
Method Article
Meccanobiologia completamente ottica Interrogazione di proteine associate al sì nel cancro umano e nelle cellule normali utilizzando un sistema multifunzionale
* Questi autori hanno contribuito in egual misura
In questo articolo
Riepilogo
Questo documento presenta un protocollo graduale dettagliato su come utilizzare un sistema integrato multifunzionale e programmabile dall'utente che consente l'imaging multicanale automatico e l'analisi meccanobiologica per chiarire la meccanosensibilità della proteina associata al Sì (YAP).
Abstract
L'imaging multifunzionale a lungo termine e l'analisi delle cellule vive richiedono un coordinamento funzionale semplificato di varie piattaforme hardware e software. Tuttavia, il controllo manuale di varie apparecchiature prodotte da diversi produttori richiede molto lavoro e molto tempo, riducendo potenzialmente l'accuratezza, la riproducibilità e la qualità dei dati acquisiti. Pertanto, un sistema all-in-one e programmabile dall'utente che consente l'acquisizione automatica, multifunzionale e a lungo termine ed è compatibile con la maggior parte delle piattaforme di microscopia fluorescente può avvantaggiare la comunità scientifica. Questo documento introduce i protocolli operativi completi dell'utilizzo di un nuovo sistema software integrato che consiste in (1) un programma software costruito in casa, intitolato "Automatic Multi-functional Integration Program (AMFIP)", che consente l'acquisizione automatica di immagini multicanale e (2) una suite di analisi di imaging quantitativo e pacchetti di calcolo della trazione cellulare.
Questo sistema integrato viene applicato per rivelare la relazione precedentemente sconosciuta tra la distribuzione spazio-temporale della proteina associata al sì meccano-sensibile (YAP) e la meccanica cellulare, compresa la diffusione e la trazione cellulare, nelle cellule normali umane (B2B) ingegnerizzate CRISPR / Cas9 e nelle cellule tumorali polmonari (PC9). Sfruttando la capacità di questo sistema di controllo e lettura multicanale, il risultato mostra: (1) le cellule normali B2B e le cellule tumorali PC9 mostrano una relazione distinta tra espressione YAP, trazione e dinamica cellulare durante i processi di diffusione e migrazione cellulare; e (2) le cellule tumorali PC9 applicano evidenti forze perinucleari sui substrati. In sintesi, questo documento presenta un protocollo graduale dettagliato su come utilizzare un sistema integrato programmabile dall'utente che consente l'imaging e l'analisi multifunzionali automatici per chiarire la meccanosensibilità YAP. Questi strumenti aprono la possibilità di esplorazioni dettagliate di dinamiche di segnalazione sfaccettate nel contesto della fisiologia e della patologia cellulare.
Introduzione
L'obiettivo generale di questo metodo è quello di consentire l'imaging multifunzionale completamente ottico e l'analisi delle cellule viventi. Un programma di imaging all-in-one che consente il coordinamento automatico di dispositivi optoelettronici multifunzionali ridurrà le operazioni manuali ad alta intensità di lavoro e soggette a errori ed è essenziale per i ricercatori per condurre l'imaging a cellule vive a lungo termine1,2,3,4. Tuttavia, la maggior parte dei programmi pubblici esistenti nella comunità di ricerca biomedica si applicano solo a dispositivi optoelettronici limitati o richiedono hardware aggiuntivo per il coordinamento di apparecchiature diverse5,6,7,8,9. Recentemente, è stato sviluppato un programma open source e basato su software, intitolato "Automatic Multi-functional Integration Program (AMFIP)", che consente l'imaging multicanale e time-lapse. Basato sul linguaggio Java e sull'Application Programming Interface (API) di μManager11,12, AMFIP è stato sviluppato come plugin in μManager che esegue script Java personalizzati per realizzare comunicazioni basate su software di più piattaforme hardware e software optoelettroniche, incluse ma non limitate a quelle di Nikon. L'istituzione di AMFIP apre la possibilità di interrogazioni programmabili e multifunzionali dei comportamenti cellulari. In questo articolo viene sviluppato un sistema sperimentale e computazionale integrato che combina AMFIP con l'analisi di imaging digitale e la microscopia della forza di trazione cellulare. Il sistema consente di chiarire la distinta meccanobiologia YAP nelle linee cellulari B2B normali umane (Figura 1) e PC9 (Figura 2) progettate da CRISPR/Cas9. Il sistema fornisce alla comunità scientifica una soluzione completa che evita la richiesta di acquistare dispositivi di controllo aggiuntivi che potrebbero non essere disponibili e/o compatibili con ogni sistema di imaging.
I protocolli presentati in questo documento introducono come (1) applicare AMFIP per condurre l'imaging automatico a lungo termine per entrambe le linee cellulari ingegnerizzate CRISPR / Cas9 che esprimono YAP con tag mNEonGreen2; e (2) combinare Fiji ImageJ, MATLAB e Origin per l'analisi quantitativa del rapporto YAP nucleare/citoplasma (N/C) in base alla loro intensità fluorescente (Figura 3 e Figura 4), campo di spostamento cellulare (Figura 1C e Figura 2C) e campo di trazione cellulare (Figura 1D e Figura 2D) ). I risultati suggeriscono che (1) durante le prime 10 ore di diffusione cellulare sui substrati che hanno rigidità meccanica fisiologicamente rilevante13,14,15,16,17,18, il rapporto YAP N/C delle singole cellule B2B mostra variazioni e fluttuazioni dipendenti dal tempo più evidenti rispetto a quelle delle singole cellule PC9 (Figura 5 e Figura 6 ); e (2) le cellule tumorali PC9 generano una notevole trazione nelle loro regioni perinucleari (Figura 7). Il sistema integrato e le metodologie descritte in questo protocollo trascendono i tipi specifici di cellule e molecole optogenetiche. I ricercatori possono applicare i protocolli per personalizzare i loro specifici esperimenti di interrogazione di cellule vive e chiarire le dinamiche di segnalazione sfaccettate nel contesto della fisiologia e della patologia cellulare.
Protocollo
1. Generazione di una linea cellulare stabile di carcinoma polmonare umano (PC9) e di una linea cellulare epiteliale bronchiale umana (Beas2B) stabile che esprimono endogenamente la proteina YAP mNeonGreen21-10/11-tagged
- Eseguire la reazione a catena della polimerasi (PCR) per amplificare la sequenza di DNA che codifica l'11° filamento della proteina di fluorescenza, mNeonGreen2, utilizzando la DNA polimerasi ad alta fedeltà (vedere la Tabella dei materiali).
- Inserire la sequenza di DNA amplificato nel locus genomico YAP delle linee cellulari PC9 e B2B utilizzando il sistema di editing genetico CRISPR-Cas9.
NOTA: Questa sequenza di DNA integra i filamenti 1-10 di mNeonGreen2 per emettere fluorescenza. La mappa della sequenza genomica di YAP-mNeonGreen21-10/11 è mostrata nella Figura supplementare S1. La mappa contiene le sequenze genomiche, donatori e mNeonGreen2 etichettate. - Controllare l'espressione di mNeonGreen2 progettata da CRISPR/Cas9 utilizzando un microscopio a epifluorescenza (vedere la Tabella dei materiali). Poiché mNeonGreen2 viene etichettato con YAP ogni volta che le cellule esprimono YAP nel contesto della sua rete di regolazione genica nativa, controllare la presenza dell'intensità di fluorescenza in entrambe le cellule ingegnerizzate CRISPR / Cas9 e confrontarla con quella nelle cellule parentali (controllo).
NOTA: per seguire questo protocollo, utilizzare (1) un laser da 488 nm (47,5 mW/mm2) per l'eccitazione, (2) un obiettivo 40x (apertura numerica (NA) = 0,95) e un filtro di emissione passa-banda (ET525/50 nm) per la misurazione della fluorescenza e (3) il software ImageJ per misurare, quantificare e confrontare le intensità di fluorescenza. - Confermare la corretta integrazione di mNeonGreen211 estraendo DNA genomico dalle linee cellulari modificate da CRISPR/Cas9; eseguire la PCR utilizzando primer che affiancano l'inserto genomico e il sequenziamento per confermare l'inserimento al corretto loci genomico19,20.
- Abbattere mNeonGreen211 utilizzando il sistema di editing genetico CRISPR/Cas9 e controllare la riduzione dell'intensità di fluorescenza nelle cellule utilizzando gli stessi sistemi di microscopio e parametri di imaging descritti nella fase 1.3.
NOTA: Questo passaggio conferma la corretta integrazione di mNeonGreen211 mediante il confronto delle intensità di fluorescenza. Le cellule ingegnerizzate CRISPR / Cas9 senza abbattere e le cellule parentali sono utilizzate come controllo. - Raccogli le cellule con la proteina di interesse etichettata attraverso lo smistamento cellulare attivato dalla fluorescenza (FACS).
- Per preparare le cellule per la cernita FACS, tripsilizzarle e risospescarle in soluzione salina tamponata con fosfato (PBS).
- Raccogli le cellule con fluorescenza mNeonGreen2 al di sopra del livello di fondo delle linee cellulari parentali in due cicli arricchenti di smistamento FACS.
NOTA: la sequenza temporale per generare le linee cellulari modificate con CRISPR/Cas9 qui descritte è dell'ordine di 1-2 mesi. Tutte le linee cellulari sono rese pubbliche su richiesta in modo che altri laboratori di ricerca possano riprodurre i risultati.
2. Manutenzione delle celle PC9 e B2B
- Mantenere entrambe le linee cellulari in incubatori di colture tissutali umidificate con il 5% di CO2 a 37 °C.
- Coltura 106 cellule PC9 e Beas2B marcate endogenamente in palloni da 75 cm2 con 12 ml di terreno RPMI-1640 integrato con il 10% di siero bovino fetale e 100 μg/ml di penicillina-streptomicina. Sottocultura di entrambe le linee cellulari quando la confluenza cellulare raggiunge ~ 80%.
- Testare entrambe le linee cellulari per il micoplasma ogni 3 mesi utilizzando un kit di rilevamento del micoplasma, seguendo rigorosamente tutti i protocolli raccomandati dal produttore.
- Conservare le linee cellulari in un congelatore a -80 °C.
- Usa le linee cellulari che sono <20 passaggi dal disgelo per tutti gli esperimenti.
3. Configurazione dell'ambiente hardware e software
- Configurazione dell'ambiente hardware dell'esperimento
- Collegare il controller confocale e il microscopio invertito al computer (vedere la tabella dei materiali).
- Installare la piattaforma software (Tabella dei materiali).
- Accendere il controller confocale e il microscopio invertito. Quindi, avvia Elements.
- Aprire i pannelli di controllo del microscopio confocale, laser e invertito in Elements. Quindi, verificare se i tre pannelli funzionano correttamente testando il movimento dello stadio motorizzato, la commutazione degli obiettivi del microscopio e la scansione spaziale delle linee laser.
- Configurazione dell'ambiente software di AMFIP
- Installare IntelliJ, Java Development Kit 14.0, μManager versione 2.0 gamma e Fiji ImageJ nel computer.
- Aprire il progetto AMFIP scaricato da GitHub (link: https://github.com/njheadshotz/AMFIP) in IntelliJ.
- Fai clic su Impostazioni | | del compilatore Elaborazioni annotazioni e selezionare Abilita elaborazione annotazioni.
- Fare clic su Struttura del progetto | Elementi e creare un file JAR. Impostare la directory di output su mmplugins nella directory μManager .
- Fare clic su Struttura del progetto | Librerie e aggiungere mmplugins e plugin nella directory μManager .
- Fare clic su Aggiungi configurazione nel menu a discesa Esegui e creare un'applicazione.
- Immettere ij. ImageJ nella classe Main.
- Immettere -Xmx3000m -Dforce.annotation.index=true nell'opzione VM.
- Impostare la directory μManager sulla directory Work.
- Fare clic su Esegui per attivare μManager con il plugin AMFIP.
- Collegare μManager con il microscopio invertito.
- Aggiungere il driver adattivo del microscopio invertito21 alla directory μManager .
- Aprire μManager. Fare clic su Dispositivi | Configurazione guidata hardware e creare una nuova configurazione.
- Aggiungere il driver Ti2 in Dispositivi disponibili.
- Selezionare tutte le periferiche e salvare il nuovo file di configurazione.
- Riavviare μManager e selezionare il file di configurazione nel passaggio 3.2.4 in Configurazione di avvio di Micro-Manager.
4. Preparazione del gel
- Trattare il coperchio di vetro con 3-aminopropilmetossisilano per 7 minuti a temperatura ambiente (24 °C).
- Utilizzare acqua deionizzata (DI) per risciacquare il coperchio di vetro e asciugare il coperchio per 20 minuti a 160 °C.
- Trattare la copertura di vetro con glutaraldeide allo 0,5% per 30 minuti e risciacquare con acqua DI.
- Mescolare la soluzione di acrilammide, la soluzione di N,N′-metilenebisacrilammide (bis) e le perle fluorescenti sospese in soluzione salina tamponata con HEPES da 10 mM. Utilizzare la soluzione di persolfato di ammonio al 10% (p/v) e N,N,N′,N′-tetrametiletilendiammina (TEMED) come iniziatori della polimerizzazione. Modificare la percentuale di ciascun componente per ottenere la rigidità meccanica desiderata degli idrogel di poliacrilammide (PAA) seguendo i protocolli stabiliti descritti in precedenza13,14.
NOTA: In questo protocollo, gel da 2 kPa: acrilammide = 12,5% e bis-acrilammide = 6,5%; Gel da 5 kPa: acrilammide = 12,5% e bis-acrilammide = 21,5%; e gel da 40 kPa: acrilammide = 12,5% e bis-acrilammide = 31,5%. Tutte le % elencate sono percentuali di volume. - Dopo 35 minuti, staccare il coperchio di vetro dall'idrogel PAA solidificato e lavare l'idrogel con soluzione salina tamponata HEPES da 50 mM due volte (5 minuti ogni volta).
- Trattare la superficie dell'idrogel con una soluzione idrazina-idrato per 6 ore.
- Risciacquare l'idrogel con acido acetico per 30 min. Rimuovere l'acido acetico e risciacquare con PBS per 30 minuti.
- Ossidare la soluzione di fibronectina (50 μg/mL in PBS) con periodato di sodio per 30 min.
- Rivestire la superficie dell'idrogel con la soluzione ossidata di fibronectina e attendere 35 minuti.
- Aggiungere PBS per immergere l'idrogel e conservare a 4 °C. Coprire tutte le piastre di Petri che contengono gli idrogel con un foglio di alluminio per evitare qualsiasi esposizione leggera agli idrogel.
5. Coltura cellulare
NOTA: Eseguire la coltura cellulare utilizzando la tecnica asettica.
- Legare le coperture di vetro con gli idrogel PAA alla capsula di Petri con fondo di vetro da 35 mm per evitare la deriva fisica dei gel durante i processi di semina e imaging cellulare.
- Utilizzando pinzette pulite sterilizzate, sollevare il coperchio (con l'idrogel PAA in cima) dalla capsula di Petri contenente i gel preparati.
- Utilizzare una salvietta asciutta per pulire le gocce d'acqua sulla superficie inferiore del coperchio di vetro.
- Utilizzare le pinzette sterilizzate per tenere la copertura in vetro.
- Posizionare piccole goccioline (1-5 μL) di colla cianoacrilata ai due angoli diagonali sulla superficie inferiore.
- Utilizzare salviette sterilizzate per rimuovere la colla in eccesso.
- Utilizzare le pinzette sterilizzate per sostituire il coperchio nella capsula di Petri con fondo di vetro. Premere leggermente gli angoli del coperchio per assicurarsi che le goccioline di colla entrino in pieno contatto con la superficie della capsula di Petri.
- Riposizionare il coperchio sulla capsula di Petri per ridurre al minimo l'evaporazione di PBS negli idrogel PAA. Attendere 3 minuti per consentire alla colla di solidificarsi e asciugarsi nella capsula di Petri.
- Riempire la capsula di Petri con 4 ml di PBS.
- Ripetere i passaggi precedenti 5.1.1-5.1.8 per i restanti campioni di idrogel PAA nelle piastre di Petri utilizzate per l'imaging.
- Utilizzare il 75% di etanolo per sterilizzare la superficie esterna di tutte le piastre di Petri e trasferirle nell'armadio di biosicurezza della coltura tissutale. Accendere la luce ultravioletta per 5 minuti e sterilizzare i campioni.
- Semina le cellule sulla superficie superiore del gel.
- Spegni la luce ultravioletta. Estrarre il matraccio (contenente cellule B2B/PC9) dall'incubatore a 37 °C nell'armadio di biosicurezza. Utilizzare una pipetta collegata a una pompa per vuoto per aspirare tutto il terreno di coltura e aggiungere 5 ml di PBS per lavare il pallone.
- Aggiungere 2 ml di tripsina allo 0,05% per staccare le cellule dal fondo del matraccio.
- Posizionare il pallone nell'incubatrice. Attendere 5 minuti.
- Trasferire il pallone nell'armadio di biosicurezza. Aggiungere 8 ml di terreno di coltura fresco al matraccio e la pipetta su e giù più volte per sospendere le cellule in modo omogeneo.
- Trasferire tutti i 10 mL della sospensione cellulare in un tubo da 15 mL e centrifugare a 300 × g per 5 min.
- Controllare il pellet di cella nella parte inferiore del tubo. Inclinare lentamente il tubo orizzontalmente e utilizzare la pipetta aspirante per rimuovere tutto il terreno di coltura dal tubo senza toccare il pellet cellulare. Quindi, aggiungere 8 ml di terreno di coltura fresco e pipettare su e giù più volte fino a quando tutte le cellule sono mescolate in modo omogeneo con il mezzo.
- Depositare 100 μL della sospensione cellulare (150 cellule/μL) sulla superficie del gel e attendere 5 min. Quindi, aggiungere lentamente 4 ml di terreno di coltura fresco alle piastre di Petri; evitare di aggiungere il mezzo fresco direttamente sul gel.
- Posizionare la capsula di Petri nell'incubatrice a 37 °C. Attendere per consentire alle cellule di attaccarsi alla superficie del gel (B2B: 0,5-1 h; PC9: 4-5 ore).
6. Imaging cellulare
NOTA: AMFIP consente l'imaging automatico, multicanale e a lungo termine coordinandosi con diversi sistemi hardware e software: (1) AMFIP manipola μManager per spostare automaticamente lo stadio motorizzato del microscopio Ti2-E su più campi visivi (FOV) e acquisire immagini in campo luminoso attraverso una telecamera monocromatica (Table of Materials); e (2) AMFIP attiva più file macro all'interno di Elements con uno script Java personalizzato per eseguire operazioni automatiche per l'imaging z-stack confocale e la commutazione di diversi canali laser (405 nm e 488 nm).
- Imposta l'ambiente per l'imaging a lungo termine.
- Posizionare la camera ambientale sullo stadio motorizzato del microscopio invertito. Impostare la portata di CO2 a 160 mL/min e regolare la temperatura della camera (in alto: 44 °C; bagno: 42 °C; stadio: 40 °C). Quindi, aggiungere 40 ml di acqua purificata nel bagno della camera.
- Estrarre la capsula di Petri con fondo di vetro con le cellule dell'incubatrice e posizionarla nella camera dell'ambiente.
- Accendere il controller confocale e il microscopio invertito. Cambiare il percorso della luce a destra e osservare le celle che si attaccano usando μManager. Se al gel sono state attaccate cellule sufficienti, trasferire la capsula di Petri nell'incubatrice. Se non abbastanza cellule si sono attaccate al gel, continuare l'incubazione cellulare per altri 30 minuti per B2B e 60 minuti per le cellule PC9.
- Tagliare due piccoli pezzi di nastro adesivo e incollarli sulla camera attorno al foro circolare. Quindi, applicare un po 'di colla adesiva sul nastro (solo sull'area che coprirà la capsula di Petri).
- Estrarre la capsula di Petri dall'incubatrice. Quindi, posizionare lentamente la capsula di Petri nella camera e lasciare che il fondo del piatto entri in contatto con la colla.
- Premere il coperchio della capsula di Petri per 1 minuto per consentire alla colla di entrare in pieno contatto con la capsula di Petri e solidificare. Quindi, spingere delicatamente la capsula di Petri orizzontalmente per confermare che la capsula di Petri è inamovibile nella camera.
- Chiudere il coperchio della camera.
- Impostare i parametri di acquisizione delle immagini per l'imaging in campo luminoso.
- Aprire IntelliJ e impostare un parametro T1 (ad esempio, 120 s) nella riga 93 del file Elements_script.java. Verificare che questo valore sia maggiore del tempo di esecuzione della macro in Elementi utilizzati per l'imaging confocale di un campo visivo (FOV). Fare clic sul pulsante Esegui per avviare il progetto AMFIP IntelliJ.
- Fare clic sul pulsante Live e Multi-D Acq. sull'interfaccia principale di μManager. Quindi, spostare il percorso della luce del microscopio invertito a destra per l'imaging a campo luminoso, passare all'obiettivo 10x e aprire la luce a diodi emettitori di luce (LED) (la sorgente luminosa per l'imaging a campo luminoso; intensità: 5%).
- Fare clic sul percorso della luce, sull'obiettivo del microscopio e sul pulsante della lampada a LED nel pannello Elements Ti2 o premere manualmente i pulsanti corrispondenti sul microscopio.
- Regolare il joystick XY e la manopola del piano Z per trovare la posizione corretta e il piano di messa a fuoco del gel sulla capsula di Petri. Utilizzare un obiettivo 10x per trovare i FOV appropriati di più singole cellule attaccate al gel.
- Selezionare la casella Posizioni multiple (XY) nella finestra Acquisizione multidimensionale . Fare clic sul pulsante Modifica elenco posizioni... e osservare la finestra Elenco posizioni stage che viene visualizzata. Quindi, modificare l'obiettivo su 40x, aumentare l'intensità della luce LED al 15%, regolare nuovamente lo stadio motorizzato XY per individuare i FOV e registrare le coordinate facendo clic sul pulsante Segna nella finestra Elenco posizione stage .
- Registra 67 FOV desiderati. Fare clic sul pulsante Salva con nome... nella finestra Elenco posizioni stage per registrare le coordinate. Immettere T1 (il parametro, ad esempio 120 s, definito nel passaggio 6.2.1) nell'intervallo di tempo di acquisizione dell'immagine a T1 nella sezione Time Points nella finestra Acquisizione multidimensionale .
- Impostare l'acquisizione dell'immagine per le immagini 2D-YAP e perline.
- Apri Elements, cambia il percorso della luce a destra per l'imaging confocale e spegni la luce a LED. Quindi, fare clic sul pulsante Rimuovi interblocco e attivare il canale laser FITC (per l'imaging YAP) selezionando la casella FITC .
- Regolare la velocità di scansione a 1 fotogramma per 2 s facendo clic sul pulsante 1/2 e ruotare la manopola del piano Z per trovare rapidamente la posizione Z delle celle collegate. Registrare i limiti inferiore e superiore per lo Z-stack.
- Fare clic su Macro sulla barra multifunzione superiore, selezionare Editor macro nel menu a discesa Macro e immettere i valori del passaggio 6.3.2 in un file macro.
- Attivare il canale laser 4′,6-diamidino-2-fenilindolo (DAPI) (per l'imaging delle perline) selezionando la casella DAPI per trovare e registrare la posizione Z focalizzata delle perline. Vai all'editor di macro e inserisci i valori registrati nel file macro.
- Impostare il compito di spostare lo stadio motorizzato utilizzando AMFIP.
- Vai su μManager e clicca su Plugins | Automazione per aprire l'interfaccia utente grafica (GUI) di AMFIP. Fare clic sui pulsanti Aggiungi punto o Rimuovi punto per acquisire il numero esatto di FOV selezionati. Immettere le coordinate registrate dei FOV nel pannello Coordinate.
- Definire il tempo totale dell'esperimento nel campo di testo Tempo totale esperimento .
- Fare clic sul pulsante Configurazione tempo aggiuntivo e definire l'intervallo di tempo T2 (ad esempio, 30 minuti) per spostare lo stadio motorizzato su ciascun FOV.
- Ingrandisci le dimensioni della finestra di Elements e trascina la GUI di AMFIP sul lato destro dello schermo per evitare che la GUI disturbi le operazioni automatiche del cursore.
- Fare clic sul pulsante Invio . Al termine della prima macro, fare clic sul pulsante Acquisisci! nella finestra Acquisizione multidimensionale .
- Sciogliere le cellule dopo l'acquisizione dell'immagine.
- Dopo aver terminato l'imaging a lungo termine, interrompere l'attività AMFIP facendo clic sul pulsante Pausa nella finestra del plug-in di automazione e sul pulsante Stop nella finestra Acquisizione multidimensionale .
- Apri Elements e imposta l'imaging Z-stack facendo clic sui pulsanti Superiore e Inferiore nella finestra Acquisizione ND (imposta la gamma Z in modo che sia più grande della gamma Z delle perline). Cambiare il percorso della luce a destra e aprire la luce a LED (intensità: 15%).
- Rimuovere lentamente e con attenzione i coperchi della camera e la capsula di Petri. Nel frattempo, monitora la vista del campo luminoso per qualsiasi deriva del FOV.
- Utilizzando una pipetta di plastica per assorbire 0,5 ml di soluzione di dodecil solfato di sodio (SDS), tenere con attenzione la pipetta di plastica un po 'sopra il terreno di coltura nella capsula di Petri e aggiungere 1-2 goccioline della soluzione SDS nel terreno di coltura.
- Una volta dissolte le celle nella vista a campo luminoso, cambiare il percorso della luce a sinistra, chiudere la luce a LED, fare clic sul pulsante Rimuovi interblocco .
- Eseguire l'imaging Z-stack. Salvare lo stack di immagini e denominarlo come Reference_N (N è il numero di sequenza di ciascun FOV).
- Fare clic sul pulsante Posizioni multiple (XY) nella finestra Acquisizione multidimensionale . Quindi, selezionare il prossimo FOV e fare clic sul pulsante Vai a per spostare lo stadio motorizzato sul secondo FOV.
- Ripetere il passaggio 6.5.7 per ogni FOV.
7. Misurazione del rapporto YAP N/C
- Eseguire l'analisi delle immagini per misurare il rapporto YAP N/C utilizzando il software Fiji ImageJ (Figura 4).
- Apri Fiji ImageJ. Importare lo stack di immagini a campo luminoso per tutti i FOV acquisiti da μManager.
- Apri il menu a discesa Immagine e seleziona Stack | Strumenti | Custode delle fette. Quindi, esporta lo stack di immagini a campo luminoso per ogni FOV.
- Importare l'immagine di fluorescenza del canale FITC e sovrapporla all'immagine a campo luminoso per lo stesso FOV. A tale scopo, scegliere l'immagine fluorescente e selezionare Sovrapposizione | Aggiungi immagine... (Immagine da aggiungere: l'immagine in campo luminoso; La posizione X e Y dipende dalle dimensioni dell'immagine in campo luminoso acquisita da diverse telecamere; Opacità: 60-70).
- Apri il menu a discesa Analizza e seleziona Imposta misure.... Seleziona area; Densità integrata e valore di grigio medio.
- Fare clic sul pulsante Selezioni a mano libera sull'interfaccia principale di ImageJ.
- Disegna il contorno del corpo cellulare e il nucleo desiderato. Quindi, fai clic su Analizza | Misurare o premere il pulsante M sulla tastiera.
- Osservare la finestra Risultati visualizzata. Si noti che i valori sotto la colonna Area rappresentano l'area della regione selezionata (μm2) e i valori sotto la colonna IntDen rappresentano l'intensità di fluorescenza della regione selezionata.
- Calcolare il rapporto YAP N/C utilizzando le seguenti formule (1), (2) e (3):
 (1)
(1) (2)
(2) (3)
(3)
Dove Inuc e Icel rappresentano l'intensità relativa del nucleo e del corpo cellulare, e Anuc e Acel rappresentano l'area del nucleo e il corpo cellulare. R è il rapporto YAP N/C. - Salva i contorni per il futuro calcolo della forza di trazione del dipolo e dello spostamento pericellale/perinucleare. Per fare ciò, fai clic su Analizza | Strumenti | Salva le coordinate XY...
8. Misurazione del campo di trazione
- Applicare la microscopia della forza di trazione tramite i plugin Fiji ImageJ22,23.
- Apri Fiji ImageJ.
- Importare la pila di immagini di perline per un FOV.
- Seleziona la sezione che mostra la distribuzione più chiara delle perline ed estraila cliccando su Immagini | Stack | Strumenti | Custode delle fette.
- Importate la pila di immagini del riferimento per lo stesso FOV.
- Scegliete la sezione con la stessa luminosità e contrasto della sezione nel passaggio 8.1.3. Quindi, estrailo come immagine di riferimento.
- Seleziona immagini | Stack | Strumenti | Concatenate per combinare le due sezioni dei passaggi 8.1.3 e 8.1.5 (selezionate l'immagine di riferimento come prima sezione).
- Seleziona Plugin | | di corrispondenza del modello Allineare le sezioni in stack o plugin | Stabilizzatore d'immagine per allineare le due sezioni.
- Seleziona immagine | Stack | Impila in immagini. Quindi, seleziona immagine | | delle tabelle di ricerca Verde per convertire il colore della prima fetta in verde e selezionare Immagine | | delle tabelle di ricerca Rosso per convertire il colore della seconda fetta in rosso.
- Seleziona immagine | | colore Unisci canali per unire le due immagini.
- Sovrapponi l'immagine con l'immagine in campo luminoso dallo stesso FOV e usa questa immagine sovrapposta per osservare lo spostamento delle perline.
- Seleziona plugin | | PIV PIV iterativo (di base).... Impostare la dimensione della finestra di interrogazione su 128/256; 64/128; 32/64 (almeno quattro perline per finestra di interrogatorio). Impostare la soglia di correlazione su 0,6.
- Fare clic su OK. Al termine del calcolo, salvare il file di testo con i dati grezzi dello spostamento del tallone in una cartella ordinaria creata dall'utente.
- Seleziona Plugin | | FTTC FTTC e scegliere il file di testo nel passaggio 8.1.9.
- Inserisci la dimensione dei pixel (μm), il modulo di Young del gel (Pascal) e la larghezza e l'altezza della trama in base all'esperimento e all'immagine delle perline.
- Fare clic su OK per salvare automaticamente il file di testo contenente i dati grezzi della forza di trazione nella stessa directory del file di testo al passaggio 8.1.12.
- Utilizzare un software di rappresentazione grafica (Table of Materials) per tracciare il campo di trazione con la stessa scala per più celle (Figura 1B,C e Figura 2B,C).
- Inserire il file di testo che contiene i dati grezzi della trazione in un foglio di calcolo.
- Create un nuovo foglio, inserite le coordinate Y di trazione nella prima riga (disponi da valori alti a valori bassi) e le coordinate X nella prima colonna (disponi da bassa ad alta).
- Immettere il valore di trazione per ogni coordinata dai dati grezzi.
- Salvare il foglio nel passaggio 8.2.2 come file *.csv.
- Apri Origin.
- Clicca su File | Aprire e importare il file *.csv nel passaggio 8.2.4. Selezionare tutte le celle e fare clic su Plot | Contorno| Contorno - Riempimento colore.
- Nella finestra Plotting: plotvm selezionare Y tra le colonne per impostare automaticamente i valori Y sulla prima riga e i valori X sulla prima colonna. Quindi, assegna un nome al titolo e fai clic su OK.
- Nella finestra del grafico che si apre, fai doppio clic sulla mappa di calore.
- Fare clic su Livelli nella finestra Colormap/Contorni . Quindi, modificare il livello della scala in un intervallo ragionevole (0300 in questa analisi) e fare clic su OK.
- Fai clic su Linee, deseleziona Mostra solo ai livelli principali e seleziona Nascondi tutto. Quindi, fare clic su OK.
- Fare clic con il pulsante destro del mouse sul grafico e selezionare Esporta grafici.... Salvare l'immagine nel percorso specificato.
- Usa MATLAB per calcolare la trazione della cella di dipolo.
- Salvare il file di testo dei dati grezzi di trazione (dal passaggio 8.1.12) e il file delle coordinate della regione di interesse limite della cella (ROI) (dal passaggio 7.1.9) nella stessa cartella definita nel passaggio 8.1.12. Trasferisci tutti i file MATLAB presenti nel pacchetto AMFIP in questa cartella.
- Apri MATLAB. Aprire la cartella definita al passaggio 8.1.12 e aprire il file della funzione di calcolo della trazione del dipolo absdipole.m trasferito in questa cartella nel passaggio 8.3.1.
- Leggi i due file di testo/csv nel passaggio 8.3.1 nello spazio di lavoro MATLAB e assegna una matrice a due variabili (ad esempio, trazione e roi).
- Eseguire la funzione absdiple (trazione, roi).
NOTA: La prima colonna dell'uscita è la forza di trazione del dipolo in nN (nano-Newton). La seconda colonna dell'uscita è l'angolo della forza di trazione del dipolo rispetto all'asse orizzontale.
Risultati
Distribuzione e dinamica YAP distinte nel cancro PC9 ingegnerizzato CRISPR / Cas9 e nelle cellule normali B2B durante la diffusione cellulare
Immagini rappresentative di fluorescenza della distribuzione YAP in singole cellule B2B e PC9 su gel PAA da 2, 5, 40 kPa e coverslip di vetro sono mostrate in Figura 1A e Figura 2A. La localizzazione nucleare di YAP nelle cellule B2B è aumentata con l'aumentare della rigidità del substrato (Figura 1A), mentre le cellule PC9 hanno mostrato una concentrazione di YAP simile nel nucleo e nel citoplasma su substrati di rigidità variabile (Figura 2A). Immagini rappresentative di fluorescenza della distribuzione YAP in singole cellule B2B e PC9 diffuse sul substrato idrogel da 5 kPa (dalla 0a h alla 10a h dopo le cellule attaccate ai substrati) sono mostrate rispettivamente nella Figura 1B e nella Figura 2B. La cella B2B ha aumentato monotonamente l'area di diffusione nel tempo insieme a una diminuzione del rapporto YAP N / C (Figura 1B), mentre la cella PC9 ha mantenuto un'area di diffusione della cella relativamente invariata, orientamento e rapporto YAP N / C durante il processo di diffusione di 10 ore (Figura 2B). Durante la durata di 10 ore della diffusione precoce, la cellula B2B rappresentativa ha deformato costitutivamente la superficie del substrato e ha applicato la trazione cellulare in evoluzione temporale su tutta l'area cellulare (Figura 1C e Figura 1D).
Al contrario, la cellula PC9 rappresentativa ha sviluppato solo spostamento e trazione alle due estremità del corpo cellulare e la sua trazione è diminuita dopo 7,5 ore (Figura 2C e Figura 2D). Altre immagini time-lapse e misurazioni di trazione delle cellule B2B e PC9 nella fase iniziale di diffusione sono fornite nella Figura supplementare S2 e nella Figura supplementare S3. Sono state osservate anche altre modalità di dinamica cellulare PC9 (Figura 6). Parallelamente a queste diverse caratteristiche di diffusione, le cellule B2B e PC9 hanno mostrato una distribuzione e una dinamica YAP distinte (Figura 3). Su un gel da 5 kPa, YAP nelle cellule B2B è stato concentrato nel nucleo alla 0a h ed è diventato più omogeneamente distribuito in tutto il corpo cellulare alla 10a h. Tuttavia, le cellule PC9 hanno mostrato una distribuzione più omogenea di YAP nel nucleo e nel citoplasma durante tutte le 10 ore del processo di diffusione. Per analizzare quantitativamente l'attività YAP e la traslocazione in celle B2B e PC9, il rapporto YAP N/C è stato calcolato utilizzando l'algoritmo descritto nella Figura 4.
Per studiare ulteriormente la dinamica YAP distinta, sono stati confrontati i cambiamenti temporali nel rapporto YAP N / C, nell'area cellula / nucleo e nella trazione di più cellule B2B singole (n = 10) e cellule PC9 (n = 5). È stato riscontrato che il rapporto YAP N/C medio delle cellule B2B è diminuito da 2,54 ± 0,22 a 1,79 ± 0,21 (n = 10; p = 0,0022**; Figura 5A), mentre il rapporto MEDIO YAP N/C delle celle PC9 è passato da 1,92 ± 0,26 a 1,57 ± 0,07 (n = 5; p = 0,187 (non significativo (ns)); Figura 5A). La trazione media del dipolo delle celle B2B è passata da 256,17 ± 123,69 nN a 287,44 ± 99,79 nN (p = 0,7593 (ns); Figura 5B). La trazione media del dipolo delle celle PC9 è passata da 141,19 ± 33,62 nN a 168,52 ± 73,01 nN (p = 0,7137 (ns); Figura 5B). L'area media di diffusione cellulare delle cellule B2B è aumentata da 613,89 ± 102,43 μm2 a 942,51 ± 226,71 μm2 (p = 0,0512 (ns); Figura 5C).
L'area media di diffusione cellulare delle cellule PC9 è passata da 495,78 ± 97,04 μm2 a 563,95 ± 89,92 μm2 (p = 0,5804 (ns); Figura 5C). L'area media di diffusione del nucleo delle cellule B2B è aumentata da 181,55 ± 36,18 μm2 a 239,38 ± 43,12 μm2 (p = 0,1217 (ns); Figura 5D) e l'area media di diffusione del nucleo delle cellule PC9 è cambiata da 133,31 ± 30,05 μm2 a 151,93 ± 22,49 μm2 (p = 0,5944 (ns); Figura 5D). Questi risultati suggeriscono che (1) le cellule B2B mostrano un rapporto YAP N/C costitutivamente substrato-rigidità-dipendenza; (2) la trazione delle cellule B2B è superiore a quella delle cellule PC9; e (3) a differenza delle cellule B2B, le cellule PC9 mostrano un aumento limitato dell'area cellulare e cambiamenti nel rapporto YAP N / C durante il processo di diffusione di 10 ore.
Correlazione della distribuzione e della dinamica YAP con gli stati di migrazione delle cellule B2B
Sono stati confrontati il rapporto YAP N/C e la trazione di dipolo di tutte le cellule B2B (n=10) e PC9 (n=5) in funzione dell'area di diffusione cellulare e dell'area di diffusione del nucleo. Il rapporto YAP N/C e la trazione del dipolo delle cellule PC9 non erano chiaramente correlati con i loro intervalli di aree di diffusione delle piccole cellule e del nucleo (Figura 6). Al contrario, il rapporto YAP N / C e la trazione del dipolo delle cellule B2B sembravano seguire due tendenze distinte (Figura 6A e Figura 6C), suggerendo che potrebbero esserci due gruppi di cellule B2B che coesistono in questo esperimento. Nel primo gruppo, il rapporto YAP N/C e la trazione del dipolo aumentano insieme all'allargamento dell'area di diffusione cellulare e raggiungono i loro massimi a ~ 1000 μm2 (Figura 6C e Figura 6D, indicata dalla linea tratteggiata gialla). Nel secondo gruppo, il rapporto YAP N/C e la trazione del dipolo aumentano a un ritmo più lento con l'allargamento dell'area di diffusione cellulare e mantengono valori quasi costanti quando l'area di diffusione cellulare continua ad aumentare (Figura 6C, D, indicata dalla linea tratteggiata verde).
Le cellule tumorali PC9 generano trazioni nelle regioni perinucleari
Singole cellule PC9 diffuse spostano i substrati nelle regioni perinucleari, a partire dalla 6a h di coltura (Figura 7C). Per visualizzare lo spostamento perinucleare causato dalla trazione cellulare, abbiamo sovrapposto le immagini di perline fluorescenti scattate prima (rosso) e dopo (verde) la rimozione delle cellule dai substrati (vedi la sezione protocollo per i dettagli). Le perline che non hanno alcuno spostamento appariranno gialle nelle immagini sovrapposte, cioè l'aggiunta di colori rosso e verde. Al contrario, le perline che vengono spostate dalle loro posizioni di riposo a causa della trazione cellulare mostreranno colori verdi e rossi separati.
In particolare, in entrambe le cellule PC9 (Figura 7C, D) e B2B (Figura 7E), lo spostamento delle perline è stato osservato nel citoplasma e all'interno del nucleo, oltre a quelli al confine cellulare. Per evidenziare lo spostamento perinucleare, l'equazione di Boussinesq della teoria dell'elasticità lineare viene utilizzata per prevedere lo spostamento teorico 2D generato da un'ipotetica forza di dipolo al confine cellulare (linea tratteggiata nera in Figura 7B)24. Confrontando questa curva teorica con lo spostamento reale del substrato misurato lungo lo stesso asse (linea tratteggiata bianca nella Figura 7D), gli spostamenti reali all'interno del nucleo sono risultati essere 1,5-8 volte più grandi del valore teorico (Figura 7B), indicando l'esistenza di forza di trazione nelle regioni perinucleari.

Figura 1: Cambiamenti nell'espressione/distribuzione yap, nel campo di spostamento del substrato e nel campo di trazione di una cellula normale B2B su substrati di rigidità variabile e durante la diffusione precoce. (A) L'espressione YAP di una cellula B2B seminata su gel PAA da 2, 5 e 40 kPa e un coperchio di vetro dopo 60 ore dall'attacco iniziale cellula-substrato. (B) La cellula B2B è stata seminata su un gel PAA da 5 kPa e fotografata oltre 10 ore dopo l'attacco iniziale del substrato cellulare. L'espressione YAP è rappresentata dall'intensità della fluorescenza verde. Nota: l'intensità YAP all'interno del nucleo diminuisce gradualmente ma rimane superiore a quella del citoplasma nel tempo. Le barre di colore indicano i livelli di espressione YAP (verde = alta espressione; nero = bassa espressione) in (A) e (B). (C) La deformazione del substrato (sovrapposta all'immagine del campo luminoso) nella posizione della cella è rappresentata dal campo di spostamento in ogni punto temporale. La direzione di spostamento e la magnitudine sono mostrate rispettivamente dalla direzione e dal colore della freccia. Lo spostamento diventa più grande alle estremità del corpo cellulare B2B man mano che aumenta l'area di diffusione cellulare. La barra di colore indica la magnitudine di spostamento (cremisi = magnitudine alta; nero = magnitudine bassa). (D) Campo di trazione (sovrapposto all'immagine in campo luminoso) calcolato dal campo di spostamento. La trazione è concentrata sul confine delle cellule B2B. I contorni punteggiati bianchi e gialli delineano rispettivamente i confini della cellula e del nucleo. La barra di colore indica la magnitudine di trazione (cremisi = magnitudine alta; nero = magnitudine bassa). Barre di scala = 20 μm. Abbreviazioni: YAP = proteina associata a Yes; PAA = poliacrilammide. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Cambiamenti nell'espressione/distribuzione di YAP, nel campo di spostamento del substrato e nel campo di trazione di una cellula tumorale PC9 su substrati di rigidità variabile e durante la diffusione precoce. (A) L'espressione YAP di una cellula PC9 seminata su gel PAA da 2, 5 e 40 kPa e coverslip di vetro dopo 65 ore dall'attacco iniziale del substrato cellulare. (B) La cellula PC9 è stata seminata su un gel PAA da 5 kPa e fotografata oltre 10 ore dopo l'attacco iniziale del substrato cellulare. L'espressione YAP è rappresentata dall'intensità della fluorescenza verde. Nota: l'intensità YAP si stabilizza da 1,5 ore in poi. Le barre di colore indicano i livelli di espressione YAP (verde = alta espressione; nero = bassa espressione) in (A) e (B). (C) La deformazione del substrato (sovrapposta all'immagine a campo luminoso) nella posizione della cella è rappresentata dal campo di spostamento del tallone fluorescente in ogni punto temporale. La direzione di spostamento e la magnitudine sono mostrate rispettivamente dalla direzione e dal colore della freccia. Il campo di spostamento causato dalle celle PC9 è più piccolo di quello causato dalla cella B2B. Durante tutto il processo di diffusione di 10 ore, l'area delle celle PC9 rimane quasi costante. La barra di colore indica la magnitudine di spostamento (cremisi = magnitudine alta; nero = magnitudine bassa). (D) Campo di trazione (sovrapposto all'immagine a campo luminoso) calcolato dal campo di spostamento. La trazione generata da questa cella PC9 rappresentativa diminuisce gradualmente dalla 6a h alla 10a h. I contorni punteggiati bianchi e gialli delineano rispettivamente i confini della cellula e del nucleo. La barra di colore indica la magnitudine di trazione (cremisi = magnitudine alta; nero = magnitudine bassa). Barre di scala = 20 μm. Abbreviazioni: YAP = proteina associata a Yes; PAA = poliacrilammide. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

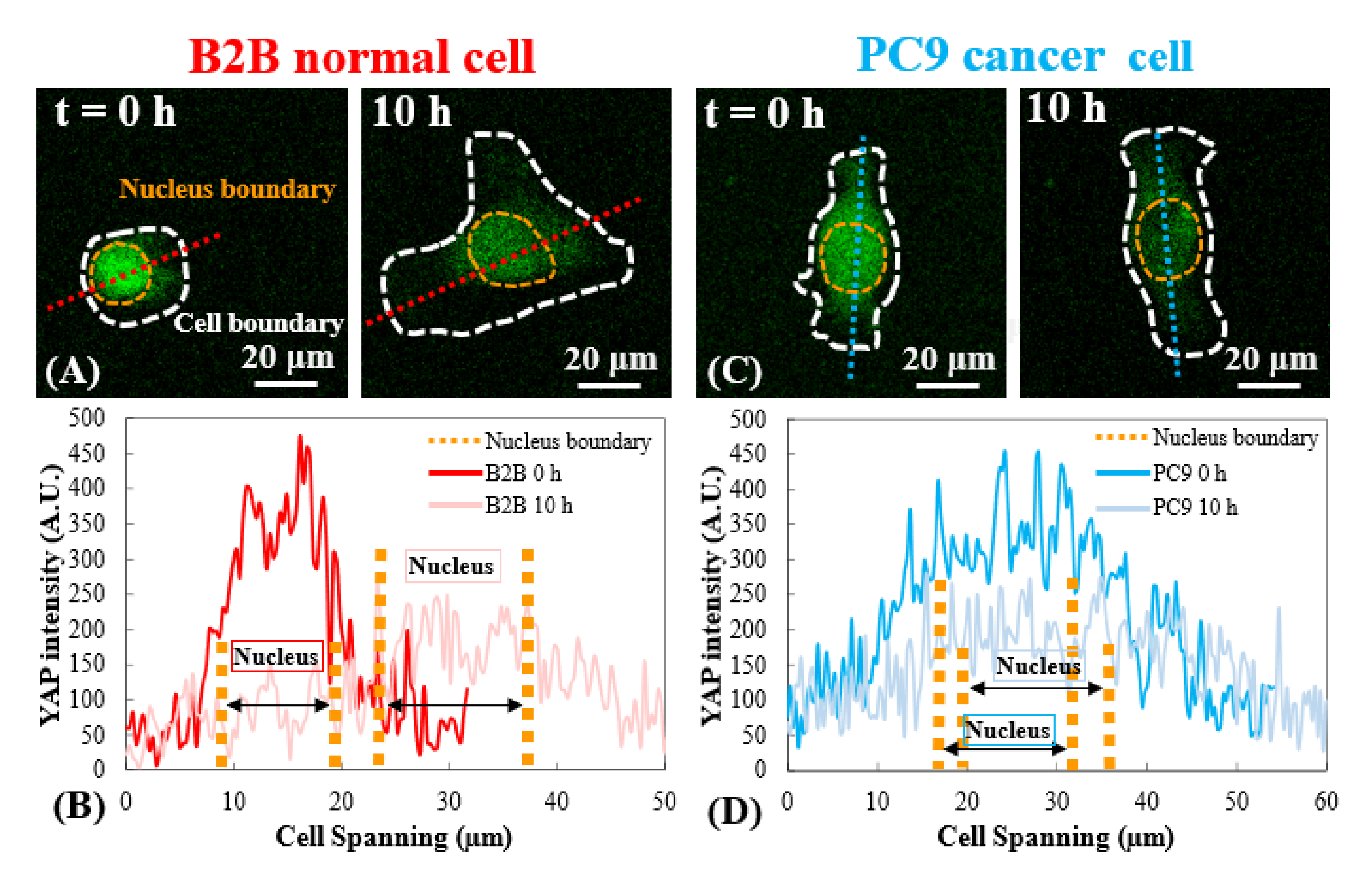
Figura 3: Distribuzione YAP in celle B2B e PC9 nella fase iniziale di diffusione. (A) L'intensità YAP della cellula B2B viene misurata lungo l'asse rosso assegnato alla 0a e alla 10a h. (B) Alla 0a h, l'intensità YAP mostra differenze di concentrazione drammatiche tra il nucleo e il citoplasma. Alla 10a ora, l'intensità yap diventa più omogenea in tutto il corpo cellulare. (C) L'intensità YAP della cellula PC9 viene misurata lungo l'asse blu assegnato alla 0a e alla 10a h. (D) Alla 0a h, l'intensità YAP nel nucleo appare superiore a quella del citoplasma, sebbene la differenza non sia così notevole come quella nelle cellule B2B. Alla 10a h, l'intensità yap nel nucleo appare ancora leggermente superiore a quella del citoplasma, con un andamento di variazione simile a quello della 0a h. Barre di scala = 20 μm (A, C). Abbreviazione: YAP = Proteina associata a Yes. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Misurazione del rapporto YAP N/C. (1) Applicare Fiji ImageJ per disegnare il contorno del nucleo e misurare la sua area proiettata in 2D Anuc. (2) Misurare l'intensità di fluorescenza all'interno del nucleo Inuc. (3) Disegna il contorno del corpo cellulare e misura la sua area proiettata Acel. (4) Misurare l'intensità di fluorescenza all'interno della cellula Icel. (5) Calcolare la densità del nucleo YAP Dnuc, la densità del citoplasma YAP Dcyto e il loro rapporto R: Dnuc = Inuc / Anuc; Dcyto=(Icel-Inuc)/(Acel-Anuc); R=Dnuc/Dcyto. La barra dei colori indica i livelli di espressione YAP (verde = alta espressione; nero = bassa espressione). Barra della scala = 20 μm. Abbreviazioni: YAP = proteina associata a Yes; N = nucleo; C = citoplasma. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Espressione distinta di YAP, morfologia cellulare/nucleo e trazione cellulare nel cancro PC9 e nelle cellule normali B2B durante la diffusione cellulare. (A) Variazione del rapporto YAP N/C durante le prime 10 ore di diffusione a singola cellula. Il rapporto N/C YAP medio delle celle B2B (colonna rossa; n = 10) è cambiato da 2,54 ± 0,22 a 1,79 ± 0,21 (n = 10; p = 0,0022**) mentre il rapporto MEDIO YAP N/C delle celle PC9 (colonna blu; n = 5) è cambiato da 1,92 ± 0,26 a 1,57 ± 0,07 (p = 0,187 (ns)). (B) La trazione media del dipolo in funzione del tempo. La trazione media del dipolo delle celle B2B è cambiata da 256,17 ± 123,69 nN a 287,44 ± 99,79 nN (p = 0,7593 (ns)) e la trazione media del dipolo delle celle PC9 è cambiata da 141,19 ± 33,62 nN a 168,52 ± 73,01 nN (p = 0,7137 (ns)). (C) L'area media della cella in funzione del tempo. L'area media di diffusione cellulare delle cellule B2B è aumentata da 613,89 ± 102,43 μm2 a 942,51 ± 226,71 μm2 (p = 0,0512 (ns)) e l'area media di diffusione cellulare delle cellule PC9 è cambiata da 495,78 ± 97,04 μm2 a 563,95 ± 89,92 μm2 (p = 0,5804 (ns)). (D) L'area media del nucleo in funzione del tempo. L'area media di diffusione del nucleo delle cellule B2B è aumentata da 181,55 ± 36,18 μm2 a 239,38 ± 43,12 μm2 (p = 0,1217 (ns)) e l'area media di diffusione del nucleo delle cellule PC9 è cambiata da 133,31 ± 30,05 μm2 a 151,93 ± 22,49 μm2 (p = 0,5944 (ns)). Abbreviazioni: YAP = proteina associata a Yes; N = nucleo; C = citoplasma; ns = non significativo. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 6: Rapporto YAP N/C e forza di trazione del dipolo in funzione dell'area di diffusione della cellula e del nucleo. Il rapporto YAP N/C e la trazione di dipolo delle celle B2B (n=10) e delle celle PC9 (n=5) sono calcolati dalla 6a h alla 10a h dopo il fissaggio al substrato. (A) Rapporto YAP N/C in funzione dell'area di diffusione cellulare. I rapporti YAP N/C delle cellule B2B variano da 1,16 a 2,53, mentre i rapporti YAP N/C delle cellule PC9 variano da 1,27 a 1,88. L'area di diffusione cellulare delle cellule B2B varia da 391,94 μm2 a 1986,40 μm2. L'area di diffusione cellulare delle celle PC9 varia da 284,46 μm2 a 830,12 μm2. (B) Rapporto YAP N/C in funzione dell'area di diffusione del nucleo. L'area di diffusione del nucleo delle cellule B2B varia da 107,09 μm2 a 514,28 μm2. L'area di diffusione del nucleo delle cellule PC9 varia da 58,03 μm2 a 259,65 μm2. Trazione a dipolo delle cellule B2B in funzione dell'area di diffusione cellulare (C) e dell'area di diffusione del nucleo (D). Le cellule B2B diffuse e non migratorie mostrano una maggiore trazione (da 47,50 nN a 1051,48 nN) con un'area cellulare e del nucleo inferiore. Durante la diffusione e la migrazione, le cellule B2B mostrano una trazione inferiore (da 105,80 nN a 310,28 nN) con intervalli più ampi di area cellulare e nucleo. Abbreviazioni: YAP = proteina associata a Yes; N = nucleo; C = citoplasma. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 7: Spostamento perinucleare in cellule normali B2B e cancerologiche PC9. (A) Schema di scorrimento laterale dello spostamento perinucleare e pericellulare misurato dallo spostamento del tallone nel substrato. (B) Lo spostamento del substrato sotto la cella PC9 viene misurato lungo l'asse della cella (linea tratteggiata bianca in 7D). Lo spostamento teorico generato dalla forza di dipolo al confine cellulare è mostrato dall'equazione di Boussinesq (curva tratteggiata nera). (C) e (D) Immagini di perline fluorescenti sovrapposte con celle (rosse) e senza celle (verdi) per le celle PC9 alla 6a ora dopo l'attacco (vista dall'alto). Le perle gialle (esatta sovrapposizione dei colori rosso e verde) non indicano alcuno spostamento. Le perle verdi e rosse separate (puntate da frecce gialle) rappresentano lo spostamento perinucleare. Le frecce gialle indicano queste macchie peri-nucleo contratte situate alla periferia del nucleo. (E) Spostamento perinucleare generato dalla cellula B2B a 1,5 ore dopo l'attacco cellula-substrato. Barre di scala = 10 μm (C–E). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Figura supplementare S1: La mappa della sequenza genomica di YAP-mNeonGreen21-10/11. Fare clic qui per scaricare questo file.
Figura supplementare S2: Cambiamenti nell'espressione/distribuzione yap, nel campo di spostamento del substrato e nel campo di trazione delle cellule normali B2B durante la diffusione precoce. (A, D, G, J, M) La cellula B2B è stata seminata su un gel PAA da 5 kPa e fotografata oltre 10 ore dopo l'attacco iniziale del substrato cellulare. L'espressione YAP è rappresentata dall'intensità della fluorescenza verde. Nota: l'intensità YAP all'interno del nucleo diminuisce gradualmente ma rimane superiore a quella del citoplasma nel tempo. Le barre di colore indicano i livelli di espressione YAP (verde = alta espressione; nero = bassa espressione) in (A, D, G, J, M). (B, E, H, K, N) La deformazione del substrato (sovrapposta all'immagine del campo luminoso) nella posizione della cella è rappresentata dal campo di spostamento in ogni punto temporale. La direzione di spostamento e la magnitudine sono mostrate rispettivamente dalla direzione e dal colore della freccia. Lo spostamento diventa più grande alla periferia del corpo cellulare B2B man mano che aumenta l'area di diffusione cellulare. Le barre di colore indicano la magnitudine di spostamento (cremisi = magnitudine alta; nero = magnitudine bassa) in (B, E, H, K, N). (C, F, I, L, O) Campo di trazione (sovrapposto all'immagine a campo luminoso) calcolato dal campo di spostamento utilizzando la microscopia della forza di trazione. La trazione è concentrata alla periferia delle cellule B2B. Le barre di colore indicano la magnitudine di trazione (cremisi = alta magnitudine; nero = bassa magnitudine) in (C, F, I, L, O). Barre di scala = 20 μm. Abbreviazioni: YAP = proteina associata a Yes; PAA = poliacrilammide. Fare clic qui per scaricare questo file.
Figura supplementare S3: Cambiamenti nell'espressione/distribuzione di YAP, nel campo di spostamento del substrato e nel campo di trazione delle cellule tumorali PC9 durante la diffusione precoce. (A, D, G, J) La cellula PC9 è stata seminata su un gel PAA da 5 kPa e fotografata oltre 10 ore dopo l'attacco iniziale del substrato cellulare. L'espressione YAP è rappresentata dall'intensità della fluorescenza verde. Nota: l'intensità YAP all'interno del nucleo diminuisce gradualmente ma rimane simile o leggermente inferiore a quella del citoplasma nel tempo. Le barre di colore indicano i livelli di espressione YAP (verde = alta espressione; nero = bassa espressione) in (A, D, G, J). (B, E, H, K) La deformazione del substrato (sovrapposta all'immagine del campo luminoso) nella posizione della cella è rappresentata dal campo di spostamento in ogni punto temporale. La direzione di spostamento e la magnitudine sono mostrate rispettivamente dalla direzione e dal colore della freccia. Lo spostamento diventa più grande alla periferia del corpo cellulare PC9 man mano che aumenta l'area di diffusione cellulare. Le barre di colore indicano la magnitudine di spostamento (cremisi = magnitudine alta; nero = magnitudine bassa) in (B, E, H, K). (C, F, I, L) Campo di trazione (sovrapposto all'immagine a campo luminoso) calcolato dal campo di spostamento. La trazione è concentrata alla periferia delle celle PC9. Le barre di colore indicano la magnitudine di trazione (cremisi = alta magnitudine; nero = bassa magnitudine) in (C, F, I, L). Barre di scala = 20 μm. Abbreviazioni: YAP = proteina associata a Yes; PAA = poliacrilammide. Fare clic qui per scaricare questo file.
Discussione
Il processo di imaging (fase 6.3) è fondamentale per garantire che le immagini di fluorescenza siano di qualità sufficientemente buona da produrre risultati di quantificazione validi. Le immagini z-stack di proteine fluorescenti o perline dovrebbero avere un intervallo z abbastanza grande da includere le immagini a fuoco per tutte le posizioni Z su cui si estende il campione. Un altro passo critico è la raccolta delle immagini di riferimento delle perle fluorescenti dopo aver sciolto le cellule (fase 6.5). Poiché le immagini di riferimento devono essere prese nelle stesse posizioni nel passaggio 6.3, non deve essere indotto alcuno spostamento relativo tra la capsula di Petri, la camera d'ambiente e il microscopio. Gli investigatori che eseguono la fase di dissoluzione devono fare attenzione a rimuovere il coperchio della capsula di Petri e assicurarsi che la perturbazione meccanica applicata non sia abbastanza grande da alterare la posizione del piatto nella camera dell'ambiente.
Di seguito vengono fornite soluzioni per risolvere alcuni errori che potrebbero verificarsi durante gli esperimenti. Se nessuna macro viene attivata dopo aver fatto clic su Invio nel passaggio 6.4, è molto probabile che l'area inferiore sinistra dello schermo sia occupata da una finestra non Elemento. In tal caso, l'area inferiore sinistra della finestra deve essere deselezionata in modo che le macro possano essere attivate in Elements. Un altro errore comune è che le immagini a campo luminoso appaiono nere. Questo problema è causato da un intervallo di tempo insufficiente tra le acquisizioni di immagini a fluorescenza e in campo luminoso. Lievi ritardi nel conteggio del tempo di imaging a fluorescenza possono accumularsi nel tempo e causare notevoli ritardi e interferire con l'imaging a campo luminoso. Una soluzione consiste nel regolare la durata di un ciclo di imaging di tutte le posizioni in modo che sia inferiore (non uguale a) l'intervallo di tempo tra l'inizio di movimenti consecutivi. Questa operazione aggiorna il conteggio del tempo ed elimina l'errore cumulativo all'inizio di ogni ciclo di imaging.
Questa tecnologia di interrogazione completamente ottica supporta (1) una vasta gamma di hardware / software, inclusi ma non limitati a Nikon, (2) diversi tipi di sistemi idrogel convalidati, tra cui gelatina, PEG, Matrigel e gel di collagene I, e (3) la personalizzazione programmabile in base alle diverse esigenze dei ricercatori. Tuttavia, se una qualsiasi delle funzioni di controllo di livello inferiore non è disponibile da un microscopio commerciale, la personalizzazione delle funzioni utilizzando AMFIP diventa difficile. Un'altra limitazione di questa tecnica è la deriva spaziale del campione sia nel piano XY che in quello di messa a fuoco (Z). Sebbene questa limitazione possa essere superata durante la post-elaborazione delle immagini, è essenziale migliorare la funzione di messa a fuoco automatica per correggere la deriva in tempo reale dei campioni. Questo miglioramento aumenterà la produttività del processo di imaging e ridurrà il potenziale errore causato dalla deriva durante gli esperimenti.
I meccanotroduttori, come YAP, possono servire come nuovi bersagli terapeutici per lo sviluppo di promettenti terapie antitumorali25,26,27. Dati emergenti suggeriscono che YAP promuove la proliferazione e l'invasione delle cellule tumorali. La traslocazione YAP indotta dalla meccanica dal citoplasma al nucleo attiva la trascrizione dei geni relativi alla migrazione cellulare, alla proliferazione, all'invasione e all'apoptosi, portando a comportamenti cellulari aberranti28,29,30,31. Questo lavoro mirava a esplorare la potenziale correlazione del rapporto YAP N / C e della meccanica cellulare in due tipiche carcinoma polmonare umano e linee cellulari normali. Durante il periodo di diffusione cellulare di 10 ore, le cellule PC9 mostrano concentrazioni yap simili nel nucleo e nel citoplasma (Figura 3D e Figura 5A). Le cellule B2B mostrano una concentrazione di YAP più elevata nel nucleo rispetto al citoplasma (Figura 3C e Figura 5A). Questa relazione trovata durante la fase di diffusione precoce è diversa dalla maggior parte dei risultati pubblicati che confrontano la concentrazione di YAP nel nucleo tra cellule normali e tumorali. Sebbene non necessariamente nella fase di diffusione precoce, la maggior parte dei risultati pubblicati mostra che YAP è più concentrato nel nucleo delle cellule tumorali che nel nucleo delle cellule normali27,28. Solo uno studio sul cancro al seno ha riportato un'eccezione32 che mostra che YAP è più concentrato nel citoplasma, che concorda con le nostre attuali osservazioni fatte nelle cellule PC9 del cancro del polmone. Per quanto a conoscenza degli autori, questo lavoro è il primo a mostrare un rapporto YAP N / C più basso in una linea cellulare di cancro del polmone umano. Gli autori ipotizzano che la ragione di un rapporto YAP N/C stabile nelle cellule PC9 potrebbe essere dovuta alla bassa variazione nell'area di diffusione cellulare/nucleo e alla trazione nelle cellule PC9 nella fase iniziale di diffusione. La dissezione dei meccanismi molecolari alla base del basso rapporto YAP N/C nelle cellule PC9 e B2B è in corso.
Durante le prime 10 ore di diffusione, queste due linee cellulari mostrano una relazione distinta tra il rapporto YAP N/C, la trazione cellulare e l'area di diffusione (Figura 5). Per le cellule B2B, un rapporto YAP N/C più elevato è correlato con un'area di diffusione cellulare e del nucleo più elevata (Figura 6A,B), che è coerente con i dati riportati di altre cellule normali33. È interessante notare che, sebbene la tendenza allo sviluppo di questa relazione si trovi generalmente in tutte le cellule B2B registrate, si trovano due diversi gradi (alto e basso) di questa relazione. Le cellule B2B che si diffondono e migrano simultaneamente mostrano una trazione inferiore e un'area di diffusione cellulare e nucleo più elevata con un rapporto YAP N / C più elevato (2,05 ± 0,32). Per le cellule B2B che si diffondono e rimangono nella stessa posizione, mostrano una maggiore trazione e un'area di diffusione cellulare e nucleo più bassa con un rapporto YAP N / C inferiore (1,74 ± 0,21). Questi due gradi di relazioni sono dimostrati nei gruppi di dati sparsi biforcati (Figura 6C,D). Come riportato in letteratura, le cellule normali stazionarie, come le cellule embrionali del fibroblasto NIH 3T3, hanno una trazione maggiore rispetto alle cellule migratorie34. I dati riportati in questo documento suggeriscono che le cellule B2B che si diffondono e non migrano hanno applicato una trazione più elevata rispetto alla diffusione e alla migrazione delle cellule B2B, suggerendo probabilmente che è necessaria un'elevata trazione affinché le cellule non migratorie si stabilizzino sul substrato.
Inoltre, questi dati mostrano che le cellule B2B normali stazionarie generano una forza perinucleare più elevata, mentre ricerche precedenti condotte da altri ricercatori hanno riportato solo una maggiore trazione cellulare generata alla periferia delle cellule stazionarie34,35,36,37. Gli autori pensano che la differenza nella tendenza intrinseca della migrazione negli esperimenti potrebbe causare questi risultati contraddittori. Negli esperimenti pubblicati, il micropatterning di forma quadrata era stato utilizzato per confinare le singole cellule dalla diffusione e inibire la migrazione; non si sa se le cellule avessero la tendenza a migrare. Poiché le cellule migratorie mostrano spesso un'elevata forza di trazione alla periferia delle cellule38, è probabile che le cellule con la tendenza a migrare manterranno comunque un'elevata trazione periferica anche se la loro migrazione è limitata. In questo studio, le cellule stazionarie non sono limitate da alcun micropattern ma non migrano, indicando che le cellule tendono a mantenere il loro stato non migratorio. Un'altra possibilità è che la forma cellulare definita dal micropattern possa influenzare la distribuzione delle aderenze focali e delle forze di trazione39. I risultati di questo studio sono stati generati senza alcun micropatterning confinante e rappresentano la distribuzione della forza delle cellule stazionarie nella loro forma originale.
Per quanto a conoscenza degli autori, solo una pubblicazione fino ad oggi ha riportato specificamente la scoperta di forze perinucleari in cellule normali (fibroblasti embrionali di topo), potenzialmente causate dal cappuccio di actina che si estende attraverso il nucleo40. La traslocazione del citoplasma-nucleo YAP è correlata con l'aumento della forza perinucleare40. Una ricerca approfondita della letteratura pertinente non ha prodotto alcuna pubblicazione che riporti una forza perinucleare o il cappuccio di actina nelle cellule tumorali. Uno studio indiretto sulle cellule tumorali del melanoma ha dimostrato che il bordo dell'actina (un'altra organizzazione di actina perinucleare situata intorno ma che non copre il nucleo) riduce i tassi di migrazione cellulare41, suggerendo indirettamente l'esistenza di una forza perinucleare. Tuttavia, non sono riportati dati sperimentali diretti. In questo studio, gli autori hanno scoperto che sia le cellule PC9 che B2B mostrano spostamento e trazione perinucleari. I meccanismi della generazione delle forze perinucleari e i loro effetti rimangono controversi. Nelle cellule normali, è stato riportato che il cappuccio di actina svolge un ruolo nella regolazione della morfologia del nucleo e dell'organizzazione della cromatina42, trasmettendo segnali meccanici dalle aderenze focali nel nucleo attraverso i linker del complesso nucleoscheletro e citoscheletro (LINC)43 e regolando la migrazione cellulare44. Lamin A/C è correlata alla formazione e all'interruzione del cappuccio di actina40,41,42,43,44. Tuttavia, il rapporto che affermava che il cappuccio di actina genera una forza perinucleare non considerava il ruolo potenziale dell'actina rim40. Nelle cellule tumorali, la sovraespressione di Lamin A facilita la formazione di un bordo di actina e limita la migrazione delle cellule tumorali. La sovraespressione di Lamin B riduce la formazione di actin rim e favorisce la migrazione. La forza perinucleare potrebbe essere coinvolta in questo processo a causa dell'esistenza di un'organizzazione di actina perinucleare e dell'effetto di Lamin A. Tuttavia, i risultati di questo studio non hanno mostrato alcuna evidenza di forze perinucleari misurate o del comportamento del cappuccio di actina. Pertanto, la scoperta di forze peri-nucleari nelle cellule PC9 in questo presente studio è il primo rapporto che mostra forze peri-nucleari e spostamenti nelle cellule tumorali polmonari. Gli autori stanno attualmente studiando i meccanismi molecolari e le funzioni delle forze perinucleari nelle cellule PC9 e B2B ingegnerizzate CRISPR / Cas9.
Oltre all'interrogazione di meccanobiologia completamente ottica che è dimostrata in questo articolo, il sistema multifunzionale integrato può essere applicato per sondare otticamente una miriade di altri segnali fisiologici e patobiologici essenziali nei sistemi viventi. Ad esempio, il laboratorio degli autori ha recentemente stabilito più linee cellulari tumorali umane stabilmente trasdotte che co-esprimono tre proteine di membrana sensibili alla luce: indicatore di tensione di membrana QuasAr2 (eccitazione: 640 nm; emissione: 660 nm-740 nm), depolarizzatore di tensione di membrana CheRiff (eccitazione: 488 nm) e iperpolarizzatore di tensione di membrana eNpHR3 (eccitazione: 590 nm). Queste tre proteine funzionali possono essere attivate da linee laser ortogonali a spettro in modo privo di diafonia, consentendo comunicazioni di segnalazione bidirezionali completamente ottiche (lettura e controllo) dell'elettrofisiologia a membrana. Utilizzando un sistema optoelettronico integrato e un patch-clamp manuale, gli autori hanno convalidato il controllo completamente ottico e la lettura della tensione di membrana (Vm) in singole cellule tumorali umane e sferoidi tumorali multicellulari. L'interrogazione di elettrofisiologia completamente ottica apre la possibilità di esplorazioni dettagliate della bioelettricità precedentemente inaccessibile nelle cellule tumorali, che possono aiutare a far progredire la biologia tumorale da un nuovo asse.
Divulgazioni
Non ci sono conflitti di interesse da dichiarare.
Riconoscimenti
Questo progetto è sostenuto finanziariamente dal Cancer Pilot Award dell'UF Health Cancer Center (X. T. e D. S.) e dal Gatorade Award Start-up Package (X. T.). Gli autori apprezzano sinceramente le discussioni intellettuali e i supporti tecnici del Dr. Jonathan Licht (UFHCC), Dr. Rolf Renne (UFHCC), Dr. Ji-Hyun Lee (Biostatistica, UF), Dr. Hugh Fan (MAE, UF), Dr. Warren Dixon (MAE, UF), Dr. Ghatu Subhash (MAE, UF), Dr. Mark Sheplak (MAE & ECE, UF), Dr. Malisa Sarntinoranont (MAE, UF), Dr. Scott Banks (MAE, UF), Dr. Matthew Traum (MAE, UF), Dr. David Hahn (Università dell'Arizona), Dr. Weihong Wang (Oracle Corporation), Dr. Youhua Tan (Hong Kong Polytechnic University) e il team di supporto di Nikon (Drs. Jose Serrano-Velez, Larry Kordon e Jon Ekman). Gli autori sono profondamente grati per il generoso ed efficace sostegno da parte di tutti i membri dei laboratori di ricerca di Tang, Siemann e Guan e di tutti i membri dello staff dei dipartimenti MAE & ECE & Physics & Radiation Oncology, UF.
Materiali
| Name | Company | Catalog Number | Comments |
| (3-Aminopropyl)triethoxysilane | Sigma-aldrich | 440140 | |
| 0.05 % Trypsin | Corning | 25-051-CI | |
| 75 cm2 flask | Corning | 430641U | |
| 8 Benchtop Centrifuge | Thermo | 75007210 | |
| A1R confocal system | Nikon | HD25 | |
| Acetic acid | Sigma-aldrich | 695092 | glacial, ACS reagent, ≥99.7% |
| BEAS-2B (B2B) cells | Sigma-aldrich | 95102433 | human epithelial cells from lung tissue |
| Carboxylate-Modified Microspheres | Invitrogen | F8797 | |
| Culture medium (RPMI-1640) | Gibco | 11875093 | |
| Desktop Computer | Dell | 2018 | with Windows 10 operating system |
| Environmental chamber TIZB | Tokai Hit | TIZB | |
| Fetal bovine serum (FBS) | Gibco | 26140 | |
| Fibronectin Human Protein, Plasma | Gibco | 33016015 | |
| Fiji ImageJ | National Institutes of Health and the Laboratory for Optical and Computational Instrumentation | 1.53k | |
| Glass-bottom petri dish | MatTek | P35G-1.5-14-C | |
| HEPES buffered saline | Sigma-aldrich | 51558 | |
| Hydrazine hydrate solution | Sigma-aldrich | 53847 | |
| IntelliJ IDEA | JetBrains | 2020 | Java development platform |
| Java Development Kit | Oracle | 14.0 | |
| Kimwipe | Kimtech Science | 3066-05 | |
| MATLAB | MathWorks | 2020b | |
| Monochrome Camera | FLIR | BFS-U3-70S7M-C | |
| MycoAlert Mycoplasma Detection Kit | Lonza | LT07-218 | |
| N,N′-Methylenebisacrylamide solution | Sigma-aldrich | M1533 | |
| NIS-Elements software platform | Nikon | 4.50 | software platform |
| Origin | OriginLab | OriginPro 2017 (Learning Edition) | data analysis and graphing software |
| Penicillin-streptomycin | Gibco | 15140122 | |
| PC9 cells | Sigma-aldrich | 90071810 | human adenocarcinoma cells from lung tissue |
| Phosphate buffered saline (PBS) | Gibco | 10010023 | |
| Phusion High-Fidelity DNA Polymerase | New England Biolabs | F-553S | high-fidelity DNA polymerase |
| Scotch tape | Scotch | adhesive tape | |
| Sodium dodecyl sulfate solution | Sigma-aldrich | 05030 | |
| Super glue | Gorilla | cyanoacrylate glue | |
| Ti2-E inverted microscope | Nikon | MEA54000 | |
| TI2-S-SE-E Motorized Stage with Encoder | Nikon | MEC56120 | |
| μManager | version 2.0 gamma | open source microscopy software (https://micro-manager.org/) |
Riferimenti
- Werley, C., Boccardo, S., Rigamonti, A., Hansson, E., Cohen, A. Multiplexed optical sensors in arrayed islands of cells for multimodal recordings of cellular physiology. Nature Communications. 11 (1), 3881 (2020).
- Yang, B., et al. Epi-illumination SPIM for volumetric imaging with high spatial-temporal resolution. Nature Methods. 16 (6), 501-504 (2019).
- Saraswathibhatla, A., Galles, E. E., Notbohm, J. Spatiotemporal force and motion in collective cell migration. Scientific Data. 7 (1), 197 (2020).
- Saraswathibhatla, A., Henkes, S., Galles, E. E., Sknepnek, R., Notbohm, J. Coordinated tractions control the size of a collectively moving pack in a cell monolayer. Extreme Mechanics Letters. 48, 101438 (2021).
- Wang, W., Kim, C. K., Ting, A. Y. Molecular tools for imaging and recording neuronal activity. Nature Chemical Biology. 15 (2), 101-110 (2019).
- Eliceiri, K. W., et al. Biological imaging software tools. Nature Methods. 9 (7), 697-710 (2012).
- Carpenter, A. E., Kamentsky, L., Eliceiri, K. W. A call for bioimaging software usability. Nature Methods. 9 (7), 666-670 (2012).
- Skylaki, S., Hilsenbeck, O., Schroeder, T. Challenges in long-term imaging and quantification of single-cell dynamics. Nature Biotechnology. 34 (11), 1137-1144 (2016).
- Lin, M. Z., Schnitzer, M. J. Genetically encoded indicators of neuronal activity. Nature Neuroscience. 19 (9), 1142-1153 (2016).
- Luo, Q., et al. Automatic multi-functional integration program (AMFIP) towards all-optical mechanobiology interrogation. bioRxiv. , (2021).
- Edelstein, A., Amodaj, N., Hoover, K., Vale, R., Stuurman, N. Computer control of microscopes using manager. Current Protocols in Molecular Biology. 92 (1), 14-20 (2010).
- Tulpule, A., et al. Kinase-mediated RAS signaling via membraneless cytoplasmic protein granules. Cell. 184 (10), 2649-2664 (2021).
- Tang, X., Tofangchi, A., Anand, S. V., Saif, T. A. A novel cell traction force microscopy to study multi-cellular system. PLOS Computational Biology. 10 (6), 1003631 (2014).
- Tang, X., et al. Mechanical force affects expression of an in vitro metastasis-like phenotype in HCT-8 cells. Biophysical Journal. 99 (8), 2460-2469 (2010).
- Guimarães, C. F., Gasperini, L., Marques, A. P., Reis, R. L. The stiffness of living tissues and its implications for tissue engineering. Nature Reviews Materials. 5, 351-370 (2020).
- Phelps, E. A., et al. Maleimide cross-linked bioactive PEG hydrogel exhibits improved reaction kinetics and cross-linking for cell encapsulation and in situ delivery. Advanced Materials. 24 (1), 64-70 (2012).
- Bajaj, P., Tang, X., Saif, T. A., Bashir, R. Stiffness of the substrate influences the phenotype of embryonic chicken cardiac myocytes. Journal of Biomedical Materials Research. Part A. 95 (4), 1261-1269 (2010).
- Temples, M. N., Adjei, I. M., Nimocks, P. M., Djeu, J., Sharma, B. Engineered three-dimensional tumor models to study natural killer cell suppression. ACS Biomaterials Science & Engineering. 6 (7), 4179-4199 (2020).
- Feng, S., et al. Improved split fluorescent proteins for endogenous protein labeling. Nature Communications. 8, 370 (2017).
- Guan, J., Liu, H., Shi, X., Feng, S., Huang, B. Tracking multiple genomic elements using correlative CRISPR imaging and sequential DNA FISH. Biophysical Journal. 112 (6), 1077-1084 (2017).
- . Micro-Manager Available from: https://micro-manager.org/wiki/NikonTi2 (2021)
- Schindelin, J., et al. Fiji: An open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Martiel, J. L., et al. Measurement of cell traction forces with ImageJ. Methods in Cell Biology. 125, 269-287 (2015).
- Okumurai, I. A. On the generalization of Cerruti's problem in an elastic half-space. Doboku Gakkai Ronbunshu. 1995, 1-10 (1995).
- Piccolo, S., Dupont, S., Cordenonsi, M. The biology of YAP/TAZ: hippo signaling and beyond. Physiological Reviews. 94 (4), 1287-1312 (2014).
- Hong, W. W., Guan, K. L. The YAP and TAZ transcription co-activators: Key downstream effectors of the mammalian Hippo pathway. Seminars in Cell and Developmental Biology. 23 (7), 785-793 (2012).
- Zanconato, F., Cordenonsi, M., Piccolo, S. YAP/TAZ at the roots of cancer. Cancer Cell. 29 (6), 783-803 (2016).
- Wang, Y., et al. Overexpression of yes-associated protein contributes to progression and poor prognosis of non-small-cell lung cancer. Cancer Science. 101 (5), 1279-1285 (2010).
- Li, H., et al. Inhibition of YAP suppresses CML cell proliferation and enhances efficacy of imatinib in vitro and in vivo. Journal of Experimental & Clinical Cancer Research. 35 (1), 134 (2016).
- Tang, X., et al. A mechanically-induced colon cancer cell population shows increased metastatic potential. Molecular Cancer. 13, 131 (2014).
- Panciera, T., Azzolin, L., Cordenonsi, M., Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nature Reviews Molecular Cell Biology. 18 (12), 758-770 (2017).
- Yuan, M., et al. Yes-associated protein (YAP) functions as a tumor suppressor in breast. Cell Death and Differentiation. 15 (11), 1752-1759 (2008).
- Koushki, N., et al. Lamin A redistribution mediated by nuclear deformation determines dynamic localization of YAP. bioRxiv. , (2020).
- Chang, S. S., Rape, A. D., Wong, S. A., Guo, W. H., Wang, Y. L. Migration regulates cellular mechanical states. Molecular Biology of the Cell. 30 (26), 3104-3111 (2019).
- Lee, J., Abdeen, A. A., Tang, X., Saif, T. A., Kilian, K. A. Geometric guidance of integrin mediated traction stress during stem cell differentiation. Biomaterials. 69, 174-183 (2015).
- Lee, J., Abdeen, A., Tang, X., Saif, T. A., Kilian, K. A. Matrix directed adipogenesis and neurogenesis of mesenchymal stem cells derived from adipose tissue and bone marrow. Acta Biomaterialia. 42, 46-55 (2016).
- Tang, X., Bajaj, P., Bashir, R., Saif, T. A. How far cardiac cells can see each other mechanically. Soft Matter. 7 (13), 6151-6158 (2011).
- Dembo, M., Wang, Y. L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophysical Journal. 76 (4), 2307-2316 (1999).
- Rape, A., Guo, W. H., Wang, Y. L. The regulation of traction force in relation to cell shape and focal adhesions. Biomaterials. 32 (8), 2043-2051 (2011).
- Shiu, J. Y., Aires, L., Lin, Z., Vogel, V. Nanopillar force measurements reveal actin-cap-mediated YAP mechanotransduction. Nature Cell Biology. 20 (3), 262-271 (2018).
- Fracchia, A., Asraf, T., Salmon-Divon, M., Gerlitz, G. Increased lamin B1 levels promote cell migration by altering perinuclear actin organization. Cells. 9 (10), 2161 (2020).
- Ramdas, N. M., Shivashankar, G. V. Cytoskeletal control of nuclear morphology and chromatin o1rganization. Journal of Molecular Biology. 427 (3), 695-706 (2015).
- Khatau, S. B., et al. A perinuclear actin cap regulates nuclear shape. Proceedings of the National Academy of Sciences of the United States of America. 106 (45), 19017-19022 (2009).
- Kim, D. H., Cho, S., Wirtz, D. Tight coupling between nucleus and cell migration through the perinuclear actin cap. Journal of Cell Science. 127 (11), 2528-2541 (2014).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati