
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
In Vitro Химическое картирование структур ДНК G-квадруплекса бис-3-хлорпиперидинами
В этой статье
Резюме
Бис-3-хлорпиперидины (B-CeP) являются полезными химическими зондами для идентификации и характеристики структур G-квадруплекса в матрицах ДНК in vitro. В этом протоколе подробно описана процедура проведения зондирующих реакций с B-CeP и растворения продуктов реакции с помощью электрофореза в полиакриламидном геле высокого разрешения.
Аннотация
G-квадруплексы (G4) — это биологически значимые, неканонические структуры ДНК, которые играют важную роль в экспрессии генов и заболеваниях, представляя собой важные терапевтические мишени. Для характеристики ДНК in vitro в потенциальных G-квадруплекс-образующих последовательностях (PQS) требуются доступные методы. B-CeP — это класс алкилирующих агентов, которые оказались полезными химическими зондами для исследования структуры нуклеиновых кислот высшего порядка. В данной работе описывается новый химический картирующий анализ, использующий специфическую реакционную способность B-CeP с N7 гуанинов с последующим прямым расщеплением цепей в алкилированных Gs.
А именно, чтобы отличить складки G4 от развернутых форм ДНК, мы используем B-CeP 1 для зондирования тромбин-связывающего аптамера (TBA), 15-мерной ДНК, способной принимать расположение G4. Реакция B-CeP-реагирующих гуанинов с B-CeP 1 дает продукты, которые могут быть разрешены с помощью электрофореза в полиакриламидном геле (PAGE) высокого разрешения на однонуклеотидном уровне путем локализации отдельных аддуктов алкилирования и расщепления цепей ДНК на алкилированных гуанинах. Картирование с использованием B-CePs является простым и мощным инструментом для характеристики in vitro последовательностей ДНК, образующих G-квадруплекс, что позволяет точно определить местоположение гуанинов, участвующих в образовании G-тетрад.
Введение
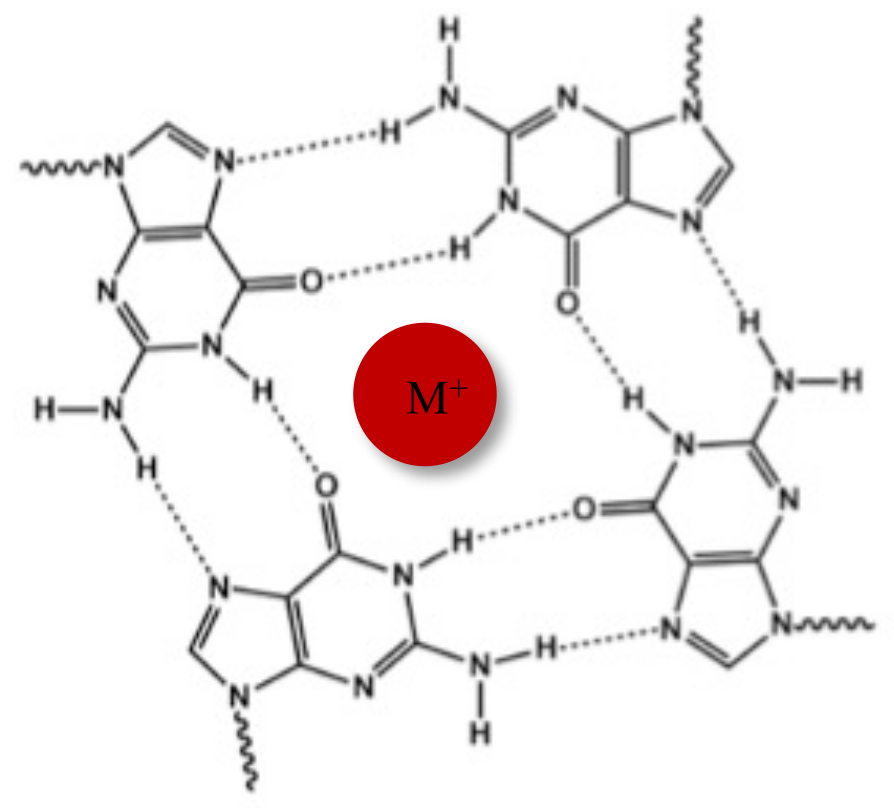
В дополнение к типичной двойной спирали Уотсона-Крика, нуклеиновые кислоты могут принимать различные вторичные структуры, такие как альтернативная форма G-квадруплекса (G4), из-за их богатых гуанином последовательностей. Структура G4 основана на образовании плоских тетрамеров, называемых G-тетрадами, в которых четыре гуанина взаимодействуют через водородные связи Хугстина. G-тетрады укладываются друг на друга и дополнительно стабилизируются одновалентными катионами, которые координируются в центре гуанинового ядра (рис. 1)1.

Рисунок 1: Схематическое изображение G-квадруплексной структуры. (A) Схематическое изображение G-тетрады. Планарная решетка стабилизируется спариванием оснований по Хугстину и центральным катионом (M+). Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
Последовательности, содержащие четыре или более последовательных прогонов, по крайней мере, двух последовательных гуаниновых нуклеотидов, являются потенциальными G-квадруплекс-образующими последовательностями (PQS), которые могут сворачиваться в G-квадруплексные структуры. PQS расположены во многих различных клеточных контекстах, таких как теломеры, промоторы генов, рибосомная ДНК и рекомбинационные сайты, и участвуют в регуляции многих биологическихпроцессов. Таким образом, идентификация и экспериментальная проверка G4 в геноме человека, которая в настоящее время осуществляется в основном с помощью вычислительных инструментов, является биологически значимойпроблемой. Для поддержки вычислительных прогнозов или обнаружения непредсказанных структур G4 здесь показан доступный метод, основанный на химическом картировании, для идентификации образования G4 в матрице ДНК, позволяющий точно идентифицировать гуанины, образующие структуру G-тетрады.
В представленном химическом картированном анализе используется различная реакционная способность бис-3-хлорпиперидинов (B-CeP) с гуанинами после образования структур G4. Благодаря своей высокой реакционной способности с нуклеофилами 4,5,6,7,8,9, B-CeP являются алкилирующими агентами нуклеиновых кислот, обладающими способностью очень эффективно реагировать с положением N7гуаниновых нуклеотидов10. Алкилирование сопровождается депуринацией и расщеплением цепей в одноцепочечных и двухцепочечных конструкциях ДНК. Напротив, гуанины, участвующие в образовании G-тетрад в G4-расположениях, невосприимчивы к алкилированию B-CeP, так как положение N7 гуанинов вовлечено в водородные связи Хугстина. Эта специфическая реакционная способность B-CeP позволяет не только обнаруживать структуры G4, но и идентифицировать гуанины, образующие тетраду (тетрады), поскольку они могут быть выведены из их относительной защиты от алкилирования по сравнению с гуанинами в одноцепочечной и двухцепочечной ДНК.
Протокол химического картирования представлен здесь с использованием B-CeP 1 (рис. 2A) в качестве зонда для характеристики тромбин-связывающего аптамера (TBA), 15-мерной ДНК, способной принимать расположение G4 в присутствии катионов калия11,12. Расположение G4 TBA (G4-TBA) напрямую сравнивается с двумя контрольными группами, а именно TBA в одноцепочечной форме (ssTBA) и TBA, отожженной до ее комплементарной последовательности с образованием двухцепочечной конструкции (dsTBA) (табл. 1). Продукты зондирующих реакций растворяются с помощью электрофореза в полиакриламидном геле (PAGE) высокого разрешения на однонуклеотидном уровне путем локализации отдельных аддуктов алкилирования и расщепления цепей ДНК на алкилированных гуанинах. Визуализация на геле обеспечивается конъюгацией олигонуклеотида TBA с флуорофором на его 3'-конце (табл. 1). В этом протоколе показано, как сворачивать TBA в различных конформациях (G4 и контроль), а также как проводить зондирующие реакции с B-CeP с последующим PAGE.
протокол
1. Подготовка нуклеиновых кислот и химических зондов
- Нуклеиновые кислоты
ПРИМЕЧАНИЕ: Олигонуклеотид под названием «TBA» представляет собой 15-мерную последовательность ДНК 5'-GGT-TGG-TGG-TGT-GGT-TGG-3', помеченную на 3'-конце флуорофором 5-карбоксифлуоресцеином (FAM) для обеспечения визуализации на геле. Немеченый олигонуклеотид "cTBA" представляет собой его комплементарную последовательность ДНК 5'-CCA-ACC-ACA-CCA-ACC-3'. TBA и cTBA используются для получения трех различных структур, как показано в таблице 1.- Наконечники для автоклавов и пробирки по 0,5 мл для получения стерильных одноразовых материалов и избежания загрязнения.
- Готовят исходные растворы, растворяющие каждый олигонуклеотид в сверхчистой воде до конечной концентрации 100 мкМ. Точную концентрацию олигонуклеотидов определяют с помощью спектрофотометра в ультрафиолетовом диапазоне видимого диапазона (UV-Vis), используя коэффициент экстинкции при 260 нм, предоставленный производителем.
ПРИМЕЧАНИЕ: Коэффициенты экстинкции: 164 300 М-1 см-1 и 138 600 М-1 см-1 были использованы для TBA и cTBA, соответственно. - Храните исходные растворы TBA и cTBA при температуре -20 °C (в течение нескольких месяцев в этих условиях).
- Соединение B-CeP 1
ПРИМЕЧАНИЕ: Соединение B-CeP 1 синтезируется, как сообщалось ранее6.- Приготовьте исходный раствор B-CeP 1 при ~10 мМ. Взвесьте ~1 мг лиофилизированного соединения с помощью аналитических весов, расположенных в вытяжном шкафу, и растворите его в 100% диметилсульфоксиде (ДМСО).
- Рассчитайте точную концентрацию соединения на основе фактического количества используемого соединения и ДМСО (d = 1,1 г/см3).
ПРИМЕЧАНИЕ: Всегда обращайтесь с соединением в перчатках (как при лиофилизации, так и при растворении в ДМСО)13,14.
Таблица 1: Олигонуклеотидные структуры, используемые в данном протоколе. Пожалуйста, нажмите здесь, чтобы скачать эту таблицу.
2. Сворачивание конструкций нуклеиновых кислот
- Подготовка буферов
- Приготовьте буферный раствор BPE (бифосфат-этилендиаминтетрауксусная кислота [ЭДТА], 5x: 2 мМ2PO 4, 6 мМ Na2 HPO 4, 1 мМNa2ЭДТА, pH7,4) и раствор 500 мМ KCl в сверхчистой воде. Отфильтруйте растворы через фильтры с размером пор 0,22 мкм.
ПРИМЕЧАНИЕ: Для достижения наилучших результатов используйте свежеприготовленные растворы. Буфер BPE можно хранить при температуре 4 °C до 15 дней.
- Приготовьте буферный раствор BPE (бифосфат-этилендиаминтетрауксусная кислота [ЭДТА], 5x: 2 мМ2PO 4, 6 мМ Na2 HPO 4, 1 мМNa2ЭДТА, pH7,4) и раствор 500 мМ KCl в сверхчистой воде. Отфильтруйте растворы через фильтры с размером пор 0,22 мкм.
- Сворачивание образцов G4-TBA, ssTBA и dsTBA методом повторного складывания
ПРИМЕЧАНИЕ: Катионы калия необходимы для сворачивания структуры G4 (G4-TBA). Не добавляйте катионы калия в раствор для сворачивания органов управления ssTBA и dsTBA.- Приготовьте 40 мкл 4 мкМ раствора G4-TBA в 1x BPE и 100 мМ KCl. Денатурируют раствор олигонуклеотида G4-TBA, нагревая пробирку до 95 °C в течение 5 мин, и медленно охлаждают ее до комнатной температуры (RT), чтобы TBA сложилась в G-квадруплексы.
- Приготовьте 40 мкл 4 мкМ раствора ssTBA в 1x BPE. Выполните процедуру термического складывания, как описано в шаге 2.2.1, чтобы сложить TBA в одноцепочечной форме.
- Приготовьте 40 мкл 4 мкМ раствора дцТБА путем смешивания эквимолярных количеств ТБА и цТБА в 1х BPE. Выполните процедуру термического складывания, как описано выше (шаг 2.2.1), для отжига TBA до его комплементарной последовательности cTBA и формирования двухцепочечной формы TBA.
ПРИМЕЧАНИЕ: Окончательный объем каждого складного раствора зависит от количества образцов для зондирующих реакций, учитывая, что для каждого образца потребуется 5 мкл раствора 4 мкМ. Приготовьте немного избыточного объема каждого раствора, чтобы избежать ошибок при пипетировании.
3. Зондирующие реакции
ПРИМЕЧАНИЕ: Зондирующие реакции должны быть выполнены сразу после процедуры термического складывания.
- Когда фальцовочные растворы G4-TBA, ssTBA и dsTBA остынут до RT, проводят центрифугирование с коротким отжимом (7 000 × г в течение 5-8 с при RT) и начинают зондирующие реакции.
- Подготовьте 21 пустую автоклавную пробирку объемом 0,5 мл. Организуйте их в три набора по семь пробирок в каждом на стойке для лабораторных образцов, как указано в таблице 2.
ПРИМЕЧАНИЕ: Каждый набор столбцов соответствует трем различным условиям свертывания TBA: G4-TBA, ssTBA и dsTBA. Каждый ряд соответствует трем различным срокам инкубации. Каждая ячейка в столбце соответствует конечной концентрации зонда B-CeP 1 (табл. 2). Убедитесь, что на тюбиках есть четкая маркировка. - Добавьте в каждую пробирку по 3 мкл сверхчистой воды.
- Добавьте 5 мкл свернутого G4-TBA в каждую пробирку первого набора (шаг 3.2). Добавьте 5 мкл свернутого ssTBA в каждую пробирку второго набора. Добавьте 5 мкл свернутого dsTBA в каждую пробирку третьего набора.
- Разбавьте исходный раствор B-CeP 1 до 250 мкМ и 25 мкМ в сверхчистой воде.
ПРИМЕЧАНИЕ: Разведения химического зонда B-CeP 1 должны быть свежеприготовленными и немедленно вступить в реакцию с субстратом ДНК, чтобы избежать конкурирующих реакций с водой. - Добавьте к образцам 2 мкл соответствующего разведения B-CeP 1 (25 мкМ и 250 мкМ). Замените соединение сверхчистой водой в трех контрольных образцах (С) для анализа по-разному свернутых TBA в отсутствие соединения. Инкубируют все образцы при температуре 37 °C.
- Через 1 ч, 4 ч и 15 ч инкубации остановите реакцию, поместив пробирки при -20 °C до следующего этапа.
ПРИМЕЧАНИЕ: Образцы можно хранить в этих условиях в течение нескольких дней. - Высушите образцы в вакуумной центрифуге.
ПРИМЕЧАНИЕ: Высушенные образцы можно хранить при температуре -20 °C в течение нескольких недель, прежде чем приступить к анализу PAGE.
Таблица 2: Образцы для зондирующих реакций (структуры, концентрации зондов и время инкубации). Каждый набор столбцов соответствует трем различным условиям свертывания TBA (G4-TBA, ssTBA и dsTBA). Каждый ряд соответствует трем различным срокам инкубации (1, 4, 15 ч). Каждая ячейка в колонке соответствует конечной концентрации зонда B-CeP 1 (5 или 50 мкМ). Контроль (С) для каждого набора соответствует образцу по-разному сложенных ТБА, инкубированных в течение более длительного времени (15 ч) в отсутствие соединения. Пожалуйста, нажмите здесь, чтобы скачать эту таблицу.
4. СТРАНИЦА с высоким разрешением
- Приготовление денатурирующего раствора полиакриламида
ПРИМЕЧАНИЕ: Заранее приготовьте 500 мл 20% денатурирующего раствора полиакриламидного геля. Для каждого эксперимента будет использовано около 80 мл этого раствора. Используйте бутылку из янтарного стекла или накройте стеклянную бутылку алюминиевой фольгой для хранения раствора в RT.
ВНИМАНИЕ: Полиакриламид нейротоксичен. На всех этапах приготовления и заливки геля надевайте перчатки и лабораторный халат. Выбросьте полимеризованный акриламид в соответствующую коробку для загрязненных материалов.- Взвесьте 210 г мочевины в стакане объемом 1 л. Добавьте 250 мл 40% раствора акриламида/бисакриламида (19/1) и 50 мл 10x клещевого энцефалита (890 мМ Tris-HCl, 890 мМ борат, 20 мМ ЭДТА, pH 8).
- Поместите стакан на пластину для перемешивания и перемешайте раствор с помощью мешалки. Накройте стакан алюминиевой фольгой во время смешивания, чтобы предотвратить разбрызгивание и загрязнение.
- Перемешивайте раствор до полного растворения мочевины и прозрачности раствора.
ПРИМЕЧАНИЕ: Этот шаг может занять много часов. Чтобы ускорить растворение мочевины, добавьте небольшую аликвоту воды, не выходя за пределы желаемого конечного объема. - Снимите стержень мешалки. Перелейте раствор в цилиндр и добавьте воды до точного конечного объема 500 мл.
- Настройка гелевого аппарата
- Очистите две пластины (одну с насечкой и одну без насечки) 70% этиловым спиртом, дайте им высохнуть, а затем обработайте пластины раствором диметилдихлорсилана.
ПРИМЕЧАНИЕ: Силанизацию можно пропустить, хотя она помогает высвобождению геля из одной из пластин при разборке гелевого сэндвича.
ВНИМАНИЕ: Работайте с раствором силанизации в перчатках и проводите обработку пластин этим раствором в вытяжном шкафу. - Поместите распорки 0,4 мм вдоль длинных краев длинной пластины, поместите короткую пластину поверх другой и совместите две пластины внизу.
- Наклейте несколько слоев бумажной ленты по всем краям, кроме верхней.
- Чтобы избежать протечек при отливке, добавьте дополнительный слой скотча на дно геля.
- Закрепите боковые стороны стеклянного сэндвича чистыми зажимами, следуя инструкциям поставщика (разные поставщики используют немного разные приспособления, зажимы для сэндвичей и прокладки).
- Очистите две пластины (одну с насечкой и одну без насечки) 70% этиловым спиртом, дайте им высохнуть, а затем обработайте пластины раствором диметилдихлорсилана.
- Заливка геля
ПРИМЕЧАНИЕ: Заливайте гель при RT (25 °C), так как полимеризация полиакриламида чувствительна к температуре.- Непосредственно перед применением в стакан наливают 80 мл заранее приготовленного денатурирующего раствора полиакриламида (шаг 4.1), 450 мкл 10% раствора персульфата аммония (APS) и 45 мкл тетраметилэтилендиамина (TEMED).
- Смешайте раствор и быстро вылейте его между стеклянными пластинами с помощью шприца объемом 50 мл. Введите расческу с нужным количеством углублений между стеклянными пластинами, избегая пузырей. При необходимости добавьте гелевый раствор, чтобы заполнить бутерброд полностью. Поместите четыре зажима на гребень, чтобы прижать и обеспечить равномерное распределение лунок.
- Дайте гелю полимеризоваться не менее 45 минут.
- Запуск геля
- После полимеризации снимите все зажимы и слои бумажной ленты. Медленно снимите гребень и тщательно промойте лунки дистиллированной водой.
- Следуйте инструкциям конкретного поставщика, чтобы правильно поместить гелевый сэндвич в вертикальный аппарат гель-электрофореза.
- Приготовьте проточный буфер для клещевого КЛЕФА (1x: 89 мМ Tris-HCl, 89 мМ борат, 2 мМ ЭДТА, pH 8) в деионизированной воде и заполните буфером верхний и нижний резервуары.
- Разогрейте пластины, выполнив предварительный прогон гель-электрофореза не менее 30 мин при мощности 50 Вт.
- Приготовьте денатурирующий гелевый загрузочный буфер (DGLB: 1 M Tris-HCl, 80% формамид, 50% глицерин, 0,05% бромфеноловый синий) в сверхчистой воде.
ПРИМЕЧАНИЕ: GLB помогает отслеживать движение образцов олигонуклеотидов в гелевую систему и позволяет загружать образцы в лунки геля. Присутствие денатурирующего агента формамида позволяет разделять виды ДНК в соответствии с размером, даже в неденатурирующем PAGE. - Суспендант высушенных образцов (образцы из этапа 3.8) в 5 мкл ГЛБ.
- Перед загрузкой образцов очистите лунки с помощью небольшого шприца и буфера КЛЕЩ в верхней буферной камере, чтобы удалить мочевину из лунок.
ПРИМЕЧАНИЕ: Повторите этот шаг несколько раз, чтобы точно очистить лунки, тем самым избегая полос, которые трудно интерпретировать. - Загрузите образцы в чистые лунки и запишите порядок загрузки.
- Выполняйте гель-электрофорез в течение 2 ч при мощности 50 Вт или, по крайней мере, до тех пор, пока бромфеноловый синий краситель не заполнит гель на 2/3.
- Визуализация геля
- После электрофореза отключите питание, снимите стеклянный сэндвич и очистите стаканы.
- Обнаружение флуоресценции олигонуклеотидных полос, меченных FAM, путем сканирования с помощью гелевого тепловизора.
Результаты
На рисунке 2 показан репрезентативный результат проведенного химического картирования, как описано в протоколе, с B-CeP 1 на олигонуклеотиде TBA, свернутом в три различные структуры. TG-квадруплексное расположение TBA (G4-TBA) было получено путем сворачивания олигонуклеотида в BP...
Обсуждение
G-квадруплексы представляют собой вторичные структуры нуклеиновых кислот, которые обычно сворачиваются в богатых гуанином последовательностях ДНК и являются важными объектами исследований из-за их связи с генетическим контролем и заболеваниями. Химическое картирование с помощью B-CeP...
Раскрытие информации
У авторов нет конфликта интересов, который можно было бы раскрыть.
Благодарности
Эта работа была поддержана Департаментом фармацевтических и фармакологических наук Университета Падуи (PRIDJ-BIRD2019).
Материалы
| Name | Company | Catalog Number | Comments |
| Acrylamide/bis-acrylamide solution 40% | Applichem | A3658 | R45-46-20/21-25-36/38-43-48/23/ 24/25-62 |
| Ammonium per-sulfate (APS) | Sigma Aldrich | A7460 | |
| Analytical balance | Mettler Toledo | ||
| Autoclave | pbi international | ||
| Boric acid | Sigma Aldrich | B0252 | |
| Bromophenol blue Brilliant blue R | Sigma Aldrich | B0149 | |
| di-Sodium hydrogen phosphate dodecahydrate | Fluka | 71649 | |
| DMSO | Sigma Aldrich | 276855 | |
| DNA oligonucleotides | Integrated DNA Technologies | synthesis of custom sequences | |
| EDTA disodium | Sigma Aldrich | E5134 | |
| Formamide | Fluka | 40248 | H351-360D-373 |
| Gel imager | GE Healtcare | STORM B40 | |
| Glycerol | Sigma Aldrich | G5516 | |
| Micro tubes 0.5 mL | Sarstedt | 72.704 | |
| Potassium Chloride | Sigma Aldrich | P9541 | |
| Sequencing apparatus | Biometra | Model S2 | |
| Silanization solution I | Fluka | 85126 | H225, 314, 318, 336, 304, 400, 410 |
| Sodium phosphate monobasic | Carlo Erba | 480086 | |
| Speedvac concentrator | Thermo Scientific | Savant DNA 120 | |
| TEMED | Fluka | 87689 | R11-21/22-23-34 |
| Tris-HCl | MERCK | 1.08387.2500 | |
| Urea | Sigma Aldrich | 51456 | |
| UV-Vis spectrophotometer | Thermo Scientific | Nanodrop 1000 |
Ссылки
- Davis, J. T. G-quartets 40 years later: from 5'-GMP to molecular biology and supramolecular chemistry. Angewandte Chemie. 43 (6), 668-698 (2004).
- Varshney, D., Spiegel, J., Zyner, K., Tannahill, D., Balasubramanian, S. The regulation and functions of DNA and RNA G-quadruplexes. Nature Reviews Molecular Cell Biology. 21 (8), 459-474 (2020).
- Chambers, V. S., et al. High-throughput sequencing of DNA G-quadruplex structures in the human genome. Nature Biotechnology. 33 (8), 877-881 (2015).
- Zuravka, I., Sosic, A., Gatto, B., Gottlich, R. Synthesis and evaluation of a bis-3-chloropiperidine derivative incorporating an anthraquinone pharmacophore. Bioorganic & Medicinal Chemistry Letters. 25 (20), 4606-4609 (2015).
- Zuravka, I., Roesmann, R., Sosic, A., Gottlich, R., Gatto, B. Bis-3-chloropiperidines containing bridging lysine linkers: Influence of side chain structure on DNA alkylating activity. Bioorganic & Medicinal Chemistry. 23 (6), 1241-1250 (2015).
- Zuravka, I., et al. Synthesis and DNA cleavage activity of bis-3-chloropiperidines as alkylating agents. ChemMedChem. 9 (9), 2178-2185 (2014).
- Sosic, A., Gottlich, R., Fabris, D., Gatto, B. B-CePs as cross-linking probes for the investigation of RNA higher-order structure. Nucleic Acids Research. 49 (12), 6660-6672 (2021).
- Sosic, A., et al. Bis-3-chloropiperidines targeting TAR RNA as a novel strategy to impair the HIV-1 nucleocapsid protein. Molecules. 26 (7), 1874 (2021).
- Sosic, A., et al. In vitro evaluation of bis-3-chloropiperidines as RNA modulators targeting TAR and TAR-protein interaction. International Journal of Molecular Sciences. 23 (2), 582 (2022).
- Sosic, A., et al. Direct and topoisomerase II mediated DNA damage by bis-3-chloropiperidines: The importance of being an earnest G. ChemMedChem. 12 (17), 1471-1479 (2017).
- Bock, L. C., Griffin, L. C., Latham, J. A., Vermaas, E. H., Toole, J. J. Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature. 355 (6360), 564-566 (1992).
- Paborsky, L. R., McCurdy, S. N., Griffin, L. C., Toole, J. J., Leung, L. L. The single-stranded DNA aptamer-binding site of human thrombin. The Journal of Biological Chemistry. 268 (28), 20808-20811 (1993).
- Carraro, C., et al. Behind the mirror: chirality tunes the reactivity and cytotoxicity of chloropiperidines as potential anticancer agents. ACS Medicinal Chemistry Letters. 10 (4), 552-557 (2019).
- Carraro, C., et al. Appended aromatic moieties in flexible bis-3-chloropiperidines confer tropism against pancreatic cancer cells. ChemMedChem. 16 (5), 860-868 (2021).
- Kypr, J., Kejnovska, I., Renciuk, D., Vorlickova, M. Circular dichroism and conformational polymorphism of DNA. Nucleic Acids Research. 37 (6), 1713-1725 (2009).
- Onel, B., Wu, G., Sun, D., Lin, C., Yang, D. Electrophoretic mobility shift assay and dimethyl sulfate footprinting for characterization of G-quadruplexes and G-quadruplex-protein complexes. Methods in Molecular Biology. 2035, 201-222 (2019).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены