
Method Article
Functional Characterization and Visualization of Espacageal Fibroblasts using organoid co-cultures (오가노이드 공동 배양을 이용한 식도 섬유아세포의 기능적 특성 분석 및 시각화)
요약
오가노이드-섬유아세포 공동배양은 생체 내 줄기세포 틈새를 연구하기 위한 모델을 제공한다. 여기서, 식도 오가노이드-섬유아세포 공동배양을 위한 프로토콜이 기재되어 있다. 또한 전체 마운트 이미징을 사용하여 섬유아세포-오가노이드 상호작용을 시각화합니다.
초록
상피 줄기 세포와 전구 세포는 평생 동안 상피 장벽의 형성과 유지에 기여합니다. 대부분의 줄기 및 전구 세포 집단은 해부학적으로 구별되는 위치에 숨겨져 있어 줄기를 유지하는 틈새 신호와 독점적인 상호 작용을 가능하게 합니다. 상피 오가노이드 배양의 발달은 항상성 및 질병에서 줄기 및 전구 세포의 역할을 이해하기 위한 강력한 도구를 제공하지만, 틈새 환경 내에서의 상호 작용은 거의 부재하여 줄기 세포 행동에 영향을 미치는 요인의 식별을 방해합니다. 섬유아세포는 상피줄기와 전구세포의 운명을 지시하는 데 중요한 역할을 합니다. 여기에서는 식도 전구 세포 재생 및 분화에서 섬유아세포 하위 집단의 묘사를 가능하게 하는 포괄적인 오가노이드-섬유아세포 공동 배양 프로토콜이 제시됩니다. 이 프로토콜에서, 상피 세포와 섬유아세포 모두를 식도로부터 병렬로 분리하는 방법이 기재되어 있다. 형질전환 리포터 또는 야생형 마우스에서 식도 전구 세포와 섬유아세포 하위 집단을 모두 분리하기 위한 뚜렷한 형광 활성화 세포 분류 전략이 설명되어 있습니다. 이 프로토콜은 특정 섬유아세포 하위 집단의 분리를 수용하도록 조정할 수 있는 다양한 접근 방식을 제공합니다. 식도 상피 오가노이드 단일 배양의 확립 및 계대 배양이 이 프로토콜에 포함되어 있어 공동 배양 시스템과 직접 비교할 수 있습니다. 또한, 상피-섬유아세포 상호작용의 상세한 이미지 분석을 가능하게 하는 3D 클리어링 접근법이 설명된다. 총체적으로, 이 프로토콜은 시험관 내에서 식도 줄기 세포 틈새 성분을 식별하고 이해하기 위한 비교 및 상대적으로 높은 처리량 방법을 설명합니다.
서문
오가노이드는 줄기 세포와 전구 세포를 특성화하고 줄기 세포 틈새 1,2,3,4의 세포 구성 요소에서 파생된 신호 신호를 이해하기 위한 3D in vitro 분석으로 사용됩니다. 마우스 식도 오가노이드는 2014년에 처음 기술되었으며 여러 논문에서 R-Spondin(RSPO), NOGGIN 및 표피 성장 인자(EGF)와 같은 성장 인자를 확인하고 식도 오가노이드 5,6,7을 유지하고 통과시키는 데 필요한 성장 인자를 확인했으며 생체 내에서 유사한 신호 신호가 필요하다고 주장했습니다 전구 세포 재생. 그러나 성장 인자는 일반적으로 비생리학적 농도로 첨가되어 생체 내 신호 전달 환경을 반드시 반영하지 않는 오가노이드 성장 조건을 초래합니다.
섬유아세포(Fibroblasts)는 많은 줄기세포 틈새에서 전구세포 특성을 지지하는 이질적인 기질 세포 집단이다8. 동일한 오가노이드 배양에서 상피 전구 세포와 섬유아세포를 결합하면 외인성으로 보충된 성장 인자의 감소된 농도에서 오가노이드 형성이 가능합니다. 장 및 간 상피로부터의 오가노이드 공동배양 시스템이 기재되어 있지만, 식도 오가노이드-섬유아세포 공동배양을 확립하기 위한 프로토콜은 여전히 탁월하다 9,10,11.
이 프로토콜에서는 형질전환 PdgfrαH2BeGFP 마우스12 또는 고전적인 항체 염색이 있는 야생형 마우스를 사용하여 식도에서 섬유아세포에 대한 두 가지 형광 활성화 세포 분류(FACS) 전략이 요약됩니다. 섬유아세포의 상이한 하위집단은 선택된 세포 표면 마커를 사용하여 분리될 수 있으며, 이에 따라 프로토콜에 유연성을 제공한다. 또한, 오가노이드 형태를 보존하는 3D 형광 이미징 기술을 사용하여 섬유아세포-오가노이드 상호작용을 특성화합니다. 오가노이드 투명화는 오가노이드의 광 투과 깊이를 증가시켜 오가노이드-섬유아세포 연결의 시각화를 개선하고 오가노이드 구조 전체를 재현할 수 있는 빠른 방법을 제공합니다. 이 프로토콜은 식도 오가노이드 공동 배양과 전체 마운트 이미징 전략을 결합하여 섬유아세포-오가노이드 상호작용의 기능적 특성화를 가능하게 합니다.
프로토콜
이 연구를 위한 동물 실험은 스톡홀름 Norra djurförsöksetiska nämnd(윤리 허가 번호 14051-2019)의 승인을 받았습니다. 동물들은 유럽 실험 동물 과학 협회 연맹 (Federation of European Laboratory Animal Science Association)의 권고에 따라 병원체가없는 상태로 사육되었다.
1. 준비
- 해리에 사용되는 효소 원액( 재료 표 참조)을 얼음 위에서 해동합니다. 성장인자 환원(GFR) 기저막 매트릭스(matrix)의 분취량을 4°C에서 해동한다.
- 배양 배지를 준비합니다. 오가노이드 배양 및 공동 배양을 위해 표 1 에 기재된 배지를 사용하고 프로토콜을 시작하기 전에 준비합니다.
2. 식도 상피와 간질의 해부 및 분리
알림: 해부 및 조직 처리에 사용되는 모든 기구가 멸균되었는지 확인하십시오. 3 식도 당 Hanks의 균형 소금 용액 (HBSS)에서 2 mL의 해리 용액 ( 재료 표 참조)을 준비합니다.
- C57BL/6J 마우스와 같은 마우스 균주를 선택하십시오. 생쥐의 식도 발달은 출생 후 (p) 70 이후에 끝나므로 p7013보다 오래된 생쥐를 사용하는 것이 좋습니다. 4-5마리의 마우스는 8-10개의 오가노이드 공동 배양을 확립하기에 충분한 재료를 제공하므로 사용하십시오.
참고: 오가노이드 형성 효율은 생쥐의 나이에 따라 감소합니다. 특정 섬유아세포 하위집단이 관심 대상인 경우, 낮은 섬유아세포 수율이 확립될 수 있는 오가노이드 공동배양의 수를 제한할 수 있다. - 섬유아세포 집단의 분리를 위해 유전자 변형(예: PdgfrαH2BeGFP) 또는 야생형(WT) 마우스를 사용합니다. WT 마우스를 사용할 때 항체 염색을 수행하여 FACS를 통해 간질에서 특정 섬유아세포 하위 집단을 분리합니다.
- 마우스를CO2 질식에 의해 안락사시킨다. 집게와 해부 가위를 사용하여 식도를 해부합니다. 식도 전체를 제거하려면 위 바로 위의 식도 말단과 기관 시작 부분의 근위 끝을 자릅니다. 식도를 PBS에 담그고 얼음 위에 올려 놓습니다.
- 해부 현미경(총 배율 범위 8x-40x)과 겸자를 사용하여 외근근을 기계적으로 제거합니다. 한 쌍의 집게를 사용하여 해부된 식도의 말단부를 잡고 다른 집게를 사용하여 해부된 식도의 원위부에서 근위부까지 근육을 잡고 당깁니다. 근육층을 제거하고 버립니다.
- 식도를 세로로 엽니다. 이것은 조직 손상을 방지하기 위해 볼 팁이 있는 미세 해부 스프링 가위를 사용하는 것이 가장 좋습니다. 식도의 한쪽 끝을 잡고 스프링 가위의 볼을 식도 내강에 삽입하고 끝을 잡고 식도를 잘라냅니다.
- 식도를 1.5mL 마이크로 원심분리 튜브 또는 24웰 플레이트에 넣습니다. 열린 식도를 HBSS의 0.5mg/mL thermolysin에서 37°C의 로커 셰이커에서 15분 동안 배양합니다. 식도를 thermolysin 용액에 완전히 담그십시오.
알림: 사용되는 Thermolysin 용액의 부피는 웰 또는 튜브 크기에 따라 다릅니다. 여러 식도를 같은 우물이나 튜브에 넣고 잠글 수 있습니다. - thermolysin 용액에서 식도를 꺼냅니다. 해부 현미경을 사용하여 식도 상피를 기질에서 조심스럽게 분리합니다. 가는 집게를 사용하여 조직의 상피 쪽과 기질 쪽을 모두 잡고 천천히 당겨 두 층을 분리합니다.
참고: 간질은 투명한 상피와 달리 흰색과 불투명한 모양으로 식별됩니다. 간질은 고유층과 점막하 층을 포함합니다. - 상피층과 기질층을 HBSS에서 200μL의 해리 용액과 함께 두 개의 분리된 1.5mL 미세 원심분리기 튜브로 옮깁니다. 얼음 위에 올려 놓으십시오.
3. 식도 전구 세포의 분리
참고: 식도 전구 세포(2단계)와 섬유아세포(3단계)의 분리를 동시에 수행할 수 있습니다. HBSS(1% FBS)에서 1% FBS의 50mL 튜브를 준비합니다.
- 분리된 식도 상피를 1.5mL 미세 원심분리 튜브(단계 2.8)에서 깨끗한 페트리 접시로 옮기고 날카로운 메스를 사용하여 다진다. 200μL의 해리 용액과 함께 페트리 접시에서 다진 조직을 수집하고 1.5mL 마이크로 원심분리기 튜브로 다시 옮깁니다.
참고: 200μL 피펫 팁을 사용하여 조각을 1.5mL 마이크로 원심분리기 튜브에 다시 넣을 수 있을 때 상피가 적절하게 다진 것입니다. - 800 μL의 신선한 해리 용액을 1.5 mL 마이크로 원심분리기 튜브에 넣어 총 부피 1 mL를 만듭니다.
알림: 충분한 양의 해리 용액을 추가하는 것이 중요합니다. 1-3 식도 당 1mL의 용액을 권장합니다. 한 번에 더 많은 식도가 처리되면 확장됩니다. - 다진 상피층이 있는 튜브를 37°C에서 60분 동안 로커-셰이커에 놓습니다. 소화를 향상시키기 위해 15분마다 200μL 피펫 팁을 사용하여 용액을 약 20회 위아래로 피펫팅합니다.
- 60분 후, 200 μL 피펫 팁을 사용하여 다시 20회 위아래로 피펫팅합니다. 용액을 40μm 셀 스트레이너를 통해 새로운 1.5mL 마이크로 원심분리기 튜브에 통과시킵니다. 4 °C에서 10분 동안 300 x g 로 원심분리합니다.
알림: 세포가 필터에 부착되는 것을 최소화하기 위해 세포를 변형시키기 전에 1% FBS로 세포 여과기를 적십니다. 상피는 완전히 소화되지 않으며 조직 조각은 여전히 볼 수 있습니다. 그러나 배양 기간이 길어지면 세포 생존율이 감소하고 생존 가능한 세포의 수율이 높아지지 않습니다. - 1mL 피펫으로 과량의 액체를 제거하여 상청액을 버리고 펠릿을 1% FBS 1mL에 재현탁합니다. 4 °C에서 10분 동안 300 x g 로 원심분리합니다.
- 원심분리 동안, 식도 전구 세포의 FACS에 대한 접합된 항체 혼합물을 준비한다.
- 100만 개의 세포당 1% FBS의 200μL에 1μL의 CD324-PE-Cy7(ECADHERIN) 및 CD104-A647(INTEGRIN-β4)을 혼합합니다.
참고: 1μL의 항체(최종 부피 200μL)는 1개 또는 2개의 식도에 충분합니다. 한 번에 더 많은 식도를 처리 할 때 항체 염색 용액의 부피를 늘리십시오.
- 펠릿을 200μL의 항체 혼합물에 재현탁하고 유세포 분석 튜브로 옮깁니다. 형광 항체를 추가한 후 신호 표백을 방지하기 위해 세포 현탁액을 어둠 속에 보관하십시오. 세포를 4°C에서 30분 동안 인큐베이션한다. 3mL의 1% FBS를 추가하고 4°C에서 10분 동안 300 x g 에서 원심분리한 다음 1% FBS의 최소 200μL에 세포를 재현탁합니다.
참고: 500 μL의 1% FBS는 최대 4개 또는 5개의 식도에 사용됩니다. 한 번에 더 많은 식도가 처리될 때 부피를 증가시켜 100-300 events/s의 FACS 유속을 달성할 수 있습니다. 이벤트/초가 많을수록 분류 효율이 감소하고 유속이 증가하면 세포 생존이 감소합니다. - 살아있는 세포를 분리하기 위해 FACS 분류 5분 전에 1:10,000의 최종 농도에 죽은 세포 염색 마커를 추가합니다. FACS 기계를 사용하여 전구 세포를 정렬합니다(게이팅 전략은 그림 1 참조). 200μL의 염기성 오가노이드 배지로 채워진 1.5mL 마이크로 원심분리 튜브에 세포를 수집합니다(표 1).
4. 기질층에서 섬유아세포 분리
- 해부 가위를 사용하여 200μL의 해리 용액이 들어 있는 1.5mL 튜브(2.8단계)에서 장루층을 미세한 조각으로 자릅니다. 200 μL 피펫 팁을 사용하여 용액을 위아래로 피펫팅할 수 있게 되면 조직을 적절하게 다집니다.
- 800 μL의 신선한 해리 용액을 1.5 mL 마이크로 원심분리기 튜브에 넣어 총 부피 1 mL를 만듭니다.
알림: 충분한 양의 해리 용액을 추가하는 것이 중요합니다. 1-3 식도 당 1mL의 용액을 권장합니다. 한 번에 더 많은 식도가 처리되면 확장됩니다. - 튜브를 37°C의 로커 셰이커에 30분 동안 놓습니다. 15분 후, 200μL 피펫 팁을 사용하여 용액을 약 20회 위아래로 피펫팅하여 소화를 개선합니다.
- 효소 분해 30분 후, 200 μL 피펫 팁을 사용하여 20회 더 위아래로 피펫팅합니다. 70μm 세포 여과기를 통해 용액을 새로운 1.5mL 마이크로 원심분리기 튜브에 걸러냅니다. 4 °C에서 10분 동안 300 x g 로 원심분리합니다.
알림: 세포가 필터에 부착되는 것을 최소화하기 위해 세포를 걸러내기 전에 1% FBS로 세포 여과기를 적십니다. - 1mL 피펫으로 과량의 액체를 제거하여 상청액을 버리고 펠릿을 1% FBS 1mL에 재현탁합니다. 4 °C에서 10분 동안 300 x g 로 원심분리합니다.
참고: 형광 표지 섬유아세포를 포함하는 유전자 변형 마우스 균주를 사용하는 경우 항체 염색은 선택 사항입니다. 항체 염색이 필요하지 않은 경우 3.7단계를 계속 진행하여 샘플을 유세포 분석 튜브로 옮깁니다.- 원심분리 동안, 섬유아세포의 FACS 분리를 위해 접합된 항체 혼합물을 준비한다. 100만 개의 세포당 1% FBS의 200μL에 1μL의 CD26-APC(DPP4)를 혼합합니다.
참고: 1μL의 항체는 1개 또는 2개의 식도에 충분합니다. 한 번에 더 많은 식도를 처리 할 때 항체 염색 용액의 부피를 늘리십시오.
- 원심분리 동안, 섬유아세포의 FACS 분리를 위해 접합된 항체 혼합물을 준비한다. 100만 개의 세포당 1% FBS의 200μL에 1μL의 CD26-APC(DPP4)를 혼합합니다.
- 1% FBS에 혼합된 접합 항체 혼합물 200μL에 펠릿을 재현탁하고 유세포 분석 튜브로 옮깁니다. 세포를 4°C에서 30분 동안 인큐베이션한다.
- 3mL의 1% FBS를 넣고 4°C에서 10분 동안 300 x g 로 원심분리합니다. FBS의 1%의 최소 200μL에 세포를 재현탁합니다.
참고: 500 μL의 1% FBS는 최대 4개 또는 5개의 식도에 사용됩니다. 한 번에 더 많은 식도를 처리할 때 부피를 증가시켜 100-300 events/s의 유속을 달성할 수 있습니다. 이벤트/초가 많을수록 분류 효율이 감소하고 유속이 증가하면 세포 생존이 감소합니다. 형광 항체를 첨가한 후 신호 표백을 방지하기 위해 세포 현탁액을 어둡게 유지해야 합니다. - 살아있는 세포를 분리하기 위해 FACS 분류 5분 전에 1:10,000의 최종 농도에 죽은 세포 염색 마커를 추가합니다. FACS 기계를 사용하여 셀을 정렬합니다(게이팅 전략은 그림 1 참조). 200μL의 염기성 오가노이드 배지로 채워진 1.5mL 마이크로 원심분리 튜브에 세포를 수집합니다(표 1).
5. 식도 오가노이드의 확립과 배양
참고: 예열ER저 (오가노이드 공동 배양), ENR(오가노이드) 배지(설명은 표 1 참조), 및 37°C에서 48-웰 플레이트. 해동된 매트릭스(1단계에서 준비됨) 분취액을 얼음 위에 놓습니다. 여기에 제공된 매트릭스( 재료 표 참조)를 마우스 식도 오가노이드 배양에 사용하는 것이 권장되는데, 이는 다른 브랜드의 매트릭스가 오가노이드 형성 효율에 부정적인 영향을 미치기 때문입니다.
- 오가노이드 공동 배양의 경우, 분류된 상피 세포와 섬유아세포를 튜브에서 1:2의 비율로 혼합합니다. 각 매트릭스 돔에 대해 5,000개의 상피 세포와 10,000개의 섬유아세포를 사용합니다. 더 많은 돔을 준비할 때 하나의 튜브에 5,000개의 상피 세포와 10,000개의 섬유아세포를 추가합니다. 섬유아세포가 없는 오가노이드 배양의 경우 매트릭스 돔당 5,000개의 상피 세포를 사용합니다.
- 혼합된 세포 집단을 300 x g 에서 5분 동안 원심분리합니다. 200 μL 피펫으로 상층액을 조심스럽게 제거하여 버립니다.
- 펠렛을 차가운 염기성 오가노이드 배지에 재현탁하고 300 x g 에서 5분 동안 원심분리하여 세포를 한 번 세척합니다. 세포를 얼음 위에 놓습니다.
- 80% 매트릭스와 20% 차가운 염기성 오가노이드 배지로 구성된 혼합물을 준비합니다. 매트릭스가 실온(RT)에서 굳어지면서 모든 것을 얼음 위에 놓습니다.
- 원심분리 후 상층액을 200 μL 피펫으로 조심스럽게 제거하여 버립니다. 10μL의 매트릭스 믹스/돔에 세포를 재현탁하고 얼음 위에 다시 놓습니다.
- 37°C 인큐베이터에서 예열된 48웰 플레이트를 가져와서 20μL 피펫을 사용하여 웰당 하나의 매트릭스 돔을 만듭니다. 각 돔에는 10μL의 80% 매트릭스, 5,000개의 상피 세포 및 10,000개의 섬유아세포가 포함되어 있습니다. 플레이트를 거꾸로 인큐베이터로 옮기고 돔을 37°C에서 20-30분 더 굳힙니다.
참고: 매트릭스 돔 부피의 감소 및/또는 세포 수의 증가는 오가노이드 형성 효율에 영향을 미칩니다. - 200μL의 예열된 ER저 배지(표 1)를 오가노이드 공동 배양이 포함된 매트릭스 돔에 추가하고 ENR 배지(표 1)를 상피 오가노이드만 포함된 각 매트릭스 돔에 추가하고 플레이트를 인큐베이터에 넣습니다.
- 오가노이드를 37°C 및 5%CO2에서 성장시킨다. 처음 2일 동안은 배지에 10μM Rock-inhibitor(Y-27623)를 보충합니다. 암석 억제제는 스트레스로 인한 세포 사멸을 방지하고 오가노이드 배양의 성공적인 확립 가능성을 높입니다.
- 2-3 일마다 매체를 새로 고칩니다. 온도에 민감한 매트릭스의 해리를 방지하기 위해 매체가 따뜻한지 확인하십시오.
- 도금 후 6-8일 후에 실험 분석을 수행한다. 오가노이드는 최대 14일 동안 배양할 수 있습니다. 14일째 경에, 돔 무결성의 손실이 관찰된다.
6. 오가노이드의 패시징
참고: 공동 배양에서 성장한 오가노이드의 계대배양은 섬유아세포의 손실을 초래합니다. 따라서 계대 시 모든 오가노이드에 대해 ENR 배지를 사용하는 것이 좋습니다. ENR, PBS, 및 48-웰 플레이트를 37°C에서 예열하였다.
- 배지를 제거하고, 매트릭스 돔을 함유하는 웰을 예열된 PBS로 세척한다. 차가운 0.25% 트립신 용액 200μL를 추가하고 위아래로 피펫을 넣어 매트릭스 돔을 부수십시오.
참고: 매트릭스가 온도에 민감하고 매트릭스 돔을 분해하는 데 도움이 되기 때문에 차가운 0.25% 트립신을 사용하는 것이 좋습니다. - 오가노이드를 트립신과 함께 37°C에서 ~20분 동안 배양합니다. 오가노이드의 해리를 증가시키기 위해 10분 후에 위아래로 피펫팅합니다. 5-10분마다 현미경으로 해리 과정을 모니터링합니다. 트립신은 세포 생존력을 감소시키기 때문에 이상적인 해리 시간을 식별하는 데 도움이 될 수 있습니다.
- 20분 후, 200μL 피펫으로 오가노이드를 위아래로 피펫팅하여 오가노이드를 해리합니다. 1.5mL 마이크로 원심분리 튜브에 세포를 모으고 1mL의 기본 오가노이드 배지를 추가하고 RT에서 300 x g 에서 5분 동안 원심분리합니다.
- 선택 사항: 최상의 오가노이드 성장 조건을 보장하고 새로운 오가노이드가 단일 세포에서 파생되도록 하려면 미리 습윤된 40μm 세포 여과기를 통해 세포를 변형시킵니다. 세포 현탁액을 여과하면 해리하기 어려운 오가노이드 코어와 세포 덩어리가 제거됩니다.
- 80% 매트릭스와 20% 저온 염기성 오가노이드 배지로 구성된 매트릭스 혼합물을 준비합니다. 매트릭스가 RT에서 굳어지면서 모든 것을 얼음 위에 놓습니다.
- 200μL 피펫으로 상층액을 조심스럽게 제거하여 버리고 10μL의 매트릭스 믹스/돔에 세포를 재현탁한 다음 혼합물을 다시 얼음에 놓습니다.
참고: 오가노이드는 돔 밀도에 따라 1:5에서 1:10 비율로 분할할 수 있습니다. 상피 세포는 또한 5,000 세포/돔에서 계수 및 재도금될 수 있습니다. - 37°C 인큐베이터에서 예열된 48웰 플레이트를 취하여 웰당 하나의 돔을 만듭니다. 플레이트를 거꾸로 인큐베이터로 옮기고 돔을 37°C에서 20-30분 더 굳힙니다.
- 예열된 ENR 배지 200μL를 각 오가노이드 돔에 추가합니다. 처음 2일 동안 배지에 10μM Rock-inhibitor(Y-27623)를 보충합니다.
- 2-3 일마다 매체를 새로 고칩니다. 온도에 민감한 매트릭스의 해리를 방지하기 위해 매체가 따뜻한지 확인하십시오.
7. 전체 마운트 염색을 위한 오가노이드 가공
알림: 오가노이드가 플라스틱에 달라붙지 않도록 사용하기 전에 팁과 튜브를 PBS에 10% FBS로 코팅하십시오. 피펫 팁의 경우 팁을 사용하기 전에 10% FBS/PBS 용액에서 위아래로 한 번 또는 두 번 피펫팅하는 것으로 충분합니다. 튜브의 경우 튜브에 10% FBS/PBS를 채운 다음 용액을 제거합니다.
- 오가노이드 배지를 제거하고 얼음처럼 차가운 PBS 200μL를 매트릭스 돔에 추가합니다. 접시를 얼음 위에 5-10분 동안 놓습니다.
- 위아래로 피펫팅하고 용액을 0.6mL 마이크로 원심분리기 비부착 튜브로 옮깁니다. 오가노이드가 바닥에 가라앉을 수 있도록 100 x g 에서 30-60초 동안 잠시 원심분리합니다.
참고: 과도한 피펫팅과 긴 원심분리는 오가노이드를 파괴하고 섬유아세포-오가노이드 상호작용을 방해합니다. - 과도한 액체를 제거하고 얼음처럼 차가운 PBS를 첨가하십시오. 오가노이드가 바닥에 가라앉을 수 있도록 100 x g 에서 30-60초 동안 잠시 원심분리합니다.
- 과도한 액체를 제거하고 얼음에서 30분 동안 PBS 용액에 차가운 4% 포름알데히드 200μL로 오가노이드를 고정합니다.
주의 : 포름알데히드는 독성이 있으므로 항상 화학 흄 후드에 사용해야 합니다. 니트릴 장갑, 보안경 및 실험복은 항상 착용해야 합니다. - 오가노이드를 고정하면 오가노이드가 가라앉고 더 이상 원심분리가 필요하지 않습니다. 튜브를 똑바로 세워 오가노이드가 가라앉도록 하고 2-3분 정도 기다렸다가 포름알데히드를 제거하고 차가운 PBS 500μL를 추가하여 포름알데히드를 씻어냅니다.
- 오가노이드를 가라앉히고 과도한 PBS를 제거한 다음 신선하고 차가운 PBS 500μL를 추가합니다. 오가노이드를 가라앉히고 과도한 PBS를 제거한 다음 500μL의 차단 완충액(PBS에서 5% 정상 당나귀 혈청, 1% BSA 및 0.5% Triton X-100)을 추가합니다. RT에서 60 분 동안 로커 셰이커에 튜브를 놓습니다.
알림: 차단은 4°C에서 하룻밤(O/N)으로 수행할 수도 있습니다. - 오가노이드를 가라앉히고 블로킹 버퍼를 제거한 다음 1차 항체가 있는 블로킹 버퍼 200μL에 오가노이드를 다시 현탁합니다( 재료 표 참조). 오가노이드를 4°C의 로커 셰이커 O/N에 보관합니다.
참고: 핵 단백질, 저발현 단백질 또는 비특이적 염색을 나타내는 항체의 염색을 개선하기 위해 4°C에서 1일 또는 2일 동안 배양 시간을 늘릴 수 있습니다. 오가노이드를 로커 셰이커에 올려놓는 것은 오가노이드가 뭉치는 것을 방지하고 염색 효율을 높이기 때문에 필수적입니다. - 오가노이드를 가라앉히고 1차 항체 혼합물을 제거한 다음 PBS(0.02% Tx) 중 0.02% Triton X-100 500μL를 사용하여 RT에서 60분 동안 오가노이드를 세척합니다.
참고: 더 긴 1차 항체 배양이 필요한 경우 4°C에서 PBS O/N에 0.02% Tx로 세척 단계를 추가합니다. - 오가노이드를 가라앉히고 세척 완충액을 제거한 다음 4°C의 블로킹 완충액 O/N에 형광 결합 2차 항체 200μL를 추가합니다. 형광 2차 항체를 추가한 후 신호의 표백을 방지하기 위해 오가노이드를 어두운 곳에 보관하십시오.
- 오가노이드를 가라앉히고 2차 항체 혼합물을 제거한 다음 RT에서 60분 동안 0.02% Tx 500μL를 사용하여 오가노이드를 세척합니다.
- 필요한 경우 핵 염색을 위해 200μL의 DAPI(0.25μg/mL) 용액으로 오가노이드를 반대 염색합니다. 로커-셰이커(rocker-shaker)의 RT에서 60분 동안 샘플을 배양합니다.
- 오가노이드를 가라앉히고 DAPI 용액을 제거하고 오가노이드를 500μL의 PBS로 RT에서 15분 동안 세척합니다.
- 오가노이드를 가라앉히고 과도한 액체를 모두 제거합니다. 오가노이드에 투명 용액 10μL를 추가하고 RT에서 15분 동안 배양합니다.
참고: 세척 용액은 점성이 있으므로 세척 용액을 피펫팅하기 전에 20μL 피펫 팁의 바깥쪽 부분을 잘라냅니다. 더 큰 스페이서가 이미징에 사용되는 경우 더 많은 투명 용액을 추가할 수 있습니다. 오가노이드를 투명화하지 않을 때 세척액 대신 마운팅 용액을 사용할 수 있습니다. 클리어링 솔루션은 이미징 시 배경을 줄이고 이미징 깊이를 증가시킵니다. - 현미경 슬라이드에 0.05mm 양면 끈적끈적한 4웰 스페이서를 놓습니다. 오가노이드와 함께 10μL의 투명 용액을 하나의 웰에 넣고 스페이서 위에 커버 슬립을 놓습니다.
알림: 오가노이드는 스페이서를 사용하지 않고도 장착할 수 있습니다. 스페이서는 오가노이드 모양을 그대로 유지하고 오가노이드가 평평해지는 것을 방지합니다. - 슬라이드를 RT O/N으로 유지하여 오가노이드를 제거합니다. 장기간 보관하려면 슬라이드를 4°C로 보관하십시오. 컨포칼 현미경 시스템을 사용하여 이미지를 획득합니다.
결과
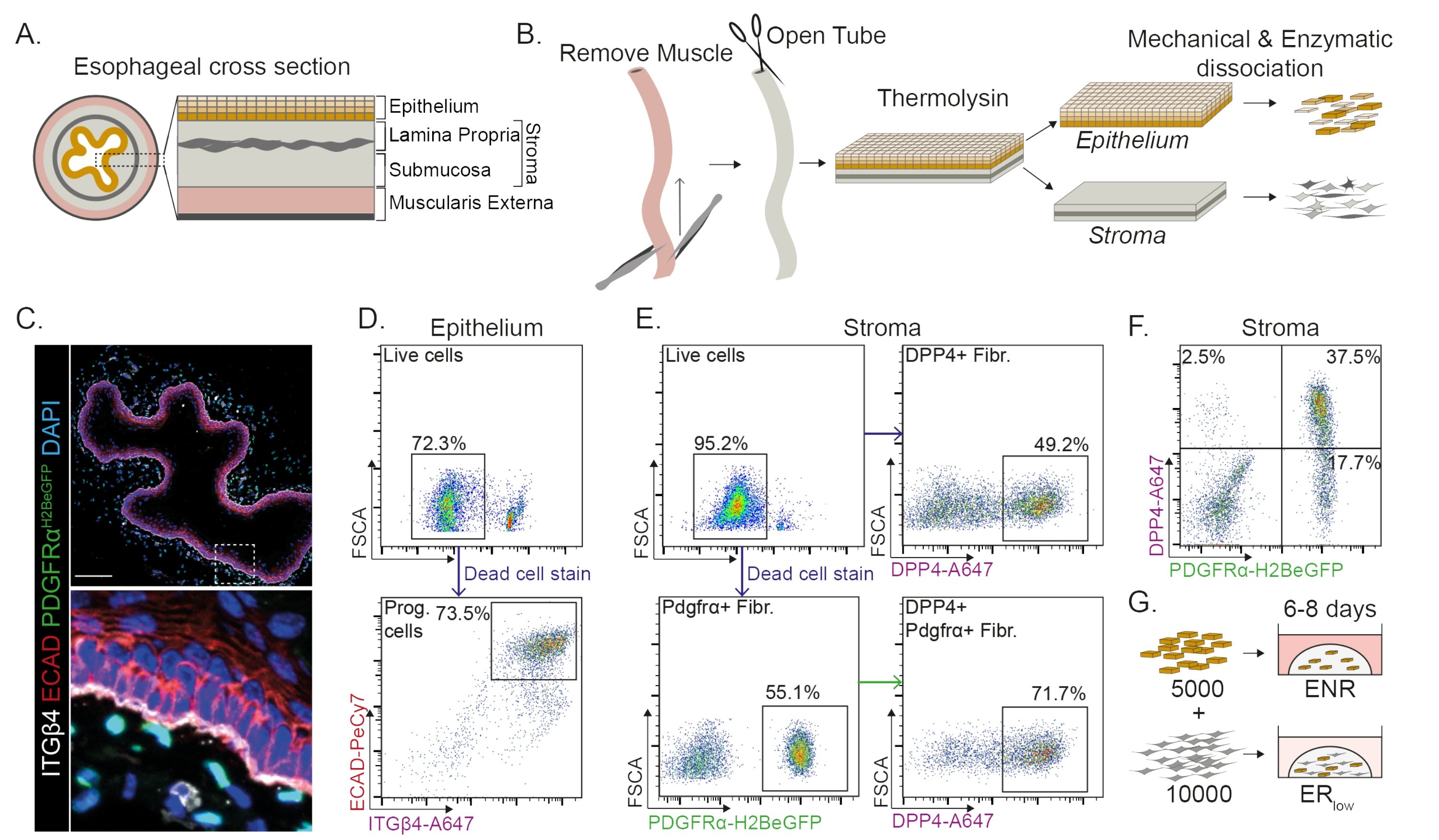
식도는 상피층, 고유판, 점막하 및 외근과 같은 여러 층으로 나뉩니다(그림 1A). 섬유아세포는 기질이라고 하는 점막하층과 고유판 내에 존재합니다. 이 프로토콜에서 근외근은 기계적으로 제거되며(그림 1B), 이는 기질에 존재하는 섬유아세포(PdgfrαH2BeGFP+)의 손실로 이어지지 않습니다(그림 1C). 해리되기 전에 상피는 간질에서 분리되어 두 개의 조직 세그먼트를 생성합니다(그림 1B). 두 층을 분리하는 것은 깨지기 쉬운 간질 층에 비해 더 견고한 상피층에 대한 해리 시간을 증가시킬 수 있는 기회를 제공합니다. 이러한 방식으로, 생존 가능한 상피 전구 세포와 기질 섬유아세포를 모두 생성하는 효율적인 분리 프로토콜이 확립됩니다(그림 1B). 식도 전구 세포는 높은 INTEGRIN-β4 및 E-CADHERIN 발현에 따라 분류됩니다(그림 1C,D).
섬유아세포의 하위집단은 별개의 마커를 사용하여 분리될 수 있다. 이 프로토콜에서는 일반적으로 사용되는 섬유아세포 마커 PDGFRα 및 DPP4(CD26)에 기초한 섬유아세포 분리를 위한 전략이 제공됩니다. PdgfrαH2BeGFP 리포터 발현 또는 DPP4 항체에 의한 분리는 분리된 세포의 약 50%가 섬유아세포임을 보여줍니다(그림 1E,F). 또한, PDGFRα+ 섬유아세포의 70%는 DPP4+이며, 이는 대체로 겹치지만 동일하지는 않은 섬유아세포 집단이 얻어진다는 것을 나타낸다. 상피 및 기질 세포 집단을 모두 분리한 후, 식도 전구 세포는 단독으로 또는 매트릭스 돔에서 섬유아세포와 함께 배양됩니다. 오가노이드 형성에 대한 섬유아세포의 기여를 연구하기 위해, 공동 배양은 성장 인자 감소 배지에서 유지된다(도 1G).
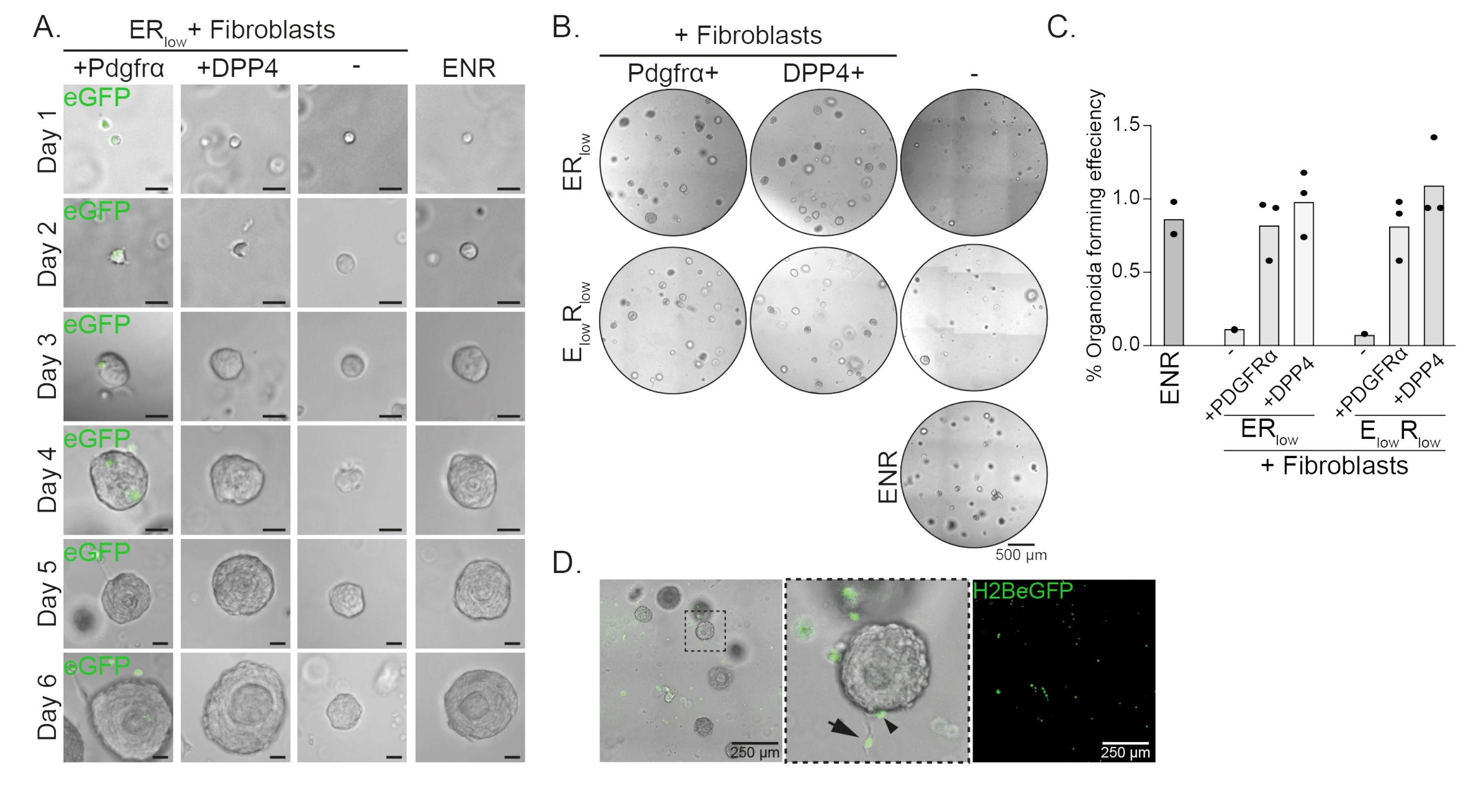
식도 전구 세포는 EGF, NOGGIN 및 RSPO(ENR)의 존재 하에 오가노이드를 형성합니다. NOGGIN 제거 및 RSPO 투여량 감소(25ng/μL; ER이낮음)은 오가노이드 형성을 방지하기에 충분하다(도 2A). 흥미롭게도, ER저배지의 식도 전구 세포에 DPP4+ 또는 PDGFRα+ 섬유아세포를 추가하면 오가노이드 형성 능력이 회복되어 두 섬유아세포 집단 모두에 대한 지지 기능을 입증합니다(그림 2A-D). PdgfrαH2BeGFP 이식유전자의 시각화는 섬유아세포가 오가노이드 형성 동안 상피 전구 세포와 밀접하게 접촉한다는 것을 보여줍니다(그림 2A). 6일째에 PdgfrαH2BeGFP+ 섬유아세포는 여전히 공동 배양에 풍부하게 존재합니다. 섬유아세포는 돔 전체에 존재하며, 오가노이드에 가까이 접하거나(full arrow), 오가노이드에 부착(arrowhead; 그림 2D).
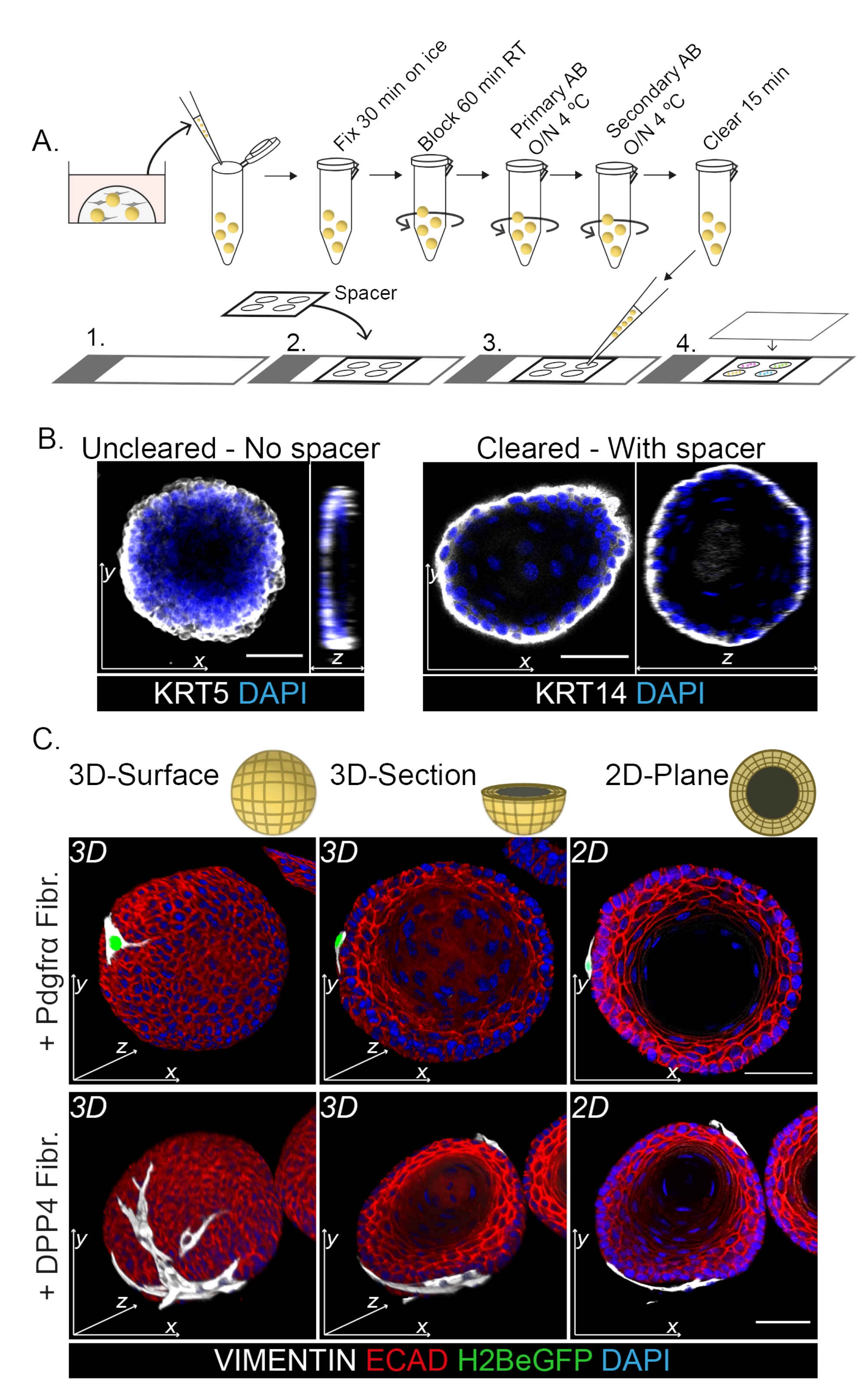
전체 마운트 염색은 섬유아세포와 오가노이드의 상호작용을 3D로 표현한 것입니다(그림 3). 투명 용액을 사용하지 않고 전체 마운트 프로토콜을 수행할 수 있지만 오가노이드의 투명도와 레이저 투과율이 감소합니다(그림 3B, z-view). 오가노이드를 장착할 때 스페이서는 오가노이드 형태를 유지하는 데 도움이 됩니다. 대조적으로, 커버슬립을 오가노이드에 직접 도금하면(스페이서 없이) 오가노이드가 평평해지고 오가노이드 구조가 손실됩니다(그림 3A, B).
DPP4+ 및 PDGFRα+ 섬유아세포는 모두 오가노이드를 감싸고 있는 것으로 밝혀졌습니다(그림 3C, 비디오 1 및 비디오 2). 식도 오가노이드의 분화는 다른 마커를 사용하여 평가할 수 있습니다. 그림 4는 제공된 염색 프로토콜이 염색하기 쉬운 케라틴(KRT14/13)과 염색하기 어려운 전사 인자(TRP63/KLF4)에 적합하다는 것을 보여줍니다. 공동 배양 프로토콜은 생체 내13,14에서 입증되고 ENR 배지에서 성장한 오가노이드에서 볼 수 있듯이 유사한 분화 패턴을 가진 오가노이드를 생성합니다. KRT14+ 또는 TRP63+ 전구세포는 외부층을 형성하고 KRT13+ 또는 KLF4+ 분화된 세포는 안쪽을 향합니다.
이 프로토콜은 시험관 내에서 식도 줄기 세포 틈새를 연구하고 오가노이드와 섬유아세포 사이의 상호 작용을 시각화하는 도구를 제공합니다. 항체를 사용하여 섬유아세포를 분리하기 위한 프로토콜을 구현함으로써, 상기 방법은 적응 가능하며, 형질전환 마우스의 필요 없이 섬유아세포 서브집단을 연구하는데 사용될 수 있다.

그림 1: 식도에서 전구 세포와 섬유아세포 하위 집단의 분리. (A) 식도의 여러 층에 대한 개략도. 간질에는 고유판과 점막하층이 있습니다. (B) 격리 프로토콜의 개략도. 근육 (muscularis externa)은 집게를 사용하여 기계적으로 제거됩니다. 나머지 식도는 절단되어 Thermolysin에서 배양되어 간질에서 상피층을 분리합니다. 상피와 간질은 분리되고, 기계적으로 다져지고, 효소에 의해 단일 세포 현탁액으로 소화됩니다. 이어서, 해리된 세포를 염색하고 FACS를 준비한다. (C) 간질에서 PdgfrαH2BeGFP+ 섬유아세포를 보여주는 근외근에서 벗겨진 식도의 단면. INTEGRIN-β4(ITGβ4) 및 E-CADHERIN(ECAD) 이중 양성 세포는 식도의 상피 전구 세포입니다. 스케일 바 = 100 μm. (D) 모든 단일 세포에서 살아있는 세포(상부 패널)의 백분율을 보여주는 상피 세포 분리의 대표적인 유세포 분석 플롯. 하단 패널은 모든 살아있는 세포에서 분리된 ITGβ4+ ECAD+ 전구세포(Prog.) 세포의 백분율을 보여줍니다. (E) 살아있는 세포의 백분율을 보여주는 기질 세포 분리의 대표적인 유세포 분석 플롯(왼쪽 상단 패널). 모든 살아있는 세포의 분리된 DPP4+ 섬유아세포(Fibr.; 우측 상단 패널) 및 Pdgfrα+ 섬유아세포(좌측 하단 패널)의 백분율을 보여주는 대표적인 유세포 분석 플롯. Pdgfrα+ 섬유아세포의 70%도 DPP4+(오른쪽 하단 패널)입니다. (F) DPP4+ 단독 세포(2.5%), DPP4+ PDGFRα+ 세포(37.5%) 및 PDGFRα+ 단독 세포(17.7%)를 보여주는 기질의 대표적인 유세포 분석 플롯. 백분율은 모든 살아있는 세포의 백분율입니다. (G) 상피 전용 세포는 50ng/μL EGF, 100ng/μL NOGGIN 및 250ng/μL RSPO(ENR)의 존재 하에 또는 EGF 및 저농도 RSPO(25ng/μL)의 존재 하에 섬유아세포와 함께 매트릭스 돔에 도말됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 2: 오가노이드 공동 배양의 대표적인 결과. (A) 1일차부터 6일차까지 오가노이드의 성장을 보여주는 명시야 이미지. PdgfrαH2BeGFP+ 섬유아세포와 공동 배양된 오가노이드의 명시야 이미지도 핵 eGFP 신호를 보여줍니다. 스케일 바 = 25 μm. (B) 6일째 전체 매트릭스 돔의 명시야 이미지. 왼쪽 열은 ER low 또는 E low R low배지에서 Pdgfrα+ 섬유아세포의 존재 하에 성장한 오가노이드 공동 배양을 보여줍니다. 중간 컬럼은 ER low 또는 E low R low배지에서 DPP4+ 섬유아세포의 존재 하에 성장한 오가노이드 공동 배양을 보여줍니다. 오른쪽 열은 ENR 배지에서 성장한 오가노이드 단일 배양을 보여줍니다. ENR 배지 = EGF(50ng/μL), NOGGIN(100ng/μL) 및 RSPO(250ng/μL). ER낮음 = EGF 및 25ng/μL RSPO. Elow Rlow = 5 ng/μL EGF 및 25 ng/μL RSPO. 스케일 바 = 500 μm. (C) 오가노이드 형성 효율(%)을 나타내는 그래프(즉, 상이한 배양 조건에서 오가노이드를 형성하는 세포의 비율). 각 점은 행렬 돔을 나타내고 막대는 조건당 모든 점의 평균을 나타냅니다. (D) PdgfrαH2BeGFP+ 섬유아세포와 공동 배양한 6일차 오가노이드의 명시야 및 형광 이미지. PdgfrαH2BeGFP+ 섬유아세포는 돔 전체에 존재하며 오가노이드에 부착되고(화살촉) 부착되지 않았지만 오가노이드와 접촉합니다(전체 화살표). 스케일 바 = 250 μm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3: 섬유아세포-오가노이드 상호작용 연구를 위한 전체 마운트 염색 프로토콜. (A) 전체 마운트 면역형광 프로토콜의 개략도. AB = 항체. (B) 투명화되지 않은 오가노이드에 비해 레이저 광의 투명성 및 침투율이 감소함을 보여주는 투명화되지 않은 전체 마운트 염색의 면역형광 사진. 스페이서가 없으면 오가노이드가 평탄화되고 오가노이드 형태가 손실됩니다. (C) 공동 배양된 오가노이드의 전체 마운트 이미지는 VIMENTIN+ 섬유아세포(Fibr.)가 오가노이드를 감싸고 밀접하게 접촉한 오가노이드의 3D 표면을 보여줍니다. 3D 단면과 2D 평면 이미지는 오가노이드의 내강을 보여줍니다. 스케일 바 = 50 μm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 4: 전체 마운트 이미지는 뚜렷한 기저 및 기저 상부 세포 집단을 보여줍니다. (A) 외층의 KRT14+ 기저 세포와 KRT13+ 분화된 기저 상부 세포를 보여주는 PdgfrαH2BeGFP+ 섬유아세포를 사용한 단일 및 공동 배양 오가노이드의 전체 마운트 염색. 스케일 바 = 50 μm. (B) 외층의 TRP63+ 기저 세포와 KLF4+ 분화된 기저 상부 세포를 보여주는 PdgfrαH2BeGFP+ 섬유아세포를 사용한 단일 및 공동 배양된 오가노이드의 전체 마운트 염색. 스케일 바 = 50 μm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
표 1: 오가노이드 배양 배지 성분을 설명하는 표. 이 표를 다운로드하려면 여기를 클릭하십시오.
동영상 1: PdgfrαH2BeGFP+ 섬유아세포가 오가노이드를 감싸고 밀접하게 접촉하고 있습니다. 비디오는 그림 3C의 상단 패널과 함께 제공됩니다. 그림 3C 의 스케일 바는 50 μm이고 오가노이드는 직경이 ~120 μm입니다. VIMENTIN은 흰색, E-CADHERIN은 빨간색, PdgfrαH2BeGFP는 녹색, DAPI는 파란색으로 표시됩니다. 이 비디오를 다운로드하려면 여기를 클릭하십시오.
비디오 2: DPP4+ 섬유아세포가 오가노이드를 감싸고 밀접하게 접촉합니다. 비디오는 그림 3C의 하단 패널과 함께 제공됩니다. 그림 3C 의 스케일 바는 50 μm이고 오가노이드는 직경이 ~120 μm입니다. VIMENTIN은 흰색, E-CADHERIN은 빨간색, DAPI는 파란색으로 표시됩니다. 이 비디오를 다운로드하려면 여기를 클릭하십시오.
토론
여기에 제시된 프로토콜은 기능적 식도 상피-섬유아세포 상호작용을 조사하기 위한 시험관 내 모델을 설정합니다.
상피층은 기질에서 분리되어 상피 및 기질 구획 모두에 대해 최적화된 해리 프로토콜을 허용합니다. 상피 해리 프로토콜의 최적화에도 불구하고 조직 덩어리는 여전히 분명합니다. 15분마다 세게 위아래로 피펫을 하면 덩어리의 수와 크기가 크게 줄어듭니다. 다른 프로토콜은 트립신을 사용하여 상피층 5,6을 추가로 해리시킵니다. 여기서, 트립신의 사용, 또는 해리 시간을 추가로 증가시키는 것은 바람직하지 않은데, 이는 상피 세포 생존율 및 오가노이드 형성 효율의 감소를 초래하는 경향이 있기 때문이다. 상피와 달리 간질은 쉽게 해리되며 해리 용액에서 30분 동안 ~90%의 섬유아세포 생존율을 가진 단일 세포 현탁액이 생성됩니다(그림 1E). 프로토콜에서 상피-기공 분리 단계를 제외하면 해리 시간이 상당히 증가하여 섬유아세포 생존율이 감소하고 상피 세포의 수율이 낮아집니다. 또한, 간질에서 상피를 분리하면 각 집단의 세포 수를 결정하고 공동 배양을 설정할 때 상피 세포와 다른 마우스 계통의 섬유아세포를 혼합할 수 있습니다.
오가노이드 성장에 대한 섬유아세포 기능을 연구하는 것은 줄기 세포 생물학에서 일반적으로 사용되는 방법이다 9,10,11,15,16. 확립된 공동 배양 배지는 10% 태아 송아지 혈청(FCS)9,15 또는 성장 인자 감소 배지10,16으로 보충된 DMEM입니다. 이 프로토콜에서, 성장 인자 환원 배지는 섬유아세포가 대체로 정지 상태인 생체내 줄기 세포 틈새의 조건을 모방하기 위해 사용된다. FCS는 공동 배양에서 섬유아세포의 활성화 및 증식을 초래하는 성장 인자 풍부 혈청이며, 생체 내 상태와 구별되는 섬유아세포 상태에 해당할 가능성이 높습니다. FCS를 배제하고 성장 인자를 감소시킴으로써, 배지 단독(ERlow low)이 오가노이드 성장을 지지하지 않고 섬유아세포 증식을 자극하지 않도록, 오가노이드 성장에 대한 섬유아세포의 효과를 단리할 수 있다. 이 배지에서 NOGGIN이 제거되고 RSPO가 최소(10% RPSO)로 감소합니다. NOGGIN과 RSPO는 모두 식도 오가노이드 성장에 필수적인 것으로 입증되었다6. EGF는 그 자체로 오가노이드 성장을 지원하지 않기 때문에 공동 배양 배지에 유지되었습니다. 그러나, 섬유아세포는 또한 EGF 환원 배지에서 오가노이드 성장을 지원할 수 있다 (ElowRlow; 그림 2B, D).
오가노이드 공동 배양은 트립신 처리 중에 섬유아세포가 손실되기 때문에 계대배양을 통해 지속될 수 없습니다. 그러나 식도 오가노이드가 유지, 확장 및 단일 배양으로 추가 실험에 사용될 수 있기 때문에 오가노이드 계대배양이 프로토콜에 포함되었습니다. 단일 배양의 계대 오가노이드는 새로 분리된 섬유아세포와 공동 배양을 설정하는 데 사용할 수 있습니다. 1차 세포 사용의 단점은 여러 오가노이드 공동 배양을 설정하는 데 필요한 마우스의 수입니다. 섬유아세포의 작은 하위 집단에 초점을 맞출 때, 얻어지는 공동 배양의 수는 제한적이다. 다른 프로토콜에서, 섬유아세포는 오가노이드 공동배양을 설정하기 위해 섬유아세포를 사용하기 전에 먼저 배양에서 확장된다10. 그러나, 섬유아세포는 패시징 동안 형태 및 정체성을 변화시키며, 이는 일차 피부 및 심장 섬유아세포를 사용하여 나타난다17,18. 식도 섬유아세포의 기존 2D 계대배양은 형태 및 표현형 변화를 모두 초래하며, 이는 섬유아세포의 시험관 내 농축이 내인성 줄기 세포 틈새의 표현화를 목표로 하는 공동 배양에 적합하지 않음을 보여줍니다.
전체 마운트 염색은 섬유아세포-오가노이드 상호작용을 유지하고 시각화하는 도구를 제공합니다. 모든 오가노이드에 섬유아세포가 직접 부착되어 있는 것은 아니지만 대부분의 오가노이드는 섬유아세포와 접촉한다는 점에 유의해야 합니다( 그림 2C 참조). 상피-섬유아세포 상호작용을 유지하려면 오가노이드를 주의해서 다루고 격렬한 피펫팅, 와류화 및 고속 회전을 피하는 것이 중요합니다. 최적의 고정은 3D 조직 구조를 유지하고 내인성 형광을 유지하는 데 중요합니다. 30분 고정은 H2BeGFP 신호를 유지하기에 충분하며 이 프로토콜에 사용되는 항체에 최적이지만 사용된 형광단과 항체에 따라 다를 수 있습니다. 오가노이드를 제거하면 광 산란이 감소하고 전체 3D 구조의 시각화가 크게 향상됩니다. 오가노이드가 작기 때문에 청소가 쉽고 빠릅니다. 그러나 레이저 스캐닝 컨포칼 현미경을 사용하여 전체 오가노이드를 이미징하는 것은 여러 Z 스택을 만들어야 하기 때문에 시간이 많이 소요될 수 있습니다. 스피닝 디스크와 같은 컨포칼 현미경을 사용하여 이미징 시간을 줄일 수 있습니다.
전반적으로, 섬유아세포가 있는 상태에서 성장한 식도 오가노이드는 식도 줄기 세포 틈새의 측면을 이해하는 데 유용한 도구를 제공합니다. 또한 전체 마운트 클리어링은 섬유아세포와 오가노이드 간의 상호 작용을 시각화할 수 있는 접근 가능한 방법을 제공합니다.
공개
저자는 이해 상충을 선언하지 않습니다.
감사의 말
이 연구는 ERC StG TroyCAN(851241)의 지원을 받았습니다. E.E.는 Cancerfonden 박사후 연구원입니다. MG는 Ragnar Söderberg 펠로우이자 Cancerfonden Junior Investigator입니다. 우리는 Biomedicum Flow Cytometry Core Facility, Biomedicum Imaging Core (BIC) 및 Comparative Medicine Biomedicum (KMB) 동물 시설을 포함한 Karolinska Institutet 핵심 시설의 기술 지원에 감사드립니다. 프로토콜을 주의 깊게 읽고 의견을 제시해 주신 Genander 연구소 구성원들에게 감사드립니다.
자료
| Name | Company | Catalog Number | Comments |
| B-27 Supplement (50X), serum free | Thermo Fisher (Gibco) | 17504001 | |
| Corning Matrigel Growth Factor Reduced (GFR) Basement Membrane Matrix | fisher scientific | 356231 | |
| Dimethyl sulfoxide | Sigma-Aldrich | 276855-100ML | |
| DMEM/F-12 | Thermo Fisher (Gibco) | 11320074 | |
| DPBS | Thermo Fisher (Gibco) | 14190250 | |
| Fetal Bovine Serum | Sigma-Aldrich | F7524 | |
| GlutaMAX Supplement | Thermo Fisher (Gibco) | 35050061 | |
| HBSS, no calcium, no magnesium, no phenol red | Thermo Fisher (Gibco) | 14175-129 | |
| Normal Donkey Serum | Jackson Immuno | 017-000-121 | |
| Penicillin-Streptomycin (10,000 U/mL) | Thermo Fisher (Gibco) | 15140122 | |
| Triton X-100 solution | Merck | 93443-100ML | |
| Trypsin-EDTA (0.25%), phenol red | Thermo Fisher (Gibco) | 25200-056 | |
| Chemicals, Peptides, and recombinant proteins | |||
| DAPI Solution | Thermo Fisher | 62248 | |
| Dissociation solution: 0.25 mg/ml Liberase TM, 0.25 mg/ml Dnase in HBSS | |||
| Dnase I | Sigma-Aldrich | 11284932001 | |
| Formaldehyde, 37%, with 10-15% methanol | Sigma-Aldrich | 252549-1L | |
| Liberase | Sigma-Aldrich | 5401127001 | |
| N-Acetyl-cysteine | Sigma-Aldrich | A9165-25G | |
| Noggin murine | Peprotech | 250-38 | |
| RapiClear 1.47 | SunJin Lab | #RC147001 | |
| Recombinant Mouse EGF Protein, CF | R&D systems | 2028-EG-200 | |
| R-spondin-1 murine | Peprotech | 315-32 | |
| SYTOX Blue Dead Cell Stain | Thermo Fisher | S34857 | |
| Thermolysin | Sigma-Aldrich | T7902-25MG | |
| Y-27632 dihydrochloride | Sigma-Aldrich | Y0503-5MG | |
| Plastic & Glassware | |||
| Corning Sterile Cell Strainers 40um | VWR | 15360801 | |
| Corning Sterile Cell Strainers 70um | VWR | 431751 | |
| Menzel Deckgläser/ cover slips | Thermo Fisher | Q10143263NR15 | |
| SafeSeal reaction tube, 1.5 mL, PP | Sarstedt | 72.706 | |
| Snap Cap Low Retention Microcentrifuge Tubes 0.6 mL | Thermo Fisher | 3446 | |
| SuperFrost Slides | VWR | 631-9483 | |
| Tools | |||
| 0.05 mm 4 circular well iSpacer | SunJin Lab | #IS204 | |
| Dumont #5 forceps, biology tip | F.S.T | 11252-20 | |
| ImmEdge Pen | VectorLaboratories | H-4000 | |
| Spring Scissors Angled to Side Ball Tip 8mm Cutting Edge | F.S.T | 15033-09 | |
| Instruments | |||
| Confocal microscope Stellaris 5 | Leica | ||
| Dissection microscope ZEISS Stemi 305 | Zeiss | ||
| FACS ARIA III | BD Biosciences | ||
| Conjugated Antibodies for FACS | |||
| Alexa Fluor 647 anti-mouse CD104 Antibody Clone: 346-11A | 123608 | 123608 | |
| APC anti-mouse CD26 (DPP-4) Antibody | H194-112 | H194-112 | |
| PE/Cy7 anti-mouse/human CD324 (E-Cadherin) Antibody | 147310 | 147310 | |
| Antibodies for Immunofluorescence | |||
| CD104 (ß-integrin 4) Clone: 346-11A | BioLegend | 553745 | |
| Cytokeratin 14 | Acris Antibodies (AbD Serotec) | BP5009 | |
| Cytokeratin13 Clone: EPR3671 | abcam | ab92551 | |
| E-cadherin (CD324) Clone: 2.40E+11 | Cell Signaling Technology | 3195 | |
| Keratin 5 Polyclonal Chicken Antibody, Purified Clone: Poly9059 | BioLegend | 905901 | |
| p63 Clone: 4a4 | abcam | ab735 | |
| Recombinant Anti-KLF4 antibod Clone: EPR20753-25 | abcam | ab214666 | |
| Vimentin | Sigma-Aldrich | AB5733 | |
| Secondary antibodies | |||
| Donkey anti-species* antibodies with fluorophore of choice | Jackson Immuno |
참고문헌
- Sachs, N., et al. Long-term expanding human airway organoids for disease modeling. The EMBO Journal. 38 (4), 100300 (2019).
- Lohmussaar, K., et al. Patient-derived organoids model cervical tissue dynamics and viral oncogenesis in cervical cancer. Cell Stem Cell. 28 (8), 1380-1396 (2021).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459 (7244), 262-265 (2009).
- Smukler, S. R., et al. The adult mouse and human pancreas contain rare multipotent stem cells that express insulin. Cell Stem Cell. 8 (3), 281-293 (2011).
- Zheng, B., et al. A new murine esophageal organoid culture method and organoid-based model of esophageal squamous cell neoplasia. iScience. 24 (12), 103440 (2021).
- DeWard, A. D., Cramer, J., Lagasse, E. Cellular heterogeneity in the mouse esophagus implicates the presence of a nonquiescent epithelial stem cell population. Cell Reports. 9 (2), 701-711 (2014).
- Kasagi, Y., et al. The esophageal organoid system reveals functional interplay between notch and cytokines in reactive epithelial changes. Cellular and Molecular Gastroenterology and Hepatology. 5 (3), 333-352 (2018).
- Plikus, M. V., et al. Fibroblasts: Origins, definitions, and functions in health and disease. Cell. 184 (15), 3852-3872 (2021).
- McCarthy, N., et al. Distinct mesenchymal cell populations generate the essential intestinal BMP signaling gradient. Cell Stem Cell. 26 (3), 391-402 (2020).
- Cordero-Espinoza, L., et al. Dynamic cell contacts between periportal mesenchyme and ductal epithelium act as a rheostat for liver cell proliferation. Cell Stem Cell. 28 (11), 1907-1921 (2021).
- Pastula, A., et al. Three-dimensional gastrointestinal organoid culture in combination with nerves or fibroblasts: a method to characterize the gastrointestinal stem cell niche. Stem Cells International. 2016, 3710836 (2016).
- Hamilton, T. G., Klinghoffer, R. A., Corrin, P. D., Soriano, P. Evolutionary divergence of platelet-derived growth factor alpha receptor signaling mechanisms. Molecular and Cellular Biology. 23 (11), 4013-4025 (2003).
- McGinn, J., et al. A biomechanical switch regulates the transition towards homeostasis in oesophageal epithelium. Nature Cell Biology. 23 (5), 511-525 (2021).
- Zhang, Y., Bailey, D., Yang, P., Kim, E., Que, J. The development and stem cells of the esophagus. Development. 148 (6), (2021).
- Ohlund, D., et al. Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. The Journal of Experimental Medicine. 214 (3), 579-596 (2017).
- Pentinmikko, N., et al. Notum produced by Paneth cells attenuates regeneration of aged intestinal epithelium. Nature. 571 (7765), 398-402 (2019).
- Janson, D., Rietveld, M., Willemze, R., El Ghalbzouri, A. Effects of serially passaged fibroblasts on dermal and epidermal morphogenesis in human skin equivalents. Biogerontology. 14 (2), 131-140 (2013).
- Landry, N. M., Rattan, S. G., Dixon, I. M. C. An improved method of maintaining primary murine cardiac fibroblasts in two-dimensional cell culture. Scientific Reports. 9 (1), 12889 (2019).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유