
JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
배양된 초파리 뇌 외래 식물에서 신경 줄기 세포 재활성화
요약
배양된 초파리 뇌 외식편에서 정동작 신경줄기세포를 재활성화시키는 방법이 확립되었다. 이 방법을 사용하여, 전신 신호의 역할은 신경 줄기 세포 정지, 출입의 조절에서 조직 내재적 신호로부터 결합되지 않을 수 있다.
초록
신경 줄기 세포 (NSCs)는 증식, 분화, 아폽토시스를 겪고, 심지어 정지를 출입 할 수있는 능력을 가지고 있습니다. 이러한 과정의 대부분은 NSC 내재적 유전 프로그램과 NSC 외인성 인자, 국소 및 전신 간의 복잡한 상호 작용에 의해 제어됩니다. 유전 모델 유기체에서, Drosophila melanogaster, NSCs, 신경 세포 (NBs)로 알려진, 정지에서 증식 동안 배아에서 애벌레로 전환. 이 기간 동안 애벌레는 달걀 껍질에서 나와서식이 영양소를 찾아 기어 다니기 시작합니다. 동물 먹이에 대한 반응으로, 지질 저장 능력을 가진 내분비 기관 인 지방 몸체는 순환하는 용혈림프로 전신적으로 방출되는 신호를 생성합니다. 지방 신체 유래 신호 (FBDS)에 반응하여 초파리 인슐린 유사 펩티드 (Dilps)가 생성되어 뇌 신경 분비 뉴런 및 글리아교에서 방출되어 NB와 그 아교 및 기관 틈새 시장에서 PI3-키나제 성장 신호 전달의 하류 활성화로 이어집니다. 이것이 NB가 정지에서 확산으로 전환하는 방법에 대한 현재 모델이지만, FBDS 외인성 큐의 본질은 여전히 애매합니다. NB 외인성 전신 단서가 정지로부터의 출구를 조절하는 방법을 더 잘 이해하기 위해, 동물 먹이를 먹기 전에 시험관 내에서 초기 애벌레 뇌를 배양하는 방법이 개발되었습니다. 이 방법을 이용하면, 외인성 인자가 배양 배지에 공급될 수 있고 NB는 정지 분석으로부터 빠져나갈 수 있다. 우리는 외인성 인슐린이 전뇌 외식식물에서 정지로부터 NB를 재활성화시키기에 충분하다는 것을 발견했습니다. 이 방법은 대규모 스크린에 매우 적합하기 때문에 NB 정지 대 증식 결정을 조절하는 추가적인 외인성 단서를 식별하는 것을 목표로합니다. NSC 증식 결정을 조절하는 유전자와 경로가 진화적으로 보존되기 때문에, 이 분석의 결과는 클리닉에서 재생 요법을 개선하는 데 대한 통찰력을 제공할 수 있다.
서문
줄기 세포는 재생 의학 1,2에 사용할 수있는 잠재력 때문에 큰 관심이 있습니다. 많은 동물, 특히 오래 사는 동물은 성인 조직 내에서 줄기 세포를 유지합니다. 이러한 상주 줄기 세포는 조직 항상성을 유지하는 기능을 하며 신체적 상해 또는 질병 3,4 이후의 복구에 활용된다. 성체 동물에서 대부분의 줄기 세포는 정지, 성장 신호 전달의 세포 주기 정지 및 불활성화를 특징으로 하는 비교적 휴면 상태이다 5. 외인성 단서에 반응하여 줄기 세포는 정지에서 빠져 나와 세포주기에 들어가 조직 유형에 특유한 딸 자손을 생성하기 시작합니다. 예를 들어, 효과적인 면역 반응을 탑재하기 위해, 항원-제시 세포는 정지된 나이브 T 세포가 세포 주기로 진입하도록 유도하고 clonally 확장6. 골격근 손상에 반응하여, 근육 위성 줄기 세포는 세포주기에 들어가 손상된 myofibrils 5,7을 대체하기 위해 딸 근아세포를 생성합니다. 정지 줄기 세포가 외인성 신호에 반응한다는 것은 분명하지만, 많은 경우 외인성 단서의 본질은 큐 유도 줄기 세포 활성화의 메커니즘뿐만 아니라 불분명하다. 정지 된 줄기 세포가 외인성 단서에 어떻게 반응하고 세포주기에 진입하는지에 대한 더 나은 이해를 얻는 것은 클리닉에서 더 나은 줄기 세포 치료법의 개발에 도움이되고 과학적 지식을 증가시킬 것입니다.
수십 년 동안 모델 유기체는 발달 및 성인기에 줄기 세포 증식을 조절하는 유전자 및 세포 신호 전달 경로를 밝히는 데 사용되어 왔습니다. 초파리에서 신경 줄기 세포 (NBs)로 알려진 신경 줄기 세포 (NSCs)는 발달 전반에 걸쳐 분열하여 궁극적으로 통합되는 모든 뉴런과 글리아교를 생성하여 뇌 기능에 필요한 신경 회로를 형성합니다 8,9. 다른 줄기 세포와 마찬가지로, NB는 비대칭적으로 분열하여 자기 갱신하고, 어떤 경우에는 대칭적으로 줄기 세포 풀을 확장합니다. NB는 배아 발생 중에 지정되며 대부분은 모성 영양소 저장소의 감소와 함께 끝까지 정지에 들어갑니다 (그림 1). 배아 발생이 완료되면 애벌레가 부화하여 먹이를 먹기 시작합니다. 동물 먹이에 대한 응답으로, NBs는 정지로부터 재활성화되고 세포 분열을 재개한다 10,11,12,13,14,15,16. 초파리 CNS는 비교적 간단하고 NB가 정해진 시간에 정해지기 때문에 초파리를 사용하여 정지, 출입 조절을 조사하는 것이 이상적임이 입증됩니다.

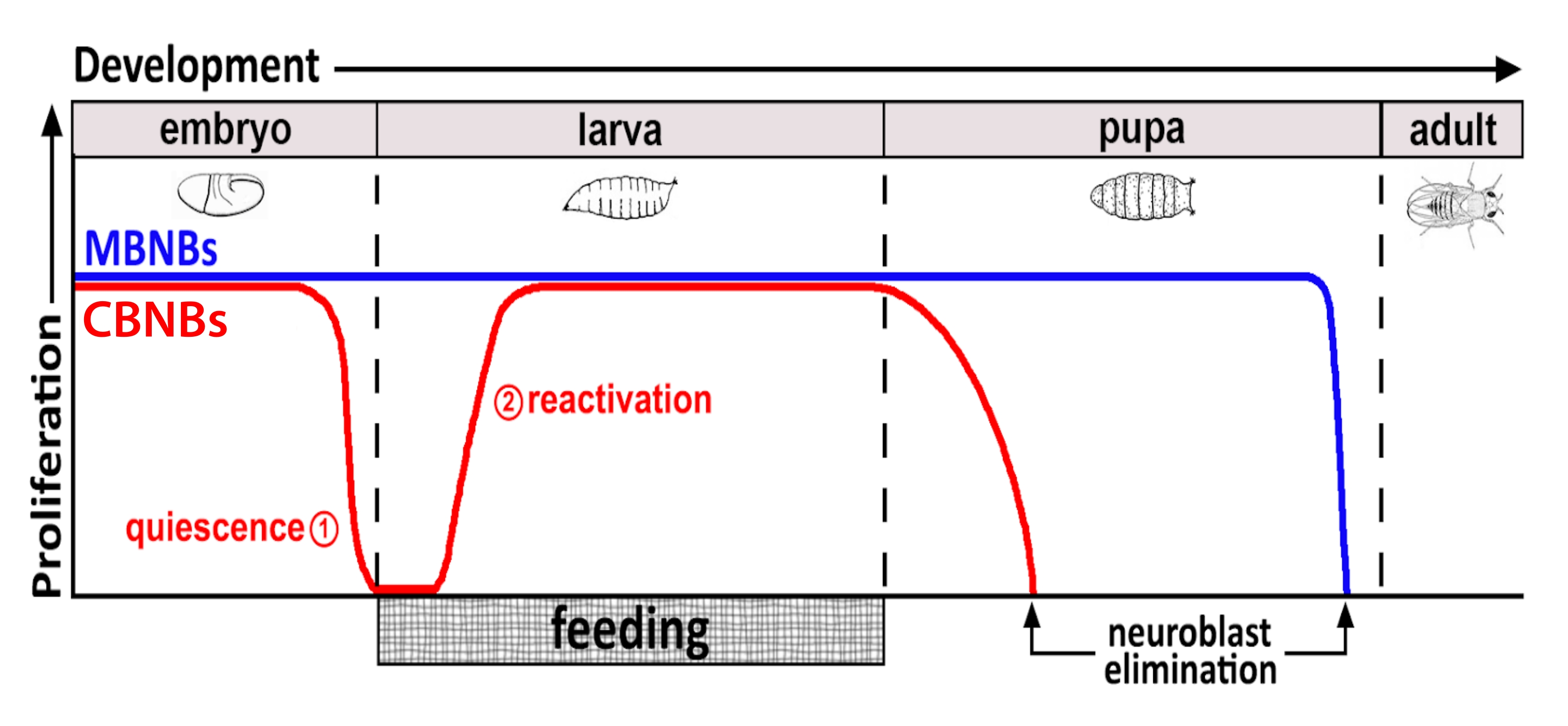
그림 1: 발달 시간 동안 CB NB(중추 뇌 신경아세포, 적색) 및 MB NB(버섯체 신경아세포, 파란색)의 상대적 증식. 배아 발생이 끝나면 대부분의 NB (빨간색 선)는 증식을 멈추고 정지에 들어갑니다. 갓 부화 한 애벌레가 첫 번째 완전한 식사를 섭취 할 때까지 정지가 계속됩니다. 이 방법론의 초점 지점은 빨간색 원 (1, 정지 및 2, 재활성화)으로 표시됩니다. MB NB (파란색)는 발달 기간 동안 지속적으로 분열하는 중추 뇌 NB의 하위 집합입니다 (뇌 반구 당 4 개). 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
동물 먹이에 대한 응답으로, PI3-키나제 및 TOR 성장 신호전달 경로는 NB와 그들의 신경교 및 기관 틈새10,11,15,16에서 활성화된다. 식이 영양소가 철회되거나 PI3-키나아제 수치가 감소하면 NB가 재활성화되지 않고 아교세포와 기관의 성장도10,11,15,16 감소합니다. 현재의 모델은 NB 재활성화가 지방 체내에 의한 애벌레 성장에 결합되며, 이는 동물 먹이12,17,18에 대한 반응으로 전신 신호를 방출한다고 가정한다. 이 신호는 여전히 애매한 상태로 남아 있으며, 뇌에서 초파리 인슐린 유사 펩티드 (Dilps)의 발현과 방출을 촉진하여 NB에서 PI3-kinase의 하류 활성화와 신경교 및 기관 틈새 시장으로 이어집니다. 전신 단서의 본질을 더 잘 이해하기 위해, 우리는 배양 된 뇌 외식에서 정동작 NB를 재활성화하는 방법을 개발했습니다. 이 방법을 사용하면 NB의 재활성화는 전체 동물 전신 단서가없는 상태에서 분석 할 수 있습니다. 외인성 인자는 티미딘 유사체, EdU의 혼입에 기초한 배양 배지 및 NB 재활성화 검정에 재공급될 수 있다. 이 방법을 사용하여, 우리는 외인성 인슐린이 뇌 외식식물에서 정동작 NB를 재활성화시키기에 충분하다는 것을 결정했다. 앞으로의 연구는 다시 추가 될 때 뇌 외식에서 NB 정지를 긍정적으로 또는 부정적으로 조절하는 추가 요인을 확인하는 것을 목표로합니다.
프로토콜
1. 초파리 유 충 수집
참고 : 시작하기 전에 효모 판, 포도 페이스트 및 플라이 콘도를 준비하십시오.
- 효모 페이스트 : 작은 용기에 활성 건조 효모 5g을 물 10mL와 섞어 땅콩 버터의 일관성이있는 페이스트를 형성하십시오. 효모 페이스트를 플라스틱 랩으로 덮고 고무 밴드를 사용하여 용기에 단단히 부착하십시오.
참고 : 신선한 효모 페이스트는 용기에서 팽창하고 단단히 부착되지 않으면 뚜껑에서 튀어 나옵니다. 효모 페이스트는 실온 (RT)에서 며칠 동안 지속됩니다. - 포도 접시 : 포도 접시를 만들기위한 조리법을 따르십시오 (표 1). 4°C에 보관된 플레이트를 사용하는 경우, 사용하기 전에 플레이트를 RT에 1시간 동안 배치하여 미리 따뜻하게 해야 합니다.
- 물(750 mL)과 한천(18.75 g)을 4 L 플라스크에 섞고, 소용돌이치고, 오토클레이브를 20분 동안 (액체 사이클) 한다.
- 포도 주스 (250 mL)와 자당 (25 g)을 가열 된 플레이트 (저열) 위에 큰 교반 막대가있는 1 L 플라스크에 섞는다. 자당이 용해되면 열을 끄고 Tegosept (10 %, 4 mL) 및 프로피온산 (5 mL)을 첨가하기 전에 플라스크가 만질 때까지 기다리십시오. 교반 막대를 켜 두십시오.
- 오토클레이빙이 완료되면 플라스크가 닿을 때까지 식히고 (~ 60 °C) 포도 주스 믹스에 섞으십시오.
- 모든 용액을 하나의 플라스크에 결합하고 접시에 저어줍니다.
- 용액을 작은 크기의 페트리 접시 (35mm)의 뚜껑에 피펫하십시오. 뚜껑 당 약 9mL의 피펫 또는 볼록한 돔이 확보 될 때까지.
- 선택 사항 : 뚜껑에 불을 붙여서 거품을 제거하십시오.
- 플레이트가 응고되면 포도 플레이트를 밀폐 된 뚜껑이있는 상자에 넣고 상자를 4 °C에 놓습니다. 플레이트는 최대 1 개월 동안 보관할 수 있습니다.
- 비행 콘도 : 18G 바늘을 사용하여 6 온스 정사각형 바닥 폴리 프로필렌 초파리 병에 ~ 20 개의 구멍을 펀치하십시오.
- 성인 파리 (~ 100 OregonR 또는 모든 유전자형)를 플라이 콘도로 옮기고 효모 페이스트를 얹은 포도 한천 접시로 콘도를 덮으십시오. 접시의 중앙쪽으로 dab을 놓고 실험실 테이프로 플레이트를 콘도에 부착하십시오.
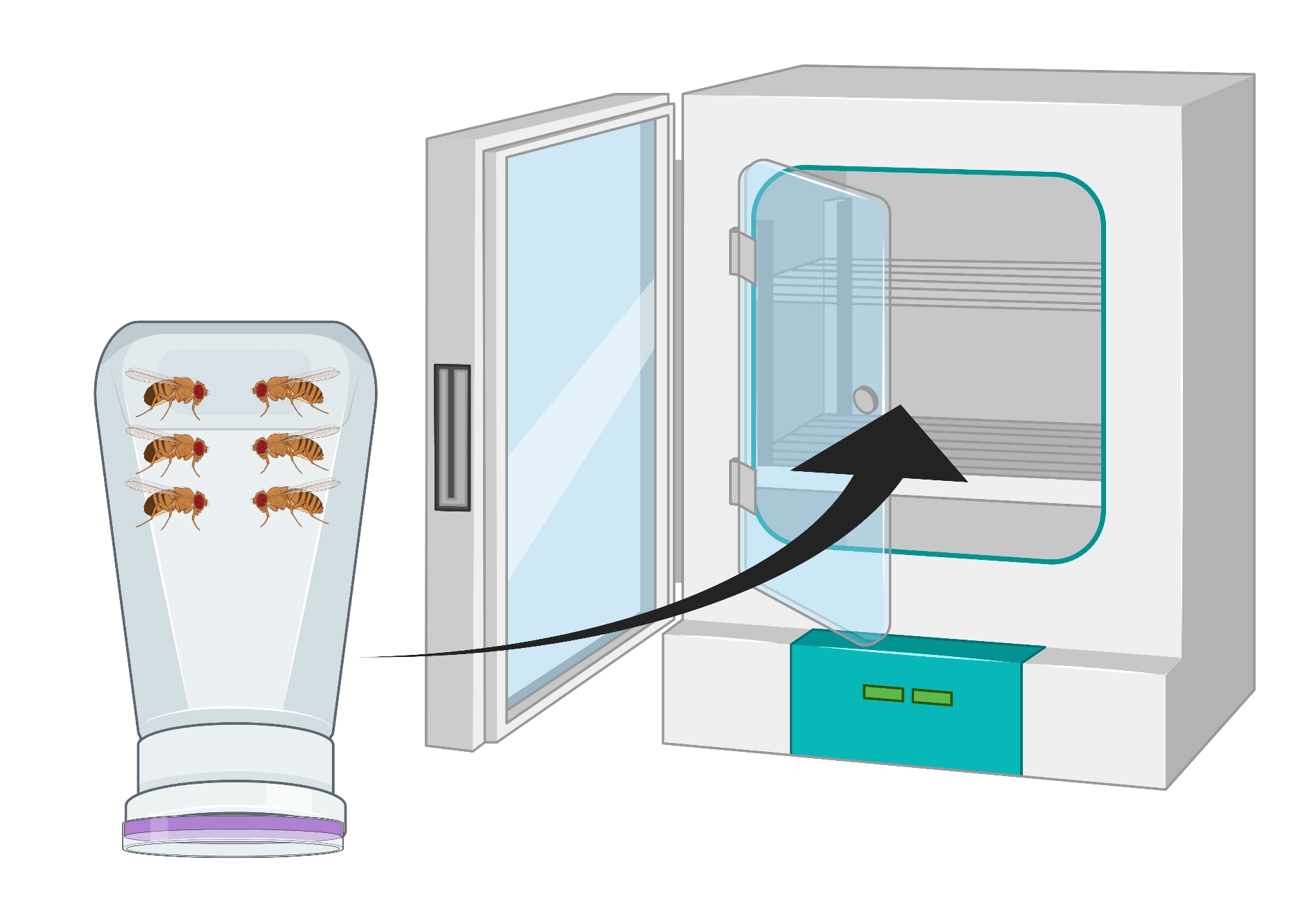
- 포도 한천 플레이트가 바닥에 오도록 용기를 반전시키고 25°C 인큐베이터에 24시간 동안 놓습니다(그림 2).

그림 2 : 남성과 여성 초파리 성인과 함께 거꾸로 된 파리 병 (콘도)의 시각적 표현. 플라스틱 병에는 산소 교환을 위해 18G 바늘로 생성 된 작은 구멍이 있습니다. 병의 입은 한천 포도 주스 뚜껑으로 밀봉되고 반전되어 25°C 인큐베이터에 저장된다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- 24 시간 후, 포도 한천 접시를 바꾸고 효모 페이스트로 얹은 새로운 접시로 교체하십시오. 성인 파리가 도망 가지 않도록 벤치에서 콘도를 가볍게 두드리면서 두 개의 접시를 빠르게 전환하십시오.
- 눈으로 플레이트를 검사하고 플레이트에있는 배아의 수를 평가하십시오. 초파리 배아는 타원형이고 흰색이며 두 개의 끈과 같은 부속기가 있습니다.
- 접시에 배아가 거의 없으면 (100 개 미만), 접시를 버리십시오 (한천을 파리 쓰레기통에 긁어 내고 재사용을 위해 플라스틱 뚜껑을 저장하십시오). 많은 경우에, 성인 암컷은 새로운 콘도에서 첫날 밤에 많은 배아를 낳지 않을 것입니다. 이 경우 성인 파리에게 적응하기 위해 또 다른 24 시간을주십시오.
- 플레이트에 많은 수의 배아가있는 경우 (적어도 100 개), 그것을 유지하고 평평한 바닥 주걱을 사용하여 효모 페이스트를 조심스럽게 제거하십시오.
- 효모 페이스트가 제거되면 금속 픽을 사용하여 해부 현미경으로 포도 판에서 모든 유충을 수동으로 제거하십시오. 해부 현미경으로 플레이트를 볼 때, 크롤링 애벌레는 배아뿐만 아니라 관찰되어야합니다.
- 한 유충의 측면을 향해 금속 픽을 닦아 모든 애벌레를 제거하십시오. 유충은 끈적 거리며 픽에 충실합니다. 일단 하나의 유충이 집어 들게되면, 더 많은 애벌레를 부착하기 위해 도구의 애벌레를 사용하여 추가 애벌레를 쉽게 집어 올릴 수 있습니다.
참고 : 애벌레는 서로 붙어 싶어합니다. 이 시점에서 애벌레가 손상되는지는 중요하지 않습니다. 이 애벌레는 버려질 것입니다. - 모든 유충을 채취 및 제거한 후, 플레이트를 다시 25°C 인큐베이터에 넣는다. 밀봉 할 수있는 더 큰 용기에 플레이트를 넣으십시오. 젖은 종이 타월을 큰 용기의 바닥에 두어 습기를 유지하십시오.
- 30-60 분 후에 플레이트를 해부 현미경으로 다시 가져 가서 동일한 포도 한천 플레이트에서 ~ 20-25 마리의 유충을 조심스럽게 선택하여 선택한 유충이 30-60 분 내에 신선하게 부화되도록하십시오.
- 20-25마리의 갓 부화한 유충과 함께 도구의 끝을 1-2mL의 1x 인산염 완충 식염수(PBS)로 채워진 페트리 접시(60mm)에 2분 동안 담그십시오.
- 2 분 후, 접시를 비스듬히 팁하여 바닥에 액체를 풀링하십시오. 작은 페인트 브러시를 사용하여 페트리 접시의 바닥 위로 액체에서 애벌레를 닦으십시오.
- 페인트 브러시에있는 모든 유충을 수집하고 유충을 70 % 에탄올 1-2 mL가 들어있는 새로운 페트리 접시 (60mm)로 옮깁니다. 1.15 단계를 반복하여 페인트 브러시로 유충을 모으고 1-2 mL의 1x PBS가있는 새로운 페트리 접시로 옮깁니다.
2. 문화 미디어 및 도구 준비
- 벤치와 작업 공간에 70 % 에탄올을 뿌리고 말리십시오.
- 해부 도구, 포셉 및 두 개의 유리 시계 접시에 70 % 에탄올을 뿌리고 벤치에서 말리십시오.
- 보충 된 슈나이더의 미디어 (SSM, 표 2)를 만들어 얼음 위에 놓습니다.
- SSM 1 mL를 각 유리 시계 접시에 피펫합니다.
- 멸균 팁이있는 마이크로 피펫을 사용하여 PBS 플레이트에서 갓 부화 한 유충을 첫 번째 유리 시계 접시의 SSM으로 옮깁니다. 멸균 팁이있는 마이크로 피펫을 사용하여 갓 부화 한 유충을 두 번째 유리 시계 접시의 SSM으로 옮깁니다.
3. 해부와 뇌 문화
- 애벌레가 SSM이있는 두 번째 유리 시계 접시에 들어가면 포셉과 해부 현미경을 사용하여 애벌레에서 뇌를 해부하십시오. 필요에 따라 배율을 조정합니다.
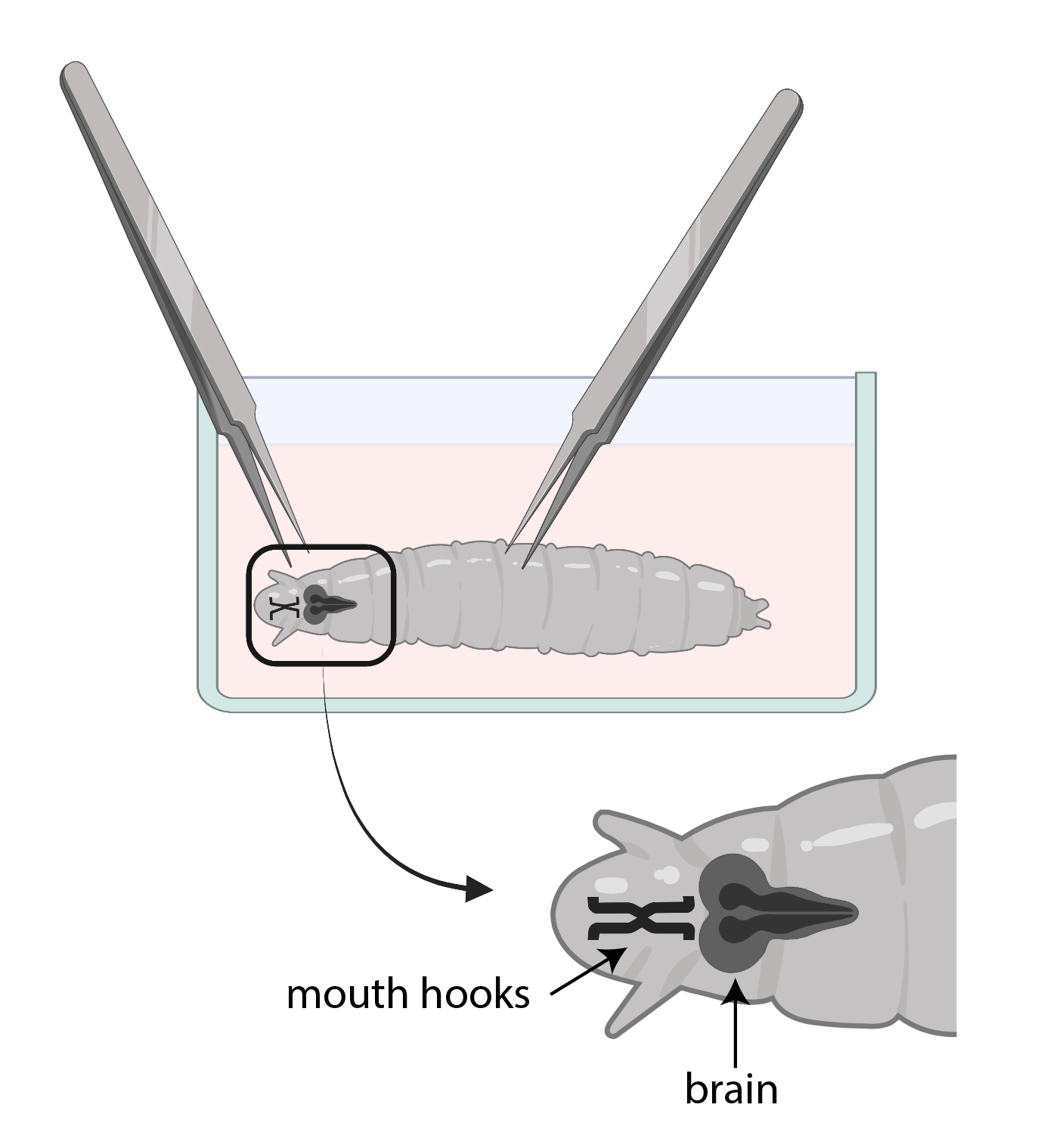
- 하나의 포셉을 사용하여 입 후크를 잡고 다른 포셉을 사용하여 몸을 반쯤 잡고 반대 방향으로 당기십시오 (그림 3).
참고 : 뇌는 입 후크 바로 뒤에 위치합니다. 뇌를 둘러싸고있는 다른 조직이있을 수 있습니다. 이러한 조직을 제거 할 때 뇌가 손상 될 수 있으므로 매우주의하십시오.

그림 3 : SSM이있는 유리 시계 접시에있는 초파리 유충. 포셉은 해부를 위해 적절하게 배치됩니다. 애벌레 두뇌 (진한 회색)의 위치는 입 후크 (검은 색)의 후방이며, 둘 다 애벌레 내부에 표시됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
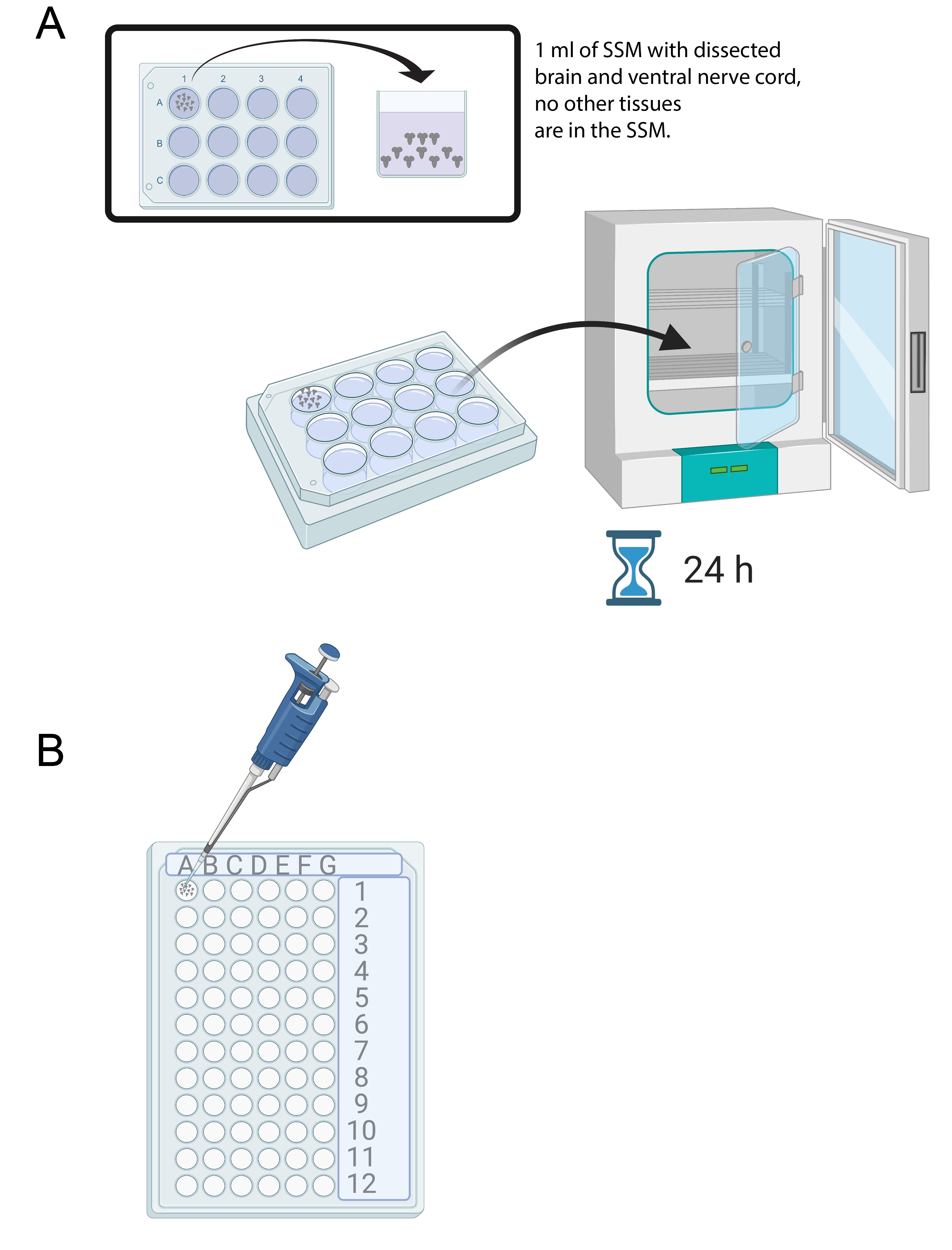
- 15-20개의 뇌가 해부되면 SSM 1mL를 멸균 12웰 배양 트레이의 한 웰에 넣습니다. 마이크로피펫과 멸균 팁을 사용하여 갓 해부된 뇌를 SSM으로 옮깁니다(그림 4A).
- 뇌를 12-웰 배양 트레이 내의 SSM 배지에 넣고 25°C에서 24시간 동안 인큐베이터에 넣는다(도 4A).

도 4: 뇌 배양 및 면역염색. (A) SSM 1mL를 함유하는 12웰 배양 접시에서 전체 뇌. 이어서, 배양 접시를 24시간 동안 25°C 인큐베이터에 넣고, (B) 면역염색 동안 뇌 외식편을 보유하는 72-웰 미니 트레이에 넣는다. 뇌는 세척되고 용액은 10 μL로 설정된 P20 마이크로피펫을 사용하여 이송된다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
4. 증식 분석, 뇌 고정 및 항체 염색
- 다음 날, EdU SSM 용액 1mL를 만드세요. 990 μL의 SSM (최종 EdU 농도는 0.1 mM과 같음)과 함께 5-에티닐-2'-데옥시우리딘 (EdU)의 10 mM 스톡의 10 μL를 멸균 마이크로원심분리 튜브에 넣고 혼합한다. 24시간 인큐베이션이 완료된 후, 1 mL의 EdU SSM을 멸균된 12-웰 배양 트레이의 하나의 웰에 피펫팅한다.
- 멸균 팁이 있는 마이크로피펫을 사용하여 뇌를 SSM을 포함하는 웰에서 EdU SSM 용액이 포함된 새로운 웰로만 옮깁니다. 25°C에서 1시간 동안 인큐베이션한다.
- 다음에, EdU 표지된 뇌를 1 mL의 고정제(4% 파라포름알데히드, 레시피에 대해서는 표 3 참조)를 함유하는 동일한 배양 트레이 내의 다른 웰로 20분 동안 옮긴다.
주의: 파라포름알데히드는 생물학적 위험이므로 적절하게 폐기해야 합니다. - 고정 후 마이크로 피펫을 사용하여 뇌를 72 웰 미니 트레이의 웰로 신속하게 옮깁니다. 각 웰은 10개의 뇌와 10-15μL의 액체를 보유할 수 있다(그림 4B). 뇌가 미니 트레이 (웰 당 10 개 이하의 뇌)로 옮겨지면 수정을 제거하고 1x PBT (인산염 완충액, 0.1 % Triton-X 100을 함유 한 pH 7.4)의 10 μL에서 뇌를 3 번 헹구십시오.
참고: 헹굼은 뇌에 1x PBT 10μL를 피펫팅하고 제거하고 3회 반복하는 것을 의미합니다. - 다음으로, 뇌를 각각 10분 동안 3회 세척하고, 다시 10 μL의 1x PBT로 세척한다. 뇌가 항상 액체로 덮여 있는지 확인하십시오.
- 세척이 완료된 후, 블로킹 용액 10 μL(10% 정상 염소 혈청을 갖는 1x PBT)를 뇌에 피펫팅한다. 트레이를 덮고 가장자리 주위에 파라필름 스트립을 사용하여 밀봉하십시오.
- 밀봉이 끝나면 미니 트레이를 젖은 수건으로 밀봉 된 상자에 넣어 습기가 많은 환경을 조성하여 증발을 방지하십시오. 트레이가 들어있는 박스를 하룻밤 사이에 4°C에 놓는다.
- 다음날, 일차 항체 용액을 만드십시오.
참고 :이 프로토콜에서 토끼 항 낙서는 세포막을 라벨링하고 쥐 항 데드 팬을 사용하여 신경 세포를 표지했지만 다른 많은 일차 항체를 사용할 수 있습니다.- 일차 항체 용액을 만들기 위해, 먼저, 차단 용액에서 일차 항체의 희석물을 만든다. 예를 들어, 토끼 낙서 방지는 1:1000의 최종 농도로 사용됩니다. 따라서, 먼저, 토끼 항-낙서 항체를 1:100에 희석한다 (항체 1 μL와 블로킹 용액 99 μL). 쥐 데드팬은 1:100의 최종 농도로 사용됩니다. 따라서, 먼저, 래트-데드팬 항체를 1:10에 희석한다 (항체 1 μL와 블로킹 용액 9 μL).
참고: 이러한 희석액은 박테리아 성장을 억제하기 위해 아지드 나트륨(0.05%)이 첨가되는 경우에도 4°C에서 장기간 보관할 수 있습니다. - 다음으로, 뇌를 포함하는 우물의 수를 계산하십시오. 웰의 수는 만들 일차 항체 용액의 부피를 결정한다. 예를 들어, 뇌의 웰이 2개 있는 경우, 20μL의 일차 항체 용액(10웰, 100μL 등)을 준비한다. 20 μL 일차 항체 용액을 만들기 위해, 각 일차 항체 희석액 2 μL와 블로킹 용액 16 μL를 첨가한다.
참고: 각 1차 항체의 최종 농도는 각각 1:1000 및 1:100이다. 요컨대, 일차 항체의 첫 번째 희석을 농도로 만들어 두 번째 희석이 항상 1:10이 되도록 하여 각각의 최종 농도에 도달하게 한다. 이 경우 토끼 낙서 방지의 경우 1:1000, 쥐 안티 데드 팬의 경우 1:100입니다.
- 일차 항체 용액을 만들기 위해, 먼저, 차단 용액에서 일차 항체의 희석물을 만든다. 예를 들어, 토끼 낙서 방지는 1:1000의 최종 농도로 사용됩니다. 따라서, 먼저, 토끼 항-낙서 항체를 1:100에 희석한다 (항체 1 μL와 블로킹 용액 99 μL). 쥐 데드팬은 1:100의 최종 농도로 사용됩니다. 따라서, 먼저, 래트-데드팬 항체를 1:10에 희석한다 (항체 1 μL와 블로킹 용액 9 μL).
- 블로킹 용액을 10 μL로 설정된 마이크로피펫으로 분리하고 10 μL의 1차 항체 용액을 각 웰에 피펫한다.
- 파라필름을 사용하여 트레이를 덮고 밀봉한 후 젖은 수건으로 밀봉된 상자에 다시 넣습니다. 4°C에서 하룻밤 동안 인큐베이션한다.
참고: 흔들림은 필요하지 않으며 강력하게 권장하지 않습니다. 항체는 흔들리거나 섞이지 않고 뇌에 침투합니다. - 다음날, 마이크로피펫을 이용하여 일차 항체 용액을 제거하고 1x PBT 10μL로 뇌를 3회 헹구었다. 다음으로, 뇌를 각각 10분 동안 10 μL의 1x PBT로 4회 세척한다. 세척 10분 동안, 이차 항체 용액을 준비한다.
- 이차 항체 용액을 만들려면 일차 항체를 인식하는 이차 항체를 선택하십시오. 이 프로토콜에서, 염소 항-토끼 알렉사 플루오르 488 및 염소 항-래트 알렉사 555가 사용되었다.
- 각각의 이차 항체 1 μL를 블로킹 용액 298 μL와 함께 마이크로원심분리 튜브에 넣고 각 이차 항체에 대해 최종 농도 1:300을 만든다.
- 마지막 10분 세척 후, 1x PBT 및 10 μL의 이차 항체 용액을 각 웰에 넣고 피펫을 제거하였다. 파라 필름을 사용하여 트레이를 밀봉하고 촉촉한 수건으로 상자에 다시 넣으십시오. 4°C에서 하룻밤 동안 인큐베이션한다.
참고: 헹굼, 세척 사이 또는 일차 및 이차 항체 용액을 추가할 때 웰의 마지막 μL를 모두 제거하는 것에 대해 걱정하지 마십시오. 두뇌는 항상 몇 μL의 액체에 잠겨 있습니다. - 다음날, 마이크로피펫을 이용하여 이차 항체 용액을 제거하고 각 1x PBT 10 μL로 뇌를 3회 헹구었다. 다음으로, 뇌를 각각 10분 동안 1x PBT를 각각 10 μL씩 10 μL로 4회 세척한다.
- 10분 세척 동안, EdU 혼입을 검출하기 위해 EdU 반응 믹스를 준비한다. 제조업체의 지침에 따라 EdU 반응 믹스를 준비하십시오.
- 최종 세척 후, 1x PBT 및 10 μL의 EdU 반응 믹스를 뇌로 각 웰에 피펫을 제거하였다. 마이크로웰 플레이트를 파라필름으로 밀봉하고 알루미늄 호일로 덮으십시오. 플레이트를 벤치에 30 분 동안 두십시오.
- 30분 후, 뇌를 각각 1x PBT 10 μL로 3회 헹구고 각각 5분 동안 1x PBT를 각각 10 μL로 3회 뇌를 세척한다.
- 마지막 세척 후, 글리세롤계 마운팅 매체 용액의 1x PBT 및 피펫 10 μL를 제거하였다. 플레이트를 밀봉하고 하룻밤 사이에 4°C에서 놓는다.
5. 두뇌 장착 및 이미징
- 다음 날, 현미경 슬라이드(25 mm x 75 mm x 1 mm)를 준비한다: 현미경 슬라이드의 양쪽 끝에 22 mm x 22 mm x 1 mm 정사각형 커버 글라스를 접착제(예: 수퍼글루)하여 슬라이드와 더 큰 커버슬립 사이의 공간을 만들기 위해 더 큰 22 mm x 50 mm x 1 mm 커버슬립을 배치하는 '브릿지'를 만듭니다(그림 5A). 이 공간은 뇌가 부서지는 것을 방지하면서 정확하게 지향 될 수있는 충분한 움직임을 허용합니다.

그림 5: 애벌레 뇌의 현미경 슬라이드, 방향 및 세포 유형을 보여주는 개략도. (A) 애벌레 뇌가 장착되고 이미징 준비가 된 현미경 슬라이드의 시각적 표현. (B) 조직 배향에 사용하기 위한 가이드라인도 도시되어 있다. (C) 공초점 현미경으로 이미징 할 준비가 된 현미경 슬라이드. (D) 애벌레 뇌의 세포 유형 중 일부를 보여주는 만화. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- 22 mm x 22 mm x 1 mm 커버 글라스를 현미경 슬라이드에 접착시킨 후, 마이크로웰 플레이트의 웰로부터 하나의 뇌를 함유하는 글리세롤계 장착 배지 용액 9.3 μL를 피펫팅하고 이를 슬라이드의 중앙에 놓는다(도 5A).
참고 : 애벌레 두뇌는 피펫 팁에 부착 될 수 있으므로 조심하십시오. 유착을 피하려면 먼저 안티페이드를 흡인 한 다음 9.3 μL 부피의 끝쪽으로 단일 뇌를 흡인하십시오. - 뇌가 슬라이드에 있으면 22mm x 50mm x 1mm 커버슬립을 위에 부드럽게 놓습니다. 도 5B에서 볼 수 있는 바와 같이 뇌를 위치시킨다. 커버슬립을 부드럽게 움직여 뇌의 방향을 잡습니다. 그런 다음 샘플을 이미징할 준비가 됩니다.
- 높은 배율과 높은 수치 조리개 대물(그림 5C)이 장착된 공초점 현미경을 사용하여 최상의 이미지를 얻을 수 있습니다. 예: 60x 또는 63x, 1.4 NA 오일 침지 렌즈.
- 커버 슬립 (및 목표)에 가장 가까운 등쪽 표면으로 두뇌를 이미지화하십시오. 1 μm 간격 또는 Z 단계 크기로 복부 표면(목표에서 가장 멀리 떨어져 있음)에서 시작하여 전체 뇌 반구를 통해 Z 스택을 획득합니다.
참고: 사용되는 레이저는 보조 항체에 따라 다릅니다. 이 프로토콜에서 사용된 레이저 라인은 낙서 염색을 검출하는 데 488nm, 데드팬을 검출하는 데 555nm, EdU를 검출하는 데 633nm였다.
- 커버 슬립 (및 목표)에 가장 가까운 등쪽 표면으로 두뇌를 이미지화하십시오. 1 μm 간격 또는 Z 단계 크기로 복부 표면(목표에서 가장 멀리 떨어져 있음)에서 시작하여 전체 뇌 반구를 통해 Z 스택을 획득합니다.
6. 데이터 분석
- 피지 오픈 소스 소프트웨어를 사용하여 뇌 반구를 분석하고 피지 세포 카운터 플러그인을 사용하여 세포를 계산하십시오.
결과
갓 부화한 오레곤R 야생형 뇌를 해부하고 인슐린으로 보충된 슈나이더 배지(SSM)에서 24시간 동안 배양하였다. Tissues는 프로토콜에 따라 고정되고 염색되었다. NBs를 검출하기 위해 데드판(Dpn)에 대해 생성된 일차 항체와 세포막을 표지하기 위한 낙서가 사용되었다. 티미딘 유사체 5-에티닐-2'-데옥시우리딘(Edu)을 첨가하여 S상 진입 및 NB 재활성화를 검출하였다. 우리는 배양물에서 24 시간 후에 큰 크?...
토론
뇌 외식편을 배양하기 위해 여기에 기술된 방법은 대부분의 실험실 환경에서 수행될 수 있다. 필요한 도구와 절차 및 데이터 수집은 간단하고 간단합니다. 이 방법을 이용하면, NB 재활성화 및 증식을 조절하는 세포 신호전달 캐스케이드 및 외인성 인자와 관련된 것들을 포함하는 다양한 가설을 시험할 수 있다. 여기에서, 야생형 OregonR 동물을 사용하여, 우리는 외인성 인슐린이 다른 동물 특이적 ?...
공개
저자는 경쟁 관심사가 없습니다.
감사의 말
우리는 LSAMP Bridges to Doctorate 프로그램 (CNK)과 NIH / NIGMS (R01-GM120421 및 R35-GM141886)를 인정합니다. 그림 1에 대해 Conor Sipe 박사님께 감사드립니다. 우리는 또한 모든 Siegrist 실험실 회원들에게 지속적인 지원과 멘토링에 감사드립니다. 우리는 특히 Chhavi Sood와 Gary Teeters에게 원고를주의 깊게 읽고 의견을 제공 해 주신 것에 감사드립니다.
자료
| Name | Company | Catalog Number | Comments |
| 10 µL Pipette tips | Denville Sci | P2102 | |
| 1000 µL Pipette tips | Denville Sci | P2103-N | |
| 1000 µL Pipettor | Gilson | P1000 | |
| 16% paraformaldehyde (10 x 10 mL) | Electron Microscopy Sciences | 2912.60.0000 | Used for Fixation of Larval Brains |
| 20 µL Pipette | Gilson | P20 | |
| 200 µL Pipette tips | Denville Sci | 1158U56 | |
| 24-well multiwell culture plates | Fisher Scientific | 50-197-4477 | |
| 35 mm Petri dishes | Fisher Scientific | 08-757-100A | Grape Plate Ingredients |
| 4 °C refrigerator | Fisher Scientific | Provides an ideal temperature for >24 h incubations in antibody solution | |
| 63x Objective | Lecia | ||
| Active dry yeast | Most supermarkets | ||
| Agarose | Fisher Scientific | 214010 | Grape Plate Ingredients |
| Click-iT EdU Cell Proliferation Kit for Imaging, Alexa Fluor 647 dye | Thermo Fisher Scientific | C10340 | to label proliferating cells |
| Confocal Microscope | Leica | SP8 | |
| Coverslips 22 mm x 22 mm x 1 mm , 10 pack of 4 oz | Fisher Scientific | 12-544-10 | Two Coverslips are super glued to the ends of the microscope slide. This creates a space that allows for the brains to float in antifade while being imaged. |
| Coverslips, 22 mm x 50 mm x 1 mm | Fisher Scientific | 12-545E | The coverslip is placed on two square coverslips on the microscope slide ensuring that the brain in the antifade does not move while imaging. |
| Dissecting microscope | Zeiss | Stemi 2000 | |
| Ethanol 200 proof (100%), Decon Labs, 1 gallon bottle | Fisher Scientific | 2701 | Used to wash off the larvae before the 24 hr hold in culture medium |
| Fetal Bovine Serum (10%) | Sigma | F4135-100ML | Supplement for cell culture media. |
| Fine forceps for dissection | Fine Science Tools | 11295-20 | Forcepts used in disections. They work best when sharpened. |
| Fly Bottles for Crossing | Genessee Scientific | 32-130 | This bottle is used as a container that lets the flies lay eggs on the grape plate. |
| Glass Dissection Dish (3 well) | These are no longer available | ||
| Glutathione | Sigma | G6013 | Provides oxidative protection during cell culture. |
| Goat Serum | Sigma | G9023- 10ML | Blocking Agent |
| Grape Plates | Made in house | Made in house | Grape juice/agarose plates for collecting freshly hatched eggs |
| Image J | Imagej.net/fiji/downloads | Free Download: https://fiji.sc | Imaging platform that is used to count cells and Edu reactivation |
| Incubator | Thermo Fisher Scientific | Ensures that the temperature, humidity, and light exposure is exactly the same throughout experiment. | |
| Insulin | Sigma | I0516 | Independant variable of the experiment |
| Laminar flow hood | For aliquoting culture media | ||
| L-Glutamine | Sigma | G7513 | Provides support during cell culture |
| Nunc 72-well Microwell Mini Trays | Fisher Scientific | 12-565-154 | Immunostaining steps are performed in this tray |
| Parafilm | Fisher Scientific | S37440 | Film used to seal plates in order to prevent evaporation |
| Pen-Strep | Sigma | P4458-100ml | Antibiodics used to prevent bacterial contamination of cells during culture. |
| Phosphate Buffer, pH7.4 | Made in house | Made in house | Solvent used to wash the brains after fixing and staining steps |
| Pick | Fine Science Tools | 10140-01 | Used to pick larvae off of the grape plate |
| Propionic acid | Fisher Scientific | A-258 | Grape Plate Ingredients |
| Rabbit 405 | Abcam | ab175653 | Antibodies used for immunostaining |
| Rat 555 | Abcam | ab150166 | Antibodies used for immunostaining |
| Rb Scribble | A Gift from Chris Doe | Antibodies used for immunostaining | |
| Rt Deadpan | Abcam | ab195173 | Antibodies used for immunostaining |
| Schneiders Culture Medium | Life Tech | 21720024 | Contains nutrients that help the cells grow and proliferate |
| SlowFade Diamond Antifade (5 x 2 mL) | Life Tech | S36963 | Reagent that provides protection against fading fluorophores |
| Sterile Water | Autoclave Milli-Q water made in house | Needed for Solutions | |
| Sucrose | Fisher | S2-12 | Grape Plate Ingredients |
| Superfrost Microscope Slides | Fisher Scientific | 12-544-7 | |
| Superglue | Most supermarkets | ||
| Tegosept | Genesee Scientific | 20-259 | Grape Plate Ingredients |
| Triton-X 100 | Sigma | T9284-100ML | PBT |
| Welch's 100% grape grape juice | Most supermarkets | Grape Plate Ingredients |
참고문헌
- Suman, S., Domingues, A., Ratajczak, J., Ratajczak, M. Z. Potential clinical applications of stem cells in regenerative medicine. Advances in Experimental Medicine and Biology. 1201, 1-22 (2019).
- Tabar, V., Studer, L. Pluripotent stem cells in regenerative medicine: challenges and recent progress. Nature Reviews Genetics. 15, 82-92 (2014).
- Daley, G. Q. Stem cells and the evolving notion of cellular identity. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370, 20140376 (2015).
- Rodrigues, M., Kosaric, N., Bonham, C. A., Gurtner, G. C. Wound healing: A cellular perspective. Physiological Reviews. 99, 665-706 (2019).
- van Velthoven, C. T. J., Rando, T. A. Stem cell quiescence: Dynamism, restraint, and cellular idling. Cell Stem Cell. 24, 213-225 (2019).
- Chapman, N. M., Boothby, M. R., Chi, H. Metabolic coordination of T cell quiescence and activation. Nature Reviews Immunology. 20, 55-70 (2020).
- Wosczyna, M. N., Rando, T. A. A muscle stem cell support group: Coordinated cellular responses in muscle regeneration. Developmental Cell. 46, 135-143 (2018).
- Homem, C. C., Knoblich, J. A. Drosophila neuroblasts: a model for stem cell biology. Development. 139, 4297-4310 (2012).
- Kang, K. H., Reichert, H. Control of neural stem cell self-renewal and differentiation in Drosophila. Cell and Tissue Research. 359, 33-45 (2015).
- Chell, J. M., Brand, A. H. Nutrition-responsive glia control exit of neural stem cells from quiescence. Cell. 143, 1161-1173 (2010).
- Sousa-Nunes, R., Yee, L. L., Gould, A. P. Fat cells reactivate quiescent neuroblasts via TOR and glial insulin relays in Drosophila. Nature. 471, 508-512 (2011).
- Britton, J. S., Edgar, B. A. Environmental control of the cell cycle in Drosophila: nutrition activates mitotic and endoreplicative cells by distinct mechanisms. Development. 125, 2149-2158 (1998).
- Lin, S., et al. Extremes of lineage plasticity in the Drosophila brain. Current biology : CB. 23, 1908-1913 (2013).
- Sipe, C. W., Siegrist, S. E. Eyeless uncouples mushroom body neuroblast proliferation from dietary amino acids in Drosophila. Elife. 6, 26343 (2017).
- Speder, P., Brand, A. H. Systemic and local cues drive neural stem cell niche remodelling during neurogenesis in Drosophila. Elife. 7, 30413 (2018).
- Yuan, X., Sipe, C. W., Suzawa, M., Bland, M. L., Siegrist, S. E. Dilp-2-mediated PI3-kinase activation coordinates reactivation of quiescent neuroblasts with growth of their glial stem cell niche. PLoS Biology. 18, 3000721 (2020).
- Colombani, J., et al. A nutrient sensor mechanism controls Drosophila growth. Cell. 114, 739-749 (2003).
- Geminard, C., Rulifson, E. J., Leopold, P. Remote control of insulin secretion by fat cells in Drosophila. Cell Metabolism. 10, 199-207 (2009).
- Siller, K. H., Serr, M., Steward, R., Hays, T. S., Doe, C. Q. Live imaging of Drosophila brain neuroblasts reveals a role for Lis1/dynactin in spindle assembly and mitotic checkpoint control. Molecular Biology of the Cell. 16, 5127-5140 (2005).
- Prithviraj, R., Trunova, S., Giniger, E. Ex vivo culturing of whole, developing Drosophila brains. Journal of Visualized Experiments: JoVE. (65), e4270 (2012).
- Bostock, M. P., et al. An immobilization technique for long-term time-lapse imaging of explanted drosophila tissues. Frontiers in Cell and Developmental Biology. 8, 590094 (2020).
- Datta, S. Activation of neuroblast proliferation in explant culture of the Drosophila larval CNS. Brain Research. 818, 77-83 (1999).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유