
Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Lipoliz Oranının Ex Vivo Murin Yağ Dokusu ve İn Vitro Diferansiye Primer Preadipositlerin Ölçülmesi
Bu Makalede
Özet
Adipositlerde trigliserit lipoliz, serbest yağ asitlerinin ve gliserolün serbest bırakılmasıyla sonuçlanan önemli bir metabolik süreçtir. Burada, adipositlerde bazal ve uyarılmış lipolizi ve farelerden ex vivo yağ dokusunu ölçmek için ayrıntılı bir protokol sunuyoruz.
Özet
Adipositler, lipit damlacıklarında trigliseritler şeklinde enerji depolar. Bu enerji, yağ asidi yan zincirlerinin gliserol omurgasından sırayla ayrıldığı ve serbest yağ asitlerinin ve gliserolün salınmasına neden olan lipoliz yoluyla harekete geçirilebilir. Beyaz adipositlerde gliserol kinazın düşük ekspresyonu nedeniyle, gliserol geri alım oranları ihmal edilebilirken, yağ asidi geri alımı, albümin gibi medya bileşenlerinin yağ asidi bağlama kapasitesi tarafından belirlenir. Hem gliserol hem de yağ asidinin ortama salınımı, lipolitik oranı belirlemek için kolorimetrik testlerle ölçülebilir. Bu faktörleri birden fazla zaman noktasında ölçerek, lipolizin doğrusal oranını yüksek güvenle belirleyebilirsiniz. Burada, farelerden in vitro diferansiye adipositlerin ve ex vivo yağ dokusunun lipoliz ölçümü için ayrıntılı bir protokol sunuyoruz. Bu protokol ayrıca diğer preadiposit hücre hatları veya diğer organizmalardan yağ dokusu için optimize edilebilir; Dikkat edilmesi gereken noktalar ve optimizasyon parametreleri ele alınmıştır. Bu protokol, fare modelleri ve tedavileri arasındaki adiposit lipoliz oranını belirlemede ve karşılaştırmada yararlı olacak şekilde tasarlanmıştır.
Giriş
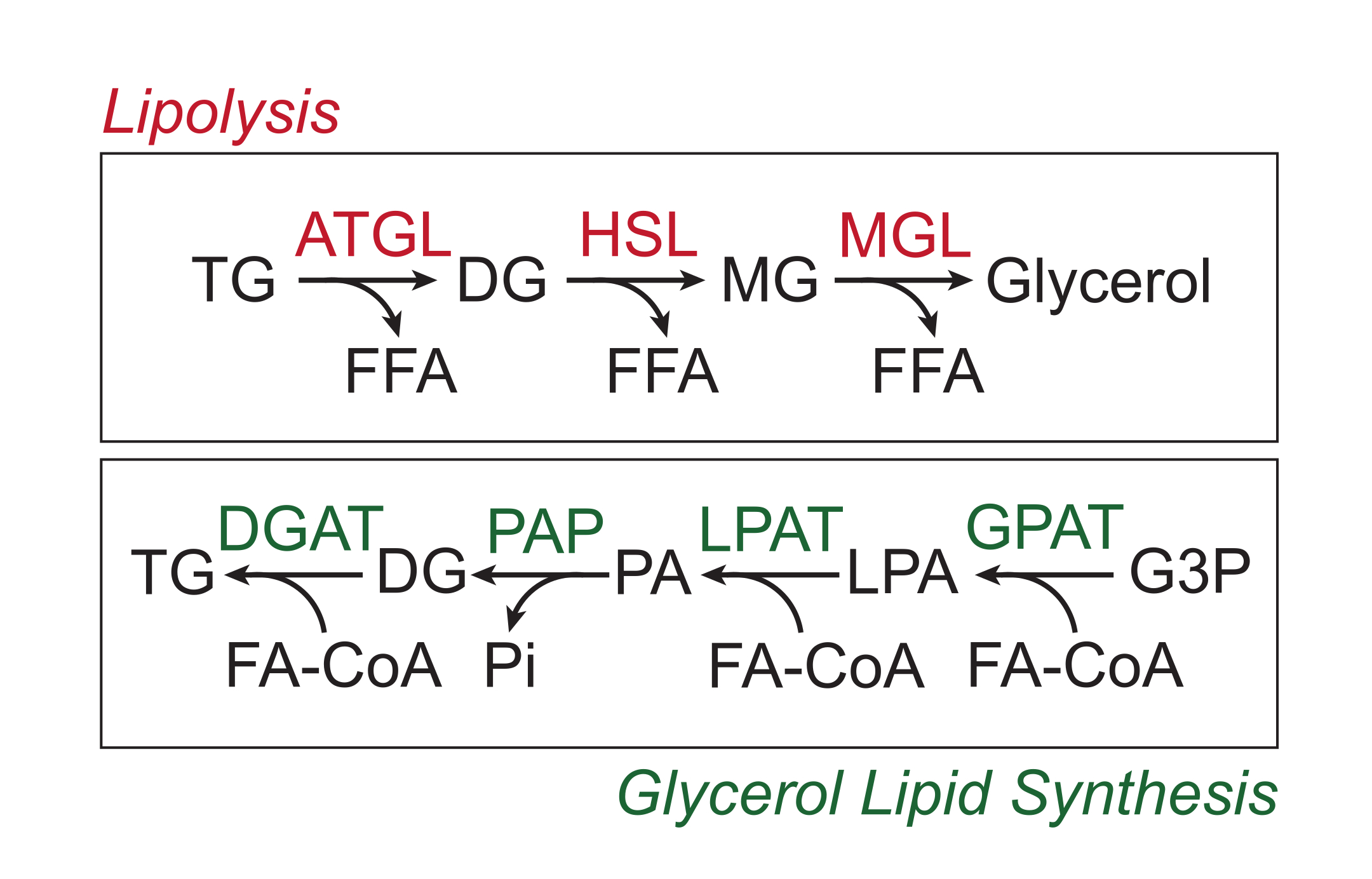
Fazla besinler, lipit damlacıklarının nötr lipit çekirdeğinde trigliseritler şeklinde beyaz yağ dokusunda depolanır. Trigliserit depoları, yağ asidi yan zincirlerinin yağ dokusu trigliserit lipaz (ATGL), hormona duyarlı lipaz (HSL) ve monogliserit lipaz (MGL) tarafından sırayla bölündüğü ve serbest yağ asitlerinin (FFA'lar) ve gliserol omurgasının 1,2 salınımına neden olan bir süreç olan lipoliz yoluyla mobilize edilir. Lipoliz, yağ dokusunda katekolamin sinyalizasyonu ile aktive edilir. Sempatik sinir terminalleri, adiposit plazma zarındaki β-adrenerjik reseptörlere bağlanan katekolaminleri lokal olarak serbest bırakır. Ligand bağlanması üzerine, bu G-protein kuplajlı reseptörler (GPCR'ler), Gαs yoluyla adenil siklazı aktive eder. Daha sonra protein kinaz A'nın (PKA) cAMP ile aktivasyonu, hem ATGL hem de HSL'nin yukarı regülasyonuna neden olur. Perilipin-1'in PKA tarafından fosforilasyonu, ATGL3'ü bağlayan ve birlikte aktive eden ABHD5'in (CGI-58 olarak da bilinir) ayrışmasına neden olur. PKA, HSL'yi doğrudan fosforile eder, sitosolden lipit damlacığına translokasyonunu teşvik eder, burada fosforile perilipin-1 ile etkileşim, lipaz aktivitesini daha da arttırır 4,5,6,7. Lipolizde yer alan üçüncü lipaz, MGL, katekolamin sinyalizasyonu8 ile düzenlenmiyor gibi görünmektedir. Önemli olarak, adipositlerde trigliserit sentezine, bir ara madde olarak monogliserit oluşumunu içermeyen gliserol lipid sentez yolu aracılık eder; Bunun yerine, gliserol-3-fosfat açil transferazlar, fosfatidik asit oluşturmak için başka bir yağ açil-CoA ile birleştirilen ve daha sonra trigliseritlerin son sentezinden önce digliseritlere izomerize edilen lizofosfatidik asit oluşumunu katalize eder (Şekil 1) 9,10,11.

Resim 1: Lipoliz ve gliserol lipid sentez yolakları. Üst: Lipolitik yolak; kırmızı renkle gösterilen enzimler: yağ dokusu trigliserit lipaz (ATGL), hormona duyarlı lipaz (HSL) ve monogliserit lipaz (MGL). Alt: gliserol lipid sentez yolu; yeşil renkte gösterilen enzimler: digliserit asiltransferaz (DGAT), fosfatidik asit fosfataz (PAP), lizofosfatidik asit asiltransferaz (LPAT, LPAAT olarak da bilinir) ve gliserol-3-fosfat asiltransferaz (GPAT). Lipitler: trigliserit (TG), digliserit (DG), monogliserit (MG), serbest yağ asidi (FFA), yağ açil-CoA (FA-CoA), lizofosfatidik asit (LPA) ve fosfatidik asit (PA). Diğer metabolitler: inorganik fosfat (Pi) ve gliserol 3-fosfat (G3P). Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}
Hücre dışı adenozin, adenil siklaz aktivitesini etkilemek için Gs ve G i-kuplajlı GPCR'ler aracılığıyla çalışan lipolizin bir başka önemli düzenleyicisidir. Adipositlerdeki baskın adenozin reseptörü ADORA1, adenilil siklazı ve böylece Gi12'nin aktivasyonu yoluyla lipolizi inhibe eder. Daha düşük seviyelerde ve öncelikle kahverengi adipositlerde eksprese edilen ADORA2A,13 sinyalleme yoluyla lipolizi aktive eder. ADORA1 hem bazal lipolizi hem de adrenerjik agonistlere yanıtı etkiler. Adenozinin lipoliz üzerindeki etkisi, adenozini nötralize etmek için adenozin deaminaz ve ADORA1'e özgü agonist fenilizopropiladenozin14,15 eklenerek kontrol edilebilir. G q-kuplajlı GPCR'lerin hormonal aktivasyonu, fosfolipaz C ve protein kinaz C 16,17,18,19'un aktivasyonu yoluyla lipolizi de etkileyebilir. Enflamatuar sinyaller ayrıca lipolitik oranları da etkiler. LPS (ve diğer endotoksinler) ile TLR4 aktivasyonu, perilipin-1 ve HSL20'yi fosforile eden ERK'yi aktive ederek lipolitik hızı arttırır. TNF-α ayrıca ERK ve NF-κB aktivasyonu yoluyla lipolizi aktive eder, ayrıca fosfodiesteraz PDE-3B ve CIDEC21,22,23'ün transkripsiyonel downregülasyonu. IL-6 ayrıca, özellikle FFA salınımı hepatik steatoz ve glukoneogenezi etkileyen mezenterik yağ dokusunda artmış adiposit lipoliz ile ilişkilendirilmiştir24,25,26.
Lipoliz, beslenen durum sırasında insülin tarafından bastırılır. AKT, cAMP sinyalini bastırmak ve PKA aktivasyonunu önlemek için PDE-3B'yi fosforile eder ve aktive eder27. İnsülin ayrıca ATGL28'i transkripsiyonel olarak aşağı regüle eder. Obezite, adipositlerde β-adrenerjik reseptörlerin aşağı regülasyonu da dahil olmak üzere çeşitli mekanizmalarla katekolamin direncini arttırır 29,30,31,32,33. Adipositler üç β-adrenerjik reseptörün tümünü eksprese eder (β-1, β-2 ve β-3). β-1 ve β-2 adrenerjik reseptörleri her yerde eksprese edilirken, β-3 adrenerjik reseptör ağırlıklı olarak farelerde adipositlerde eksprese edilir34,35. Adrb3 ekspresyonu, adipogenez36 sırasında C / EBPα tarafından indüklenir. β-3 adrenerjik reseptör olgun adipositlerde yüksek oranda eksprese edilir. β-1 ve β-2 adrenerjik reseptörlerin aktivasyonu, β-arrestin37 ile geri besleme inhibisyonu nedeniyle kendi kendini sınırlar. β-3 adrenerjik reseptörün geri besleme inhibisyonuna, Adrb3 ekspresyonunu 33,38,39 azaltan diğer sinyal yolları aracılık eder.
Adiposit lipolizi aktive etmek için çok sayıda bileşik kullanılabilir. Katekolaminler lipolizin başlıca fizyolojik aktivatörleridir. Norepinefrin (veya noradrenalin) ve epinefrin (veya adrenalin) üç β-adrenerjik reseptörün hepsini aktive eder40. Norepinefrin ve epinefrin ayrıca α-adrenerjik reseptör sinyalinin aktivasyonu yoluyla lipolizi etkiler41. Yaygın olarak kullanılan β-adrenerjik reseptör agonistleri, seçici olmayan bir β-adrenerjik reseptör agonisti olan izoproterenol ve β-3 adrenerjik reseptör agonistleri CL-316,243 ve mirabegron42'yi içerir. Adipositlerin ağırlıklı olarak β-3 adrenerjik reseptörü ifade ettiği göz önüne alındığında, burada örnek olarak CL-316,243 kullanıyoruz. β-3 adrenerjik reseptör için özgüllüğü, onu in vivo olarak güvenle kullanılabilen adiposit katekolamin sinyallemesinin nispeten spesifik bir aktivatörü yapar. Hücre kültüründe yaygın olarak kullanılan 10 μM CL-316,243 konsantrasyonunun, maksimum yanıt33'e ulaşmak için gereken ~ 0.1 μM dozundan daha yüksek büyüklük sıraları olduğunu unutmayın. Forskolin, adrenerjik reseptörü atlayarak adenil siklaz ve aşağı akış lipolitik sinyallemesini doğrudan aktive eder. Lipolizin baskılayıcılarının yanı sıra daha birçok aktivatör vardır. Lipolizi uyarmak için bir bileşik seçerken, reseptör özgüllüğü ve aşağı akış sinyal yolları deneysel tasarım içinde dikkatlice düşünülmelidir.
Beyaz yağ dokusunda lipoliz oranı, açlık veya egzersiz sırasında soğuk toleransını ve besin mevcudiyetini etkileyen önemli bir metabolik faktördür43,44,45,46. Bu protokolün amacı, adiposit metabolizmasının anlaşılmasını ve çeşitli murin modellerinin metabolik fenotipini nasıl etkileyebileceğini kolaylaştıracak adiposit ve yağ dokusundaki lipoliz oranını ölçmektir. Lipolitik oranı ölçmek için, lipolitik ürünlerin medyadaki görünümünü (yani, FFA'lar ve gliserol) ölçüyoruz. Yöntem, lipolitik ürünlerin adipositten medyaya salınmasına dayanır. Beyaz adipositler düşük gliserol kinaz seviyelerini eksprese ettiğinden, gliserol geri alım oranları düşüktür47. Tersine, FFA'ların ve gliserolün lipoliz dışındaki metabolik yollarla üretimi de göz önünde bulundurulmalıdır. Adipositler, gliserol-3 fosfata karşı aktiviteye sahip bir fosfatazı eksprese ediyor gibi görünmektedir ve glikoz 48,49,50'den türetilen gliserol-3-fosfattan gliserol üretimini sağlar. Glikoliz, beyaz adipositlerde FFA reesterifikasyonu için kullanılan bir gliserol-3-fosfat kaynağıdır. Glikoz seviyeleri sınırlı olduğunda, gliseroneogenez, laktat ve piruvat51 gibi diğer 3 karbonlu kaynakları gerektirir. Lipoliz ile salınan FFA'ların hücre içinde kanalize edilmesi ve metabolik kaderleri tam olarak anlaşılamamıştır; Lipoliz tarafından salınan FFA'lar, yeniden esterleştirilmeden veya β-oksidasyona uğramadan önce yağlı açil-CoA'ya dönüştürülmelidir. Lipoliz tarafından salınan FFA'ların muhtemelen geri alınmadan ve yağlı açil-CoA'ya dönüştürülmeden önce hücreden çıktığı görülmektedir 52,53,54,55,56,57,58,59,60,61,62 . FFA'lar albümin ile hücrenin dışında tutulabilir. Önemli olarak, uzun zincirli FFA'ların, albümin63,64,65,66,67 tarafından tutulmazlarsa lipolizi geri besleme-inhibe ettiği bilinmektedir. Bu nedenle, lipoliz testi sırasında ortamın FFA tamponlama kapasitesinin optimize edilmesi kritik öneme sahiptir. Burada açıklanan prosedür, farelerden ve insanlardan adipositlerde ve ex vivo yağ dokusunda lipolitik oranı ölçmek için daha önce yayınlanmış yöntemlere benzer 15,68,69,70,71. Bu protokol, seri örnekleme kullanımına göre farklılık gösterir; Seri örnekleme yaparak, lipolizin doğrusal fazda ölçüldüğünü dahili olarak doğrulayabilir ve lipoliz oranını hesaplamak için birden fazla ölçüm kullanabilir, böylece nihai hesaplanan değere olan güveni artırmak için ölçüm hatasını azaltabiliriz. Seri örneklemenin dezavantajı, tahlilin daha fazla zaman ve reaktif gerektirmesidir; Bununla birlikte, daha uzun zaman dilimi, ölçüm hatasının oran tahminlerinin standart hatası üzerindeki etkisini azaltır. Ek olarak, bu protokol hem FFA hem de gliserol salınımını ölçer ve FFA: gliserol salınımının oranını, tam lipoliz ve lipolitik ürünlerin medyaya salınmasından beklendiği gibi 3: 1 oranına ulaşmak amacıyla dikkate alır72.
Protokol
Tüm hayvanların kullanımı, Cornell Üniversitesi Weill Cornell Tıp Fakültesi'ndeki Kurumsal Hayvan Bakımı ve Kullanımı Komitesi (IACUC) tarafından onaylanmıştır.
1. Tamponların ve toplama plakalarının hazırlanması
- 5 g BSA'yı 100 mL Dulbecco'nun modifiye Eagle's medium (DMEM) içinde fenol kırmızısı olmadan çözerek% 5 sığır serum albümini (BSA) yapın. BSA'yı çözünmesi için hafifçe karıştırın (sallamak verimsizdir). BSA tamamen çözündükten sonra, ortamı 0,2 μm filtre ile filtreleyerek sterilize edin. BSA ortamını 4 °C'de 1 aya kadar saklayın.
- Kontrol ve stimülasyon ortamının çalışma konsantrasyonlarını yapın. Kontrol ortamı: Araç kontrollü %5 BSA ortamı. Stimülasyon ortamı: 0,5 μM CL-316.243 ile %5 BSA ortamı. Her deney için yeni stimülasyon ortamı oluşturun.
- Kullanılacak ortamı 37 °C'ye ısıtın. Medya koleksiyonu için 96 delikli bir plakayı etiketleyin.
2. Numune hazırlama
- Hücre kültürünü aşağıda açıklandığı gibi gerçekleştirin. Dış kontaminasyonu en aza indirmek için tüm hücre çalışmalarını steril bir duman davlumbazında üstlenin.
- Primer preadipositleri 73,74'te olduğu gibi izole edin ve ayırt edin.
- 1 mL/kuyucuklu kültür ortamında 24 delikli bir plakada 1 x 105 hücre/kuyu gibi yüksek yoğunlukta primer preadipositleri plakalayın (DMEM/F12'de %15 fetal sığır serumu (FBS) ve 1x penisilin-streptomisin-glutamin).
- Hücreler% 100 akıcılığa ulaştıktan sonra, 3 gün boyunca kültür ortamında 5 μM deksametazon, 0.5 mM 3-izobütil-1-metilksantin, 1 μg / mL insülin ve 1 μM tiazolidinedion (TZD) ile farklılaşır. Daha sonra, lipit damlacıklarını büyütmek için en az 3 gün boyunca 1 μg / mL insülin içeren kültür ortamına geçin. 24 delikli plakada 1 mL/kuyucuklu ortam kullanın.
- Kültür ortamını (1 ml / kuyu) her 2 veya 3 günde bir insülin ile değiştirin. Hücreler 2 haftaya kadar insülin içeren ortamlarda tutulabilir. Bu test için yalnızca farklılaşma oranlarının% 90'ın üzerinde olduğu ve gruplar arasında benzer olduğu kültürleri kullanın, çünkü azaltılmış farklılaşma lipolitik orandaki bir azalma olarak yanlış yorumlanabilir.
- Lipoliz ölçümünden önce 24 saat boyunca insülin içermeyen ortamda hücreleri kültürleyin.
NOT: Ortamdaki insülin lipit damlacıklarını korur, ancak aynı zamanda lipolizi de inhibe eder. 24 saat boyunca insülin olmadan inkübasyon, lipit damlacık hacminde bir kayıp olmadan tam lipolitik aktivasyona izin verir. Bazı sistemlerde, insülinsiz kültür süresinin kısaltılması veya uzatılması gerekebilir.
- Artık serumu kültür ortamından çıkarmak için hücreleri DPBS ile bir kez yıkayın.
NOT: Bu protokol, lipolizi aktive edebilen serum açlığını içermez. Serum açlığı, araştırmacının takdirine bağlı olarak kullanılabilir.
- Primer preadipositleri 73,74'te olduğu gibi izole edin ve ayırt edin.
- Aşağıda açıklandığı gibi ex vivo kültürü gerçekleştirin.
- Her fareden toplanacak her doku için bir kuyucuk içeren 6 kuyucuklu bir plaka hazırlayın. Kullanılacak her bir kuyucuğa 4 mL oda sıcaklığında DMEM yerleştirin.
NOT: Koleksiyon ortamında BSA gerekli değildir. - Lipoliz testi için 48 delikli bir plaka hazırlayın, her çoğaltma için bir kuyucuk hazırlayın. Kullanılacak her bir kuyucuğa 400 μL oda sıcaklığında DMEM yerleştirin. Fare başına doku başına iki ila dört kontrol ve iki ila dört uyarılmış kuyucuk kullanın.
- Bilateral pnömotoraks gibi ikincil bir yöntemle anestezi altında servikal çıkık ile fareyi ötenazi yapın. Burada, 4 ay boyunca% 45 yüksek yağlı bir diyetle beslenen 32 g, 7 aylık dişi bir C57BL / 6J fare kullandık.
NOT: Bu protokol erkeklerin yanı sıra diğer suşlar, diyetler ve yaşlar için de kullanılabilir. - % 70 etanol ile püskürtün ve karın derisinin merkezinde küçük (~ 1 cm) bir lateral kesi yapmak için makas kullanın, her iki tarafı başparmak ve işaret parmağıyla sıkıştırarak cildi ayırın ve posterior subkutan depoları ortaya çıkarmak için alt karın derisini katlayın. Kasık lenf nodunu bulup çıkarın ve forseps kullanarak inguinal lenf nodunun hemen arkasındaki inguinal yağ dokusunu künt diseke edin.
- Gonadal yağ dokusunu toplamak için, periton boşluğuna erişmek için peritonda lateral ve dikey bir kesi yapın. Gonadal yağ yastığını cımbızla tutun ve gonadal yağ dokusunu çıkarmak için uterus boyunca (veya erkekler için epididim) kesin. Toplanan depoları 6 delikli bir plakaya yerleştirin.
- Dokuyu iyice çıkarın, silikon bir paspas üzerine yerleştirin ve makasla 5 ila 7 mg'lık parçalar halinde kesin.
- Her bir tahlil kuyusu için 25 ila 30 mg (beş veya altı parça) tartın ve 48 kuyucuklu bir tahlil plakasına yerleştirin. Herhangi bir ortamı çıkarmak için tartmadan önce dokuyu temiz bir havlu üzerinde lekeleyin. Dokuyu çıkardıktan sonra ağırlık teknesini tartın ve geride kalan kalıntıların ağırlığını kaydedin. Numuneler arasında ağırlık teknesini temizleyin ve gerekirse tekrar dara yapın. Her doku için yeni bir ağırlık teknesi kullanın.
- Tüm doku örnekleri tartıldıktan sonra, 48 kuyucuklu tahlil plakasını 15 dakika boyunca 37 ° C,% 10 CO2 inkübatöre yerleştirin.
- Her fareden toplanacak her doku için bir kuyucuk içeren 6 kuyucuklu bir plaka hazırlayın. Kullanılacak her bir kuyucuğa 4 mL oda sıcaklığında DMEM yerleştirin.
3. Lipoliz tahlili
- Medya toplama işlemini gerçekleştirin. Dış kaynaklardan kaynaklanan potansiyel kontaminasyonu en aza indirmek için ortamın transferini ve ardından numunenin steril bir duman davlumbazında toplanmasını üstlenin.
- T = 0'da, ortamı çıkarın ve kontrol veya stimülasyon ortamının kuyucuğu başına 400 μL ekleyin ve plakayı 37 ° C,% 10 CO2 inkübatöre yerleştirin. Ex vivo doku kültürü için, bir pipet kullanarak ortamı dikkatlice çıkarın; emme asla kullanılmamalıdır.
NOT: Alternatif olarak, kontrol ve stimülasyon ortamı ile ikinci bir plaka hazırlayın ve dokuları aktarın. - t = 1, 2, 3 ve 4 h'de, 200 μL ortam toplayın, uygun kontrol veya stimülasyon ortamının 200 μL'si ile değiştirin ve tahlil plakasını inkübatöre geri getirin. Toplama plakasını 4 °C'de saklayın. BSA ortamının FFA arabelleğe alma kapasitesini belirlemek için, 24 saatte ek bir koleksiyon kullanın.
NOT: Deneyler burada durdurulabilir ve toplanan ortam -20 ° C'de saklanabilir.
- T = 0'da, ortamı çıkarın ve kontrol veya stimülasyon ortamının kuyucuğu başına 400 μL ekleyin ve plakayı 37 ° C,% 10 CO2 inkübatöre yerleştirin. Ex vivo doku kültürü için, bir pipet kullanarak ortamı dikkatlice çıkarın; emme asla kullanılmamalıdır.
4. FFA kolorimetrik tahlil
- Reaktifleri oda sıcaklığına ısıtın ve bir şişe renk reaktifi A'yı bir şişe çözücü A ile ve bir şişe renk reaktifi B'yi bir şişe çözücü B ile çözün. Sulandırma tarihinden itibaren, bu reaktifler en iyi 1 hafta içinde kullanılır. Sulandırmadan 1 ay sonra atın.
- Numuneleri çözün ve karıştırın.
- FFA standart eğrisi oluşturun. Standart çözüm 1 mM'dir. Standart eğri için reaktiflerle birlikte şu hacmi kullanın: maksimum aralık için 25 μL, 20 μL, 15 μL, 10 μL, 10 μL (1:2 seyreltme), 10 μL (1:4 seyreltme), 10 μL (1:8 seyreltme) ve 10 μL su. Düşük FFA seviyeleri için, 1 mM, 0,8 mM, 0,6 mM, 0,4 mM, 0,2 mM, 0,1 mM ve 0,05 mM'lik 10 μL standardı daha uygulanabilir olabilir.
- Pipet standartları ve numuneleri 96 delikli bir tahlil plakasına yerleştirin. Önerilen numune hacmi 10 μL'dir. Arka plan düzeltmesi için numunelerle aynı hacimde BSA ortamına sahip üç kuyucuk ekleyin.
NOT: Numune konsantrasyonları standart eğri aralığının dışına düşerse, numune hacmini 2-25 μL'ye ayarlayarak testi tekrarlayın. - Her bir oyuğa 150 μL reaktif A ekleyin ve karıştırın. Kabarcık oluşturmaktan kaçının. İnce bir gösterge iğnesi ile herhangi bir kabarcığı patlatın. Tahlil plakasını 37 ° C'de 5 dakika boyunca inkübe edin.
- Plakanın emilimini 550 nm ve 660 nm referansta okuyun (Okuma A).
- Her bir oyuğa 75 μL B reaktifi ekleyin ve karıştırın. Kabarcık oluşturmaktan kaçının. İnce bir gösterge iğnesi ile herhangi bir kabarcığı patlatın. Tahlil plakasını 37 ° C'de 5 dakika boyunca inkübe edin.
- Plakanın emilimini 550 nm ve 660 nm referansta tekrar okuyun (Okuma B).
5. Gliserol kolorimetrik tahlil
- Serbest gliserol reaktifini 36 mL ultra saf su ile yeniden oluşturun ve oda sıcaklığına alıştırın. Bu reaktifler en iyi birkaç hafta içinde kullanılır. Sulandırmadan 2 ay sonra atın.
- Numuneleri çözün ve karıştırın.
- Gliserol standart çözeltisinin yedi noktalı, 2 kat seri seyreltilmesini ve bir su boşluğunu yaparak bir gliserol standart eğrisi oluşturun.
NOT: Standart eğri, 2,8 mM gliserolün 25 μL'sine kadar nispeten doğrusaldır, ancak daha yüksek konsantrasyonlarda doğrusal değildir. - Pipet, her biri 25 μL standart ve numuneleri 96 delikli tahlil plakasına yerleştirin. Arka plan düzeltmesi için BSA ortamına üç kuyu ekleyin.
- Her bir oyuğa 175 μL serbest gliserol reaktifi ekleyin ve karıştırın. Kabarcık oluşturmaktan kaçının. İnce bir gösterge iğnesi ile herhangi bir kabarcığı patlatın. Tahlil plakasını 37 ° C'de 5 dakika boyunca inkübe edin.
- Plakanın emilimini 540 nm'de okuyun.
6. Lipolitik oranın hesaplanması
- Optik yoğunluk (OD) değerleriyle başlayın. Gliserol için, doğrudan A540 OD değerlerini kullanın. FFA testinin OD'sini aşağıdaki formüle göre hesaplayın:
OD = (Okuma B: A 550 - A 660) - (Okuma A: A550 - A660) - Toplanan örneklerdeki FFA ve gliserol seviyelerini hesaplamak için standart eğriyi kullanın. Standart OD değerlerini y ekseninde çizin ve x ekseninde, numune hacmine göre standart konsantrasyonlar kullanın (yani, 10 μL numuneli bir plaka üzerinde 20 μL 1 mM FFA standardına sahip kuyucukların konsantrasyonu 2 mM'ye eşittir). Doğrusal bir eğilim çizgisine uyun:
y = mx + b - Standart eğriyi görsel olarak inceleyin ve testin doğrusal aralığının dışındaki noktaları kaldırın. Denklemi kullanarak numune konsantrasyonlarını hesaplayın:
Numune konsantrasyonu: x = (OD - b) ÷ m - Doğrusal tahlil aralığının dışında kalan numuneleri ayarlayın ve yeniden analiz edin. Son numune konsantrasyonunu elde etmek için, numunelerin konsantrasyonundan sadece BSA ortamı içeren arka plan kuyucuklarının konsantrasyonunu çıkarın.
- Her numune tarafından üretilen FFA ve gliserol mollerini her zaman noktasında, formüle göre hesaplayın:

burada C n = t = n zamanındaki konsantrasyon; Vt = kuyudaki toplam hacim; Vs = numune toplama hacmi; ve M n = t = n zamanında üretilen moller (konsantrasyonlar mM cinsinden ve hacimler mL cinsinden olduğunda, çıktı μMol'dür).
Örneğin, çeşitli zaman noktalarında:
M 1 = C1 × Vt
M 4 = C4 × Vt + (C1 + C2+ C3) Vs
veya
M 4 = C4 × Vt + C3 × V s + C2 × Vs + C1 ×Vs - μmol/g birimleri elde etmek için her numune için gram cinsinden doku ağırlığına bölerek doku ağırlığına normalleştirin. Kültürlenmiş hücreler için değerler μmol/well olarak sunulur. Hücre sayısının ve farklılaşma verimliliğinin iyiden iyiye karşılaştırılabilir olduğundan emin olun.
NOT: Proliferasyon veya farklılaşma verimliliğindeki farklılıklar, sonuçların yorumlanmasını zorlaştıracak ve başka bir normalleştirme yöntemi gerektirecektir (örneğin, proteine normalleştirme; tartışmaya bakınız). - Her numune için üretilen μmol/g'nin (y ekseni) zamana (x ekseni) karşı eğimini ayrı ayrı hesaplayın.
- Bir elektronik tabloda, bu =SLOPE(known_ys,known_xs) işlevi kullanılarak yapılabilir. Yeni bir hücreye "=SLOPE" yazın (ardından imleci kullanarak örnek gliserol veya FFA değerlerini μmol/g cinsinden vurgulayın, ardından karşılık gelen zaman değerlerini vurgulayın).
- Verilerin doğrusallığını doğrulayın. R2 değerleri, numunelerin doğrusallığını belirlemenin hızlı bir yoludur. Bir elektronik tabloda bu, adım 6.7.1'de açıklandığı gibi =RSQ(known_ys,known_xs) işlevi kullanılarak yapılabilir, ancak ilk giriş =RSQ'dur. R2 değerlerinin > 0,98 olduğundan emin olun; Düşük değerler doğrusallıktan sapmayı gösterir. Bu, bir ölçüm/örnekleme hatasından veya doğrusallık kaybından kaynaklanabilir.
- Doğrusallığı test etmenin bir başka yolu da her örnek için doğrusal bir regresyon gerçekleştirmek ve artıkları çizmektir. İstatistiksel bir analiz yazılımında, her zaman noktası için tek bir Y değerine sahip bir XY tablosu oluşturun. Analyze > Simple linear regression (Basit doğrusal regresyonu analiz et) seçeneğini belirleyin ve OK (Tamam) düğmesine basmadan önce Residual Plot (Artık Grafik) kutusunu işaretleyin. Kalan grafik yeni bir grafik olarak görünür.
- İstatistiksel analiz yapmak için her numune için FFA ve gliserol üretim hızını (yani, eğim [(μmol / g / h]) ayrı bir veri noktası olarak kullanın ve farklı lipolitik koşullar karşılaştırılıyorsa değerleri çizin. Lipolitik oranlar genotipler arasında karşılaştırılıyorsa, teknik kopyalar olarak hayvan başına iki veya üç örnek kullanın ve hayvan başına bir veri noktası için ortalamayı kullanın, böylece örneklem büyüklüğü hayvan sayısına eşit olur.
Sonuçlar
İn vitro diferansiye adipositlerin bazal ve uyarılmış lipolitik oranlarını ölçtük. Kasık beyaz yağ dokusundan primer preadipositler, 5 μM deksametazon, 0.5 mM IBMX, 1 μg / mL insülin ve 1 μM troglitazon ile 4 gün boyunca konfluent hücrelerin tedavisi ile adipositlere farklılaştırıldı ve ardından 1 μg / mL insülin ile 3 günlük ek bir tedavi uygulandı. Hücreler lipoliz tahlilinden önce 24 saat boyunca insülinsülsüz ortamda inkübe edildi. Zaman = 0h'de, hücreler bir kez PBS ile y?...
Tartışmalar
Burada, adipositlerde ve ex vivo yağ dokusunda lipoliz oranını ölçmek için temel bir protokol sunuyoruz. Lipolizi ölçmek için, doğrusal fazda lipolitik hızı ölçmek önemlidir. Ortamın büyük bir kısmının toplandığı ve düzenli aralıklarla taze ortamlarla değiştirildiği seri örnekleme tekniği kullanıyoruz. Bu yarı konservatif yöntem, FFA tamponlama kapasitesine sahip taze BSA'nın eklenmesine izin verir ve lineer lipoliz süresini uzatarak geri besleme inhibisyonunu geciktirir. Bu d...
Açıklamalar
Yazarların açıklayacak hiçbir şeyi yok.
Teşekkürler
Bu çalışma, ABD Ulusal Sağlık Enstitüleri tarafından S.M.R.'ye R01DK126944 hibesi ile desteklenmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 24-Well tissue culture treated plate | Corning Inc | 3527 | Must be tissue culture treated for adipocyte differntiation |
| 48-Well flat bottom plate with lid | Corning Inc | 353078 | Can be tissue culture treated |
| 6-Well flat bottom plate with lid | Corning Inc | 353046 | Can be tissue culture treated |
| 96-Well PCR Plate | USA sceintific | 1402-9100 | Any conical 0.2 mL PCR plate will be convenient |
| Bovine Serum Albumin | Sigma Aldrich | A9418 | FFA free BSA such as A8806, is also commonly used. The BSA should not have detectable FFA, also lot to lot variations in BSA can impact the observed rate of lipolysis |
| CL-316,243 | Sigma Aldrich | C5976 | CAS #: 138908-40-4 availaible from other suppliers |
| CO2 incubator | PHCBI | MCO-170AICUVH | CO2 should ideally be set to 10% for adipose tissue, however 5% CO2 will also work |
| DMEM, low glucose, no phenol red | Thermofischer | 11054020 | Any phenol red free media should work, DMEM/F12, RPMI, but should contain volatile buffering capacity, i.e. biocarbonate |
| FFA-free Bovine serum albumin | Equitech-Bio, Inc, | BAH66 | |
| Free Glycerol Reagent | Sigma Aldrich | F6428 | |
| Glycerol Standard Solution | Sigma Aldrich | G7793 | This can also be made by diluting glycerol to the desired concentration |
| HR Series NEFA Standard Solution | Fujifilm | 276-76491 | |
| HR Series NEFA-HR (2) Color Reagent A | Fujifilm | 999-34691 | |
| HR Series NEFA-HR (2) Color Reagent B | Fujifilm | 991-34891 | |
| HR Series NEFA-HR (2) Solvent A | Fujifilm | 995-34791 | |
| HR Series NEFA-HR (2) Solvent B | Fujifilm | 993-35191 | |
| Microbiological Incubator | Fischer Scientific | S28668 | Any incubator at 37C can be used |
| Nunc MicroWell 96-Well Plates | Thermo Scientific | 269620 | Any optically clear, flat bottom 96-well plate works |
| Silicone Laboratory Benchtop Mat | VWR | 76045-300 | Glass plate can also be used. Absorbant surfaces are not recommended |
| Spectrophotometer/Microplate Reader | Molecular devices | SpectraMax i3x | Any plate reader that can read at 540, 550 and 660 mm will work |
| V Bovine serum albumin | Sigma-Aldrich | 810531 | |
| WypAll X70 Wipers | Kimberly-Clark | 41200 | Any high quality paper towel will work |
Referanslar
- Vaughan, M., Berger, J. E., Steinberg, D. Hormone-sensitive lipase and monoglyceride lipase activities in adipose tissue. The Journal of Biological Chemistry. 239, 401-409 (1964).
- Zimmermann, R., et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science. 306 (5700), 1383-1386 (2004).
- Lass, A., et al. Adipose triglyceride lipase-mediated lipolysis of cellular fat stores is activated by CGI-58 and defective in Chanarin-Dorfman syndrome. Cell Metabolism. 3 (5), 309-319 (2006).
- Stralfors, P., Bjorgell, P., Belfrage, P. Hormonal regulation of hormone-sensitive lipase in intact adipocytes: identification of phosphorylated sites and effects on the phosphorylation by lipolytic hormones and insulin. Proceedings of the National Academy of Sciences. 81 (11), 3317-3321 (1984).
- Miyoshi, H., et al. Perilipin promotes hormone-sensitive lipase-mediated adipocyte lipolysis via phosphorylation-dependent and -independent mechanisms. The Journal of Biological Chemistry. 281 (23), 15837-15844 (2006).
- Sztalryd, C., et al. Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. The Journal of Cell Biology. 161 (6), 1093-1103 (2003).
- Lafontan, M., Langin, D. Lipolysis and lipid mobilization in human adipose tissue. Progress in Lipid Research. 48 (5), 275-297 (2009).
- Grabner, G. F., Xie, H., Schweiger, M., Zechner, R. Lipolysis: cellular mechanisms for lipid mobilization from fat stores. Nature Metabolism. 3 (11), 1445-1465 (2021).
- Weiss, S. B., Kennedy, E. P., Kiyasu, J. Y. The enzymatic synthesis of triglycerides. The Journal of Biological Chemistry. 235, 40-44 (1960).
- Kennedy, E. P. Biosynthesis of complex lipids. Federation Proceedings. 20, 934-940 (1961).
- Wendel, A. A., Lewin, T. M., Coleman, R. A. Glycerol-3-phosphate acyltransferases: rate limiting enzymes of triacylglycerol biosynthesis. Biochimica et Biophysica Acta. 1791 (6), 501-506 (2009).
- Johansson, S. M., Lindgren, E., Yang, J. N., Herling, A. W., Fredholm, B. B. Adenosine A1 receptors regulate lipolysis and lipogenesis in mouse adipose tissue-interactions with insulin. European Journal of Pharmacology. 597 (1-3), 92-101 (2008).
- Gnad, T., et al. Adenosine activates brown adipose tissue and recruits beige adipocytes via A2A receptors. Nature. 516 (7531), 395-399 (2014).
- Fried, S. K., et al. Resistance to the antilipolytic effect of insulin in adipocytes of African-American compared to Caucasian postmenopausal women. Journal of Lipid Research. 51 (5), 1193-1200 (2010).
- Lee, M. J., Fried, S. K. Optimal protocol for the differentiation and metabolic analysis of human adipose stromal cells. Methods in Enzymology. 538, 49-65 (2014).
- Fricke, K., Heitland, A., Maronde, E. Cooperative activation of lipolysis by protein kinase A and protein kinase C pathways in 3T3-L1 adipocytes. Endocrinology. 145 (11), 4940-4947 (2004).
- Bergan, H. E., Kittilson, J. D., Sheridan, M. A. PKC and ERK mediate GH-stimulated lipolysis. Journal of Molecular Endocrinology. 51 (2), 213-224 (2013).
- Schmitz-Peiffer, C. The tail wagging the dog--regulation of lipid metabolism by protein kinase C. The FEBS Journal. 280 (21), 5371-5383 (2013).
- Carmen, G. Y., Victor, S. M. Signalling mechanisms regulating lipolysis. Cellular Signalling. 18 (4), 401-408 (2006).
- Zu, L., et al. Bacterial endotoxin stimulates adipose lipolysis via toll-like receptor 4 and extracellular signal-regulated kinase pathway. The Journal of Biological Chemistry. 284 (9), 5915-5926 (2009).
- Zhang, H. H., Halbleib, M., Ahmad, F., Manganiello, V. C., Greenberg, A. S. Tumor necrosis factor-alpha stimulates lipolysis in differentiated human adipocytes through activation of extracellular signal-related kinase and elevation of intracellular cAMP. Diabetes. 51 (10), 2929-2935 (2002).
- Tan, X., et al. TNF-α downregulates CIDEC via MEK/ERK pathway in human adipocytes. Obesity. 24 (5), 1070-1080 (2016).
- Laurencikiene, J., et al. NF-kappaB is important for TNF-alpha-induced lipolysis in human adipocytes. Journal of Lipid Research. 48 (5), 1069-1077 (2007).
- van Hall, G., et al. Interleukin-6 stimulates lipolysis and fat oxidation in humans. The Journal of Clinical Endocrinology and Metabolism. 88 (7), 3005-3010 (2003).
- Wueest, S., et al. Mesenteric fat lipolysis mediates obesity-associated hepatic steatosis and insulin resistance. Diabetes. 65 (1), 140-148 (2016).
- Trujillo, M. E., et al. Interleukin-6 regulates human adipose tissue lipid metabolism and leptin production in vitro. The Journal of Clinical Endocrinology and Metabolism. 89 (11), 5577-5582 (2004).
- Kitamura, T., et al. Insulin-induced phosphorylation and activation of cyclic nucleotide phosphodiesterase 3B by the serine-threonine kinase Akt. Molecular and Cellular Biology. 19 (9), 6286-6296 (1999).
- Chakrabarti, P., et al. Insulin inhibits lipolysis in adipocytes via the evolutionarily conserved mTORC1-Egr1-ATGL-mediated pathway. Molecular and Cellular Biology. 33 (18), 3659-3666 (2013).
- Collins, S., Daniel, K. W., Petro, A. E., Surwit, R. S. Strain-specific response to beta 3-adrenergic receptor agonist treatment of diet-induced obesity in mice. Endocrinology. 138 (1), 405-413 (1997).
- Surwit, R. S., Dixon, T. M., Petro, A. E., Daniel, K. W., Collins, S. Diazoxide restores beta3-adrenergic receptor function in diet-induced obesity and diabetes. Endocrinology. 141 (10), 3630-3637 (2000).
- Gettys, T. W., et al. Age-dependent changes in beta-adrenergic receptor subtypes and adenylyl cyclase activation in adipocytes from Fischer 344 rats. Endocrinology. 136 (5), 2022-2032 (1995).
- Mowers, J., et al. Inflammation produces catecholamine resistance in obesity via activation of PDE3B by the protein kinases IKKε and TBK1. eLife. 2, e01119 (2013).
- Valentine, J. M., et al. β3-Adrenergic receptor downregulation leads to adipocyte catecholamine resistance in obesity. The Journal of Clinical Investigation. 132 (2), e153357 (2022).
- Collins, S., et al. Impaired expression and functional activity of the beta 3- and beta 1-adrenergic receptors in adipose tissue of congenitally obese (C57BL/6J ob/ob) mice. Molecular Endocrinology. 8 (4), 518-527 (1994).
- Collins, S., Surwit, R. S. The beta-adrenergic receptors and the control of adipose tissue metabolism and thermogenesis. Recent Progress in Hormone Research. 56, 309-328 (2001).
- Dixon, T. M., Daniel, K. W., Farmer, S. R., Collins, S. CCAAT/enhancer-binding protein alpha is required for transcription of the beta 3-adrenergic receptor gene during adipogenesis. The Journal of Biological Chemistry. 276 (1), 722-728 (2001).
- Lohse, M. J., Benovic, J. L., Codina, J., Caron, M. G., Lefkowitz, R. J. beta-Arrestin: a protein that regulates beta-adrenergic receptor function. Science. 248 (4962), 1547-1550 (1990).
- Nantel, F., et al. The human beta 3-adrenergic receptor is resistant to short term agonist-promoted desensitization. Molecular Pharmacology. 43 (4), 548-555 (1993).
- Liggett, S. B., Freedman, N. J., Schwinn, D. A., Lefkowitz, R. J. Structural basis for receptor subtype-specific regulation revealed by a chimeric beta 3/beta 2-adrenergic receptor. Proceedings of the National Academy of Sciences. 90 (8), 3665-3669 (1993).
- Baker, J. G. The selectivity of beta-adrenoceptor agonists at human beta1-, beta2- and beta3-adrenoceptors. British Journal of Pharmacology. 160 (5), 1048-1061 (2010).
- Lafontan, M. Inhibition of epinephrine-induced lipolysis in isolated white adipocytes of aging rabbits by increased alpha-adrenergic responsiveness. Journal of Lipid Research. 20 (2), 208-216 (1979).
- Baker, J. G. The selectivity of beta-adrenoceptor antagonists at the human beta1, beta2 and beta3 adrenoceptors. British Journal of Pharmacology. 144 (3), 317-322 (2005).
- Jensen, M. D., Nielsen, S. Insulin dose response analysis of free fatty acid kinetics. Metabolism. 56 (1), 68-76 (2007).
- Jensen, M. D., Haymond, M. W., Gerich, J. E., Cryer, P. E., Miles, J. M. Lipolysis during fasting. Decreased suppression by insulin and increased stimulation by epinephrine. The Journal of Clinical Investigation. 79 (1), 207-213 (1987).
- Heckmann, B. L., et al. Defective adipose lipolysis and altered global energy metabolism in mice with adipose overexpression of the lipolytic inhibitor G0/G1 switch gene 2 (G0S2). The Journal of Biological Chemistry. 289 (4), 1905-1916 (2014).
- Shin, H., et al. Lipolysis in brown adipocytes is not essential for cold-induced thermogenesis in mice. Cell Metabolism. 26 (5), 764.e5-777.e5 (2017).
- Treble, D. H., Mayer, J. Glycerolkinase activity in white adipose tissue of obese-hyperglycaemic mice. Nature. 200, 363-364 (1963).
- Possik, E., et al. New mammalian glycerol-3-phosphate phosphatase: role in beta-cell, liver and adipocyte metabolism. Frontiers in Endocrinology. 12, 706607 (2021).
- Romero Mdel, M., Sabater, D., Fernandez-Lopez, J. A., Remesar, X., Alemany, M. Glycerol production from glucose and fructose by 3T3-L1 cells: a mechanism of adipocyte defense from excess substrate. PLoS One. 10 (10), e0139502 (2015).
- Mugabo, Y., et al. Identification of a mammalian glycerol-3-phosphate phosphatase: Role in metabolism and signaling in pancreatic beta-cells and hepatocytes. Proceedings of the National Academy of Sciences. 113 (4), E430-E439 (2016).
- Hanson, R. W., Reshef, L. Glyceroneogenesis revisited. Biochimie. 85 (12), 1199-1205 (2003).
- Vaughan, M. The production and release of glycerol by adipose tissue incubated in vitro. The Journal of Biological Chemistry. 237, 3354-3358 (1962).
- Jensen, M. D., Ekberg, K., Landau, B. R. Lipid metabolism during fasting. American Journal of Physiology-Endocrinology and Metabolism. 281 (4), E789-E793 (2001).
- Ballard, F. J., Hanson, R. W., Leveille, G. A. Phosphoenolpyruvate carboxykinase and the synthesis of glyceride-glycerol from pyruvate in adipose tissue. The Journal of Biological Chemistry. 242 (11), 2746-2750 (1967).
- Reshef, L., Hanson, R. W., Ballard, F. J. A possible physiological role for glyceroneogenesis in rat adipose tissue. The Journal of Biological Chemistry. 245 (22), 5979-5984 (1970).
- Gorin, E., Tal-Or, Z., Shafrir, E. Glyceroneogenesis in adipose tissue of fasted, diabetic and triamcinolone treated rats. European Journal of Biochemistry. 8 (3), 370-375 (1969).
- Elia, M., Zed, C., Neale, G., Livesey, G. The energy cost of triglyceride-fatty acid recycling in nonobese subjects after an overnight fast and four days of starvation. Metabolism. 36 (3), 251-255 (1987).
- Reshef, L., et al. Glyceroneogenesis and the triglyceride/fatty acid cycle. Journal of Biological Chemistry. 278 (33), 30413-30416 (2003).
- Edens, N. K., Leibel, R. L., Hirsch, J. Mechanism of free fatty acid re-esterification in human adipocytes in vitro. Journal of Lipid Research. 31 (8), 1423-1431 (1990).
- Vaughan, M., Steinberg, D. Effect of hormones on lipolysis and esterification of free fatty acids during incubation of adipose tissue in vitro. Journal of Lipid Research. 4, 193-199 (1963).
- Brooks, B., Arch, J. R., Newsholme, E. A. Effects of hormones on the rate of the triacylglycerol/fatty acid substrate cycle in adipocytes and epididymal fat pads. Federation of European Biochemical Societies Letters. 146 (2), 327-330 (1982).
- Bjorntorp, P., Karlsson, M., Hovden, A. Quantitative aspects of lipolysis and reesterification in human adipose tissue in vitro. Acta Medica Scandinavica. 185 (1-2), 89-97 (1969).
- Angel, A., Desai, K., Halperin, M. L. Free fatty acid and ATP levels in adipocytes during lipolysis. Metabolism. 20 (1), 87-99 (1971).
- Husted, A. S., et al. Autocrine negative feedback regulation of lipolysis through sensing of NEFAs by FFAR4/GPR120 in WAT. Molecular Metabolism. 42, 101103 (2020).
- Fain, J. N., Shepherd, R. E. Free fatty acids as feedback regulators of adenylate cyclase and cyclic 3':5'-AMP accumulation in rat fat cells. The Journal of Biological Chemistry. 250 (16), 6586-6592 (1975).
- Burns, T. W., Langley, P. E., Terry, B. E., Robinson, G. A. The role of free fatty acids in the regulation of lipolysis by human adipose tissue cells. Metabolism. 27 (12), 1755-1762 (1978).
- Kalderon, B., et al. Suppression of adipose lipolysis by long-chain fatty acid analogs. Journal of Lipid Research. 53 (5), 868-878 (2012).
- Schweiger, M., et al. Measurement of lipolysis. Methods in Enzymology. 538, 171-193 (2014).
- Decaunes, P., Bouloumie, A., Ryden, M., Galitzky, J. Ex vivo analysis of lipolysis in human subcutaneous adipose tissue explants. Bio-Protocol. 8 (3), e2711 (2018).
- Roy, D., Myers, J. M., Tedeschi, A. Protocol for assessing ex vivo lipolysis of murine adipose tissue. STAR Protocols. 3 (3), 101518 (2022).
- Baskaran, P., Thyagarajan, B. Measurement of basal and forskolin-stimulated lipolysis in inguinal adipose fat pads. Journal of Visualized Experiments. 125 (125), 55625 (2017).
- Reilly, S. M., et al. Catecholamines suppress fatty acid re-esterification and increase oxidation in white adipocytes via STAT3. Nature Metabolism. 2 (7), 620-634 (2020).
- Liu, L., et al. Isolation of mouse stromal vascular cells for monolayer culture. Methods in Molecular Biology. 1566, 9-16 (2017).
- DeLuca, J. H., Reilly, S. M. . Methods in Molecular Biology. , (2023).
- Richard, G., Vernon, R. A. C. New Perspectives in Adipose Tissue. Butterworth-Heinemann. , (1985).
- Brito, M. N., Botion, L. M., Brito, N. A., Kettelhut, I. C., Migliorini, R. H. Lipolysis and glycerokinase activity in brown adipose tissue of rat fed a high protein, carbohydrate-free diet. Hormone and Metabolic Research. 26 (1), 51-52 (1994).
- Bertin, R. Glycerokinase activity and lipolysis regulation in brown adipose tissue of cold acclimated rats. Biochimie. 58 (4), 431-434 (1976).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır