
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
מדידת שיעור הליפוליזה ברקמת השומן Ex Vivo Murine ובפראדיפוציטים ראשוניים מובחנים במבחנה
In This Article
Summary
טריגליצרידים ליפוליזה באדיפוציטים היא תהליך מטבולי חשוב המביא לשחרור חומצות שומן חופשיות וגליצרול. כאן, אנו מספקים פרוטוקול מפורט למדידת ליפוליזה בסיסית ומגורה באדיפוציטים וברקמת שומן ex vivo מעכברים.
Abstract
אדיפוציטים מאחסנים אנרגיה בצורה של טריגליצרידים בטיפות שומנים. אנרגיה זו יכולה להיות מגויסת באמצעות ליפוליזה, שבה שרשראות הצד של חומצות השומן נבקעות ברצף מעמוד השדרה של הגליצרול, וכתוצאה מכך משחררים חומצות שומן חופשיות וגליצרול. בשל הביטוי הנמוך של גליצרול קינאז באדיפוציטים לבנים, שיעורי הספיגה מחדש של גליצרול זניחים, בעוד ספיגה מחדש של חומצות שומן מוכתבת על ידי יכולת הקישור של חומצות שומן של רכיבי מדיה כגון אלבומין. ניתן לכמת הן גליצרול והן שחרור חומצות שומן למדיה על ידי בדיקות קולורימטריות כדי לקבוע את הקצב הליפוליטי. על ידי מדידת גורמים אלה בנקודות זמן מרובות, ניתן לקבוע את הקצב הליניארי של ליפוליזה בביטחון גבוה. כאן, אנו מספקים פרוטוקול מפורט למדידת ליפוליזה במבחנה אדיפוציטים מובחנים ורקמת שומן ex vivo מעכברים. פרוטוקול זה עשוי להיות מותאם גם עבור שורות תאים preadipocyte אחרים או רקמת שומן מאורגניזמים אחרים; שיקולים ופרמטרים אופטימיזציה נדונים. פרוטוקול זה נועד להיות שימושי בקביעה והשוואה של שיעור ליפוליזה אדיפוציט בין מודלים וטיפולים בעכברים.
Introduction
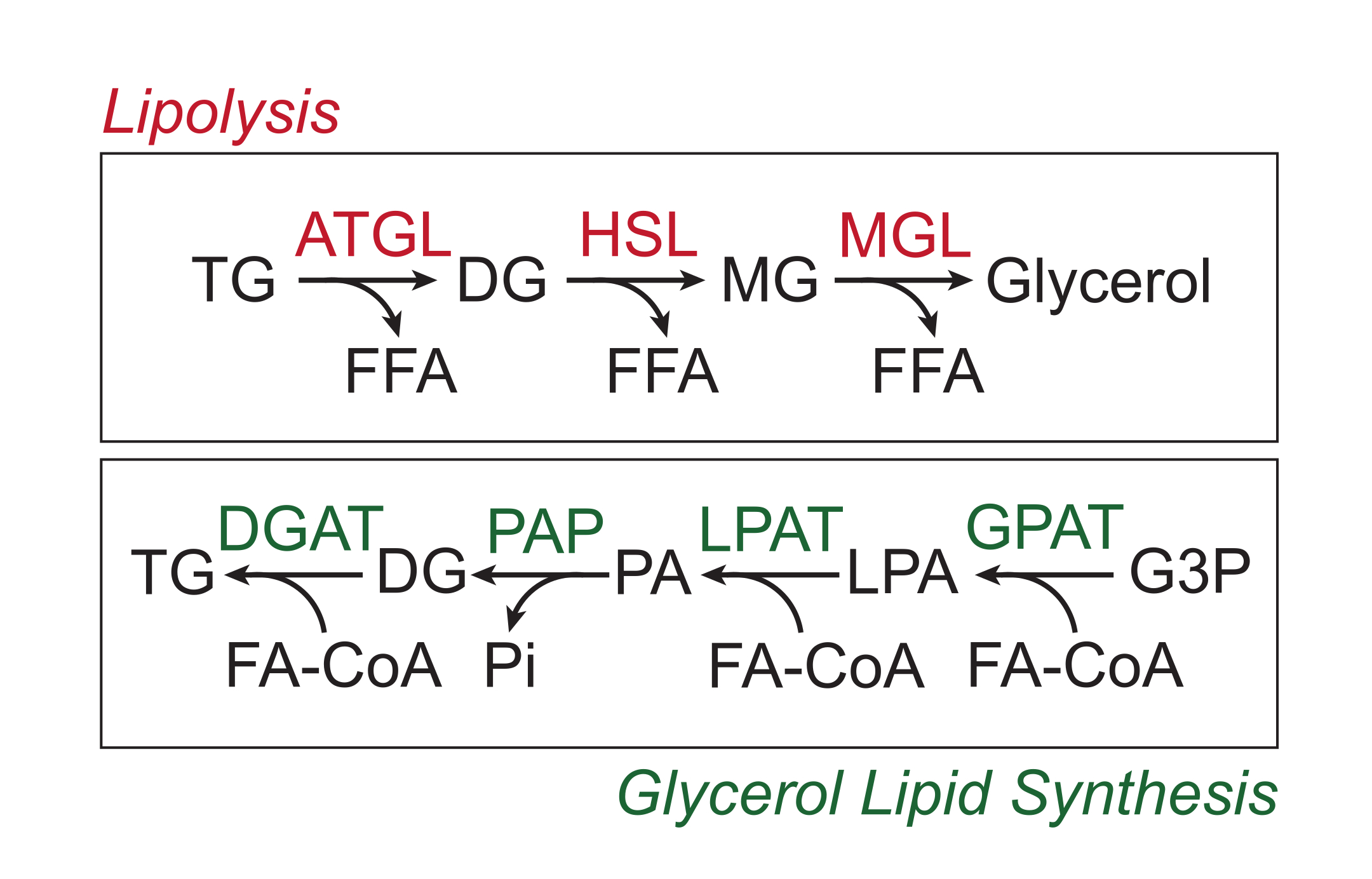
חומרים מזינים עודפים מאוחסנים ברקמת שומן לבן בצורה של טריגליצרידים בליבת השומנים הניטרלית של טיפות השומנים. מאגרי הטריגליצרידים מגויסים באמצעות ליפוליזה, תהליך שבו שרשראות הצד של חומצות השומן נבקעות ברצף על ידי ליפאז טריגליצרידים ברקמת השומן (ATGL), ליפאז רגיש להורמונים (HSL) ומונוגליצרידים ליפאז (MGL), וכתוצאה מכך משחררים חומצות שומן חופשיות (FFAs) ואת עמוד השדרה של גליצרול 1,2. ליפוליזה מופעלת על ידי איתות קטכולאמין ברקמת השומן. מסופי עצבים סימפתטיים משחררים קטכולאמינים באופן מקומי, הנקשרים לקולטנים β-אדרנרגיים על קרום הפלזמה האדיפוציטית. עם קשירת ליגנד, קולטנים מצומדים אלה של חלבון G (GPCRs) מפעילים ציקלאז אדניליל באמצעות Gαs. הפעלה עוקבת של חלבון קינאז A (PKA) על ידי cAMP גורמת להגברת הרגולציה הן של ATGL והן של HSL. הזרחן של perilipin-1 על ידי PKA גורם לדיסוציאציה של ABHD5 (הידוע גם בשם CGI-58), אשר קושר ומפעיל ATGL3. PKA ישירות phosphorylates HSL, קידום טרנסלוקציה שלה מן cytosol אל טיפת השומנים, שבו אינטראקציה עם phosphorylated perilipin-1 עוד יותר מקדם את פעילות ליפאז שלה 4,5,6,7. נראה כי הליפאז השלישי המעורב בליפוליזה, MGL, אינו מוסדר על ידי איתות קטכולאמין8. חשוב לציין, סינתזת טריגליצרידים באדיפוציטים מתווכת על ידי מסלול סינתזת השומנים גליצרול, שאינו כרוך בהיווצרות מונוגליצרידים כמתווך; במקום זאת, גליצרול-3-פוספט אציל טרנספראזות מזרז את היווצרותה של חומצה ליזופוספטידית, אשר משולבת עם אציל-CoA שומני אחר ליצירת חומצה פוספטידית, ולאחר מכן איזומריזציה לדיגליצרידים לפני הסינתזה הסופית של טריגליצרידים (איור 1)9,10,11.

איור 1: מסלולי סינתזת ליפוליזה וגליצרול. למעלה: מסלול ליפוליטי; אנזימים המוצגים באדום: טריגליצרידים ליפאז מרקמת השומן (ATGL), ליפאז רגיש להורמונים (HSL) ומונוגליצריד ליפאז (MGL). למטה: מסלול סינתזת שומנים גליצרול; אנזימים המוצגים בירוק: דיגליצריד אצילטרנספראז (DGAT), חומצה פוספטידית פוספטאז (PAP), חומצה ליזופוספטידית אצילטרנספראז (LPAT, הידוע גם בשם LPAATs) וגליצרול-3-פוספט אצילטרנספראז (GPAT). שומנים: טריגליצרידים (TG), דיגליצרידים (DG), מונוגליצרידים (MG), חומצת שומן חופשית (FFA), אציל-CoA שומני (FA-CoA), חומצה ליזופוספטידית (LPA) וחומצה פוספטידית (PA). מטבוליטים אחרים: פוספט אנאורגני (Pi) וגליצרול 3-פוספט (G3P). אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}
אדנוזין חוץ-תאי הוא וסת חשוב נוסף של ליפוליזה, הפועל באמצעות GPCRs מצומדים Gs ו-Gi כדי להשפיע על פעילות ציקלז אדניל. קולטן האדנוזין השולט באדיפוציטים, ADORA1, מעכב ציקלז אדניליל, ובכך ליפוליזה באמצעות הפעלת Gi12. ADORA2A, המתבטא ברמות נמוכות יותר, ובעיקר באדיפוציטים חומים, מפעיל ליפוליזה באמצעות איתות Gs 13. ADORA1 משפיע הן על ליפוליזה בסיסית והן על התגובה לאגוניסטים אדרנרגיים. ההשפעה של אדנוזין על ליפוליזה יכולה להיות נשלטת על ידי הוספת אדנוזין deaminase כדי לנטרל אדנוזין, כמו גם אגוניסט ספציפי ADORA1 phenylisopropyladenosine14,15. הפעלה הורמונלית של GPCRs מצומדים ל-Gq יכולה להשפיע גם על ליפוליזה באמצעות הפעלה של פוספוליפאז C וחלבון קינאז C16,17,18,19. אותות דלקתיים משפיעים גם על שיעורים ליפוליטיים. הפעלת TLR4 על ידי LPS (ואנדוטוקסינים אחרים) מגבירה את הקצב הליפוליטי על ידי הפעלת ERK, אשר פוספורילטים perilipin-1 ו- HSL20. TNF-α גם מפעיל ליפוליזה באמצעות הפעלת ERK ו- NF-κB, כמו גם הפחתת שעתוק של פוספודיאסטראז PDE-3B ו- CIDEC21,22,23. IL-6 נקשר גם לליפוליזה מוגברת של אדיפוציטים, במיוחד ברקמת שומן מזנטרית, ששחרור FFA שלה משפיע על סטאטוזיס בכבד וגלוקונאוגנזה24,25,26.
ליפוליזה מדוכאת במהלך מצב ההזנה על ידי אינסולין. AKT פוספורילטים ומפעיל PDE-3B כדי לדכא איתות cAMP ולמנוע הפעלת PKA27. אינסולין גם משעתק למטה ATGL28. השמנת יתר מקדמת עמידות לקטכולאמין באמצעות מגוון מנגנונים, כולל הפחתת ויסות של קולטני β-אדרנרגי באדיפוציטים 29,30,31,32,33. אדיפוציטים מבטאים את כל שלושת הקולטנים β-אדרנרגיים (β-1, β-2 ו-β-3). בעוד קולטנים אדרנרגיים β-1 ו-β-2 מתבטאים בכל מקום, הקולטן האדרנרגי β-3 מתבטא בעיקר באדיפוציטים בעכברים34,35. ביטוי Adrb3 מושרה על ידי C/EBPα במהלך אדיפוגנזה36. הקולטן האדרנרגי β-3 מתבטא מאוד באדיפוציטים בוגרים. ההפעלה של קולטנים אדרנרגיים β-1 ו-β-2 מגבילה את עצמה עקב עיכוב משוב על ידי β-arrestin37. עיכוב משוב של הקולטן האדרנרגי β-3 מתווך על ידי מסלולי איתות אחרים, המפחיתים את ביטוי Adrb3 33,38,39.
תרכובות רבות ניתן להשתמש כדי להפעיל ליפוליזה אדיפוציט. קטכולאמינים הם מפעילים פיזיולוגיים עיקריים של ליפוליזה. נוראפינפרין (או נוראדרנלין) ואפינפרין (או אדרנלין) מפעילים את כל שלושת קולטני β-אדרנרגי40. נוראפינפרין ואפינפרין משפיעים גם על ליפוליזה באמצעות הפעלת איתות קולטן α-אדרנרגי41. אגוניסטים נפוצים לקולטן β-אדרנרגי כוללים איזופרוטרנול, שהוא אגוניסט קולטן β-אדרנרגי לא סלקטיבי, ואגוניסטים לקולטן β-3 אדרנרגי CL-316,243 ומירבגרון42. בהתחשב בכך שהאדיפוציטים מבטאים בעיקר את הקולטן האדרנרגי β-3, אנו משתמשים ב-CL-316,243 כדוגמה כאן. הספציפיות שלו לקולטן האדרנרגי β-3 הופכת אותו גם למפעיל ספציפי יחסית של איתות קטכולאמין אדיפוציט, שניתן להשתמש בו בבטחה גם in vivo. שימו לב שהריכוז הנפוץ של 10 מיקרומטר CL-316,243 בתרבית תאים גבוה בסדרי גודל מהמינון ~0.1 מיקרומטר הנדרש להשגת תגובה מקסימלית33. פורסקולין עוקף את הקולטן האדרנרגי, מפעיל ישירות ציקלז אדניליל ואיתות ליפוליטי במורד הזרם. ישנם הרבה יותר activators, כמו גם מדכאים של ליפוליזה. בעת בחירת תרכובת לגירוי ליפוליזה, יש לשקול בזהירות את ספציפיות הקולטן ואת מסלולי האיתות במורד הזרם במסגרת תכנון הניסוי.
שיעור הליפוליזה ברקמת השומן הלבנה הוא גורם מטבולי חשוב המשפיע על סבילות לקור וזמינות חומרים מזינים במהלך צום או פעילות גופנית43,44,45,46. מטרת פרוטוקול זה היא למדוד את קצב הליפוליזה באדיפוציטים וברקמת השומן, מה שיקל על הבנת חילוף החומרים של האדיפוציט וכיצד הוא עשוי להשפיע על הפנוטיפ המטבולי של מודלים שונים של מורין. כדי לכמת את השיעור הליפוליטי, אנו מודדים את הופעתם של מוצרים ליפוליטיים בתקשורת (כלומר, FFAs וגליצרול). השיטה מסתמכת על שחרור מוצרים ליפוליטיים מהאדיפוציט לתקשורת. מכיוון שאדיפוציטים לבנים מבטאים רמות נמוכות של גליצרול קינאז, שיעורי ספיגה חוזרת של גליצרול נמוכים47. לעומת זאת, יש לשקול גם את הייצור של FFAs וגליצרול על ידי מסלולים מטבוליים שאינם ליפוליזה. נראה כי אדיפוציטים מבטאים פוספטאז עם פעילות נגד גליצרול-3 פוספט, מה שמאפשר ייצור גליצרול מגליצרול-3-פוספט המופק מגלוקוז48,49,50. גליקוליזה היא מקור לגליצרול-3-פוספט המשמש לאסטריפיקציה מחדש של FFA באדיפוציטים לבנים. כאשר רמות הגלוקוז מוגבלות, גליצרונוגנזה דורשת מקורות אחרים של 3 פחמן, כגון לקטט ופירובט51. התקשור של FFAs המשוחררים על ידי ליפוליזה בתוך התא וגורלם המטבולי אינו מובן היטב; FFAs המשוחררים על ידי ליפוליזה חייבים להיות מומרים לאציל-CoA שומני, לפני שהם עוברים אסטריפיקציה מחדש או עוברים חמצון β. נראה כי FFAs המשוחררים על ידי ליפוליזה ככל הנראה יוצאים מהתא לפני שהם נלקחים בחזרה ומומרים לשומן acyl-CoA 52,53,54,55,56,57,58,59,60,61,62 . FFAs יכולים להיות מבודדים מחוץ לתא על ידי אלבומין. חשוב לציין, FFAs ארוכי שרשרת ידועים כמעכבים ליפוליזה אם הם אינם מוחזקים על ידי אלבומין 63,64,65,66,67. לכן, אופטימיזציה של יכולת האגירה FFA של התקשורת במהלך בדיקת ליפוליזה היא קריטית. ההליך המתואר כאן דומה לשיטות שפורסמו בעבר למדידת השיעור הליפוליטי באדיפוציטים וברקמת שומן ex vivo מעכברים ובני אדם 15,68,69,70,71. פרוטוקול זה שונה באמצעות שימוש בדגימה סדרתית; על ידי ביצוע דגימה סדרתית, אנו יכולים לאמת פנימית שליפוליזה נמדדת בפאזה ליניארית ולהשתמש במדידות מרובות כדי לחשב את קצב הליפוליזה, ובכך להפחית את טעות המדידה כדי להגביר את הביטחון בערך המחושב הסופי. החיסרון של דגימה סדרתית הוא כי הבדיקה דורשת יותר זמן ריאגנטים; עם זאת, מסגרת הזמן הארוכה יותר מפחיתה את השפעת טעות המדידה על שגיאת התקן של אומדני הקצב. בנוסף, פרוטוקול זה מודד הן FFA והן שחרור גליצרול, ומשקלל את היחס בין שחרור FFA:גליצרול במטרה להשיג יחס של 3:1, כפי שניתן היה לצפות מליפוליזה מלאה ושחרור מוצרים ליפוליטיים למדיה72.
Protocol
השימוש בכל בעלי החיים אושר על ידי הוועדה המוסדית לטיפול ושימוש בבעלי חיים (IACUC) בקולג' הרפואי וייל קורנל באוניברסיטת קורנל.
1. הכנת חוצצים וצלחות איסוף
- הכינו אלבומין בסרום בקר 5% (BSA) על ידי המסת 5 גרם BSA ב-100 מ"ל של מדיום הנשר המעובד של דולבקו (DMEM) ללא פנול אדום. ערבבו בעדינות את ה-BSA כדי להתמוסס (ניעור אינו מועיל). לאחר המסת ה-BSA במלואה, יש לעקר את המדיה באמצעות מסנן של 0.2 מיקרומטר. אחסן את מדיית ה- BSA ב- 4 ° C למשך עד חודש אחד.
- הפוך את העבודה לריכוזי בקרה וגירוי. מדיית בקרה: 5% מדיית BSA עם בקרת רכב. אמצעי גירוי: 5% חומרי הדפסה BSA עם 0.5 מיקרומטר CL-316,243. צור מדיה חדשה לגירוי עבור כל ניסוי.
- יש לחמם את המדיה לשימוש בטמפרטורה של 37°C. תייגו צלחת של 96 בארות לאיסוף מדיה.
2. הכנת מדגם
- בצע תרבית תאים כמתואר להלן. בצע את כל עבודת התאים במכסה אדים סטרילי כדי למזער זיהום חיצוני.
- בודד והבדיל בין פרדיפוציטים ראשוניים, כמו ב-73,74.
- צלחת preadipocytes ראשוני בצפיפות גבוהה, כגון 1 x 105 תאים / באר בצלחת 24 באר ב 1 מ"ל / באר תרבית מדיה (15% סרום בקר עוברי (FBS) ו 1x פניצילין-סטרפטומיצין-גלוטמין ב DMEM/F12).
- לאחר שהתאים מגיעים למפגש של 100%, התמינו עם 5 מיקרומטר דקסמתזון, 0.5 מילימטר 3-איזובוטיל-1-מתיל-קסנתין, 1 מיקרוגרם / מ"ל אינסולין, ו 1 מיקרומטר תיאזולידינדיון (TZD) במדיה תרבית במשך 3 ימים. לאחר מכן, יש לעבור לתרבית עם 1 מיקרוגרם/מ"ל אינסולין למשך 3 ימים לפחות כדי לגדל טיפות שומנים. יש להשתמש ב-1 מ"ל/באר של מדיה בצלחת בעלת 24 הקידוחים.
- שנה את מצע התרבית (1 מ"ל / באר) עם אינסולין כל 2 או 3 ימים. התאים יכולים להישמר במדיה עם אינסולין עד שבועיים. השתמש רק בתרבויות שבהן שיעורי הבידול הם מעל 90% והם דומים בין קבוצות עבור בדיקה זו, שכן דיפרנציאציה מופחתת עלולה להתפרש בטעות כירידה בשיעור הליפוליטי.
- תרבית את התאים במדיה נטולת אינסולין במשך 24 שעות לפני מדידת ליפוליזה.
הערה: אינסולין בתקשורת שומר על טיפות שומנים, אך גם מעכב ליפוליזה. דגירה ללא אינסולין למשך 24 שעות מאפשרת הפעלה ליפוליטית מלאה ללא אובדן נפח טיפת השומנים. במערכות מסוימות, ייתכן שיהיה צורך לקצר או להאריך את זמן התרבית ללא אינסולין.
- שטפו את התאים עם DPBS פעם אחת כדי להסיר שאריות סרום מתרבית.
הערה: פרוטוקול זה אינו כולל רעב בסרום, אשר יכול להפעיל ליפוליזה. ניתן להשתמש בהרעבת נסיוב על פי שיקול דעתו של החוקר.
- בודד והבדיל בין פרדיפוציטים ראשוניים, כמו ב-73,74.
- בצע תרבות ex vivo כמתואר להלן.
- הכינו צלחת בת 6 בארות, עם באר אחת לכל רקמה שתיאסף מכל עכבר. יש להניח 4 מ"ל DMEM בטמפרטורת החדר בכל באר לשימוש.
הערה: BSA במדיית האיסוף אינו הכרחי. - הכינו צלחת של 48 בארות לבדיקת הליפוליזה, עם באר אחת לכל שכפול. יש להניח 400 μL של DMEM בטמפרטורת החדר בכל באר לשימוש. השתמש בשתיים עד ארבע בקרות ושתיים עד ארבע בארות מגורות לכל רקמה לכל עכבר.
- להרדים את העכבר על ידי נקע צוואר הרחם תחת הרדמה, עם שיטה משנית כגון pneumothorax דו צדדי. כאן, השתמשנו בעכבר נקבה C57BL/6J בן 32 גרם, 7 חודשים, שהוזן בתזונה עתירת שומן של 45% במשך 4 חודשים.
הערה: פרוטוקול זה יכול לשמש גם עבור גברים, כמו גם זנים, דיאטות וגילאים אחרים. - רססו עם 70% אתנול והשתמשו במספריים כדי לבצע חתך רוחבי קטן (~ 1 ס"מ) במרכז עור הבטן, פרקו את העור על ידי צביטה משני הצדדים עם האגודל והאצבע וקפלו את עור הבטן התחתונה כדי לחשוף את המחסנים התת עוריים האחוריים. אתר והסר את בלוטת הלימפה המפשעתית והקהה נתח את רקמת השומן המפשעתית מיד אחורית לבלוטת הלימפה המפשעתית באמצעות מלקחיים.
- כדי לאסוף את רקמת השומן גונדלי, לבצע חתך לרוחב אנכי בצפק כדי לגשת לחלל הצפק. החזיקו את כרית שומן הגונדל בפינצטה וחתכו לאורך הרחם (או אפידידימיס לזכרים) כדי להסיר את רקמת השומן הגונדלית. מניחים את המחסנים שנאספו בצלחת בת 6 בארות.
- מוציאים את הרקמה היטב, מניחים על משטח סיליקון וחותכים לגושים של 5 עד 7 מ"ג עם מספריים.
- שקלו היטב 25 עד 30 מ"ג (חמישה או שישה גושים) עבור כל בדיקה והכניסו לצלחת בדיקה של 48 בארות. כתם את הרקמה על מגבת נקייה לפני השקילה כדי להסיר מדיה כלשהי. שקול את סירת המשקולות לאחר הסרת הרקמה ורשום את המשקל של כל השאריות שנותרו מאחור. נקו את סירת המשקולות בין הדגימות וטפחו מחדש במידת הצורך. השתמש בסירת משקל חדשה עבור כל רקמה.
- לאחר שכל דגימות הרקמה נשקלו, הניחו את צלחת הבדיקה 48 בארות באינקובטור של 37 מעלות צלזיוס, 10% CO2 למשך 15 דקות.
- הכינו צלחת בת 6 בארות, עם באר אחת לכל רקמה שתיאסף מכל עכבר. יש להניח 4 מ"ל DMEM בטמפרטורת החדר בכל באר לשימוש.
3. בדיקת ליפוליזה
- ביצוע איסוף מדיה. לבצע העברה של המדיה ואיסוף הדגימות לאחר מכן במכסה אדים סטרילי כדי למזער זיהום פוטנציאלי ממקורות חיצוניים.
- ב t = 0, להסיר את המדיה ולהוסיף 400 μL לכל באר של מדיה בקרה או גירוי, ומניחים את הבדיקה לתוך 37 ° C, 10% CO2 אינקובטור. עבור תרבית רקמות ex vivo , בזהירות להסיר מדיה באמצעות פיפטה; אין להשתמש ביניקה.
הערה: לחלופין, הכינו צלחת שנייה עם אמצעי בקרה וגירוי, והעבירו את הרקמות. - ב t = 1, 2, 3 ו 4 שעות, לאסוף 200 μL של מדיה, להחליף עם 200 μL של אמצעי הבקרה או גירוי המתאים, ולהחזיר את צלחת הבדיקה לאינקובטור. אחסנו את צלחת האיסוף בטמפרטורה של 4°C. כדי לקבוע את קיבולת האגירה של FFA של מדיית BSA, השתמש באוסף נוסף בשעה 24 שעות.
הערה: ניתן לעצור את הניסויים כאן, ואת המדיה שנאספה ניתן לאחסן ב -20 ° C.
- ב t = 0, להסיר את המדיה ולהוסיף 400 μL לכל באר של מדיה בקרה או גירוי, ומניחים את הבדיקה לתוך 37 ° C, 10% CO2 אינקובטור. עבור תרבית רקמות ex vivo , בזהירות להסיר מדיה באמצעות פיפטה; אין להשתמש ביניקה.
4. בדיקת FFA colorimetric
- מחממים את הריאגנטים לטמפרטורת החדר וממיסים בקבוק אחד של מגיב צבע A עם בקבוק אחד של ממס A, ובקבוק אחד של מגיב צבע B עם בקבוק אחד של ממס B. מתאריך החוקה מחדש, ריאגנטים אלה משמשים בצורה הטובה ביותר בתוך שבוע אחד. יש להשליך חודש לאחר החוקה מחדש.
- מפשירים ומערבבים את הדגימות.
- צור עקומה סטנדרטית של FFA. הפתרון הסטנדרטי הוא 1 mM. השתמש בנפח הבא עם הריאגנטים עבור העקומה הסטנדרטית: 25 μL, 20 μL, 15 μL, 10 μL, 10 μL (דילול 1:2), 10 μL (דילול 1:4), 10 μL (דילול 1:8) ו- 10 μL מים לטווח מרבי. עבור רמות FFA נמוכות, תקן 10 μL של 1 mM, 0.8 mM, 0.6 mM, 0.4 mM, 0.2 mM, 0.1 mM ו- 0.05 mM עשוי להיות ישים יותר.
- פיפטה סטנדרטית ודגימות לתוך צלחת בדיקה 96 באר. נפח הדגימה המומלץ הוא 10 μL. כלול שלוש בארות עם נפח זהה של מדיה BSA כמו הדגימות לתיקון רקע.
הערה: אם ריכוזי הדגימה חורגים מטווח העקומה הסטנדרטית, חזור על הבדיקה והתאמת נפח הדגימה ל- 2-25 μL. - מוסיפים 150 μL של מגיב A לכל באר ומערבבים. הימנעו מיצירת בועות. פוצצו את כל הבועות בעזרת מחט מד עדינה. לדגור את צלחת הבדיקה ב 37 ° C במשך 5 דקות.
- קרא את ספיגת הצלחת בסימוכין של 550 ננומטר ו- 660 ננומטר (קריאה A).
- מוסיפים 75 μL של מגיב B לכל באר ומערבבים. הימנעו מיצירת בועות. פוצצו את כל הבועות בעזרת מחט מד עדינה. לדגור את צלחת הבדיקה ב 37 ° C במשך 5 דקות.
- קרא שוב את ספיגת הצלחת בנקודת ייחוס של 550 ננומטר ו- 660 ננומטר (קריאה B).
5. בדיקת גליצרול קולורימטרי
- הרכיבו מחדש את מגיב הגליצרול החופשי עם 36 מ"ל מים טהורים במיוחד והתאקלמו לטמפרטורת החדר. ריאגנטים אלה משמשים בצורה הטובה ביותר בתוך מספר שבועות. יש להשליך חודשיים לאחר החוקה מחדש.
- מפשירים ומערבבים את הדגימות.
- צור עקומה סטנדרטית של גליצרול על ידי ביצוע דילול סדרתי של שבע נקודות, פי 2 של תמיסת הגליצרול הסטנדרטית וריק במים.
הערה: העקומה הסטנדרטית היא ליניארית יחסית עד 25 μL של 2.8 מילימטר גליצרול, אך לא ליניארית בריכוזים גבוהים יותר. - פיפטה 25 μL כל אחד של תקן דגימות לתוך צלחת הבדיקה 96-well. כלול שלוש בארות עם מדיית BSA לתיקון רקע.
- מוסיפים 175 μL של מגיב גליצרול חופשי לכל באר ומערבבים. הימנעו מיצירת בועות. פוצצו את כל הבועות בעזרת מחט מד עדינה. לדגור את צלחת הבדיקה ב 37 ° C במשך 5 דקות.
- קרא את ספיגת הצלחת ב 540 ננומטר.
6. חישוב שיעור ליפוליטי
- התחל עם ערכי צפיפות אופטית (OD). עבור גליצרול, השתמש ישירות בערכי ODA 540 . חשב את OD של בדיקת FFA לפי הנוסחה הבאה:
OD = (קריאה B: A 550 - A 660) - (קריאה A: A550 - A660) - השתמש בעקומה הסטנדרטית כדי לחשב את רמות ה- FFA והגליצרול בדגימות שנאספו. התווה את ערכי OD הסטנדרטיים על ציר y, ועל ציר x, השתמש בריכוזים סטנדרטיים ביחס לנפח הדגימה (כלומר, ריכוז הבארות עם 20 μL של תקן FFA של 1 mM על צלחת עם דגימות 10 μL שווה ל 2 mM). התאם לקו מגמה ליניארי:
y = mx + b - בדוק חזותית את העקומה הסטנדרטית והסר נקודות מחוץ לטווח הליניארי של הבדיקה. חישוב ריכוזי מדגם באמצעות המשוואה:
ריכוז מדגם: x = (OD - b) ÷ m - התאם ובחן מחדש דגימות שאינן בטווח הבדיקה הליניארית. כדי לקבל את ריכוז הדגימה הסופי, הפחת את ריכוז בארות הרקע המכילות רק מדיה BSA מריכוז הדגימות.
- חישוב השומות של FFA וגליצרול המיוצרות על ידי כל מדגם בכל נקודת זמן, על פי הנוסחה:

כאשר C n = ריכוז בזמן t = n; Vt = נפח כולל בבאר; Vs = נפח איסוף המדגם; ו- M n = שומות המיוצרות בזמן t = n (כאשר הריכוזים הם ב- mM והנפחים הם ב- mL הפלט הוא μMol).
לדוגמה, בנקודות זמן שונות:
M 1 = C1 × Vt
M 4 = C4 × Vt + (C1 + C2+ C3)Vs
או
M 4 = C4 × Vt + C3 × V s + C2 × V s + C1 ×V s - נרמלו למשקל הרקמה על ידי חלוקה במשקל הרקמה עבור כל דגימה בגרמים לקבלת יחידות של μmol/g. עבור תאים בתרבית, הערכים מוצגים כ-μmol/well. ודא שמספר התא ויעילות ההתמיינות דומים מבאר לבאר.
הערה: הבדלים ביעילות ההתרבות או ההתמיינות יסבכו את פירוש התוצאות ויחייבו שיטה אחרת של נורמליזציה (למשל, נורמליזציה לחלבון; ראו דיון). - חשב את השיפוע של μmol/g שנוצר (ציר y) לעומת זמן (ציר x) עבור כל דגימה בנפרד.
- בגיליון אלקטרוני, ניתן לעשות זאת באמצעות הפונקציה =SLOPE(known_ys,known_xs). בתא חדש, הקלד "=SLOPE" (ולאחר מכן השתמש בסמן כדי להדגיש את ערכי הגליצרול או FFA לדוגמה ב- μmol/g, ולאחר מכן כדי להדגיש את ערכי הזמן המתאימים).
- אמת את הליניאריות של הנתונים. ערכי R2 הם דרך מהירה לקבוע ליניאריות של הדגימות. בגיליון אלקטרוני, ניתן לעשות זאת באמצעות הפונקציה =RSQ(known_ys,known_xs), באותו אופן המתואר בשלב 6.7.1, אך הקלט הראשוני הוא =RSQ. ודא שערכי R2 הם >- 0.98; ערכים נמוכים יותר מצביעים על סטייה מלינאריות. הדבר יכול לנבוע משגיאת מדידה/דגימה או מאובדן ליניאריות.
- דרך נוספת לבדוק ליניאריות היא לבצע רגרסיה ליניארית עבור כל דגימה ולשרטט את השאריות. בתוכנת ניתוח סטטיסטי, צור טבלת XY עם ערך Y יחיד עבור כל נקודת זמן. בחר נתח > רגרסיה ליניארית פשוטה ובחר את התיבה עבור העלילה השיורית לפני שתלחץ על אישור. התרשים השיורי יופיע כגרף חדש.
- השתמש בקצב ייצור FFA וגליצרול (כלומר, שיפוע [(μmol/g/h]) עבור כל דגימה כנקודת נתונים נפרדת לביצוע ניתוח סטטיסטי, וערכי תרשים אם משווים תנאים ליפוליטיים שונים. אם משווים שיעורים ליפוליטיים בין גנוטיפים, השתמשו בשתיים או שלוש דגימות לכל חיה כשכפולים טכניים, והשתמשו בממוצע עבור נקודת נתונים אחת לכל חיה, כך שגודל הדגימה יהיה שווה למספר בעלי החיים.
תוצאות
מדדנו את הקצב הליפוליטי הבסיסי והמגורה של אדיפוציטים ממוינים במבחנה. קדם-אדיפוציטים ראשוניים מרקמת השומן הלבן המפשעה התמינו לאדיפוציטים על ידי טיפול בתאים מתמזגים עם 5 מיקרומטר דקסמתזון, 0.5 מילימטר IBMX, 1 מיקרוגרם / מ"ל אינסולין ו -1 מיקרומטר טרוגליטזון במשך 4 ימים, ולאחר מכן טיפול נוסף ש...
Discussion
כאן, אנו מספקים פרוטוקול בסיסי למדידת שיעור הליפוליזה באדיפוציטים וברקמת השומן ex vivo . כדי לכמת ליפוליזה, חשוב למדוד את הקצב הליפוליטי בשלב הלינארי. אנו משתמשים בטכניקת דגימה סדרתית, שבה חלק גדול מהמדיה נאסף ומוחלף במדיה טרייה במרווחי זמן קבועים. שיטה שמרנית למחצה זו מאפשרת תוספת של BSA ...
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
עבודה זו נתמכה על ידי מענק R01DK126944 של המכונים הלאומיים לבריאות בארה"ב ל- S.M.R.
Materials
| Name | Company | Catalog Number | Comments |
| 24-Well tissue culture treated plate | Corning Inc | 3527 | Must be tissue culture treated for adipocyte differntiation |
| 48-Well flat bottom plate with lid | Corning Inc | 353078 | Can be tissue culture treated |
| 6-Well flat bottom plate with lid | Corning Inc | 353046 | Can be tissue culture treated |
| 96-Well PCR Plate | USA sceintific | 1402-9100 | Any conical 0.2 mL PCR plate will be convenient |
| Bovine Serum Albumin | Sigma Aldrich | A9418 | FFA free BSA such as A8806, is also commonly used. The BSA should not have detectable FFA, also lot to lot variations in BSA can impact the observed rate of lipolysis |
| CL-316,243 | Sigma Aldrich | C5976 | CAS #: 138908-40-4 availaible from other suppliers |
| CO2 incubator | PHCBI | MCO-170AICUVH | CO2 should ideally be set to 10% for adipose tissue, however 5% CO2 will also work |
| DMEM, low glucose, no phenol red | Thermofischer | 11054020 | Any phenol red free media should work, DMEM/F12, RPMI, but should contain volatile buffering capacity, i.e. biocarbonate |
| FFA-free Bovine serum albumin | Equitech-Bio, Inc, | BAH66 | |
| Free Glycerol Reagent | Sigma Aldrich | F6428 | |
| Glycerol Standard Solution | Sigma Aldrich | G7793 | This can also be made by diluting glycerol to the desired concentration |
| HR Series NEFA Standard Solution | Fujifilm | 276-76491 | |
| HR Series NEFA-HR (2) Color Reagent A | Fujifilm | 999-34691 | |
| HR Series NEFA-HR (2) Color Reagent B | Fujifilm | 991-34891 | |
| HR Series NEFA-HR (2) Solvent A | Fujifilm | 995-34791 | |
| HR Series NEFA-HR (2) Solvent B | Fujifilm | 993-35191 | |
| Microbiological Incubator | Fischer Scientific | S28668 | Any incubator at 37C can be used |
| Nunc MicroWell 96-Well Plates | Thermo Scientific | 269620 | Any optically clear, flat bottom 96-well plate works |
| Silicone Laboratory Benchtop Mat | VWR | 76045-300 | Glass plate can also be used. Absorbant surfaces are not recommended |
| Spectrophotometer/Microplate Reader | Molecular devices | SpectraMax i3x | Any plate reader that can read at 540, 550 and 660 mm will work |
| V Bovine serum albumin | Sigma-Aldrich | 810531 | |
| WypAll X70 Wipers | Kimberly-Clark | 41200 | Any high quality paper towel will work |
References
- Vaughan, M., Berger, J. E., Steinberg, D. Hormone-sensitive lipase and monoglyceride lipase activities in adipose tissue. The Journal of Biological Chemistry. 239, 401-409 (1964).
- Zimmermann, R., et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science. 306 (5700), 1383-1386 (2004).
- Lass, A., et al. Adipose triglyceride lipase-mediated lipolysis of cellular fat stores is activated by CGI-58 and defective in Chanarin-Dorfman syndrome. Cell Metabolism. 3 (5), 309-319 (2006).
- Stralfors, P., Bjorgell, P., Belfrage, P. Hormonal regulation of hormone-sensitive lipase in intact adipocytes: identification of phosphorylated sites and effects on the phosphorylation by lipolytic hormones and insulin. Proceedings of the National Academy of Sciences. 81 (11), 3317-3321 (1984).
- Miyoshi, H., et al. Perilipin promotes hormone-sensitive lipase-mediated adipocyte lipolysis via phosphorylation-dependent and -independent mechanisms. The Journal of Biological Chemistry. 281 (23), 15837-15844 (2006).
- Sztalryd, C., et al. Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. The Journal of Cell Biology. 161 (6), 1093-1103 (2003).
- Lafontan, M., Langin, D. Lipolysis and lipid mobilization in human adipose tissue. Progress in Lipid Research. 48 (5), 275-297 (2009).
- Grabner, G. F., Xie, H., Schweiger, M., Zechner, R. Lipolysis: cellular mechanisms for lipid mobilization from fat stores. Nature Metabolism. 3 (11), 1445-1465 (2021).
- Weiss, S. B., Kennedy, E. P., Kiyasu, J. Y. The enzymatic synthesis of triglycerides. The Journal of Biological Chemistry. 235, 40-44 (1960).
- Kennedy, E. P. Biosynthesis of complex lipids. Federation Proceedings. 20, 934-940 (1961).
- Wendel, A. A., Lewin, T. M., Coleman, R. A. Glycerol-3-phosphate acyltransferases: rate limiting enzymes of triacylglycerol biosynthesis. Biochimica et Biophysica Acta. 1791 (6), 501-506 (2009).
- Johansson, S. M., Lindgren, E., Yang, J. N., Herling, A. W., Fredholm, B. B. Adenosine A1 receptors regulate lipolysis and lipogenesis in mouse adipose tissue-interactions with insulin. European Journal of Pharmacology. 597 (1-3), 92-101 (2008).
- Gnad, T., et al. Adenosine activates brown adipose tissue and recruits beige adipocytes via A2A receptors. Nature. 516 (7531), 395-399 (2014).
- Fried, S. K., et al. Resistance to the antilipolytic effect of insulin in adipocytes of African-American compared to Caucasian postmenopausal women. Journal of Lipid Research. 51 (5), 1193-1200 (2010).
- Lee, M. J., Fried, S. K. Optimal protocol for the differentiation and metabolic analysis of human adipose stromal cells. Methods in Enzymology. 538, 49-65 (2014).
- Fricke, K., Heitland, A., Maronde, E. Cooperative activation of lipolysis by protein kinase A and protein kinase C pathways in 3T3-L1 adipocytes. Endocrinology. 145 (11), 4940-4947 (2004).
- Bergan, H. E., Kittilson, J. D., Sheridan, M. A. PKC and ERK mediate GH-stimulated lipolysis. Journal of Molecular Endocrinology. 51 (2), 213-224 (2013).
- Schmitz-Peiffer, C. The tail wagging the dog--regulation of lipid metabolism by protein kinase C. The FEBS Journal. 280 (21), 5371-5383 (2013).
- Carmen, G. Y., Victor, S. M. Signalling mechanisms regulating lipolysis. Cellular Signalling. 18 (4), 401-408 (2006).
- Zu, L., et al. Bacterial endotoxin stimulates adipose lipolysis via toll-like receptor 4 and extracellular signal-regulated kinase pathway. The Journal of Biological Chemistry. 284 (9), 5915-5926 (2009).
- Zhang, H. H., Halbleib, M., Ahmad, F., Manganiello, V. C., Greenberg, A. S. Tumor necrosis factor-alpha stimulates lipolysis in differentiated human adipocytes through activation of extracellular signal-related kinase and elevation of intracellular cAMP. Diabetes. 51 (10), 2929-2935 (2002).
- Tan, X., et al. TNF-α downregulates CIDEC via MEK/ERK pathway in human adipocytes. Obesity. 24 (5), 1070-1080 (2016).
- Laurencikiene, J., et al. NF-kappaB is important for TNF-alpha-induced lipolysis in human adipocytes. Journal of Lipid Research. 48 (5), 1069-1077 (2007).
- van Hall, G., et al. Interleukin-6 stimulates lipolysis and fat oxidation in humans. The Journal of Clinical Endocrinology and Metabolism. 88 (7), 3005-3010 (2003).
- Wueest, S., et al. Mesenteric fat lipolysis mediates obesity-associated hepatic steatosis and insulin resistance. Diabetes. 65 (1), 140-148 (2016).
- Trujillo, M. E., et al. Interleukin-6 regulates human adipose tissue lipid metabolism and leptin production in vitro. The Journal of Clinical Endocrinology and Metabolism. 89 (11), 5577-5582 (2004).
- Kitamura, T., et al. Insulin-induced phosphorylation and activation of cyclic nucleotide phosphodiesterase 3B by the serine-threonine kinase Akt. Molecular and Cellular Biology. 19 (9), 6286-6296 (1999).
- Chakrabarti, P., et al. Insulin inhibits lipolysis in adipocytes via the evolutionarily conserved mTORC1-Egr1-ATGL-mediated pathway. Molecular and Cellular Biology. 33 (18), 3659-3666 (2013).
- Collins, S., Daniel, K. W., Petro, A. E., Surwit, R. S. Strain-specific response to beta 3-adrenergic receptor agonist treatment of diet-induced obesity in mice. Endocrinology. 138 (1), 405-413 (1997).
- Surwit, R. S., Dixon, T. M., Petro, A. E., Daniel, K. W., Collins, S. Diazoxide restores beta3-adrenergic receptor function in diet-induced obesity and diabetes. Endocrinology. 141 (10), 3630-3637 (2000).
- Gettys, T. W., et al. Age-dependent changes in beta-adrenergic receptor subtypes and adenylyl cyclase activation in adipocytes from Fischer 344 rats. Endocrinology. 136 (5), 2022-2032 (1995).
- Mowers, J., et al. Inflammation produces catecholamine resistance in obesity via activation of PDE3B by the protein kinases IKKε and TBK1. eLife. 2, e01119 (2013).
- Valentine, J. M., et al. β3-Adrenergic receptor downregulation leads to adipocyte catecholamine resistance in obesity. The Journal of Clinical Investigation. 132 (2), e153357 (2022).
- Collins, S., et al. Impaired expression and functional activity of the beta 3- and beta 1-adrenergic receptors in adipose tissue of congenitally obese (C57BL/6J ob/ob) mice. Molecular Endocrinology. 8 (4), 518-527 (1994).
- Collins, S., Surwit, R. S. The beta-adrenergic receptors and the control of adipose tissue metabolism and thermogenesis. Recent Progress in Hormone Research. 56, 309-328 (2001).
- Dixon, T. M., Daniel, K. W., Farmer, S. R., Collins, S. CCAAT/enhancer-binding protein alpha is required for transcription of the beta 3-adrenergic receptor gene during adipogenesis. The Journal of Biological Chemistry. 276 (1), 722-728 (2001).
- Lohse, M. J., Benovic, J. L., Codina, J., Caron, M. G., Lefkowitz, R. J. beta-Arrestin: a protein that regulates beta-adrenergic receptor function. Science. 248 (4962), 1547-1550 (1990).
- Nantel, F., et al. The human beta 3-adrenergic receptor is resistant to short term agonist-promoted desensitization. Molecular Pharmacology. 43 (4), 548-555 (1993).
- Liggett, S. B., Freedman, N. J., Schwinn, D. A., Lefkowitz, R. J. Structural basis for receptor subtype-specific regulation revealed by a chimeric beta 3/beta 2-adrenergic receptor. Proceedings of the National Academy of Sciences. 90 (8), 3665-3669 (1993).
- Baker, J. G. The selectivity of beta-adrenoceptor agonists at human beta1-, beta2- and beta3-adrenoceptors. British Journal of Pharmacology. 160 (5), 1048-1061 (2010).
- Lafontan, M. Inhibition of epinephrine-induced lipolysis in isolated white adipocytes of aging rabbits by increased alpha-adrenergic responsiveness. Journal of Lipid Research. 20 (2), 208-216 (1979).
- Baker, J. G. The selectivity of beta-adrenoceptor antagonists at the human beta1, beta2 and beta3 adrenoceptors. British Journal of Pharmacology. 144 (3), 317-322 (2005).
- Jensen, M. D., Nielsen, S. Insulin dose response analysis of free fatty acid kinetics. Metabolism. 56 (1), 68-76 (2007).
- Jensen, M. D., Haymond, M. W., Gerich, J. E., Cryer, P. E., Miles, J. M. Lipolysis during fasting. Decreased suppression by insulin and increased stimulation by epinephrine. The Journal of Clinical Investigation. 79 (1), 207-213 (1987).
- Heckmann, B. L., et al. Defective adipose lipolysis and altered global energy metabolism in mice with adipose overexpression of the lipolytic inhibitor G0/G1 switch gene 2 (G0S2). The Journal of Biological Chemistry. 289 (4), 1905-1916 (2014).
- Shin, H., et al. Lipolysis in brown adipocytes is not essential for cold-induced thermogenesis in mice. Cell Metabolism. 26 (5), 764.e5-777.e5 (2017).
- Treble, D. H., Mayer, J. Glycerolkinase activity in white adipose tissue of obese-hyperglycaemic mice. Nature. 200, 363-364 (1963).
- Possik, E., et al. New mammalian glycerol-3-phosphate phosphatase: role in beta-cell, liver and adipocyte metabolism. Frontiers in Endocrinology. 12, 706607 (2021).
- Romero Mdel, M., Sabater, D., Fernandez-Lopez, J. A., Remesar, X., Alemany, M. Glycerol production from glucose and fructose by 3T3-L1 cells: a mechanism of adipocyte defense from excess substrate. PLoS One. 10 (10), e0139502 (2015).
- Mugabo, Y., et al. Identification of a mammalian glycerol-3-phosphate phosphatase: Role in metabolism and signaling in pancreatic beta-cells and hepatocytes. Proceedings of the National Academy of Sciences. 113 (4), E430-E439 (2016).
- Hanson, R. W., Reshef, L. Glyceroneogenesis revisited. Biochimie. 85 (12), 1199-1205 (2003).
- Vaughan, M. The production and release of glycerol by adipose tissue incubated in vitro. The Journal of Biological Chemistry. 237, 3354-3358 (1962).
- Jensen, M. D., Ekberg, K., Landau, B. R. Lipid metabolism during fasting. American Journal of Physiology-Endocrinology and Metabolism. 281 (4), E789-E793 (2001).
- Ballard, F. J., Hanson, R. W., Leveille, G. A. Phosphoenolpyruvate carboxykinase and the synthesis of glyceride-glycerol from pyruvate in adipose tissue. The Journal of Biological Chemistry. 242 (11), 2746-2750 (1967).
- Reshef, L., Hanson, R. W., Ballard, F. J. A possible physiological role for glyceroneogenesis in rat adipose tissue. The Journal of Biological Chemistry. 245 (22), 5979-5984 (1970).
- Gorin, E., Tal-Or, Z., Shafrir, E. Glyceroneogenesis in adipose tissue of fasted, diabetic and triamcinolone treated rats. European Journal of Biochemistry. 8 (3), 370-375 (1969).
- Elia, M., Zed, C., Neale, G., Livesey, G. The energy cost of triglyceride-fatty acid recycling in nonobese subjects after an overnight fast and four days of starvation. Metabolism. 36 (3), 251-255 (1987).
- Reshef, L., et al. Glyceroneogenesis and the triglyceride/fatty acid cycle. Journal of Biological Chemistry. 278 (33), 30413-30416 (2003).
- Edens, N. K., Leibel, R. L., Hirsch, J. Mechanism of free fatty acid re-esterification in human adipocytes in vitro. Journal of Lipid Research. 31 (8), 1423-1431 (1990).
- Vaughan, M., Steinberg, D. Effect of hormones on lipolysis and esterification of free fatty acids during incubation of adipose tissue in vitro. Journal of Lipid Research. 4, 193-199 (1963).
- Brooks, B., Arch, J. R., Newsholme, E. A. Effects of hormones on the rate of the triacylglycerol/fatty acid substrate cycle in adipocytes and epididymal fat pads. Federation of European Biochemical Societies Letters. 146 (2), 327-330 (1982).
- Bjorntorp, P., Karlsson, M., Hovden, A. Quantitative aspects of lipolysis and reesterification in human adipose tissue in vitro. Acta Medica Scandinavica. 185 (1-2), 89-97 (1969).
- Angel, A., Desai, K., Halperin, M. L. Free fatty acid and ATP levels in adipocytes during lipolysis. Metabolism. 20 (1), 87-99 (1971).
- Husted, A. S., et al. Autocrine negative feedback regulation of lipolysis through sensing of NEFAs by FFAR4/GPR120 in WAT. Molecular Metabolism. 42, 101103 (2020).
- Fain, J. N., Shepherd, R. E. Free fatty acids as feedback regulators of adenylate cyclase and cyclic 3':5'-AMP accumulation in rat fat cells. The Journal of Biological Chemistry. 250 (16), 6586-6592 (1975).
- Burns, T. W., Langley, P. E., Terry, B. E., Robinson, G. A. The role of free fatty acids in the regulation of lipolysis by human adipose tissue cells. Metabolism. 27 (12), 1755-1762 (1978).
- Kalderon, B., et al. Suppression of adipose lipolysis by long-chain fatty acid analogs. Journal of Lipid Research. 53 (5), 868-878 (2012).
- Schweiger, M., et al. Measurement of lipolysis. Methods in Enzymology. 538, 171-193 (2014).
- Decaunes, P., Bouloumie, A., Ryden, M., Galitzky, J. Ex vivo analysis of lipolysis in human subcutaneous adipose tissue explants. Bio-Protocol. 8 (3), e2711 (2018).
- Roy, D., Myers, J. M., Tedeschi, A. Protocol for assessing ex vivo lipolysis of murine adipose tissue. STAR Protocols. 3 (3), 101518 (2022).
- Baskaran, P., Thyagarajan, B. Measurement of basal and forskolin-stimulated lipolysis in inguinal adipose fat pads. Journal of Visualized Experiments. 125 (125), 55625 (2017).
- Reilly, S. M., et al. Catecholamines suppress fatty acid re-esterification and increase oxidation in white adipocytes via STAT3. Nature Metabolism. 2 (7), 620-634 (2020).
- Liu, L., et al. Isolation of mouse stromal vascular cells for monolayer culture. Methods in Molecular Biology. 1566, 9-16 (2017).
- DeLuca, J. H., Reilly, S. M. . Methods in Molecular Biology. , (2023).
- Richard, G., Vernon, R. A. C. New Perspectives in Adipose Tissue. Butterworth-Heinemann. , (1985).
- Brito, M. N., Botion, L. M., Brito, N. A., Kettelhut, I. C., Migliorini, R. H. Lipolysis and glycerokinase activity in brown adipose tissue of rat fed a high protein, carbohydrate-free diet. Hormone and Metabolic Research. 26 (1), 51-52 (1994).
- Bertin, R. Glycerokinase activity and lipolysis regulation in brown adipose tissue of cold acclimated rats. Biochimie. 58 (4), 431-434 (1976).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved