
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
测量体外分化的离体小鼠脂肪组织和原代前脂肪细胞的脂肪分解率
摘要
脂肪细胞中的甘油三酯脂肪分解是导致游离脂肪酸和甘油释放的重要代谢过程。在这里,我们提供了一个详细的方案来测量小鼠脂肪细胞和 离体 脂肪组织中的基础和刺激脂肪分解。
摘要
脂肪细胞以甘油三酯的形式在脂滴中储存能量。这种能量可以通过脂解 动员 ,其中脂肪酸侧链从甘油骨架上依次裂解,导致游离脂肪酸和甘油的释放。由于甘油激酶在白色脂肪细胞中的低表达,甘油再摄取率可以忽略不计,而脂肪酸再摄取由白蛋白等培养基成分的脂肪酸结合能力决定。甘油和脂肪酸释放到培养基中都可以通过比色测定法进行定量,以确定脂解速率。通过在多个时间点测量这些因素,可以高置信度地确定脂肪分解的线性速率。在这里,我们提供了一个详细的方案,用于测量来自小鼠的体 外 分化脂肪细胞和 离体 脂肪组织中的脂肪分解。该协议也可以针对其他前脂肪细胞系或来自其他生物体的脂肪组织进行优化;讨论了考虑因素和优化参数。该协议旨在用于确定和比较小鼠模型和治疗之间的脂肪细胞脂肪分解率。
引言
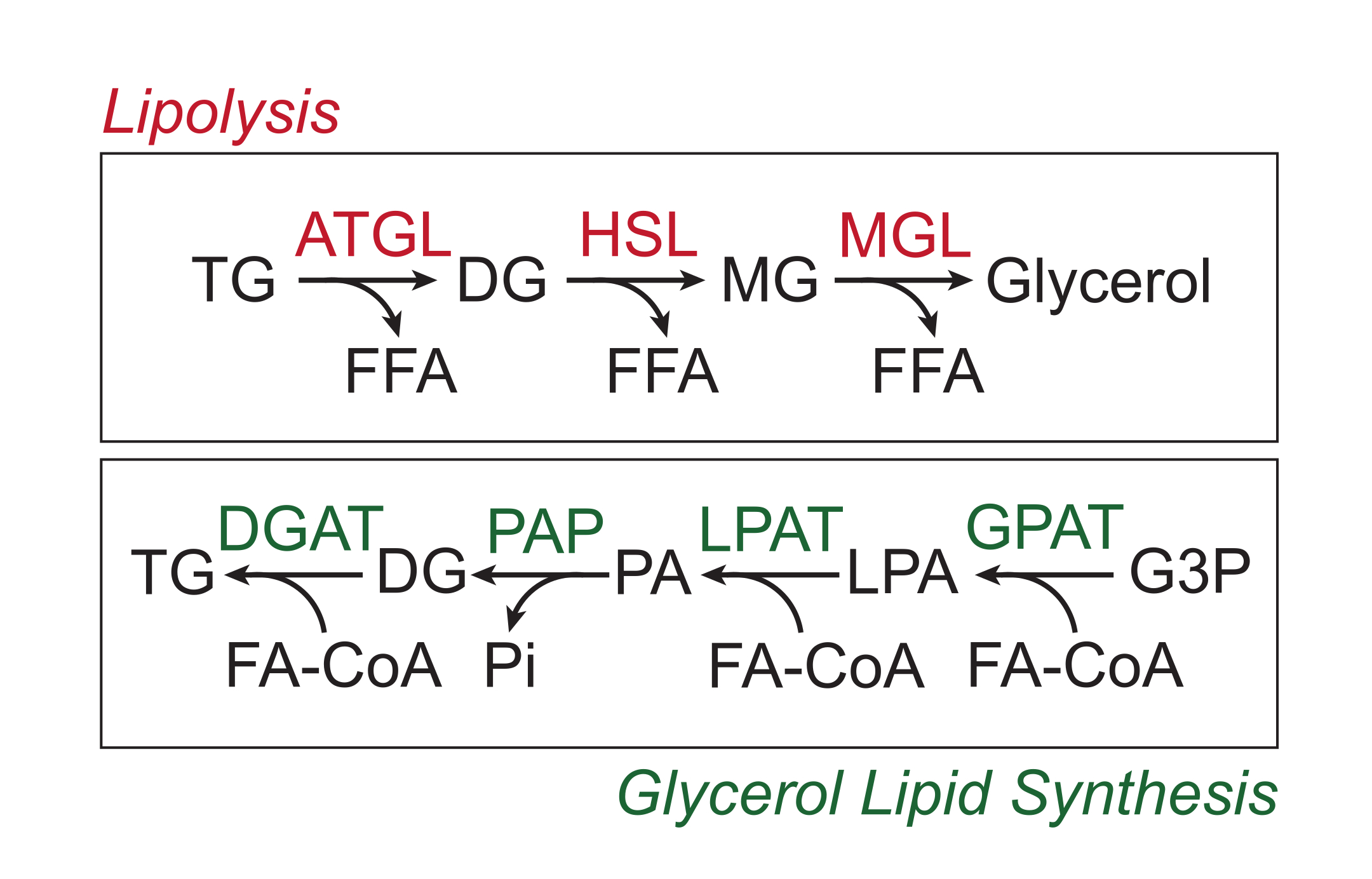
多余的营养物质以甘油三酯的形式储存在脂滴的中性脂质核心中的白色脂肪组织中。甘油三酯储存通过脂解动员,脂肪分解是脂肪酸侧链被脂肪组织甘油三酯脂肪酶(ATGL),激素敏感脂肪酶(HSL)和甘油单酯脂肪酶(MGL)顺序切割的过程,导致游离脂肪酸(FFA)和甘油骨架的释放1,2。脂肪分解由脂肪组织中的儿茶酚胺信号激活。交感神经末梢局部释放儿茶酚胺,儿茶酚胺与脂肪细胞质膜上的β-肾上腺素能受体结合。配体结合后,这些G蛋白偶联受体(GPCR)通过Gαs激活腺苷酸环化酶。随后通过cAMP激活蛋白激酶A(PKA)导致ATGL和HSL的上调。PKA对perilipin-1的磷酸化导致ABHD5(也称为CGI-58)的解离,其结合并共激活ATGL3。PKA直接磷酸化HSL,促进其从细胞质质到脂滴的易位,其中与磷酸化的perilipin-1相互作用进一步促进其脂肪酶活性4,5,6,7。参与脂解的第三种脂肪酶MGL似乎不受儿茶酚胺信号传导的调节8。重要的是,脂肪细胞中的甘油三酯合成是由甘油脂质合成途径介导的,其不涉及甘油单酯作为中间体的形成;相反,甘油-3-磷酸酰基转移酶催化溶血磷脂酸的形成,溶血磷脂酸与另一种脂肪酰基辅酶A结合形成磷脂酸,然后在最终合成甘油三酯之前异构化为甘油二酯(图1)9,10,11。

图1:脂解和甘油脂质合成途径。 上图:脂解通路;红色显示的酶:脂肪组织甘油三酯脂肪酶(ATGL),激素敏感脂肪酶(HSL)和甘油单酯脂肪酶(MGL)。下图:甘油脂质合成途径;绿色显示的酶:甘油二酰基转移酶 (DGAT)、磷脂酸磷酸酶 (PAP)、溶血磷脂酸酰基转移酶 (LPAT,也称为 LPAAT) 和甘油-3-磷酸酰基转移酶 (GPAT)。脂质:甘油三酯 (TG)、甘油二酯 (DG)、甘油单酯 (MG)、游离脂肪酸 (FFA)、脂肪酰辅酶 A (FA-CoA)、溶血磷脂酸 (LPA) 和磷脂酸 (PA)。其他代谢物:无机磷酸盐(Pi)和甘油3-磷酸(G3P)。 请点击此处查看此图的大图。
{kind=link}
细胞外腺苷是脂肪分解的另一种重要调节因子,通过Gs和Gi偶联GPCR影响腺苷酸环化酶活性。脂肪细胞中的主要腺苷受体ADORA1抑制腺苷酸环化酶,从而通过激活Gi12来抑制脂肪分解。ADORA2A以较低水平表达,主要在棕色脂肪细胞中表达,通过Gs信号传导激活脂肪分解13。ADORA1影响基础脂肪分解和对肾上腺素能激动剂的反应。腺苷对脂肪分解的作用可以通过添加腺苷脱氨酶来中和腺苷,以及ADORA1特异性激动剂苯基异丙基腺苷14,15来控制。Gq偶联GPCR的激素激活也可以通过磷脂酶C和蛋白激酶C16,17,18,19的活化来影响脂肪分解。炎症信号也会影响脂解率。LPS(和其他内毒素)的TLR4激活通过激活ERK来增加脂解率,ERK磷酸化perilipin-1和HSL20。TNF-α还通过ERK和NF-κB活化激活脂肪分解,以及磷酸二酯酶PDE-3B和CIDEC21,22,23的转录下调。IL-6 也与脂肪细胞脂肪分解增加有关,特别是在肠系膜脂肪组织中,其 FFA 释放影响肝脂肪变性和糖异生24,25,26。
脂肪分解在进食状态下被胰岛素抑制。AKT 磷酸化并激活 PDE-3B 以抑制 cAMP 信号传导并阻止 PKA 活化27。胰岛素也转录下调ATGL28。肥胖通过多种机制促进儿茶酚胺耐药性,包括下调脂肪细胞29,30,31,32,33中的β-肾上腺素能受体。脂肪细胞表达所有三种β-肾上腺素能受体(β-1、β-2 和 β-3)。虽然β-1和β-2肾上腺素能受体普遍表达,但β-3肾上腺素能受体主要在小鼠34,35的脂肪细胞中表达。Adrb3 表达由 C/EBPα 在脂肪生成36 期间诱导。β-3肾上腺素能受体在成熟脂肪细胞中高表达。由于β-arrestin37的反馈抑制,β-1和β-2肾上腺素能受体的激活是自限性的。β-3肾上腺素能受体的反馈抑制由其他信号通路介导,其降低Adrb3表达33,38,39。
许多化合物可用于激活脂肪细胞脂质分解。儿茶酚胺是脂解的主要生理激活剂。去甲肾上腺素(或去甲肾上腺素)和肾上腺素(或肾上腺素)激活所有三种β肾上腺素能受体40。去甲肾上腺素和肾上腺素也通过激活α-肾上腺素能受体信号传导 来 影响脂肪分解41。常用的β-肾上腺素能受体激动剂包括异丙肾上腺素能激动剂,它是一种非选择性β-肾上腺素能受体激动剂,以及β-3肾上腺素能受体激动剂CL-316,243和米拉贝隆42。鉴于脂肪细胞主要表达β-3肾上腺素能受体,我们在这里使用CL-316,243作为一个例子。它对β-3肾上腺素能受体的特异性也使其成为脂肪细胞儿茶酚胺信号传导的相对特异性激活剂,也可以安全地用于 体内。请注意,细胞培养物中常用的10μM CL-316,243浓度比达到最大反应所需的~0.1μM剂量高几个数量级33。佛司可林绕过肾上腺素能受体, 直接激活腺苷酸环化酶和下游脂解信号.还有更多的激活剂,以及脂肪分解的抑制剂。在选择刺激脂解的化合物时,应在实验设计中仔细考虑受体特异性和下游信号通路。
白色脂肪组织中脂肪分解的速度是影响禁食或运动期间耐寒性和营养可用性的重要代谢因素43,44,45,46。该协议的目的是测量脂肪细胞和脂肪组织中脂肪分解的速率,这将有助于了解脂肪细胞代谢以及它如何影响各种小鼠模型的代谢表型。为了量化脂解率,我们测量了脂解产物在培养基中的外观(即FFA和甘油)。该方法依赖于脂解产物从脂肪细胞释放到培养基中。由于白色脂肪细胞表达低水平的甘油激酶,因此甘油再摄取率较低47。相反,还应考虑通过脂肪分解以外的代谢途径产生FFA和甘油。脂肪细胞似乎表达一种具有抗甘油-3磷酸活性的磷酸酶,使得能够从葡萄糖48,49,50衍生的甘油-3-磷酸产生甘油。糖酵解是用于白色脂肪细胞中FFA再酯化的甘油-3-磷酸的来源。当葡萄糖水平有限时,甘油新生需要其他3碳来源,如乳酸和丙酮酸51。细胞内脂肪分解释放的FFA的通道及其代谢命运知之甚少;脂肪分解释放的FFA必须在再酯化或经历β氧化之前转化为脂肪酰基辅酶A。看来脂肪分解释放的FFAs可能会在被带回并转化为脂肪酰辅酶A 52,53,54,55,56,57,58,59,60,61,62之前离开细胞.FFA可以通过白蛋白隔离在细胞外。重要的是,已知长链FFAs如果不被白蛋白63,64,65,66,67隔离,则反馈抑制脂肪分解。因此,在脂解测定过程中优化培养基的FFA缓冲能力至关重要。这里描述的程序类似于先前发表的测量小鼠和人类脂肪细胞和离体脂肪组织中脂解率的方法15,68,69,70,71。该协议通过使用串行采样而有所不同;通过执行连续采样,我们可以在内部验证线性阶段测量脂肪分解,并利用多次测量来计算脂肪分解速率,从而减少测量误差,以提高最终计算值的置信度。连续取样的缺点是检测需要更多的时间和试剂;然而,较长的时间范围减少了测量误差对速率估计标准误差的影响。此外,该协议测量FFA和甘油释放,并考虑FFA:甘油释放的比例,目标是实现3:1的比例,正如完全脂解和脂解产物释放到培养基中的预期一样72。
研究方案
所有动物的使用都得到了康奈尔大学威尔康奈尔医学院机构动物护理和使用委员会(IACUC)的批准。
1. 缓冲液和收集板的制备
- 通过将 5 g BSA 溶解在 100 mL 不含酚红的 Dulbecco 改性鹰培养基 (DMEM) 中,制成 5% 牛血清白蛋白 (BSA)。轻轻搅拌BSA溶解(摇晃适得其反)。BSA完全溶解后,用0.2μm过滤器对培养基进行过滤灭菌。将BSA培养基在4°C下储存长达1个月。
- 使控制和刺激介质的工作浓度。控制介质:5% BSA 介质,带车辆控制。刺激培养基:含有0.5μM CL-316,243的5%BSA培养基。为每个实验制作新鲜的刺激培养基。
- 将要使用的培养基加热至37°C。 标记用于培养基收集的 96 孔板。
2. 样品制备
- 如下所述进行细胞培养。在无菌通风橱中进行所有细胞工作,以尽量减少外部污染。
- 分离和分化原代前脂肪细胞,如73,74。
- 高密度接种原代前脂肪细胞,例如 1 x 105 个细胞 /孔在 1 mL/孔培养基中的 24 孔板中(DMEM/F12 中的 15% 胎牛血清 (FBS) 和 1x 青霉素-链霉素-谷氨酰胺)。
- 细胞达到 100% 汇合后,在培养基中用 5 μM 地塞米松、0.5 mM 3-异丁基-1-甲基黄嘌呤、1 μg/mL 胰岛素和 1 μM 噻唑烷二酮 (TZD) 分化 3 天。然后,换成含有 1 μg/mL 胰岛素的培养基至少 3 天以生长脂滴。在 24 孔板中使用 1 mL/孔培养基。
- 每2或3天用胰岛素更换培养基(1ml /孔)。细胞可以在含有胰岛素的培养基中维持长达2周。在此测定中,仅使用分化率超过90%且各组相似的培养物,因为分化的减少可能会被误解为脂解率的降低。
- 在测量脂解之前,将细胞在无胰岛素培养基中培养24小时。
注意:培养基中的胰岛素维持脂滴,但也抑制脂解。在没有胰岛素的情况下孵育24小时允许完全脂解活化而不会损失脂滴体积。在某些系统中,可能需要缩短或延长不使用胰岛素的培养时间。
- 用DPBS洗涤细胞一次,以去除培养基中的残留血清。
注意:此协议不包括血清饥饿,这可以激活脂肪分解。血清饥饿可由研究人员自行决定。
- 分离和分化原代前脂肪细胞,如73,74。
- 如下所述进行 离体 培养。
- 准备一个6孔板,每个组织都有一个孔,用于从每只小鼠收集。将 4 mL 室温 DMEM 放入要使用的每个孔中。
注:收集介质中的 BSA 不是必需的。 - 准备一个用于脂肪分解测定的48孔板,每个重复一个孔。将 400 μL 室温 DMEM 放入要使用的每个孔中。每只小鼠每个组织使用两到四个对照和两到四个刺激孔。
- 在麻醉下通过颈椎脱位对小鼠实施安乐死,使用双侧气胸等辅助方法。在这里,我们使用一只 32 克、7 个月大的雌性 C57BL/6J 小鼠,用 45% 的高脂肪饮食喂养 4 个月。
注意:此协议也可用于男性,以及其他菌株,饮食和年龄。 - 喷洒70%乙醇,用剪刀在腹部皮肤中心做一个小(~1厘米)侧切口,用拇指和食指捏住两侧将皮肤拉开,将下腹部皮肤折叠起来,露出后皮下库。定位并切除腹股沟淋巴结,并使用镊子钝化解剖紧邻腹股沟淋巴结的腹股沟脂肪组织。
- 为了收集性腺脂肪组织,在腹膜上做一个横向和垂直切口以进入腹膜腔。用镊子握住性腺脂肪垫,沿着子宫(或男性附睾)切开,以去除性腺脂肪组织。将收集的仓库放入6孔板中。
- 从孔中取出纸巾,放在硅胶垫上,用剪刀切成 5 至 7 毫克的大块。
- 为每个测定孔称出 25 至 30 mg(五块或六块),并放入 48 孔测定板中。称量前,用干净的毛巾吸干纸巾以除去任何介质。取出组织后称量重量舟,并记录留下的任何残留物的重量。在样品之间擦拭配重舟,必要时重新去皮。为每张纸巾使用新的配重船。
- 称量所有组织样品后,将48孔测定板置于37°C,10%CO2 培养箱中15分钟。
- 准备一个6孔板,每个组织都有一个孔,用于从每只小鼠收集。将 4 mL 室温 DMEM 放入要使用的每个孔中。
3. 脂肪分解试验
- 执行媒体收集。在无菌通风橱中进行培养基转移和随后的样品收集,以最大程度地减少来自外部来源的潜在污染。
- 在t = 0时,除去培养基,每孔加入400μL对照或刺激培养基,并将测定板放入37°C,10%CO2 培养箱中。对于 离体 组织培养,使用移液管小心地去除培养基;切勿使用抽吸。
注意:或者,用对照和刺激培养基准备第二块板,并转移组织。 - 在t = 1,2,3和4小时时,收集200μL培养基,用200μL适当的对照或刺激培养基替换,然后将测定板返回到培养箱中。将收集板储存在4°C。 要确定BSA培养基的FFA缓冲容量,请在24小时使用额外的集合。
注意:实验可以在这里停止,收集的培养基可以储存在-20°C。
- 在t = 0时,除去培养基,每孔加入400μL对照或刺激培养基,并将测定板放入37°C,10%CO2 培养箱中。对于 离体 组织培养,使用移液管小心地去除培养基;切勿使用抽吸。
4. FFA比色法
- 将试剂加热至室温,将一瓶颜色试剂A与一瓶溶剂A溶解,将一瓶颜色试剂B与一瓶溶剂B溶解。从复溶之日起,这些试剂最好在1周内使用。复溶后1个月丢弃。
- 解冻并混合样品。
- 创建 FFA 标准曲线。标准溶液为1 mM。将以下体积与标准曲线试剂一起使用:25 μL、20 μL、15 μL、10 μL、10 μL(1:2 稀释)、10 μL(1:4 稀释)、10 μL(1:8 稀释)和 10 μL 水以获得最大范围。对于低FFA水平,10 μL 的 1 mM、0.8 mM、0.6 mM、0.4 mM、0.2 mM、0.1 mM 和 0.05 mM 标准品可能更适用。
- 将标准品和样品移液到96孔测定板中。推荐的样品体积为 10 μL,包括三个具有与样品相同体积的 BSA 培养基的孔进行背景校正。
注意:如果样品浓度超出标准曲线的范围,请重复测定,将样品体积调整为 2-25 μL。 - 向每个孔中加入 150 μL 试剂 A 并混合。避免产生气泡。用细针戳破任何气泡。将测定板在37°C孵育5分钟。
- 读取板在550nm和660nm参考处的吸光度(读数A)。
- 向每个孔中加入 75 μL 试剂 B 并混合。避免产生气泡。用细针戳破任何气泡。将测定板在37°C孵育5分钟。
- 在550nm和660nm参考处再次读取板的吸光度(读数B)。
5. 甘油比色法
- 用 36 mL 超纯水复溶游离甘油试剂并适应室温。这些试剂最好在几周内使用。复溶后2个月丢弃。
- 解冻并混合样品。
- 通过对甘油标准溶液和水空白进行7点,2倍连续稀释来创建甘油标准曲线。
注意:标准曲线在高达 25 μL 的 2.8 mM 甘油时相对线性,但在较高浓度下不线性。 - 将标准品和样品各 25 μL 移液到 96 孔测定板中。包括三个带有BSA培养基的孔,用于背景校正。
- 向每个孔中加入 175 μL 游离甘油试剂并混合。避免产生气泡。用细针戳破任何气泡。将测定板在37°C孵育5分钟。
- 读取板在540nm处的吸光度。
6. 脂解率的计算
- 从光密度 (OD) 值开始。对于甘油,直接使用A 540 OD 值。根据以下公式计算FFA测定的OD:
OD = (读数 B: A 550 - A 660) - (读数 A: A550 - A660) - 使用标准曲线计算收集样品中的FFA和甘油水平。在y轴上绘制标准OD值,在x轴上,使用相对于样品体积的标准浓度(即,在含有10 μL样品的平板上含有20 μL1 mM FFA标准的孔的浓度等于2 mM)。拟合线性趋势线:
y = mx + b - 目视检查标准曲线并去除测定线性范围之外的任何点。使用以下公式计算样品浓度:
样品浓度:x = (OD - b) ÷ m - 调整并重新测定超出线性测定范围的样品。要获得最终样品浓度,请从样品浓度中减去仅含有BSA培养基的背景孔浓度。
- 根据公式计算每个样品在每个时间点产生的FFA和甘油的摩尔数:

其中 C n = 时间 t = n 时的浓度;Vt = 井中的总体积;Vs = 样品收集体积;M n = 在时间 t = n 时产生的摩尔数(当浓度以 mM 为单位且体积以 mL 为单位时,输出为 μMol)。
例如,在不同的时间点:
M 1 = C1 × Vt
M 4 = C4 × Vt + (C1 + C 2+ C3)Vs
或
M 4 = C4 × Vt + C3 × V s + C2 × V s + C1 ×V s - 通过将每个样品的组织重量除以克为单位,获得μmol/g单位,归一化为组织重量。对于培养的细胞,值以μmol/孔表示。确保细胞数量和分化效率在孔与孔之间具有可比性。
注意:增殖或分化效率的差异将使结果的解释复杂化,需要另一种标准化方法(例如,标准化为蛋白质;参见讨论)。 - 单独计算每个样品产生的μmol/g(y轴)与时间(x轴)的斜率。
- 在电子表格中,这可以使用 =SLOPE(known_ys,known_xs) 函数来完成。在新单元格中,键入"=SLOPE"(然后使用光标突出显示样品甘油或 FFA 值(以μmol/g为单位),然后突出显示相应的时间值)。
- 验证数据的线性度。R2 值是确定样品线性的快速方法。在电子表格中,可以使用 =RSQ(known_ys,known_xs) 函数完成此操作,方法与步骤 6.7.1 中所述相同,但初始输入为 =RSQ。确保R 2 值> 0.98;较低的值表示与线性度的偏差。这可能是由测量/采样误差或线性度损失引起的。
- 测试线性的另一种方法是对每个样本执行线性回归并绘制残差图。在统计分析软件中,生成一个XY表,每个时间点都有一个Y值。选择分析>简单线性回归,然后在点击确定之前选中残差图框。残差图将显示为新图形。
- 使用每个样品的FFA和甘油产生速率(即斜率[(μmol/g/h])作为单独的数据点进行统计分析,并在比较不同的脂解条件时绘制值。如果要比较不同基因型的脂解率,则每只动物使用两个或三个样本作为技术重复,并使用每只动物一个数据点的平均值,以便样本量等于动物的数量。
结果
我们测量了体外分化脂肪细胞的基础和刺激脂解率。通过用5μM地塞米松、0.5mM IBMX、1μg/mL胰岛素和1μM曲格列酮处理融合细胞4天,将来自腹股沟白色脂肪组织的原代前脂肪细胞分化为脂肪细胞,然后用1μg/mL胰岛素再处理3天。在脂肪分解测定之前,将细胞在没有胰岛素的培养基中孵育24小时。在时间= 0h时,用PBS洗涤细胞一次,然后将含有10μM CL-316,243或载体对照的2%BSA的无酚红DMEM加入到12孔?...
讨论
在这里,我们提供了一种用于测量脂肪细胞和离体脂肪组织中脂肪分解速率的基本方案。为了量化脂解,重要的是测量线性相中的脂解速率。我们使用连续采样技术,其中收集大部分培养基并定期更换为新鲜培养基。这种半保守的方法允许添加具有FFA缓冲能力的新鲜BSA,并延迟反馈抑制,延长线性脂解的持续时间。该实验设计试图概括体内脂肪组织的血管化,其提供新鲜白蛋白以结?...
披露声明
作者没有什么可透露的。
致谢
这项工作得到了美国国立卫生研究院向S.M.R.拨款R01DK126944的支持。
材料
| Name | Company | Catalog Number | Comments |
| 24-Well tissue culture treated plate | Corning Inc | 3527 | Must be tissue culture treated for adipocyte differntiation |
| 48-Well flat bottom plate with lid | Corning Inc | 353078 | Can be tissue culture treated |
| 6-Well flat bottom plate with lid | Corning Inc | 353046 | Can be tissue culture treated |
| 96-Well PCR Plate | USA sceintific | 1402-9100 | Any conical 0.2 mL PCR plate will be convenient |
| Bovine Serum Albumin | Sigma Aldrich | A9418 | FFA free BSA such as A8806, is also commonly used. The BSA should not have detectable FFA, also lot to lot variations in BSA can impact the observed rate of lipolysis |
| CL-316,243 | Sigma Aldrich | C5976 | CAS #: 138908-40-4 availaible from other suppliers |
| CO2 incubator | PHCBI | MCO-170AICUVH | CO2 should ideally be set to 10% for adipose tissue, however 5% CO2 will also work |
| DMEM, low glucose, no phenol red | Thermofischer | 11054020 | Any phenol red free media should work, DMEM/F12, RPMI, but should contain volatile buffering capacity, i.e. biocarbonate |
| FFA-free Bovine serum albumin | Equitech-Bio, Inc, | BAH66 | |
| Free Glycerol Reagent | Sigma Aldrich | F6428 | |
| Glycerol Standard Solution | Sigma Aldrich | G7793 | This can also be made by diluting glycerol to the desired concentration |
| HR Series NEFA Standard Solution | Fujifilm | 276-76491 | |
| HR Series NEFA-HR (2) Color Reagent A | Fujifilm | 999-34691 | |
| HR Series NEFA-HR (2) Color Reagent B | Fujifilm | 991-34891 | |
| HR Series NEFA-HR (2) Solvent A | Fujifilm | 995-34791 | |
| HR Series NEFA-HR (2) Solvent B | Fujifilm | 993-35191 | |
| Microbiological Incubator | Fischer Scientific | S28668 | Any incubator at 37C can be used |
| Nunc MicroWell 96-Well Plates | Thermo Scientific | 269620 | Any optically clear, flat bottom 96-well plate works |
| Silicone Laboratory Benchtop Mat | VWR | 76045-300 | Glass plate can also be used. Absorbant surfaces are not recommended |
| Spectrophotometer/Microplate Reader | Molecular devices | SpectraMax i3x | Any plate reader that can read at 540, 550 and 660 mm will work |
| V Bovine serum albumin | Sigma-Aldrich | 810531 | |
| WypAll X70 Wipers | Kimberly-Clark | 41200 | Any high quality paper towel will work |
参考文献
- Vaughan, M., Berger, J. E., Steinberg, D. Hormone-sensitive lipase and monoglyceride lipase activities in adipose tissue. The Journal of Biological Chemistry. 239, 401-409 (1964).
- Zimmermann, R., et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science. 306 (5700), 1383-1386 (2004).
- Lass, A., et al. Adipose triglyceride lipase-mediated lipolysis of cellular fat stores is activated by CGI-58 and defective in Chanarin-Dorfman syndrome. Cell Metabolism. 3 (5), 309-319 (2006).
- Stralfors, P., Bjorgell, P., Belfrage, P. Hormonal regulation of hormone-sensitive lipase in intact adipocytes: identification of phosphorylated sites and effects on the phosphorylation by lipolytic hormones and insulin. Proceedings of the National Academy of Sciences. 81 (11), 3317-3321 (1984).
- Miyoshi, H., et al. Perilipin promotes hormone-sensitive lipase-mediated adipocyte lipolysis via phosphorylation-dependent and -independent mechanisms. The Journal of Biological Chemistry. 281 (23), 15837-15844 (2006).
- Sztalryd, C., et al. Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. The Journal of Cell Biology. 161 (6), 1093-1103 (2003).
- Lafontan, M., Langin, D. Lipolysis and lipid mobilization in human adipose tissue. Progress in Lipid Research. 48 (5), 275-297 (2009).
- Grabner, G. F., Xie, H., Schweiger, M., Zechner, R. Lipolysis: cellular mechanisms for lipid mobilization from fat stores. Nature Metabolism. 3 (11), 1445-1465 (2021).
- Weiss, S. B., Kennedy, E. P., Kiyasu, J. Y. The enzymatic synthesis of triglycerides. The Journal of Biological Chemistry. 235, 40-44 (1960).
- Kennedy, E. P. Biosynthesis of complex lipids. Federation Proceedings. 20, 934-940 (1961).
- Wendel, A. A., Lewin, T. M., Coleman, R. A. Glycerol-3-phosphate acyltransferases: rate limiting enzymes of triacylglycerol biosynthesis. Biochimica et Biophysica Acta. 1791 (6), 501-506 (2009).
- Johansson, S. M., Lindgren, E., Yang, J. N., Herling, A. W., Fredholm, B. B. Adenosine A1 receptors regulate lipolysis and lipogenesis in mouse adipose tissue-interactions with insulin. European Journal of Pharmacology. 597 (1-3), 92-101 (2008).
- Gnad, T., et al. Adenosine activates brown adipose tissue and recruits beige adipocytes via A2A receptors. Nature. 516 (7531), 395-399 (2014).
- Fried, S. K., et al. Resistance to the antilipolytic effect of insulin in adipocytes of African-American compared to Caucasian postmenopausal women. Journal of Lipid Research. 51 (5), 1193-1200 (2010).
- Lee, M. J., Fried, S. K. Optimal protocol for the differentiation and metabolic analysis of human adipose stromal cells. Methods in Enzymology. 538, 49-65 (2014).
- Fricke, K., Heitland, A., Maronde, E. Cooperative activation of lipolysis by protein kinase A and protein kinase C pathways in 3T3-L1 adipocytes. Endocrinology. 145 (11), 4940-4947 (2004).
- Bergan, H. E., Kittilson, J. D., Sheridan, M. A. PKC and ERK mediate GH-stimulated lipolysis. Journal of Molecular Endocrinology. 51 (2), 213-224 (2013).
- Schmitz-Peiffer, C. The tail wagging the dog--regulation of lipid metabolism by protein kinase C. The FEBS Journal. 280 (21), 5371-5383 (2013).
- Carmen, G. Y., Victor, S. M. Signalling mechanisms regulating lipolysis. Cellular Signalling. 18 (4), 401-408 (2006).
- Zu, L., et al. Bacterial endotoxin stimulates adipose lipolysis via toll-like receptor 4 and extracellular signal-regulated kinase pathway. The Journal of Biological Chemistry. 284 (9), 5915-5926 (2009).
- Zhang, H. H., Halbleib, M., Ahmad, F., Manganiello, V. C., Greenberg, A. S. Tumor necrosis factor-alpha stimulates lipolysis in differentiated human adipocytes through activation of extracellular signal-related kinase and elevation of intracellular cAMP. Diabetes. 51 (10), 2929-2935 (2002).
- Tan, X., et al. TNF-α downregulates CIDEC via MEK/ERK pathway in human adipocytes. Obesity. 24 (5), 1070-1080 (2016).
- Laurencikiene, J., et al. NF-kappaB is important for TNF-alpha-induced lipolysis in human adipocytes. Journal of Lipid Research. 48 (5), 1069-1077 (2007).
- van Hall, G., et al. Interleukin-6 stimulates lipolysis and fat oxidation in humans. The Journal of Clinical Endocrinology and Metabolism. 88 (7), 3005-3010 (2003).
- Wueest, S., et al. Mesenteric fat lipolysis mediates obesity-associated hepatic steatosis and insulin resistance. Diabetes. 65 (1), 140-148 (2016).
- Trujillo, M. E., et al. Interleukin-6 regulates human adipose tissue lipid metabolism and leptin production in vitro. The Journal of Clinical Endocrinology and Metabolism. 89 (11), 5577-5582 (2004).
- Kitamura, T., et al. Insulin-induced phosphorylation and activation of cyclic nucleotide phosphodiesterase 3B by the serine-threonine kinase Akt. Molecular and Cellular Biology. 19 (9), 6286-6296 (1999).
- Chakrabarti, P., et al. Insulin inhibits lipolysis in adipocytes via the evolutionarily conserved mTORC1-Egr1-ATGL-mediated pathway. Molecular and Cellular Biology. 33 (18), 3659-3666 (2013).
- Collins, S., Daniel, K. W., Petro, A. E., Surwit, R. S. Strain-specific response to beta 3-adrenergic receptor agonist treatment of diet-induced obesity in mice. Endocrinology. 138 (1), 405-413 (1997).
- Surwit, R. S., Dixon, T. M., Petro, A. E., Daniel, K. W., Collins, S. Diazoxide restores beta3-adrenergic receptor function in diet-induced obesity and diabetes. Endocrinology. 141 (10), 3630-3637 (2000).
- Gettys, T. W., et al. Age-dependent changes in beta-adrenergic receptor subtypes and adenylyl cyclase activation in adipocytes from Fischer 344 rats. Endocrinology. 136 (5), 2022-2032 (1995).
- Mowers, J., et al. Inflammation produces catecholamine resistance in obesity via activation of PDE3B by the protein kinases IKKε and TBK1. eLife. 2, e01119 (2013).
- Valentine, J. M., et al. β3-Adrenergic receptor downregulation leads to adipocyte catecholamine resistance in obesity. The Journal of Clinical Investigation. 132 (2), e153357 (2022).
- Collins, S., et al. Impaired expression and functional activity of the beta 3- and beta 1-adrenergic receptors in adipose tissue of congenitally obese (C57BL/6J ob/ob) mice. Molecular Endocrinology. 8 (4), 518-527 (1994).
- Collins, S., Surwit, R. S. The beta-adrenergic receptors and the control of adipose tissue metabolism and thermogenesis. Recent Progress in Hormone Research. 56, 309-328 (2001).
- Dixon, T. M., Daniel, K. W., Farmer, S. R., Collins, S. CCAAT/enhancer-binding protein alpha is required for transcription of the beta 3-adrenergic receptor gene during adipogenesis. The Journal of Biological Chemistry. 276 (1), 722-728 (2001).
- Lohse, M. J., Benovic, J. L., Codina, J., Caron, M. G., Lefkowitz, R. J. beta-Arrestin: a protein that regulates beta-adrenergic receptor function. Science. 248 (4962), 1547-1550 (1990).
- Nantel, F., et al. The human beta 3-adrenergic receptor is resistant to short term agonist-promoted desensitization. Molecular Pharmacology. 43 (4), 548-555 (1993).
- Liggett, S. B., Freedman, N. J., Schwinn, D. A., Lefkowitz, R. J. Structural basis for receptor subtype-specific regulation revealed by a chimeric beta 3/beta 2-adrenergic receptor. Proceedings of the National Academy of Sciences. 90 (8), 3665-3669 (1993).
- Baker, J. G. The selectivity of beta-adrenoceptor agonists at human beta1-, beta2- and beta3-adrenoceptors. British Journal of Pharmacology. 160 (5), 1048-1061 (2010).
- Lafontan, M. Inhibition of epinephrine-induced lipolysis in isolated white adipocytes of aging rabbits by increased alpha-adrenergic responsiveness. Journal of Lipid Research. 20 (2), 208-216 (1979).
- Baker, J. G. The selectivity of beta-adrenoceptor antagonists at the human beta1, beta2 and beta3 adrenoceptors. British Journal of Pharmacology. 144 (3), 317-322 (2005).
- Jensen, M. D., Nielsen, S. Insulin dose response analysis of free fatty acid kinetics. Metabolism. 56 (1), 68-76 (2007).
- Jensen, M. D., Haymond, M. W., Gerich, J. E., Cryer, P. E., Miles, J. M. Lipolysis during fasting. Decreased suppression by insulin and increased stimulation by epinephrine. The Journal of Clinical Investigation. 79 (1), 207-213 (1987).
- Heckmann, B. L., et al. Defective adipose lipolysis and altered global energy metabolism in mice with adipose overexpression of the lipolytic inhibitor G0/G1 switch gene 2 (G0S2). The Journal of Biological Chemistry. 289 (4), 1905-1916 (2014).
- Shin, H., et al. Lipolysis in brown adipocytes is not essential for cold-induced thermogenesis in mice. Cell Metabolism. 26 (5), 764.e5-777.e5 (2017).
- Treble, D. H., Mayer, J. Glycerolkinase activity in white adipose tissue of obese-hyperglycaemic mice. Nature. 200, 363-364 (1963).
- Possik, E., et al. New mammalian glycerol-3-phosphate phosphatase: role in beta-cell, liver and adipocyte metabolism. Frontiers in Endocrinology. 12, 706607 (2021).
- Romero Mdel, M., Sabater, D., Fernandez-Lopez, J. A., Remesar, X., Alemany, M. Glycerol production from glucose and fructose by 3T3-L1 cells: a mechanism of adipocyte defense from excess substrate. PLoS One. 10 (10), e0139502 (2015).
- Mugabo, Y., et al. Identification of a mammalian glycerol-3-phosphate phosphatase: Role in metabolism and signaling in pancreatic beta-cells and hepatocytes. Proceedings of the National Academy of Sciences. 113 (4), E430-E439 (2016).
- Hanson, R. W., Reshef, L. Glyceroneogenesis revisited. Biochimie. 85 (12), 1199-1205 (2003).
- Vaughan, M. The production and release of glycerol by adipose tissue incubated in vitro. The Journal of Biological Chemistry. 237, 3354-3358 (1962).
- Jensen, M. D., Ekberg, K., Landau, B. R. Lipid metabolism during fasting. American Journal of Physiology-Endocrinology and Metabolism. 281 (4), E789-E793 (2001).
- Ballard, F. J., Hanson, R. W., Leveille, G. A. Phosphoenolpyruvate carboxykinase and the synthesis of glyceride-glycerol from pyruvate in adipose tissue. The Journal of Biological Chemistry. 242 (11), 2746-2750 (1967).
- Reshef, L., Hanson, R. W., Ballard, F. J. A possible physiological role for glyceroneogenesis in rat adipose tissue. The Journal of Biological Chemistry. 245 (22), 5979-5984 (1970).
- Gorin, E., Tal-Or, Z., Shafrir, E. Glyceroneogenesis in adipose tissue of fasted, diabetic and triamcinolone treated rats. European Journal of Biochemistry. 8 (3), 370-375 (1969).
- Elia, M., Zed, C., Neale, G., Livesey, G. The energy cost of triglyceride-fatty acid recycling in nonobese subjects after an overnight fast and four days of starvation. Metabolism. 36 (3), 251-255 (1987).
- Reshef, L., et al. Glyceroneogenesis and the triglyceride/fatty acid cycle. Journal of Biological Chemistry. 278 (33), 30413-30416 (2003).
- Edens, N. K., Leibel, R. L., Hirsch, J. Mechanism of free fatty acid re-esterification in human adipocytes in vitro. Journal of Lipid Research. 31 (8), 1423-1431 (1990).
- Vaughan, M., Steinberg, D. Effect of hormones on lipolysis and esterification of free fatty acids during incubation of adipose tissue in vitro. Journal of Lipid Research. 4, 193-199 (1963).
- Brooks, B., Arch, J. R., Newsholme, E. A. Effects of hormones on the rate of the triacylglycerol/fatty acid substrate cycle in adipocytes and epididymal fat pads. Federation of European Biochemical Societies Letters. 146 (2), 327-330 (1982).
- Bjorntorp, P., Karlsson, M., Hovden, A. Quantitative aspects of lipolysis and reesterification in human adipose tissue in vitro. Acta Medica Scandinavica. 185 (1-2), 89-97 (1969).
- Angel, A., Desai, K., Halperin, M. L. Free fatty acid and ATP levels in adipocytes during lipolysis. Metabolism. 20 (1), 87-99 (1971).
- Husted, A. S., et al. Autocrine negative feedback regulation of lipolysis through sensing of NEFAs by FFAR4/GPR120 in WAT. Molecular Metabolism. 42, 101103 (2020).
- Fain, J. N., Shepherd, R. E. Free fatty acids as feedback regulators of adenylate cyclase and cyclic 3':5'-AMP accumulation in rat fat cells. The Journal of Biological Chemistry. 250 (16), 6586-6592 (1975).
- Burns, T. W., Langley, P. E., Terry, B. E., Robinson, G. A. The role of free fatty acids in the regulation of lipolysis by human adipose tissue cells. Metabolism. 27 (12), 1755-1762 (1978).
- Kalderon, B., et al. Suppression of adipose lipolysis by long-chain fatty acid analogs. Journal of Lipid Research. 53 (5), 868-878 (2012).
- Schweiger, M., et al. Measurement of lipolysis. Methods in Enzymology. 538, 171-193 (2014).
- Decaunes, P., Bouloumie, A., Ryden, M., Galitzky, J. Ex vivo analysis of lipolysis in human subcutaneous adipose tissue explants. Bio-Protocol. 8 (3), e2711 (2018).
- Roy, D., Myers, J. M., Tedeschi, A. Protocol for assessing ex vivo lipolysis of murine adipose tissue. STAR Protocols. 3 (3), 101518 (2022).
- Baskaran, P., Thyagarajan, B. Measurement of basal and forskolin-stimulated lipolysis in inguinal adipose fat pads. Journal of Visualized Experiments. 125 (125), 55625 (2017).
- Reilly, S. M., et al. Catecholamines suppress fatty acid re-esterification and increase oxidation in white adipocytes via STAT3. Nature Metabolism. 2 (7), 620-634 (2020).
- Liu, L., et al. Isolation of mouse stromal vascular cells for monolayer culture. Methods in Molecular Biology. 1566, 9-16 (2017).
- DeLuca, J. H., Reilly, S. M. . Methods in Molecular Biology. , (2023).
- Richard, G., Vernon, R. A. C. New Perspectives in Adipose Tissue. Butterworth-Heinemann. , (1985).
- Brito, M. N., Botion, L. M., Brito, N. A., Kettelhut, I. C., Migliorini, R. H. Lipolysis and glycerokinase activity in brown adipose tissue of rat fed a high protein, carbohydrate-free diet. Hormone and Metabolic Research. 26 (1), 51-52 (1994).
- Bertin, R. Glycerokinase activity and lipolysis regulation in brown adipose tissue of cold acclimated rats. Biochimie. 58 (4), 431-434 (1976).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。