv 作为 pcr 模板的阳性对照;和 c2 = 无模板控制 (ntc)。b. 使用与 a 中相同的引物和从埃及采集的病罗非鱼头肾组织样本的一步 rt-pcr 15.m = 标记以基对显示;s1-s5 = 样品1-5。控制 c1-c2 是负逆转录酶对照, c3-c4 是 ntc. 底部面板是一个单步 rt-pcr 使用针对罗非鱼 actinb38 的引物 (详见文本 ) 产生217基对的 pcr 产品。请点击这里查看此图的较大版本.
{kind=link}
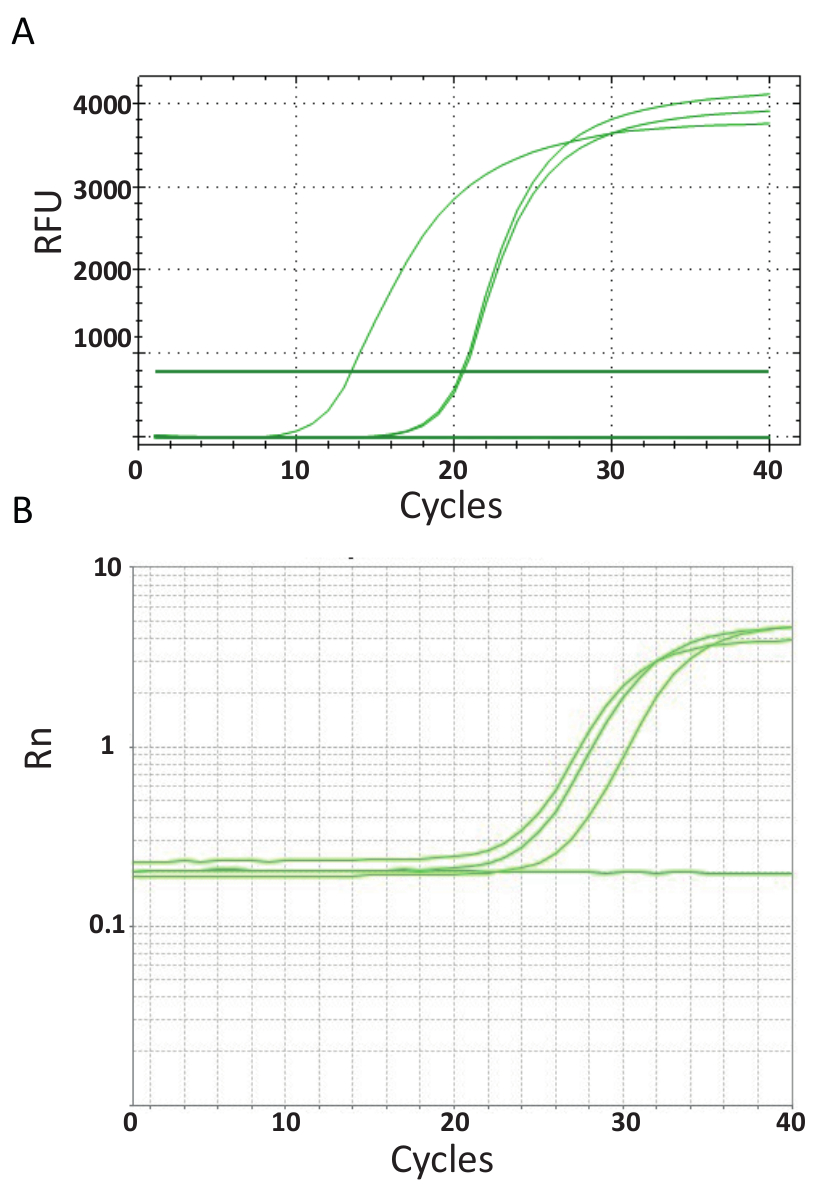
与图 3中所示的端点 pcr 不同, 协议6中解释的 qpcr 检测在每个 pcr 周期后测量 pcr 产物的数量。目标 dna 的扩增是通过荧光分子与每一轮反应产生的 dna 相互作用的。在这里, 使用了 sybr 绿色 i 染料, 它与双链 dna 交织在一起。在反应过程中跟随荧光信号, 其强度与形成的产品数量为 39、40、41、42、43.tilv qpcr 检测是按照第6号议定书所述在不同的实验室使用不同的 sybr 绿色试剂、qpcr 机器和来自不同国家的样品进行的。由此产生的放大曲线如图 4 a 和4 b所示。可以看出, 对于每一个检测, 实验过程有四个阶段: 线性地相、早期指数相、晚期指数相和高原相。线性地相发生在早期周期, 由于 dna 数量不充分而无法确定 dna 重复, 而 dna 重复产生的信号与背景比不足。在此阶段计算基线荧光。此后, 目标 dna 开始加倍集中, 每个周期诱导信号变得可检测到的背景以上, 并呈指数级增长。优化后的 qpcr 检测方法的扩增效率 (e) 在反应开始时非常高 (接近 100%), 并且在扩增的早期指数阶段保持稳定, 此时进行量化, 当反应效率仍然稳定。在以后的周期中, 信号开始趋于平稳, 荧光的强度不再与起始模板复制号相关, 因为反应成分已耗尽44。由于再退火反应、组分浓度比的变化或酶单位与 dna 底物分子的数量的竞争, 也可能发生饱和。这些参数可能会解释图 4 a 和4 b所示的检测放大曲线之间的差异。包含的控件没有产生这些特征放大曲线。

图 4.扩增图显示产品在实时 pcr 检测期间的积累情况。a. 从泰国获得的 tiv 阳性样品的扩增曲线、ntc 和使用 sybr-gre-grei 2 步 qpcr 法的正质粒控制。图表是通过绘制相对荧光 (rfu) 与循环数生成的。b. 从埃及获得的 tilv 阳性样品的扩增曲线, 如图 3b和 ntc 所示。扩增曲线是将报告信号归一化为分析 (rn) 中的被动 rox 染料相对于循环数的荧光的荧光.请点击这里查看此图的较大版本.
{kind=link}
在每个实验室的不同机器上进行 qpcr 热循环结束时, 对数据进行了采集和分析。图 5a和5A 显示了在每个实验室进行的检测的典型熔融曲线。每台 qpcr 机器都经过编程, 在最后进行熔融曲线分析。这是通过增量增加温度和监测荧光作为温度的函数来实现的。当温度足够高, 足以使 dsdna 变性时, 由于荧光分子被释放, 荧光会有很大的下降。每个 qpcr 仪器的软件通过绘制负一阶导数与温度来计算熔融曲线数据中的退火温度 (tm) (图 5)。可以看出, 在图 5a和5A 中, 在不同样品集中形成的产品在约80°c 的预期温度下具有均匀的熔融转变。在较低的温度下没有观察到其他峰值。由于其体积小, 引物二聚体的 tm 通常低于目标 dna 序列的 t m。因此, tm s之间的这种差异使得识别潜在的引物二聚体或其他非特异性放大产品变得很容易。这些控件没有生成像 tilv 阳性样本和标准这样的熔体曲线, 可以看作是图 5a和5A图表底部的一条几乎水平线。

图 5.熔融曲线分析, 以确保检测特异性和不同的 pcr 产品可以通过其熔融特征进行鉴别。泰国tilv 阳性样品的熔融曲线分析、阴性对照和正质粒控制。b. 从埃及获得的 tilv 阳性样品的熔融曲线分析、ptilv 标准和 ntc。a 和 b 中的图表都显示了荧光的变化与根据温度绘制的温度变化的变化, 从而清晰地反映了熔融动力学。请点击这里查看此图的较大版本.
{kind=link}
大多数 qpcr 机器都配备了一个软件, 便于进一步评估 qpcr 运行情况, 并将通过自动绘制针对 ptilv 标准模板对数的周期阈值 (ct)来生成标准曲线来量化样本两个独立实验室的副本编号, 如图 6a和6A所示。简单地说, ct 是用于评估 qpcr 结果的单位。ct 值表示达到设定阈值荧光信号电平所需的周期数。启动模板的数量越大, 达到可检测到的荧光水平所需的周期就越少。事实上, 具有高负载的 tilv 的样品将具有较低的 ct 值, 而不是具有较低负载的 tilv 的样品, 例如在具有亚临床感染的鱼类中。为了确定 ct 值, 首先从原始数据中扣除背景荧光水平。接下来, 与 qpcr 设备关联的软件将通过搜索每个样本的数据曲线并合并一个 ct 表示样本跨越阈值的位置来自动选择荧光阈值。这是为每个检测单独进行的, 每个阈值都应仔细评估, 确保阈值已在放大曲线的对数部分和所有曲线平行的位置设置。因此, 获得的特定 ct 是一个相对值, 它是相对于起始模板拷贝编号45, 但它也是特定的 qpcr 机器和使用的试剂, pcr 扩增的效率和检测的敏感性。这些参数有助于使用图 6中相同的分析观察到的差异。
从图 6中的标准曲线来看, 进行了回归分析, 包括标准曲线斜率 (m) 和拦截的计算、放大效率 (100 x (10-1 ))和反应的线性度。还使用了标准曲线分析来确认检测的灵敏度 (检测极限)、重复性和重现性。从理论上讲, 每个 pcr 周期的 dna 量都增加了一倍, 这意味着效率 (e) 等于100%。然而, 在实践中, 由于 dna 聚合酶抑制、污染物、过多的 cdna 和移液错误47等 pcr 条件,这种理想的效率很少达到。通常情况下, 扩增 e 的范围从90-110% 的良好检测, 在图6a 中, 94.5% 的效率是使用8个连续稀释的 ptilv 样品计算的, 而在图 6A中使用7连续稀释的 ptilv 的检测效率样本为101.2。效率超过100% 通常是由于检测中存在 pcr 抑制剂。标准图的线性回归分析还允许计算每个样本中的 tilv 副本数量 41、42、45, 如图6b 中红色显示的三个 tilv 样本所示这与图 3b 所示的样本 s1、s3 和 s5 的结果一致。

图 6.rt-qpcr 标准曲线。双系列 ptilv 的10倍连续稀释的实时 pcr, 这两个实验室都使用标准。对 a. 8个连续稀释的 ptilv 样品进行了测试, 所有已知浓度均与 tilv 铜/反应的数量相关。标准曲线是通过绘制日志副本号与周期阈值 (ct) 生成的。斜率 =-3.462, r2 = 0.9992, 效率为94.47。b. 与 a 一样, 测试了7个连续稀释的 ptilv 样品 (绿色), 图表在 y 轴上显示阈值周期, 在 x 轴上显示 tilv (数量) 的复制编号。y-截距 = 32.327, 斜率 =-3.292, r2 = 0.98, 效率为101.2。对于 a 和 b 中的两个标准曲线, 利用斜率、y 截距和相关系数值(r2) 来了解检测的性能。重要的是,r2值应接近 1, 因为它是标准曲线线性度的度量。斜率用于测量 pcr 效率, 其中100% 效率对应于-3.32 的斜率, 请参见主要文本的方程和进一步的细节。良好的 qpcr 反应的效率一般在与-3.58 和-3.58 之间的斜率相关的 90%-110% 之间。标准曲线用于对未知的 tilv 阳性样品进行绝对定量, 并确定 tilv 拷贝/反应的确切数量, 如在 b 中着色为红色的三个 tilv 阳性样品。
讨论
tilv 于2014年首次在以色列报告,自那时以来, 已在多个国家发现, 包括埃及、哥伦比亚、印度、马来西亚、乌干达、坦桑尼亚和泰国15、16、18、35,48. 全球认识, 特别是罗非鱼生产国的认识, 更加关注该病毒, 政府当局实施了各种限制和控制措施, 试图防止 tilv 的传播。本文详细介绍了罗非鱼组织中的 tilv 检测方案, 包括样本采集、rna 分离、cdna 合成、pcr 和 qpcr 检测。这些方法有各个方面值得具体讨论。到目前为止, 在大小为 9、12、14、15、49的鱼类中已经发现了 tilv 和罗非鱼品种, 包括养殖的杂交罗非鱼(o. o。11,14, 尼罗河罗非鱼 ( o. niloticus)9,10, 14, 15, 16,33,35,36,49,50和红色罗非鱼 (oro虫夏子sp.)16,33,48, 51,以及在野生尼罗河罗非鱼9,12, 黑色罗非鱼51, t. zilli14,15, s.galilaeus, o. aureus 和 t. simonis中间体 14 , 最近在野生鲤鱼 (barbonyus) 中发现了 tilv斯瓦南费迪)52。来自内脏器官 (刺、脾、肝、心、头肾) 或粘液37的组织样本可以从健康和垂死的罗非鱼中采集, 而不论其年龄、大小或物种如何, 并进行 rna 分离处理。这里概述的总 rna 提取协议使用苯酚和硫氰酸 guanidinium 的单相溶液, 它是一种各向异性变性剂。组织在这种溶液中直接同质化, 然后加入氯仿和离心, 以实现相分离, 其中含有上水相、间相和较低的有机相的透明 rna。通过异丙醇沉淀法从水相中分离 rna, 然后清洗回收的 rna 以去除污染物。采用这种方法分离 rna 是由 piotr chomczynski 和 nicoletta sacchi 首创的, 被称为 guanidinium 硫氰酸酯-苯酚-氯仿提取物53,54。这种类型的试剂用于 rna 提取可以商业购买或在实验室制造 (见材料表的进一步信息)。与硅基纯化等柱基方法相比, 该协议所需的时间稍长, 但一般来说, 它更具成本效益, 产生更多的 rna。
在该协议中, 概述了使用 a260 值定量 rna 的方法, 即分光光度法值可以指示 rna 质量 (a260\ a280 = 1.9-2.1)。虽然这种方法将提供一个很好的样品纯度的指示, 它不能绝对告知提取的 rna 的质量。为了正确地确定 rRNA 是否完整或部分降解, 样品可以通过琼脂糖凝胶电泳分离, 其中 etbr 染色18s 和 28s rrna 带的涂抹表明 rRNA 降解。rna 质量的进一步验证可能包括使用片上实验室仪器。此外, 用 dnase i 消化纯化的 rna 以去除污染宿主基因组 dna 也很重要, 这取决于下游应用可能会导致错误的结果。如果宿主 gdna 仍在很大程度上污染 rna 样品, 也可以在 rna 提取过程结束时进行额外的 dnasei 处理 (见材料表)。
互补 dna 合成可以极大地影响整个 qpcr 结果, 是该方法的一个方面, 可以引入变异。这里提倡的 cdna 协议包括使用寡核苷酸 (dt) 的单个组件设置, 因此只转录含有多 a 尾的 mrna。它允许用户控制在逆转录酶中究竟使用哪些成分, 这种 cdna 合成模式已被证明是成功的 tilv 检测 32.这种设置的另一种选择是商业购买的主混料, 其中包含逆转录酶反应所需的所有组件, 并且非常快速和简单, 无需通常的多步骤、移液和多温度协议。这是有利的, 因为它最大限度地减少了处理, 并促进了所有样品的一致性。这种主混物通常包括寡核苷酸 (dt) 和随机引物, 使其适用于不同的 rna 模板, 并从种群中的整个 rna 长度 (病毒和罗非鱼宿主 mrna) 生成具有代表性的 cdna 序列副本, 理论上,然后, 可以通过常规 pcr 或 qpcr 从这样的样本中测量每个所需的 rna 物种。这种多功能性是两步 rt-pcr 方法的主要优点;它提供了一个长期的池, 可以用于许多不同的实验。结果表明, 采用了一步 rt-pcr 方法, 其中使用了序列特异性引物 (表 1), 并在一个管中进行了 rt 和 pcr (见材料清单)。一般来说, 序列特定引物比使用随机启动更高的特定目标 rna rna rt 效率, 但特定目标 rna 是唯一可以在这样的 cdna 样本中量化的, 而这可能是某些实验室的唯一目的 (见cdna 合成产品的材料表)。
虽然到目前为止, 常规的 rt-pcr 似乎在 tilv 诊断中普遍使用,13,14,15,16,17, 18,33,35,48,55. rt-qpcr 已被证明是检测和定量鱼类组织或粘液中少量 tilv 的更有力工具 32、37。qpcr 以其灵敏度高、特异性强、重现性好、动态范围宽、速度21等优点, 在临床病毒学诊断实验室中得到了广泛的应用。虽然 qpcr 最初的实施成本可能高于传统的 rt-pcr, 但与传统的 pcr 相比, 它确实提供了许多重要的优势;它具有更快的周转时间从样品到结果, 它不需要任何 pcr 后步骤。后一点意味着实验室污染的风险最小, 它可以更容易地适应高通量的情况, 例如在爆发的情况下。此外, 它本质上比传统的 rt-pcr 更敏感, 它对于检测亚临床感染21中的低病毒载量至关重要。这将需要一个嵌套的 pcr 方法, 需要逆转录酶链转录, 两个进一步的 pcr 反应, 然后通过琼脂糖凝胶电泳分析。这些许多步骤需要大量的时间, 并增加错误或污染的机会。尽管如此, 由于其高灵敏度, rt-qpcr 要求细致的实验设计和对定量技术的透彻理解, 以产生精确的结果56,57。
该协议演示了 dna 结合荧光体 sybr green i。它是一种 dsdna 非特异性 dna 结合染料, 因此该检测的特异性完全在于引物集, 这可能会产生假阳性58。因此, 虽然在每次 pcr 结束时进行的 dsdna 熔融曲线分析是 pcr 反应的一个特别重要的部分, 因为它证实只产生了一个正确的 t m pcr 放大器 (这也应该通过凝胶来实现)在实施新的检测时进行电泳)。dna 片段的 tm 取决于多种特征, 如长度、气相色谱组成、序列、链互补、浓度以及缓冲成分和 pcr 增强剂。两个实验室在有代表性的结果中进行的熔融曲线分析没有发现初级二聚体或其他不需要的 pcr 产物的存在, 但如果用其他样品和实验装置观察到这一点, 那么检测结果应该是重新优化。更先进的 qpcr 技术不需要这样的熔融曲线步骤, 事实上, 自从编写了这篇论文以来, 利用两个引物和一个探针开发了一个基于 taqman 的 tilv rt-qpcr, 使其具有高度的 tilv 特异性34。
毫无疑问, 为 rt-qpcr 检测设计的引物是检测成功的基础, 这里的引物是根据当时公开的 tilv 基因组数据设计的。然而, rna 病毒是众所周知的表现出很高的突变率和可能的菌株将逃脱目前的诊断测试, 正如观察到 isav59.这类病毒类型将始终难以生成通用的泛 tilv rt-qpcr 检测方法, 只有在有更多来自影响深远的位置和时间段的 tilv 基因组数据可用的情况下, 此类检测才会不断得到改进。
最后, 在 qpcr 内和 qpcr 内检测中进行重复的或在可能的情况下运行三文三联反应是非常重要的。如果 ct 值非常高, 则使用复制物对于确保 pcr 反应是可靠和可重现的尤为重要。一般情况下, 如果复制反应的数据变化超过0.5个周期, 则应重复反应, 如果复制中的 ct 值持续变化 & gt; 0.5个周期, 则应重新优化检测结果。使用集成的 qpcr 移液机器人对解决这个问题有很大的帮助, 但它是一个豪华的工具。与任何实验一样, 纳入适当和适当的控制对于开发强大的分子检测至关重要, 特别是在必须认可此类检测的诊断实验室。控制应包括阳性 (阳性 tilv 样本、tilv 质粒标准) 和阴性对照 (ntc 和-rt) 样本, 以及内源性罗非鱼内务基因的检测。这种控制不可低估, 应包括在每个检测中, 以正确理解每个步骤的质量, 并正确解释结果。
披露声明
作者没有什么可透露的。
致谢
我们感谢伯尔尼大学 vetsuisse 学院兽医细菌学研究所的支持。这项工作由伯尔尼大学威特苏塞学院早期职业研究人员和两性平等学术促进委员会资助, 向 pn 提供了120% 的示范资金。ws 和 pr 由泰国曼谷 kasetsart 大学农业和粮食高级研究中心、高级研究所、泰国高等教育研究促进和国家研究大学项目提供支持, 办公室泰国教育部高等教育委员会主席。我们要感谢 kwanrawee sirikanchana 博士的叙述和 piyawatchara sikarin 编辑的视频。
材料
| Name | Company | Catalog Number | Comments |
| Tissue collection | Step 1 | ||
| Tricaine methanesulfonate | Sigma-Aldrich | E10521 | An alternative to clove oil. Step 1.1 |
| RNAlater stabilization solution | Thermo Fisher Scientific | AM7020 | For storing tissues if they cannot be processed immediately Step 1.3 |
| RNA extraction | Step 2 | ||
| TRIreagent | Sigma-Aldrich | Step 2.1 | |
| TRIzol | Thermo Fisher Scientific (Invitrogen) | 15596026 | Step 2.1 |
| GENEzol | Geneaid | GZR100 | Step 2.1 |
| Trisure | Bioline | BIO-38032 | Step 2.1 |
| Homemade solution | - | - | 94.53 g/L (800 mM) guanidine thiocyanate 30.45 g/L (400 mM) ammonium thiocyanate 8.20 g/L (100 mM) sodium acetate 380 mL/L (38 % v/v) phenol 50 mL/L (5 % v/v) glycerol 1.0 g/L (0.1 % w/v) 8-quinolinol, pH 5.0 Store up to 2 years at 4oC Step 2.1 |
| MagNA Lyser Green Beads | Roche | 3358941001 | An alternative tissue homogenization method used in conjunction with tissue lysing machines detailed below Step 2.2 |
| Lysing Matrix D, 2 mL Tube | MP BIOMEDICALS | 116913050 | |
| Chloroform | Sigma-Aldrich | C2432 | Step 2.3 |
| Chloroform | RCI Labscan | AR1027E-G2.5L | Step 2.3 |
| 1-Bromo-3-chloropropane | Sigma-Aldrich | B9673 | A less toxic alternative to chloroform Step 2.3 |
| Isopropanol (GC) ≥ 99.8 % | Sigma-Aldrich | 59300 | Step 2.6 |
| Isopropanol (ACS, ISO Reag. Ph Eur) | Merck KGaA (EMSURE) | 1.09634.2500 | Step 2.6 |
| Glycogen, molecular biology grade (e.g., Sigma, cat. no. G1767) | Thermo Fisher Scientific (Thermo Scientific) | R0551 | Useful step if tissue starting material is small to maximise RNA precipitation optional |
| Ethanol (purity (GC) ≥ 99.9 % | Sigma-Aldrich (EMD Millipore) | 1.00983 | Step 2.9 |
| Ethanol (ACS, ISO Reag. Ph Eur) | Merck (EMSURE) | 1.00983.2500 | Step 2.9 |
| Nuclease-free water | Promega | P1193 | Step 2.13 |
| Nuclease-free water | Multicell | 809-115-CL | Step 2.13 |
| Ambion TURBO DNA-free kit | Thermo Fisher Scientific (Invitrogen) | AM1907 | Can be performed at the end of the RNA extraction protocol optional |
| cDNA synthesis | Step 4 | ||
| Viva cDNA Synthesis Kit | Vivantis | cDSK01 | Step 4.1 & 4.3 |
| ReverTra Ace qPCR RT MasterMix with gDNA remover | Toyobo | A1172K | An alternative option see discussion |
| ReverTra Ace qPCR RT Kit | Toyobo | FSQ-101 | An alternative option see discussion |
| AffinityScript Multiple Temperature Reverse Transcriptase | Agilent Technologies | 600107 | An alternative option |
| PCR | Step 5 | ||
| DNA polymerase systems: | Step 5.2 | ||
| - Platinum II Hot-Start Green PCR Master Mix (2X) | Thermo Fisher Scientific (Invitrogen) | 14001012a | Step 5.2 |
| - GoTaq Mastermix | Promega | M7122 | Step 5.2 |
| Separate PCR mixture components: | Step 5.2 | ||
| 10mM dNTP Mix | Vivantis | NP2409 | Step 5.2 |
| 25mM MgCl2 | Thermo Fisher Scientific | R0971 | Step 5.2 |
| 10X Taq Buffer with KCl | Thermo Fisher Scientific | 00348114 | Step 5.2 |
| Taq DNA polymerase | Vivantis | PL1202 | Step 5.2 |
| - Verso 1-step RT-PCR ReddyMix with ThermoPrime Taq | Thermo Fisher Scientific | AB1454 | One step RT-PCR exemplified in Figure 3B |
| Gel electrophoresis: | For visulation of PCR products from steps 5.1-5.4 | ||
| Ethidium Bromide solution (10 mg/mL) | Thermo Fisher Scientific | 17898 | Step 5.5 |
| Tris/Acetic/EDTA (TAE) buffer: | Step 5.5 | ||
| - Tris | Vivantis | PR0612-1KG | Step 5.5 |
| - Acetic acid (glacial) (ACS, ISO, Reag. Ph Eur) | Merck KGaA (EMSURE) | 1.00063.2500 | Step 5.5 |
| - Ethylenediaminetetraacetic acid (EDTA) | BIO-RAD | 161-0729 | Step 5.5 |
| Agarose | Vivantis | PC0701-100G | Step 5.5 |
| DNA ladders and markers | Vivantis | NL1405 | Step 5.5 |
| DNA gel loading dye (6X) | Thermo Fisher Scientific | R0611 | Step 5.5 |
| qPCR | Step 6 | ||
| PowerUP SYBR Green Master Mix | Thermo Fisher Scientific (Applied Biosystems) | A25779 | Exemplified in Figures 4-6B Step 6.2 |
| iTaq Universal SYBR Green Supermix | BIO-RAD | 1725120 | Exemplified in the video and in Figures 4-6A Step 6.2 |
| Equipment | |||
| Dounce tissue grinder pestle | Sigma-Aldrich | P1110 | Protocol 2 |
| MagNA Lyser Instrument | Roche | 3358976001 | An alternative tissue homogenizing option for protocol 2 which are used in conjunction with the lysing beads detailed above Step 2.2 |
| FastPrep-24 5G Homogenizer | MP BIOMEDICALS | 116005500 | |
| Refrigerated microcentrifuge | Eppendorf | Eppendorf 5427R | Protocol 2 Step 2.4, 2.7 & 2.10 |
| Refrigerated microcentrifuge | Eppendorf | Eppendorf 5418R | |
| Heat box | Labnet | AccuBlock Digital Dry Bath | Protocol 2 Step 2.13 |
| Microvolume spectrophotometer | Thermo Fisher Scientific (Applied Biosystems) | Nanodrop 2000 | Protocol 3 Step 3.1 - 3.4 |
| PCR machine | BIO-RAD | T100 Thermal Cycler | Protocol 5 Step 5.4 |
| Power supply | BIO-RAD | PowerPac HC | Protocol 5 Step 5.5 |
| Horizontal gel electrophoresis | BIO-RAD | Mini ReadySub-Cell GT Cell #1704487edu | Protocol 5 Step 5.5 |
| Mini microcentrifuge | Corning | LSE 6766 | Useful to quickly spin down PCR reaction tubes in protocols 4, 5 & 6 Step 6.5.1 |
| Microcentrifuge | LioFuge | LM-60 | Step 6.5.1 |
| qPCR machine and software | Thermo Fisher Scientific | 7500 Fast Real-Time PCR System with 7500 Software v2.0 | Protocol 6 Step 6.6-6.8 |
| qPCR machine and software | BIO-RAD | CFX96 Touch Real-Time PCR Detection System with CFX Manager software | |
| General Materials | |||
| Mayo scissors | Step 1.1-1.2 | ||
| Forceps | Step 1.1-1.2 | ||
| Pipette | Rainin | Pipette-Lite XLS | |
| Aerosol-barrier pipette tips | Sigma-Aldrich | Z333328, Z333336, Z333344 | |
| Nuclease-free 1.5-ml microcentrifuge tubes | Eppendorf |
参考文献
- FAO. . The State of World Fisheries and Aquaculture, 2014. Opportunities and Challenges. , (2014).
- FAO. . The State of World Fisheries and Aquaculture, 2016. Contributing to Food Security and Nutrition for all. , (2016).
- WorldBank. . FISH TO 2030: Prospects for Fisheries and Aquaculture. Agriculture and Environmental Services Discussion Paper 03. , (2013).
- Wing-Keong, N., Nicholas, R. A review of the nutrition and feeding management of farmed tilapia throughout the culture cycle. Reviews in Aquaculture. 5 (4), 220-254 (2013).
- Cleasby, N., et al. The socio-economic context for improving food security through land based aquaculture in Solomon Islands: A peri-urban case study. Marine Policy. 45, 89-97 (2014).
- Ponzoni Raul, W., et al. Genetic improvement of Nile tilapia (Oreochromis niloticus) with special reference to the work conducted by the WorldFish Center with the GIFT strain. Reviews in Aquaculture. 3 (1), 27-41 (2011).
- Hounmanou, Y. M. G., et al. Tilapia lake virus threatens tilapiines farming and food security: Socio-economic challenges and preventive measures in Sub-Saharan Africa. Aquaculture (Amsterdam, Netherlands). 493, 123-129 (2018).
- OIE. . Tilapia Lake Virus (TiLV) - a novel orthomyxo-like virus. OIE technical disease cards. , (2018).
- Mugimba, K. K., et al. Detection of tilapia lake virus (TiLV) infection by PCR in farmed and wild Nile tilapia (Oreochromis niloticus) from Lake Victoria. Journal of Fish Diseases. , (2018).
- Koesharyani, I., Gardenia, L., Widowati, Z., Khumaira, D. D., Rustianti, Studi kasus infeksi tilapia lake virus (tilv) pada ikan nila (Oreochromis niloticus). Jurnal Riset Akuakultur. 13 (1), 85-92 (2018).
- OIE. . Tilapia lake virus disease (TiLV), Chinese Taipei. Immediate Notification. , (2017).
- OIE. . Tilapia Lake Virus Disease (TiLV), Peru. Immediate Notification. , (2018).
- Bacharach, E., et al. Characterization of a Novel Orthomyxo-like Virus Causing Mass Die-Offs of Tilapia. MBio. 7 (2), e00431-e00416 (2016).
- Eyngor, M., et al. Identification of a novel RNA virus lethal to tilapia. Journal of Clinical Microbiology. 52 (12), 4137-4146 (2014).
- Nicholson, P., et al. Detection of Tilapia Lake Virus in Egyptian fish farms experiencing high mortalities in 2015. Journal of Fish Diseases. 40 (12), 1925-1928 (2017).
- Surachetpong, W., et al. Outbreaks of Tilapia Lake Virus Infection, Thailand, 2015-2016. Emerging Infectious Diseases. 23 (6), 1031-1033 (2017).
- Tattiyapong, P., Dachavichitlead, W., Surachetpong, W. Experimental infection of Tilapia Lake Virus (TiLV) in Nile tilapia (Oreochromis niloticus) and red tilapia (Oreochromis spp.). Veterinary Microbiology. 207, 170-177 (2017).
- Kembou Tsofack, J. E., et al. Detection of Tilapia Lake Virus in Clinical Samples by Culturing and Nested Reverse Transcription-PCR. Journal of Clinical Microbiology. 55 (3), 759-767 (2017).
- Thangaraj, R. S., et al. Derivation of two tilapia (Oreochromis niloticus) cell lines for efficient propagation of Tilapia Lake Virus (TiLV). Aquaculture (Amsterdam, Netherlands). 492, 206-214 (2018).
- Hanson, L. A., Rudis, M. R., Vasquez-Lee, M., Montgomery, R. D. A broadly applicable method to characterize large DNA viruses and adenoviruses based on the DNA polymerase gene. Virology Journal. 3, 28-28 (2006).
- Josko, D. Molecular virology in the clinical laboratory. Clinical Laboratory Science. 23 (4), 231-236 (2010).
- Munir, K., Kibenge, F. S. Detection of infectious salmon anaemia virus by real-time RT-PCR. Journal of Virological Methods. 117 (1), 37-47 (2004).
- Snow, M., et al. Developement, application and validation of a Taqman real-time RT-PCR assay for the detection of infectious salmon anaemia virus (ISAV) in Atlantic salmon (Salmo salar). Developments in Biologicals. 126, 133-145 (2006).
- Matejusova, I., McKay, P., McBeath, A. J., Collet, B., Snow, M. Development of a sensitive and controlled real-time RT-PCR assay for viral haemorrhagic septicaemia virus (VHSV) in marine salmonid aquaculture. Diseases of Aquatic Organisms. 80 (2), 137-144 (2008).
- Garver, K. A., et al. Development and validation of a reverse transcription quantitative PCR for universal detection of viral hemorrhagic septicemia virus. Diseases of Aquatic Organisms. 95 (2), 97-112 (2011).
- Dalla Valle, L., et al. Development of a sensitive and quantitative diagnostic assay for fish nervous necrosis virus based on two-target real-time PCR. Veterinary Microbiology. 110 (3-4), 167-179 (2005).
- Hodneland, K., Garcia, R., Balbuena, J. A., Zarza, C., Fouz, B. Real-time RT-PCR detection of betanodavirus in naturally and experimentally infected fish from Spain. Journal of Fish Diseases. 34 (3), 189-202 (2011).
- Hodneland, K., Endresen, C. Sensitive and specific detection of Salmonid alphavirus using real-time PCR (TaqMan). Journal of Virological Methods. 131 (2), 184-192 (2006).
- Wang, X. W., Ao, J. Q., Li, Q. G., Chen, X. H. Quantitative detection of a marine fish iridovirus isolated from large yellow croaker, Pseudosciaena crocea, using a molecular beacon. Journal of Virological Methods. 133 (1), 76-81 (2006).
- van Beurden, S. J., et al. Development and validation of a real-time PCR assay for the detection of anguillid herpesvirus 1. Journal of Fish Diseases. 39 (1), 95-104 (2016).
- Ciulli, S., et al. Development and application of a real-time PCR assay for the detection and quantitation of lymphocystis disease virus. Journal of Virological Methods. 213, 164-173 (2015).
- Tattiyapong, P., Sirikanchana, K., Surachetpong, W. Development and validation of a reverse transcription quantitative polymerase chain reaction for tilapia lake virus detection in clinical samples and experimentally challenged fish. Journal of Fish Diseases. 41 (2), 255-261 (2018).
- Dong, H. T., et al. Emergence of tilapia lake virus in Thailand and an alternative semi-nested RT-PCR for detection. Aquaculture (Amsterdam, Netherlands). 476, 111-118 (2017).
- Waiyamitra, P., et al. A TaqMan RT-qPCR assay for tilapia lake virus (TiLV) detection in tilapia. Aquaculture (Amsterdam, Netherlands). 497, 184-188 (2018).
- Behera, B. K., et al. Emergence of Tilapia Lake Virus associated with mortalities of farmed Nile Tilapia Oreochromis niloticus (Linnaeus 1758) in India. Aquaculture (Amsterdam, Netherlands). 484, 168-174 (2018).
- Ferguson, H. W., et al. Syncytial hepatitis of farmed tilapia, Oreochromis niloticus (L.): a case report. Journal of Fish Diseases. 37 (6), 583-589 (2014).
- Liamnimitr, P., Thammatorn, W., U-thoomporn, S., Tattiyapong, P., Surachetpong, W. Non-lethal sampling for Tilapia Lake Virus detection by RT-qPCR and cell culture. Aquaculture (Amsterdam, Netherlands). 486, 75-80 (2018).
- Yang, C. G., et al. Evaluation of reference genes for quantitative real-time RT-PCR analysis of gene expression in Nile tilapia (Oreochromis niloticus). Gene. 527 (1), 183-192 (2013).
- Bustin, S. A. Real-time, fluorescence-based quantitative PCR: a snapshot of current procedures and preferences. Expert Review of Molecular Diagnostics. 5 (4), 493-498 (2005).
- Fleige, S., Pfaffl, M. W. RNA integrity and the effect on the real-time qRT-PCR performance. Molecular Aspects of Medicine. 27 (2-3), 126-139 (2006).
- Kubista, M., et al. The real-time polymerase chain reaction. Molecular Aspects of Medicine. 27 (2-3), 95-125 (2006).
- Mackay, I. M., Arden, K. E., Nitsche, A. Real-time PCR in virology. Nucleic Acids Research. 30 (6), 1292-1305 (2002).
- Wong, M. L., Medrano, J. F. Real-time PCR for mRNA quantitation. Biotechniques. 39 (1), 75-85 (2005).
- Bustin, S. A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. Journal of Molecular Endocrinology. 25 (2), 169-193 (2000).
- Heid, C. A., Stevens, J., Livak, K. J., Williams, P. M. Real time quantitative PCR. Genome Research. 6 (10), 986-994 (1996).
- Rutledge, R. G., Côté, C. Mathematics of quantitative kinetic PCR and the application of standard curves. Nucleic Acids Research. 31 (16), e93-e93 (2003).
- Svec, D., Tichopad, A., Novosadova, V., Pfaffl, M. W., Kubista, M. How good is a PCR efficiency estimate: Recommendations for precise and robust qPCR efficiency assessments. Biomolecular Detection and Quantification. 3, 9-16 (2015).
- Amal, M. N. A., et al. A case of natural co-infection of Tilapia Lake Virus and Aeromonas veronii in a Malaysian red hybrid tilapia (Oreochromis niloticus × O. mossambicus) farm experiencing high mortality. Aquaculture (Amsterdam, Netherlands). 485, 12-16 (2018).
- Fathi, M., et al. Identification of Tilapia Lake Virus in Egypt in Nile tilapia affected by ‘summer mortality’ syndrome. Aquaculture (Amsterdam, Netherlands). 473, 430-432 (2017).
- OIE. . Tilapia Lake Virus disease (TiLV), Philippines. Immediate Notification. , (2017).
- OIE. . Tilapia lake virus disease (TiLV), Malaysia. Immediate Notification. , (2017).
- Abdullah, A., et al. First detection of tilapia lake virus (TiLV) in wild river carp (Barbonymus schwanenfeldii) at Timah Tasoh Lake, Malaysia. Journal of Fish Diseases. 41 (9), 1459-1462 (2018).
- Chomczynski, P., Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Analytical Biochemistry. 162 (1), 156-159 (1987).
- Chomczynski, P., Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: twenty-something years on. Nature Protocols. 1 (2), 581-585 (2006).
- Del-Pozo, J., et al. Syncytial Hepatitis of Tilapia ( Oreochromis niloticus L.) is Associated With Orthomyxovirus-Like Virions in Hepatocytes. Veterinary Pathology. 54 (1), 164-170 (2017).
- Bustin, S. A., et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clinical Chemistry. 55 (4), 611-622 (2009).
- Purcell, M. K., Getchell, R. G., McClure, C. A., Garver, K. A. Quantitative polymerase chain reaction (PCR) for detection of aquatic animal pathogens in a diagnostic laboratory setting. Journal of Aquatic Animal Health. 23 (3), 148-161 (2011).
- Simpson, D. A., Feeney, S., Boyle, C., Stitt, A. W. Retinal VEGF mRNA measured by SYBR green I fluorescence: A versatile approach to quantitative PCR. Molecular Vision. 6, 178-183 (2000).
- Kibenge, M. J., et al. Discovery of variant infectious salmon anaemia virus (ISAV) of European genotype in British Columbia, Canada. Virology Journal. 13, 3 (2016).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。