
Method Article
Évaluation de la réponse auditive du tronc cérébral chez les nouveau-nés de poulet
Dans cet article
Résumé
Nous avons utilisé des techniques standard de réponse auditive du tronc cérébral (ABR) et les avons appliquées à des poulets nouveau-nés, un modèle aviaire précoce pour la fonction auditive. Le protocole décrit en détail les techniques de préparation animale et d’acquisition ABR, avec des étapes qui pourraient se traduire par d’autres modèles aviaires ou rongeurs.
Résumé
La réponse auditive du tronc cérébral (ABR) est un test inestimable en audiologie clinique, chez les animaux non humains et dans la recherche humaine. Malgré l’utilisation généralisée des APR dans la mesure de la synchronisation neuronale auditive et l’estimation de la sensibilité auditive dans d’autres systèmes de modèles de vertébrés, les méthodes d’enregistrement des APR chez le poulet n’ont pas été rapportées depuis près de quatre décennies. Les poulets fournissent un modèle de recherche animal robuste parce que leur système auditif est proche de la maturation fonctionnelle à la fin de l’embryon et au début de l’éclosion. Nous avons démontré des méthodes utilisées pour obtenir des enregistrements ABR à un ou deux canaux à l’aide de réseaux d’électrodes à aiguilles sous-dermiques chez les nouveau-nés de poulet. Quelle que soit la configuration d’enregistrement de l’électrode (c.-à-d. montage), les enregistrements ABR comprenaient 3 à 4 formes d’onde de crête positives dans les 6 premières ms d’un stimulus de clic supra-seuil. Les amplitudes de forme d’onde de crête à creux variaient de 2 à 11 μV à des niveaux de haute intensité, les pics positifs présentant des fonctions d’intensité de latence attendues (c.-à-d. augmentation de la latence en fonction de la diminution de l’intensité). La position normalisée des écouteurs était essentielle pour des enregistrements optimaux, car la peau lâche peut obstruer le conduit auditif et le mouvement des animaux peut déloger le transducteur de stimulus. Les amplitudes de crête étaient plus faibles et les latences étaient plus longues à mesure que la température corporelle des animaux diminuait, ce qui favorisait la nécessité de maintenir la température corporelle physiologique. Pour les jeunes nouveau-nés (<3 h après le jour 1 de l’éclosion), les seuils ont été relevés d’environ 5 dB, les latences maximales ont augmenté d’environ 1 à 2 ms et les amplitudes des pics au creux ont diminué d’environ 1 μV par rapport aux nouveau-nés plus âgés. Cela suggère un problème potentiel lié à la conductivité (c.-à-d. du liquide dans la cavité de l’oreille moyenne) et devrait être envisagé pour les jeunes nouveau-nés. Dans l’ensemble, les méthodes ABR décrites ici permettent un enregistrement précis et reproductible de la fonction auditive in vivo chez les nouveau-nés de poulet qui pourrait être appliqué à différents stades de développement. De tels résultats sont facilement comparés à des modèles humains et mammifères de perte auditive, de vieillissement ou d’autres manipulations auditives.
Introduction
L’étude des réponses neuronales évoquées aux stimuli sonores remonte à plus d’un demi-siècle1. La réponse auditive du tronc cérébral (ABR) est un potentiel évoqué qui a été utilisé comme mesure de la fonction auditive chez les animaux non humains et les humains depuis des décennies. L’ABR humain présente cinq à sept pics de forme d’onde classiquement étiquetés par des chiffres romains (I-VII)2. Ces pics sont analysés en fonction de leur latence (temps d’occurrence en millisecondes) et de l’amplitude (taille du pic au creux en microvolts) des réponses neuronales. L’ABR joue un rôle déterminant dans l’évaluation de la fonction et de l’intégrité du nerf auditif ainsi que de la sensibilité du tronc cérébral et du seuil auditif. Les déficits du système auditif entraînent des latences et des amplitudes ABR absentes, réduites, prolongées ou anormales. Remarquablement, ces paramètres sont presque identiques chez les humains et les autres animaux, ce qui en fait un test objectif cohérent de la fonction auditive à travers les modèles de vertébrés3.
L’un de ces systèmes modèles est le poulet, et il est particulièrement utile pour diverses raisons. Les oiseaux peuvent être classés comme altriciels ou précoces4. Les oiseaux altriciels éclosent avec des sens encore en développement; par exemple, les chouettes effraies ne présentent pas un ABR constant avant quatre jours après l’éclosion5. Les animaux précoces comme le poulet éclosent avec des sens presque matures. Le début de l’audition se produit dans le développement embryonnaire, de sorte que quelques jours avant l’éclosion (jour embryonnaire 21), le système auditif est proche de la maturation fonctionnelle 6,7,8. Les oiseaux altriciels et la plupart des modèles de mammifères sont sensibles aux facteurs extrinsèques qui influencent le développement et nécessitent un élevage jusqu’à ce que l’audition soit mature. Les APR de poulet peuvent être effectués le même jour que l’éclosion, en renonçant au besoin d’alimentation ou d’un environnement enrichi.
Le poulet embryonnaire a été un modèle bien étudié pour la physiologie et le développement, en particulier dans le tronc cérébral auditif. Les structures spécifiques comprennent le noyau cochléaire du poulet, divisé en noyau magnocellularis (NM) et noyau angularis (NA), et le corrélat aviaire de l’olive supérieure médiale connue sous le nom de noyau laminaire (NL)6,7. L’ABR est idéal pour se concentrer sur la fonction auditive centrale avant le niveau du cerveau antérieur et du cortex. La traduction entre les mesures ABR in vivo et les études neuronales in vitro du développement8, de la physiologie9, de la tonotopie10 et de la génétique11,12 offre des possibilités de recherche idéales qui soutiennent les études de la fonction auditive globale.
Bien que l’ABR ait été largement étudié dans les modèles de mammifères, il y a eu moins d’attention pour les oiseaux. Les études antérieures de l’ABR aviaire comprennent des caractérisations de la perruche13, du pic14, de la mouette15, des oiseaux plongeurs16, du pinson zèbre17, des rapaces diurnes18, des canaris19, de trois espèces dehiboux 5,20,21,22 et de poulet23. Compte tenu des près de quatre décennies qui se sont écoulées depuis la dernière caractérisation approfondie du poulet ABR, de nombreux équipements et techniques précédemment utilisés ont changé. Les enseignements tirés d’études menées dans d’autres modèles aviaires peuvent aider à développer une méthodologie ABR moderne du poulet tout en servant de comparaison avec l’ABR du poulet. Cet article décrira la configuration expérimentale et la conception permettant l’enregistrement ABR chez les poulets nouveau-nés qui pourraient également être appliqués aux stades embryonnaires de développement et à d’autres modèles de petits rongeurs et d’oiseaux. De plus, étant donné le développement précoce du poulet, des manipulations du développement peuvent être effectuées sans élevage extensif. Les manipulations d’un embryon en développement peuvent être évaluées quelques heures seulement après l’éclosion de l’animal avec des capacités auditives presque matures.
Protocole
Les expériences décrites ici ont été approuvées par les comités institutionnels de soins et d’utilisation des animaux (IACUC) de l’Université Northwestern et menées conformément au National Institutes of Health Guide for the Care and Use of Laboratory Animals.
1. Élevage de poulet
- Acquérir des œufs de poule de livourne blanc fécondés.
REMARQUE: Il existe plusieurs races de poulet utilisées dans la recherche scientifique, mais les résultats présentés ici proviennent de poulets à livournes blanches (Gallus gallus domesticus). Bien que la variabilité de l’ABR entre les races soit inconnue, certaines différences ont été trouvées en comparant les poulets pondeurs adultes aux poulets de chair producteurs de viande24,25. - Incuber les œufs à 38 °C, humidité à 50 %, pendant 21 jours avant la date d’essai souhaitée.
REMARQUE: Si les œufs ne sont pas immédiatement incubés à 38 ° C, ils peuvent être conservés à 14 ° C, humidité à 40%. Cependant, plus les œufs sont maintenus longtemps à 14 °C, moins ils sont susceptibles de devenir des nouveau-nés viables. Après 7 jours, la viabilité des œufs peut chuter jusqu’à 50 % selon la durée de conservation des œufs à 14 °C. La viabilité des œufs diminuera également pendant les mois d’hiver. - Tournez périodiquement les œufs 2 à 3 fois par jour. La plupart des incubateurs ont un mécanisme pour effectuer cela automatiquement.
- Si vous utilisez un incubateur en polystyrène ou un incubateur contenant plus de 6 œufs, transférer les œufs dans un petit incubateur à 38 °C la veille de l’éclosion, jour embryonnaire 20 (E20). Les œufs doivent éclore 21 jours (E21) après avoir été mis dans l’incubateur.
REMARQUE: Dans le processus d’éclosion, l’animal commencera à « piquer » hors de l’œuf, faisant un petit trou qui finit par contourner tout l’œuf. Si les conditions sont trop sèches, l’œuf peut se dessécher et l’animal sera incapable d’éclore. L’humidité doit être maintenue autour de 50%, sur la base d’études antérieures sur la viabilité de l’éclosion des œufs 26,27,28,29. - Déterminez l’âge de l’animal. Si l’éclosion n’est pas observée en personne, la seule indication de l’âge est les 2-3 heures qu’il faut pour que le liquide amniotique sèche.
REMARQUE: L’incubateur à nouveau-né doit être soigneusement nettoyé quotidiennement avec de l’alcool isopropylique à 70% en fonction du nombre de nouveau-nés traités. Les nouveau-nés de poulet laissent souvent des excréments, des plumes et du liquide amniotique dans l’incubateur, ce qui peut contaminer les conditions et la qualité de l’air.
2. Préparation de médicaments
- Pesez l’animal en le plaçant dans un grand bateau de pesage. Avec un placement assez doux, l’animal ne doit pas bouger.
REMARQUE: La masse peut varier de 30 à 45 g. Les animaux plus jeunes sont souvent plus lourds en raison des réserves de jaunes et du fait qu’ils n’excrètent pas encore de déchets. Les animaux plus âgés approchant l’âge de 24 heures et P2 pèsent généralement moins. - Préparer un cocktail anesthésique de kétamine (100 mg/mL) et de xylazine (20 mg/mL) de telle sorte que la posologie soit de 50 mg/kg de kétamine et de 16,68 mg/kg de xylazine en fonction du poids de l’animal.
REMARQUE: Ce cocktail médicamenteux peut être fait avec 1 mL de kétamine (100 mg / mL), 1,5 mL de xylazine (20 mg / mL) et 2,5 mL de H2O. Les injections de cocktail anesthésique varieront de 0,05 à 0,1 mL en fonction de la plage de 30 à 45 g de poids animal.
3. Injection de drogues et préparation des animaux
- Tenez l’animal dans une main, en veillant à maintenir les pattes vers le bas.
- Sentez le sternum de l’animal, la quille. De chaque côté de la quille se trouvera le muscle mammaire.
- Utilisez une aiguille et une seringue de 29 G pour pénétrer 5 mm dans la peau et injectez le cocktail kétamine/xylazine dans le muscle mammaire. Injecter entre 0,05 et 0,1 mL en fonction du poids de l’animal.
- Replacez l’animal dans l’incubateur après l’injection. Maintenez la température corporelle de l’animal pendant quelques minutes pendant que l’anesthésique prend effet.
- Utilisez une pince pour pincer l’orteil de l’animal et vérifiez si le cou est mou. S’il n’y a pas de réflexe et un cou boiteux, l’animal est inconscient.
- Déterminez le sexe du poulet à l’aide de ses plumes d’ailes. Si les plumes ont toutes la même longueur, l’animal est mâle. Si la longueur des plumes varie, l’animal est femelle30.
REMARQUE: Une autre méthode de sexage de l’animal est la ventilation. Les organes génitaux masculins peuvent être vus dans lecloaque 31. Cette méthode est très difficile et peut nuire à l’animal si elle n’est pas faite correctement. Il est recommandé d’utiliser la méthode de la plume d’aile. - Appliquez la crème dépilatoire avec un applicateur à pointe de coton sur la tête et le cou, en particulier près de l’ouverture de l’oreille pour l’oiseau.
- Utilisez des lingettes à 70% d’alcool isopropylique pour essuyer les plumes, toute crème dépilatoire restante et la peau de la tête et du cou.
- Utilisez une lingette à 70% d’alcool isopropylique pour stériliser les électrodes sous-cutanées et la sonde rectale.
- Placez l’animal dans une chambre insonorisée et blindée électriquement. Assurez-vous que l’environnement a un minimum de bruit électrique et acoustique pour les meilleurs enregistrements.
REMARQUE: Les expériences ici ont été faites dans un boîtier isolé du son personnalisé mesurant 24 x 24 x 25 pouces. Toute chambre ou pièce qui élimine le bruit acoustique, ainsi que le bruit électrique du courant électrique alternatif (60 Hz aux États-Unis), est suffisante. - Utilisez un coussin chauffant ou un système de contrôle de la température pour maintenir la température corporelle de l’animal.
- Insérez la sonde rectale lubrifiée pour vous assurer que la température de l’animal est maintenue entre 37-41 °C (98,6-105 °F)32,33.
REMARQUE: Si la sonde est d’une taille incorrecte, l’animal peut se coucher sur le dessus de la sonde de température. - Fixez la tête de l’animal en place ou posez le bec contre un objet pour éviter tout mouvement indésirable. Cela peut être fait avec de la pâte à modeler si la respiration n’est pas obstruée.
- Administrer une injection supplémentaire de cocktail anesthésique qui représente la moitié de la dose initiale si l’animal commence à reprendre conscience pendant les tests.
REMARQUE: Tout mouvement du corps ou vocalisation est un signe qu’un supplément doit être administré. De minuscules mouvements du bec indiquent la respiration et sont acceptables.
4. Placement des électrodes
- Utilisez trois électrodes à aiguille en acier inoxydable et en chlorure d’argent avec les désignations suivantes : l’électrode de référence, l’électrode active et l’électrode de masse commune.
REMARQUE: L’électrode de référence est également appelée inversion ou « -« . L’électrode active est également appelée non inversante ou « + ». - Placez chaque électrode sous-dermiquement de 2-3 mm dans la tête, mais pas assez profondément pour pénétrer dans le crâne. Utilisez des électrodes de 7 mm de longueur et de 0,4 mm de diamètre.
- Sortez l’électrode de la peau, exposant la pointe. Cela permet de minimiser le contact avec la peau et d’assurer une profondeur d’insertion constante chez les animaux34.
REMARQUE: Le fil de l’électrode doit avoir un mou suffisant pour qu’après avoir placé l’électrode, il n’y ait pas de tension qui le tirera ou tirera la peau tendue. - Pour l’enregistrement à canal unique, placez l’électrode active au-dessus du crâne à la ligne médiane, aussi loin caudale que le conduit auditif.
- Placez l’électrode de référence derrière l’oreille où le stimulus sera délivré et placez l’électrode de terre derrière le conduit auditif controlatéral dans le cou.
REMARQUE: Si vous effectuez une intervention chirurgicale au niveau du crâne ou du conduit auditif de l’animal, placez l’électrode de référence dans le cou à la ligne médiane de l’animal. Cette étape et l’étape 4.4.1 sont considérées comme des montages d’enregistrement d’électrodes horizontales.
- Placez l’électrode de référence derrière l’oreille où le stimulus sera délivré et placez l’électrode de terre derrière le conduit auditif controlatéral dans le cou.
- Pour l’enregistrement à deux canaux, utilisez deux électrodes négatives et une électrode positive combinée qui nécessite un câble adaptateur. Placez l’électrode de masse sous le cordon dans le cou et une électrode de référence derrière chaque conduit auditif.
- Vérifiez l’impédance de l’électrode. Assurez-vous que l’impédance globale de l’électrode ne dépasse pas 5,0 kΩ. Maintenir l’impédance interélectrode en dessous de 3,0 kΩ.
5. Enregistrement ABR
- En fonction du matériel et des logiciels d’acquisition, assurez-vous d’effectuer l’étalonnage pour obtenir des niveaux sonores corrects sur les fréquences de stimulus utilisées.
REMARQUE : Les techniques d’étalonnage varient en fonction de l’équipement (voir la discussion). Pour certains programmes, l’atténuation du son peut être modifiée dans le logiciel. Les procédures d’étalonnage effectuées ici impliquaient l’utilisation d’un microphone à condensateur B&K 4138 de 1/8 pouce pour enregistrer les stimuli de fréquence dans un système de coupleur fermé qui se rapprochait du conduit auditif poussin (~ 5 mm). Une table d’étalonnage de l’éclosion de poulet est fournie en tant que table supplémentaire. - Déplacez l’appareil du transducteur sonore vers l’oreille active de l’animal. Placez le transducteur sonore à une faible profondeur de 2 mm dans le conduit auditif.
REMARQUE: Selon le transducteur de son, un spéculum en plastique peut être attaché et inséré dans le conduit auditif. Le placement du spéculum est critique. Si le son est bloqué par la paroi du canal ou si le conduit auditif est pincé, les APR seront absents ou ressembleront à un décalage d’environ 40 dB dans le seuil. - Vérifiez sur l’animal pendant les tests si les résultats semblent anormaux ou absents. Si c’est le cas, repositionnez le transducteur sonore dans le conduit auditif.
REMARQUE: Étant donné que la peau est lâche et que le mouvement des animaux est possible, le placement du spéculum peut changer pendant l’enregistrement. Cependant, avec une injection anesthésique correcte et l’animal complètement inconscient, l’enregistrement peut rester ininterrompu pendant 30 à 45 minutes.
6. Acquisition de données
- Utilisez suffisamment d’équipement ou de logiciel pour générer des stimuli sonores et enregistrer/acquérir des enregistrements ABR.
REMARQUE: Il existe de nombreux systèmes disponibles dans le commerce ou personnalisés pour l’acquisition d’ABR. Pour ces expériences, la plate-forme USB SmartEP intelligent des systèmes auditifs intelligents (IHS) disponible dans le commerce a été utilisée. La capacité de manipuler les paramètres d’enregistrement est essentielle; ceux-ci incluent, sans toutefois s’y limiter, l’intensité du stimulus, la durée du stimulus, la fréquence du stimulus, le taux de présentation du stimulus, le filtre passe-haut et passe-bas, le rejet d’artefact, le nombre de balayages, le taux d’échantillonnage, la forme de l’enveloppe et la polarisation du stimulus. - Réglez les limites supérieure et inférieure du rejet d’artefact (RA) sur ±25 μV, de sorte que le mouvement ou le bruit des animaux pendant un balayage exclura ce balayage de l’analyse. Dans l’ensemble de la population testée, moins de 1 % du total des balayages ont été rejetés en raison d’artefacts.
- Recueillez au moins 1024 balayages pour obtenir une réponse moyenne. Cela peut être fait en deux enregistrements de 512 balayages chacun. Cela garantit également que la réponse est évoquée par un stimulus et reproductible.
- Réglez le gain sur 100 000, le filtre passe-bas sur 100 Hz et le filtre passe-haut sur 3000 Hz.
REMARQUE: Les paramètres de filtre passe-bas et passe-haut étaient optimaux pour les enregistrements utilisant le système IHS. Par conséquent, ces paramètres sont des recommandations. Les enregistrements ABR chez d’autres espèces aviaires à l’aide du logiciel BIOSIG ont filtré le signal entre 30 et 3000 Hz 5,13,14,16,22. - Définissez le taux de présentation du stimulus entre 10 et 20 stimuli par seconde. Des taux de présentation élevés modifieront la latence de crête ABR, en particulier pour les picsultérieurs 13. De faibles taux de présentation augmenteront le temps nécessaire à l’acquisition de l’ABR.
- Réglez la durée du stimulus de clic sur 100 μs.
- Si vous utilisez un stimulus d’éclatement de tonalité, modifiez la fréquence et la durée du stimulus en fonction de l’effet souhaité. Une plage de 100-4000 Hz a été utilisée pour les stimuli de tonalité, bien que la gamme d’audition comportementale chez les poulets adultes varie de 2-9000 Hz35.
REMARQUE: Dans le système IHS, le temps de montée et de descente d’un stimulus de sursaut de tonalité ne peut être modifié que si la forme de l’enveloppe spectrale est un trapèze. Cependant, les enveloppes cosinus carrées et Blackman fournissent un temps de montée et de chute prédéfini qui est couramment utilisé dans les expériences ABR animales. Le système IHS peut afficher l’enveloppe spectrale d’un sursaut de tonalité pour assurer des temps de montée et de descente appropriés. Le temps de montée et de chute d’un stimulus de clic ne peut pas être modifié dans IHS.
- Si vous utilisez un stimulus d’éclatement de tonalité, modifiez la fréquence et la durée du stimulus en fonction de l’effet souhaité. Une plage de 100-4000 Hz a été utilisée pour les stimuli de tonalité, bien que la gamme d’audition comportementale chez les poulets adultes varie de 2-9000 Hz35.
- Réglez la fréquence d’échantillonnage sur la valeur la plus élevée autorisée (généralement 40 kHz) pour obtenir les meilleures données de résolution.
REMARQUE: Certains systèmes, y compris IHS, utilisent un nombre limité de points d’échantillonnage et modifient la durée de la fenêtre d’enregistrement. Une fréquence d’échantillonnage de 40 kHz (période de 25 μs) ne peut permettre qu’une fenêtre d’enregistrement de 12 ms, donc pour capturer un ABR de sursaut de tonalité, une fréquence d’échantillonnage de 20 kHz (période de 50 μs) a été utilisée pour permettre une fenêtre d’enregistrement de 24 ms. Si vous comparez directement les ARN de clic et de tonalité, maintenez le taux d’échantillonnage constant pour maintenir la même résolution. - Réglez la polarisation du stimulus sur l’alternance. Ceci est fait pour éliminer la visualisation de la microphonique cochléaire des enregistrements ABR. Pour visualiser la microphonie cochléaire, utilisez la raréfaction ou la condensation pour la polarité du stimulus.
REMARQUE: De nombreux paramètres peuvent être modifiés lors de la sélection des stimuli. Les paramètres de gain et de filtre fournis peuvent ne pas être optimaux pour d’autres configurations d’équipement. Les paramètres d’usine par défaut sur la plupart des machines ABR ne sont pas configurés pour l’enregistrement dans le poulet nouveau-né. - Si vous enregistrez 512 balayages, combinez deux tests distincts pour créer une moyenne de balayage de 1024.
- Pour un stimulus de clic ou d’éclatement de tonalité, acquérez un ABR à une intensité supra-seuil.
- Continuez à enregistrer à des intensités de plus en plus faibles jusqu’à ce que la réponse évoquée ne puisse plus être identifiée.
- Définissez le seuil ABR comme l’intensité de stimulus la plus faible qui suscite une réponse évoquée détectable. Abaissez l’intensité du stimulus par paliers de 5 dBSPL pour trouver l’intensité de stimulus la plus faible qui déclenche un pic détectable.
7. Euthanasie et fin de l’expérience
- Une fois les ABR acquis, préparer un surdosage (0,1 mL) de solution d’euthanasie (Pentobarbital Sodium 390 mg/mL Phénytoïne Sodique 50 mg/mL).
- Après avoir utilisé un pincement d’orteil pour confirmer qu’il n’y a pas de réflexe, injecter la solution d’euthanasie dans le muscle mammaire avec une aiguille 29-G à une profondeur de 5 mm. La technique d’injection est la même que l’injection anesthésique.
REMARQUE: L’animal expirera après quelques minutes. Ne manipulez pas ou ne décapitez pas l’animal jusqu’à ce qu’aucun mouvement ne soit détecté. Une autre technique d’euthanasie consiste à effectuer une injection intraveineuse dans la veine brachiale sous l’aile. - Dès que l’animal n’est pas réflexif et que la respiration et le rythme cardiaque ont cessé, décapitez rapidement avec des ciseaux ou des cisailles tranchants.
- Nettoyez le coussin chauffant, la sonde rectale et les électrodes de chlorure d’argent avec des lingettes à 70% d’alcool isopropylique.
- Assurez-vous que toutes les traces acquises ont été enregistrées. Pour une analyse plus approfondie, exportez les fichiers sous forme de fichiers .txt qui peuvent être affichés dans le bloc-notes ou importés dans une feuille de calcul.
Résultats
Enregistrements ABR représentatifs pour les poussins nouveau-nés
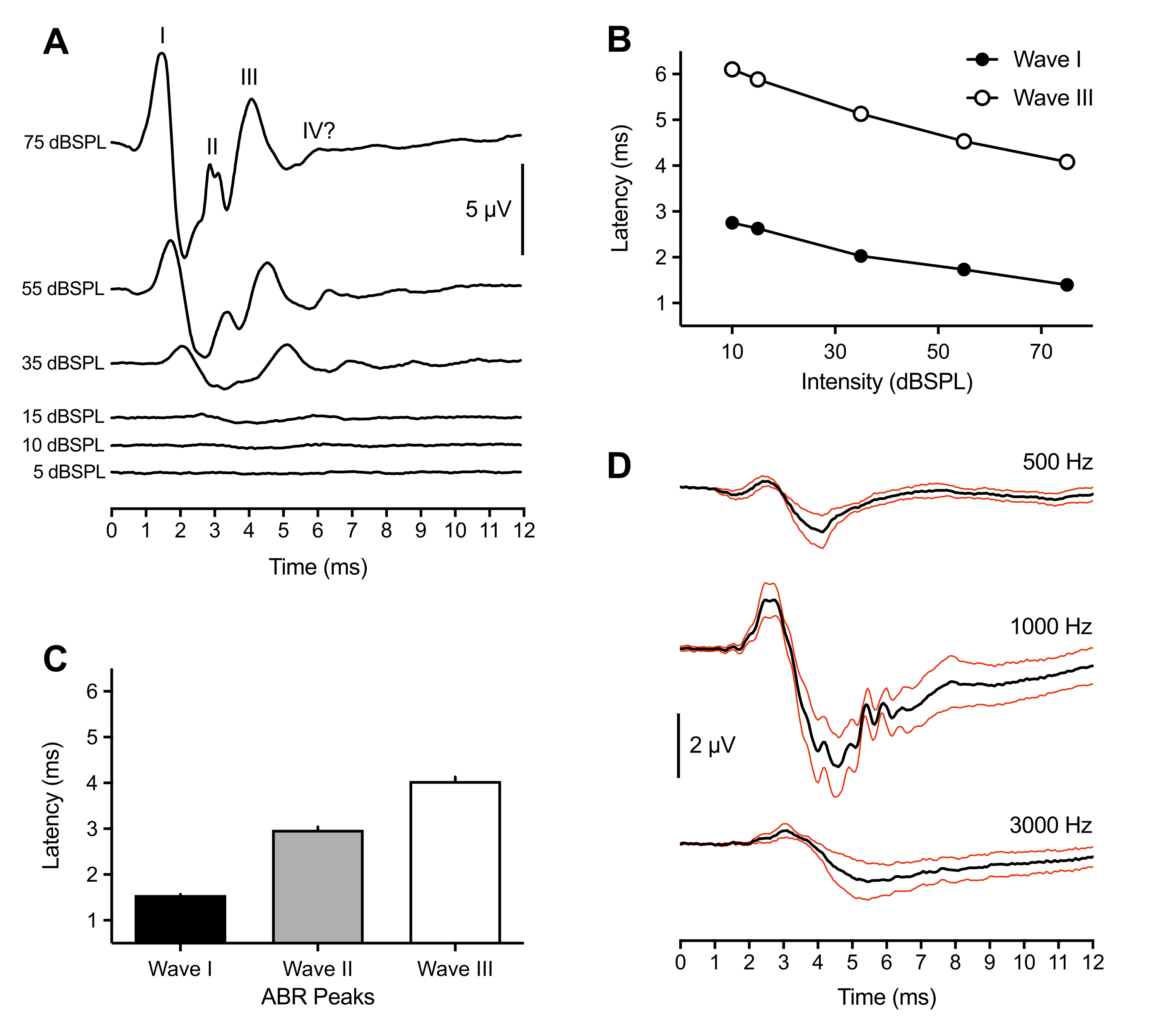
Les résultats représentatifs et de population suivants proviennent d’enregistrements ABR effectués chez 43 animaux. En réponse à un stimulus de clic supra-seuil (75 dBSPL), trois pics positifs ont été systématiquement observés chez tous les nouveau-nés. Ces pics se sont produits dans les 6 ms suivant l’apparition du stimulus. Rarement, un quatrième pic a également été observé à ~6 ms. Alors que l’identification des pics ABR chez les oiseaux variait d’un animal à l’autre (voir la discussion), les pics ont été étiquetés et identifiés comme des vagues I à IV en chiffres romains. Une forme d’onde ABR représentative avec des pics marqués est illustrée à la figure 1A (trace supérieure). La figure 1B montre la fonction latence-intensité pour les ondes I et III étiquetées dans la trace représentative. La latence de crête de l’onde I a augmenté d’environ 0,3 ms pour chaque diminution de 20 dB de l’intensité du stimulus. En moyenne, les vagues I-III se sont produites à 1,50 ms (±0,02 ms), 3,00 ms (±0,06 ms) et 4,13 ms (±0,09 ms) à 75 dBSPL, respectivement (Figure 1C). Les vagues I et III se présentent toujours comme un pic singulier. Occasionnellement, pour la vague II, plusieurs petits pics ont été observés entre 2,5 et 3,2 ms. Chaque pic avait un creux correspondant, et l’amplitude de crête à creux de l’onde I - le plus grand de tous les pics - était en moyenne de 7 μV et approchait une amplitude maximale de 11 μV à 75 dB SPL.
En plus de la plus grande amplitude, l’onde I du poussin ABR présentait la moindre variabilité de la latence de crête chez les animaux. Par conséquent, ce pic a été utilisé pour estimer la sensibilité du seuil auditif. Les seuils ABR ont été définis comme l’intensité de stimulus la plus faible qui a provoqué un pic de forme d’onde identifiable et reproductible. Cela a été déterminé subjectivement par l’expérimentateur et recoupé par un deuxième expérimentateur pour l’accord de seuil. Les pics étaient mieux définis et plus faciles à identifier lors de l’utilisation de stimuli de clic, mais les sursauts de tonalité généraient également des pics définis et identifiables qui variaient en fonction de la fréquence du stimulus et de ses paramètres (Figure 1D, n = 4 poussins). Le seuil ABR évoqué par clic était inférieur au seuil évoqué par sursaut de tonalité, à l’exception de 1000 Hz. Les seuils variaient entre 10 et 30 dBSPL pour les stimuli de clic. Les APR évoqués par clic qui ne montraient pas de pics identifiables >30 dBSPL étaient souvent le résultat du délogement du spéculum du conduit auditif en raison du mouvement des animaux.
La diminution de la température corporelle augmente les latences ABR
La vitesse de l’activité neuronale - mesurée par l’occurrence maximale d’une amplitude de forme d’onde (c.-à-d. latence) - est connue pour diminuer à des températures corporelles plus basses36,37. Ce phénomène a été observé chez les APR de poulet nouveau-né en utilisant un stimulus de clic de 75 dBSPL. Une trace représentative est illustrée à la figure 2A. Lorsque la température corporelle a diminué à partir de 39 ° C, la latence des pics ABR s’est produite plus tard dans le temps, malgré le même niveau d’intensité du stimulus. La figure 2B montre la latence des ondes I et III en fonction de températures corporelles plus basses pour la trace représentative. Il y avait une forte corrélation (R2 = 0,89) entre la baisse de la température corporelle et l’apparition de la latence maximale de la vague I (figure 2C, n = 5 poussins). Ces résultats démontrent la nécessité de maintenir une température corporelle proche de la normale pendant les enregistrements ABR. Si la température corporelle proche de la normale n’est pas maintenue, les fonctions d’intensité de latence et les mesures d’amplitude de l’ABR sont très variables et souvent inexactes.
Différences de latence et d’amplitude chez les premiers nouveau-nés
La recherche a montré que l’activité neuronale liée au début de l’audition chez le poussin est proche de la maturation à la fin de l’âge embryonnaire8. Cependant, pour un sous-ensemble de nouveau-nés très précoces (<3 h après l’éclosion), nous avons observé un décalage de latence maximale des formes d’onde ABR (n = 4) en réponse à un stimulus de clic SPL de 75 dB ou les potentiels évoqués n’étaient pas identifiables (n = 2 poussins). Chez 2 jeunes nouveau-nés, aucun ABR de tonalité n’a pu être déclenché et les seuils de clic ont été élevés de 50 dBSPL. Cela pourrait être dû à un problème de conduction où il y a encore du liquide dans le conduit auditif / cavité de l’oreille moyenne de l’animal, ou à un composant neuronal sous-développé. Des études sur les mammifères ont rapporté des changements de seuil de 50 dB chez les nouveau-nés38,39. Les animaux représentatifs utilisés ici étaient âgés de >3 h, ce qui coïncidait également avec le temps nécessaire au séchage des plumes. La figure 3A montre les APR enregistrés chez les jeunes (P1, <3 h) et les nouveau-nés plus âgés (P2). Pour l’analyse, seuls 3 jeunes nouveau-nés ont présenté les trois pics ABR. Les latences de forme d’onde de crête ont été significativement prolongées et les amplitudes de forme d’onde ont été légèrement réduites par rapport aux nouveau-nés plus âgés (figure 3B-C, respectivement).
Placement de l’électrode de référence et enregistrements ABR à deux canaux
Dans la figure 4, le placement de l’électrode de référence a été modifié entre 2 emplacements différents, mais a tout de même donné lieu à des enregistrements ABR comparables. Une comparaison entre 75 traces de clic dBSPL chez le même animal avec les deux placements d’électrodes de référence a montré des différences minimes dans les amplitudes de forme d’onde de crête à creux et les latences de forme d’onde de crête (Figure 4A). Le placement des mastoïdes était méthodologiquement similaire aux expériences ABR sur les mammifères qui placent l’électrode de référence sur la mastoïde ou le pavillon. L’utilisation d’un placement du cou pour l’électrode de référence serait bénéfique si une manipulation ou une intervention chirurgicale était effectuée sur l’une ou l’autre oreille. Fait intéressant, l’amplitude du pic de la vague II pour le placement mastoïdien (trace rouge) s’est produite 1 ms après le pic de la vague II pour le placement du cou (trace noire). Ce décalage horaire reflète probablement le(s) site(s) de génération neuronale ABR par rapport au placement de l’électrode.
À l’aide d’une configuration à deux canaux, une électrode d’enregistrement active (placement du haut de la tête) et deux électrodes de référence (placements mastoïdiens) ont été utilisées pour obtenir des APR pour les oreilles gauche et droite (figure 4B). Les réponses entre les deux oreilles étaient similaires, avec des changements mineurs dans les amplitudes de crête probablement dus au positionnement des écouteurs. La latence de l’oreille gauche et de l’oreille droite étant équivalente, elle a soutenu le fonctionnement tout aussi sain des deux oreilles et des hémisphères du tronc cérébral chez le poulet nouveau-né. Le montage d’enregistrement à deux canaux pourrait également être utilisé pour les ARN binauraux, mais des considérations supplémentaires seraient nécessaires pour ces enregistrements.

Figure 1 : Enregistrements représentatifs de poussins nouveau-nés à des stimuli évoqués par des clics et des tonalités. (A) Enregistrements ABR représentatifs d’un poussin nouveau-né (P2) en fonction de différents niveaux d’intensité de stimulus. Trois à quatre pics positifs en microvolts (μV) peuvent être identifiés dans les 6 ms suivant le début du stimulus (temps = 0 ms). Les ondes ont été identifiées à l’aide de chiffres romains. Les amplitudes de crête à creux diminuent à des niveaux d’intensité de stimulus plus faibles. (B) Fonctions latence-intensité des ondes I et III pour la trace représentative indiquée au point (A). Seuls ces pics ont été analysés, car la vague II n’a généralement pas été observée à des intensités <45 dBSPL. (C) Latence des formes d’onde de crête ABR évoquées par clic (n = 43 poussins). Les barres d’erreur désignent l’erreur-type de la moyenne (SEM). (D) Moyenne des ABR (traces noires) évoquées par tonalité pour quatre poussins nouveau-nés à trois fréquences différentes. Traces rouges = erreur type des stimuli moyens (SEM) = 75 dBSPL. Dans cette figure et les suivantes, les barres d’erreur désignent le SEM, et l’oreille droite était l’oreille de stimulation. (exception pour la figure 4B où les deux oreilles ont été stimulées). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2: Effet de la température corporelle sur les enregistrements ABR. (A) Enregistrements ABR représentatifs d’un poussin nouveau-né (P2) en fonction de la température corporelle. Pour les températures corporelles plus basses, les latences de forme d’onde maximale ont augmenté tandis que les amplitudes de crête à creux sont restées relativement inchangées. (B) Fonction latence-température des ondes I et III pour les traces représentatives indiquées en (A). (C) Données démographiques montrant la relation entre la latence et les changements de température pour 5 poussins (p < 0,01, R2 = 0,89). Une tendance similaire a été observée pour les vagues II et III (données non présentées). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Différences liées à l’âge sur les enregistrements ABR. (A) Enregistrements ABR représentatifs (chevauchement) d’un poussin nouveau-né représentatif à P2 (trace noire) et P1 (<3 h après l’éclosion, trace rouge). (B) Latences de forme d’onde maximale pour les ondes I, II et III en fonction de l’âge. Les latences des vagues I à III étaient significativement différentes d’un âge à l’autre (P < 0,05, n = 6 poussins). (C) Amplitudes de forme d’onde de crête à creux des ondes I, II et III en fonction de l’âge. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Placement de l’électrode et enregistrements ABR à deux canaux : (A) Enregistrements ABR représentatifs (superposés) du même poussin nouveau-né (P2) avec l’électrode de référence placée dans le cou (trace noire) ou mastoïde (trace rouge). L’électrode active a été placée à la ligne médiane du crâne pour les deux montages d’enregistrement d’électrodes. La latence des ondes I et III, et l’amplitude des ondes I et III sont presque identiques dans les deux conditions. La latence de l’onde II est plus précoce et l’amplitude est plus grande pour l’électrode placée dans le tissu du cou. (B) Enregistrement à deux canaux tout en stimulant séquentiellement les oreilles droite et gauche. Enregistrements ABR représentatifs (superposés) du même poussin nouveau-né (P2) avec les électrodes de référence placées dans la mastoïde de l’oreille gauche (traces bleues) et de l’oreille droite (traces rouges) à trois niveaux d’intensité différents. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Tableau supplémentaire : Table d’étalonnage de l’éclosion de poulet. Veuillez cliquer ici pour télécharger ce tableau.
Discussion
Le tronc cérébral auditif des oiseaux est bien étudié et de nombreuses structures sont analogues à la voie auditive des mammifères. Le nerf auditif fournit des entrées excitatrices sur les deux noyaux centraux de premier ordre, le noyau cochléaire magnocellularis (NM) et angularis (NA). NM envoie une projection excitatrice bilatéralement à sa cible auditive, le noyau laminaire (NL)7. NL projette le noyau mesencephalicus lateralis, pars dorsalis (MLd)40,41. NL projette également sur le noyau olivaire supérieur (SON), qui fournit une inhibition de rétroaction à NM, NA et NL42. Ce microcircuit auditif inférieur du tronc cérébral est conservé de manière exquise pour la fonction qu’il sous-sert, la localisation du son et l’audition binaurale33. Les régions du tronc cérébral auditif supérieur de l’oiseau ont également des noyaux analogues au lemniscus latéral des mammifères et au colliculus inférieur dans le mésencéphale. Compte tenu de ces similitudes, la composition de l’ABR aviaire jusqu’au mésencéphale auditif est comparable chez tous les vertébrés.
Alors que plusieurs espèces aviaires présentent trois pics positifs dans les 6 ms suivant l’apparition du stimulus, la corrélation des pics ABR avec les structures auditives centrales présente une certaine variabilité. On peut raisonnablement supposer que l’onde I est la première réponse neuronale de la papille basilaire périphérique et du nerf auditif et présente peu de variabilité entre les individus (Figure 1C). L’identification ultérieure des vagues est moins certaine et peut différer d’une espèce à l’autre. Kuokkanen et coll.17 ont récemment déterminé que la vague III de l’ABR de la chouette effraie est générée par NL; il est donc raisonnable de soutenir que l’onde II provient de NM et NA du noyau cochléaire20. Cependant, la vague III de la chouette a été définie comme le pic positif généré 3 ms après le début du stimulus. Cela correspond à la vague II telle que définie dans l’ABR du poulet nouveau-né. Dans l’ABR de la chouette effraie, les vagues I et II ont été combinées.
Alors que le poulet nouveau-né présentait habituellement trois pics dans les 6 ms, un quatrième pic a été observé à l’occasion (p. ex., voir la figure 1A). Des données sur la population, une plus grande taille d’échantillon et des paradigmes expérimentaux supplémentaires seraient nécessaires pour soutenir une quatrième vague et, dans certains cas, un ABR de poulet à cinq vagues. La constatation la plus cohérente a été les trois représentations de pic présentées ici.
Étant donné que l’ABR est défini comme une mesure de la synchronie neuronale, les principaux noyaux de la voie auditive pourraient représenter chaque pic positif de l’ABR. Le signal passant du nerf auditif à NM/NA puis à NL peut définir les ondes I, II et III dans l’ABR de poulet nouveau-né, respectivement. De plus, le quatrième pic tardif de l’ABR du poulet pourrait représenter un tronc cérébral supérieur ou une structure auditive du mésencéphale. La caractérisation des ABR aviaires devrait également tenir compte de la différence entre les oiseaux précoces et les oiseaux altriciaux. La maturation des réponses auditives varie d’une espèce à l’autre et est également affectée par d’autres traits critiques tels que le comportement des prédateurs et / ou l’apprentissage vocal4. Quoi qu’il en soit, les méthodes et techniques décrites sont facilement appliquées à une variété d’espèces aviaires et vertébrées.
L’importance du maintien de la température corporelle des animaux est illustrée à la figure 2. À mesure que la température interne du corps diminuait, la latence des réponses ABR augmentait pour le même niveau d’intensité du stimulus. Ceci est plus prononcé lorsque la température corporelle tombe en dessous de 32 °C36,37. L’augmentation de la latence d’environ 1 ms dans l’ABR est inférieure à celle précédemment rapportée dans le poulet23. Cependant, Katayama23 a utilisé un nouveau-né de 12 jours qui a été refroidi puis réchauffé sur une période de 4 heures. Les données de la figure 2 ont été enregistrées pendant le processus de refroidissement sur une période de 20 minutes. Pour obtenir la meilleure qualité et les enregistrements les plus cohérents, la température corporelle de l’animal doit être maintenue et tous les enregistrements doivent être effectués à la même température physiologique chez les animaux.
L’effet de l’âge sur l’ABR est léger mais important à considérer. Bien que seule la latence des ondes I et II de l’ABR soit significativement différente, c’est en partie parce que seuls trois jeunes nouveau-nés ont été utilisés dans la figure 3; les trois autres ne présentaient pas trois pics ABR identifiables. Les changements d’amplitude et de seuil de l’ABR peuvent également être évidents si vous utilisez de grandes tailles d’échantillon ou si vous comparez des APR spécifiques à la fréquence. Cet effet lié à l’âge pourrait être causé par du liquide dans l’oreille moyenne du poulet. De tels changements de conduction entraînent une augmentation marquée des seuils ABR pour les modèles humains et d’autres mammifères38,39.
À l’aide de deux montages d’enregistrement différents, des réponses similaires ont été observées (figure 4A). Alors que le montage le plus courant place l’électrode de référence derrière l’oreille réceptrice du stimulus, avoir l’électrode de référence dans le tissu du cou peut être utile s’il y a une intervention chirurgicale accompagnant l’ABR. Toutefois, si des enregistrements ABR à deux canaux sont utilisés, les électrodes de référence doivent être placées séparément et symétriquement, ce qui est difficile si vous placez l’électrode de référence dans le cou. La position mastoïdienne de l’électrode de référence est recommandée pour normaliser autant d’aspects de l’enregistrement que possible. L’enregistrement ABR à deux canaux est un outil efficace nécessitant peu de préparation supplémentaire et entraîne des réponses similaires entre les oreilles. Des différences d’amplitude mineures étaient probablement dues au positionnement de l’écouteur. L’enregistrement à deux canaux permet une comparaison facile entre une oreille ou un hémisphère cérébral manipulé expérimentalement et un contrôle. Cette configuration serait également nécessaire pour tester les ABR binauraux. Les expériences futures utilisant l’ABR de poulet peuvent se référer à la littérature antérieure sur les configurations d’enregistrement et les montages34.
Cette méthodologie comporte plusieurs limites. Comme mentionné à l’étape 5.1, un mauvais placement du spéculum peut entraîner un changement de 40 dBSPL en réponse. Cela pourrait entraîner une interprétation incorrecte d’un animal manipulé ou modifié. Les précautions suivantes sont recommandées : acquérir un large échantillon de données de contrôle avant d’acquérir les APR de modèles manipulés ou mutants. Ne diminuez pas l’intensité du stimulus de plus de 20 dBSPL entre les enregistrements. Si l’amplitude ou la latence se déplace plus que prévu, vérifiez la position de l’animal et du spéculum. Répétez ce stimulus ABR pour observer les changements. Si le spéculum a bougé, réacquisez les tests précédents. Une autre limitation est l’étalonnage des APR. Sans étalonnage approprié pour enregistrer le niveau de pression acoustique, l’intensité présentée à l’animal est inconnue. Lorsque vous mesurez la sortie sonore, utilisez le même spéculum que dans l’enregistrement expérimental et un petit microphone à l’intérieur d’une cavité qui se rapproche de la longueur du conduit auditif de l’animal (~ 5 mm). Mesurez les mêmes fréquences de tonalité utilisées dans les expériences, car les étalonnages sont spécifiques à la fréquence. Le manuel pour les systèmes matériels et logiciels peut être accompagné d’instructions pour l’étalonnage. Il existe également des filtres supplémentaires tels que les filtres de phase linéaire et de phase minimale, qui peuvent améliorer les ARN de clic et de tonalité43. Ces filtres n’ont pas été utilisés dans la présente étude. Des considérations supplémentaires, telles que le temps de montée et de descente d’une enveloppe spectrale de sursaut de tonalité changeant en fonction de la fréquence ou modifiant le temps de montée et de descente des stimuli de clic n’ont pas non plus été examinées. Il s’agit de bonnes enquêtes futures une fois que des APR fiables et cohérents pourront être acquis.
La comparaison du poulet nouveau-né avec d’autres modèles aviaires est prometteuse. Les perruches et les hiboux de l’Est affichent également trois pics positifs de microvolts dans les 6 premiers ms de l’ABR13,22. Chez différentes espèces de pics, trois pics sont également observés, mais leur latence est plus tardive. De plus, la plage de la meilleure sensibilité à la fréquence chez les pics se situe entre 1500 et 4000 Hz, ce qui est un peu plus élevé que le meilleur seuil du poulet à 1000 Hz. Chez le poulet adulte, la meilleure sensibilité est à 2000 Hz35, de sorte qu’il peut y avoir une meilleure audition des hautes fréquences à mesure que les nouveau-nés de poulet se développent en adultes. Ce développement différera d’une espèce d’oiseau à l’autre, en tenant compte du développement altricial ou précoce de l’animal4.
Les méthodes expérimentales décrites ici peuvent aider à déterminer quels facteurs entraînent des préjudices ou des changements dans les réponses auditives et les seuils, ainsi que des études à différents stades du développement embryonnaire. La manipulation génétique, le vieillissement et l’exposition au bruit sont tous des manipulations connues chez les animaux et autres modèles aviaires 24,25,44,45. Ces méthodes devraient être étendues au modèle du poulet maintenant que des techniques comme l’électroporation in-ovo permettent l’expression de protéines qui sont contrôlées focalement et temporellement d’un côté du tronc cérébral auditif12,46. Cela permet la comparaison directe des APR de l’oreille génétiquement manipulée à l’oreille de contrôle controlal controlaire en utilisant un paradigme d’enregistrement à deux canaux.
Dans l’ensemble, l’ABR des poulets nouveau-nés est une méthode de recherche utile, presque identique aux mesures de la fonction auditive chez les humains et d’autres modèles de mammifères. Il s’agit également d’une méthodologie in vivo non invasive. En dehors de l’injection anesthésique et du placement d’électrodes sous-cutanées de quelques millimètres, aucune autre manipulation physique n’est nécessaire. Un nouveau-né pourrait théoriquement être testé plusieurs fois au cours d’une période de développement de plusieurs jours ou semaines s’il est conservé dans un environnement approprié. Ce protocole définit non seulement les étapes et les paramètres d’enregistrement nécessaires pour l’ABR du poulet nouveau-né, mais il propose également les caractéristiques d’un ABR aviaire qui peuvent éclairer d’autres tests sur la fonction auditive du tronc cérébral.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Ce travail est soutenu par le NIH/NIDCD R01 DC017167
matériels
| Name | Company | Catalog Number | Comments |
| 1/8 inch B&K Microphone | Brüel & Kjær | 4138 | Type 4138-A-015 also works |
| Auditory Evoked Potential Universal Smart Box | Intelligent Hearing Systems | M011110 | |
| Custom Sound Isolation Chamber | GK Soundbooth Inc | N/A | Custom built |
| DC Power Supply | CSI/Speco | PSV-5 | |
| ER3 Insert Earphone | Intelligent Hearing Systems | M015302 | Used as sound transducer |
| Euthasol | Virbac | 710101 | Controlled Substance; euthanasia solution |
| Insulin Syringe (29 G) | Comfort Point | 26028 | |
| Ketamine | Covetrus | 11695-0703-1 | Controlled Substance |
| Power Supply | Powervar | 93051-55R | |
| Rectal Probe | YSI | 401 (10-09010) | Any 400 series probe will work with the YSI temperatuer monitor |
| Subdermal needles | Rhythmlink | RLSND107-1.5 | |
| Temperature Monitor | YSI | 73ATA 7651 | Works with any 400 series rectal probe |
| Xylazine | Anased | 59399-110-20 | Used with ketamine and water for anesthetic |
Références
- Wever, E. G., Bray, C. W. Action currents in the auditory nerve in response to acoustical stimulation. Proceedings of the National Academy of Sciences of the United States of America. 16 (5), 344-350 (1930).
- Jewett, D. L., Williston, J. S. Auditory-evoked far fields averaged from the scalp of humans. Brain. 94 (4), 681-696 (1971).
- Corwin, J. T., Bullock, T. H., Schweitzer, J. The auditory brain stem response in five vertebrate classes. Electroencephalography and Clinical Neurophysiology. 54 (6), 629-641 (1982).
- Carey, C. Avian Growth and Development. Evolution within the Altricial Precocial Spectrum. , Oxford University Press. Oxford. (1998).
- Kraemer, A., Baxter, C., Hendrix, A., Carr, C. E. Development of auditory sensitivity in the barn owl. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural and Behavioral Physiology. 203 (10), 843-853 (2017).
- Rebillard, G., Rubel, E. W. Electrophysiological study of the maturation of auditory responses from the inner ear of the chick. Brain Research. 229 (1), 15-23 (1981).
- Parks, T. N., Rubel, E. W. Organization and development of brain stem auditory nuclei of the chicken: organization of projections from n. magnocellularis to n. laminaris. Journal of Comparative Neurology. 164 (4), 435-448 (1975).
- Hong, H., Rollman, L., Feinstein, B., Sanchez, J. T. Developmental profile of ion channel specializations in the avian nucleus magnocellularis. Frontiers in Cellular Neuroscience. 10, 80(2016).
- Leao, R. M. The ion channels and synapses responsible for the physiological diversity of mammalian lower brainstem auditory neurons. Hearing Research. 376, 33-46 (2019).
- Oline, S. N., Ashida, G., Burger, R. M. Tonotopic optimization for temporal processing in the cochlear nucleus. Journal of Neuroscience. 36 (32), 8500-8515 (2016).
- Kopp-Scheinpflug, C. Your genes decide what you are listening to. Channels. 11 (5), 355-356 (2017).
- Sid, H., Schusser, B. Applications of gene editing in chickens: A new era is on the horizon. Frontiers in Genetics. 9, 456(2018).
- Brittan-Powell, E. F., Dooling, R. J., Gleich, O. Auditory brainstem responses in adult budgerigars (Melopsittacus undulatus). The Journal of the Acoustical Society of America. 112 (3), Pt 1 999-1008 (2002).
- Lohr, B., Brittan-Powell, E. F., Dooling, R. J. Auditory brainstem responses and auditory thresholds in woodpeckers. The Journal of the Acoustical Society of America. 133 (1), 337-342 (2013).
- Counter, S. A. Brain-stem evoked potentials and noise effects in seagulls. Comparative Biochemistry and Physiology. A, Comparative Physiology. 81 (4), 837-845 (1985).
- Crowell, S. E., et al. A comparison of auditory brainstem responses across diving bird species. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural and Behavioral Physiology. 201 (8), 803-815 (2015).
- Noirot, I. C., Brittan-Powell, E. F., Dooling, R. J. Masked auditory thresholds in three species of birds, as measured by the auditory brainstem response (L). Journal of the Acoustical Society of America. 129 (6), 3445-3448 (2011).
- McGee, J., et al. Auditory performance in bald eagles and red-tailed hawks: a comparative study of hearing in diurnal raptors. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural and Behavioral Physiology. 205 (6), 793-811 (2019).
- Brittan-Powell, E. F., Dooling, R. J., Ryals, B., Gleich, O. Electrophysiological and morphological development of the inner ear in Belgian Waterslager canaries. Hearing Research. 269 (1-2), 56-69 (2010).
- Kuokkanen, P. T., Kraemer, A., Kempter, R., Koppl, C., Carr, C. E. Auditory brainstem response wave iii is correlated with extracellular field potentials from nucleus laminaris of the barn owl. Acta Acustica United with Acustica. The Journal of the European Acoustics Association (EEIG). 104 (5), 874-877 (2018).
- Beatini, J. R., Proudfoot, G. A., Gall, M. D. Frequency sensitivity in Northern saw-whet owls (Aegolius acadicus). Journal of Comparative Physiology. A, Neuroethology Sensory Neural and Behavioral Physiology. 204 (2), 145-154 (2018).
- Brittan-Powell, E. F., Lohr, B., Hahn, D. C., Dooling, R. J. Auditory brainstem responses in the Eastern Screech Owl: an estimate of auditory thresholds. The Journal of the Acoustical Society of America. 118 (1), 314-321 (2005).
- Katayama, A. Postnatal development of auditory function in the chicken revealed by auditory brainstem responses (ABRs). Electroencephalography and Clinical Neurophysiology. 62 (5), 388-398 (1985).
- Durham, D., Park, D. L., Girod, D. A. Breed differences in cochlear integrity in adult, commercially raised chickens. Hearing Research. 166 (1-2), 82-95 (2002).
- Kaiser, C. L., Girod, D. A., Durham, D. Breed-dependent susceptibility to acute sound exposure in young chickens. Hearing Research. 203 (1-2), 101-111 (2005).
- Hamdy, A. M., Vander Hel, W., Henken, A. M., Galal, A. G., Abd-Elmoty, A. K. Effects of air humidity during incubation and age after hatch on heat tolerance of neonatal male and female chicks. Poultry Science. 70 (7), 1499-1506 (1991).
- Bruzual, J. J., Peak, S. D., Brake, J., Peebles, E. D. Effects of relative humidity during the last five days of incubation and brooding temperature on performance of broiler chicks from young broiler breeders. Poultry Science. 79 (10), 1385-1391 (2000).
- vander Pol, C. W., van Roovert-Reijrink, I. A. M., Maatjens, C. M., vanden Brand, H., Molenaar, R. Effect of relative humidity during incubation at a set eggshell temperature and brooding temperature posthatch on embryonic mortality and chick quality. Poultry Science. 92 (8), 2145-2155 (2013).
- Buhr, R. J. Incubation relative humidity effects on allantoic fluid volume and hatchability. Poultry Science. 74 (5), 874-884 (1995).
- Galli, R., et al. Sexing of chicken eggs by fluorescence and Raman spectroscopy through the shell membrane. PLoS One. 13 (2), 0192554(2018).
- Otsuka, M., Miyashita, O., Shibata, M., Sato, F., Naito, M. A novel method for sexing day-old chicks using endoscope system. Poultry Science. 95 (11), 2685-2689 (2016).
- Kaiser, A. The ontogeny of homeothermic regulation in post-hatching chicks: its influence on the development of hearing. Comparative Biochemistry and Physiology. A, Comparative Physiology. 103 (1), 105-111 (1992).
- Kuba, H., Yamada, R., Ohmori, H. Evaluation of the limiting acuity of coincidence detection in nucleus laminaris of the chicken. The Journal of Physiology. 552, Pt 2 611-620 (2003).
- Shaheen, L. A., Valero, M. D., Liberman, M. C. Towards a diagnosis of cochlear neuropathy with envelope following responses. Journal of the Association for Research in Otolaryngology. 16 (6), 727-745 (2015).
- Hill, E. M., Koay, G., Heffner, R. S., Heffner, H. E. Audiogram of the chicken (Gallus gallus domesticus) from 2 Hz to 9 kHz. Journal of Comparative Physiology. A, Neuroethology Sensory Neural and Behavioral Physiology. 200 (10), 863-870 (2014).
- Rossi, G. T., Britt, R. H. Effects of hypothermia on the cat brainstem auditory evoked response. Electroencephalography and Clinical Neurophysiology. 57 (2), 143-155 (1984).
- Doyle, W. J., Fria, T. J. The effects of hypothermia on the latencies of the auditory brainstem response (ABR) in the rhesus monkey. Electroencephalography and Clinical Neurophysiology. 60 (3), 258-266 (1985).
- Guan, X., Gan, R. Z. Effect of middle ear fluid on sound transmission and auditory brainstem response in guinea pigs. Hearing Research. 277 (1-2), 96-106 (2011).
- Ravicz, M. E., Rosowski, J. J., Merchant, S. N. Mechanisms of hearing loss resulting from middle-ear fluid. Hearing Research. 195 (1-2), 103-130 (2004).
- Wang, Y., Zorio, D. A. R., Karten, H. J. Heterogeneous organization and connectivity of the chicken auditory thalamus (Gallus gallus). Journal of Comparative Neurology. 525 (14), 3044-3071 (2017).
- Wang, Y., Karten, H. J. Three subdivisions of the auditory midbrain in chicks (Gallus gallus) identified by their afferent and commissural projections. Journal of Comparative Neurology. 518 (8), 1199-1219 (2010).
- Fukui, I., Burger, R. M., Ohmori, H., Rubel, E. W. GABAergic inhibition sharpens the frequency tuning and enhances phase locking in chicken nucleus magnocellularis neurons. Journal of Neuroscience. 30 (36), 12075-12083 (2010).
- Beutelmann, R., Laumen, G., Tollin, D., Klump, G. M. Amplitude and phase equalization of stimuli for click evoked auditory brainstem responses. The Journal of the Acoustical Society of America. 137 (1), 71-77 (2015).
- Liberman, M. C. Noise-induced and age-related hearing loss: new perspectives and potential therapies. F1000Research. 6, 927(2017).
- Efrati, A., Gutfreund, Y. Early life exposure to noise alters the representation of auditory localization cues in the auditory space map of the barn owl. Journal of Neurophysiology. 105 (5), 2522-2535 (2011).
- Lu, T., Cohen, A. L., Sanchez, J. T. In ovo electroporation in the chicken auditory brainstem. Journal of Visualized Experiments: JoVE. (124), e55628(2017).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.