Method Article
Полностью оптический механобиологический опрос Yes-ассоциированного белка в раковых и нормальных клетках человека с использованием многофункциональной системы
* Эти авторы внесли равный вклад
В этой статье
Резюме
В этой статье представлен подробный пошаговый протокол о том, как использовать интегрированную многофункциональную и программируемую пользователем систему, которая обеспечивает автоматическую многоканальную визуализацию и механобиологический анализ для выяснения механочувствительности Yes-ассоциированного белка (YAP).
Аннотация
Долгосрочная многофункциональная визуализация и анализ живых клеток требуют упорядоченной функциональной координации различных аппаратных и программных платформ. Однако ручное управление различным оборудованием, выпускаемым разными производителями, является трудоемким и трудоемким, что потенциально снижает точность, воспроизводимость и качество получаемых данных. Таким образом, универсальная и программируемая пользователем система, которая обеспечивает автоматическое, многофункциональное и долгосрочное получение изображений и совместима с большинством платформ флуоресцентной микроскопии, может принести пользу научному сообществу. В этой статье представлены полные операционные протоколы использования новой интегрированной программной системы, состоящей из (1) собственной программной программы под названием «Автоматическая многофункциональная программа интеграции (AMFIP)», которая обеспечивает автоматическое многоканальное получение изображений, и (2) набора пакетов количественного анализа изображений и вычислений клеточной тяги.
Эта интегрированная система применяется для выявления ранее неизвестной взаимосвязи между пространственно-временным распределением механочувствительного Yes-ассоциированного белка (YAP) и клеточной механикой, включая распространение и вытяжение клеток, в нормальных клетках человека (B2B), спроектированных CRISPR / Cas9, и клетках рака легких (PC9). Используя возможности этой системы многоканального контроля и считывания, результат показывает: (1) нормальные клетки B2B и раковые клетки PC9 демонстрируют четкую связь между экспрессией YAP, тракцией и динамикой клеток во время процессов распространения и миграции клеток; и (2) раковые клетки PC9 оказывают заметные периядерные силы на субстраты. Таким образом, в этом документе представлен подробный пошаговый протокол о том, как использовать интегрированную программируемую пользователем систему, которая позволяет автоматически создавать многофункциональные изображения и анализировать для выяснения механочувствительности YAP. Эти инструменты открывают возможность для детального исследования многогранной динамики сигнализации в контексте физиологии и патологии клеток.
Введение
Общая цель этого метода заключается в том, чтобы обеспечить полностью оптическую многофункциональную визуализацию и анализ живых клеток. Универсальная программа визуализации, которая обеспечивает автоматическую координацию многофункциональных оптоэлектронных устройств, сократит трудоемкие и подверженные ошибкам ручные операции и имеет важное значение для исследователей для проведения долгосрочной визуализации живых клеток1,2,3,4. Однако большинство существующих государственных программ в биомедицинском исследовательском сообществе либо применяются только к ограниченным оптоэлектронным устройствам, либо требуют дополнительного оборудования для координации различного оборудования5,6,7,8,9. Недавно была разработана программа с открытым исходным кодом и программное обеспечение под названием «Автоматическая многофункциональная программа интеграции (AMFIP)», позволяющая создавать многоканальные и покадровые изображения. Основанный на языке Java и интерфейсе прикладного программирования (API) μManager11,12, AMFIP был разработан как плагин в μManager, который выполняет настраиваемые Java-скрипты для выполнения программных коммуникаций нескольких оптоэлектронных аппаратных и программных платформ, включая, но не ограничиваясь, от Nikon. Создание AMFIP открывает возможность для программируемого и многофункционального опроса поведения клеток. В этой статье разработана интегрированная экспериментальная и вычислительная система, сочетающая AMFIP с цифровым анализом изображений и силовой микроскопией тяги клеток. Система позволяет прояснить различные метанобиологии YAP в клеточных линиях нормального B2B (рисунок 1) и рака легких PC9 (рисунок 2), спроектированного CRISPR / Cas9. Система предоставляет научному сообществу комплексное решение, которое позволяет избежать необходимости приобретения дополнительных управляющих устройств, которые могут быть недоступны и/или совместимы с каждой системой визуализации.
Протоколы, представленные в этой статье, представляют, как (1) применять AMFIP для проведения автоматической долгосрочной визуализации для обеих клеточных линий, спроектированных CRISPR / Cas9, которые экспрессируют YAP с тегами mNEonGreen2; и (2) объединить Fiji ImageJ, MATLAB и Origin для количественного анализа соотношения YAP ядерной/цитоплазмы (N/C) на основе их флуоресцентной интенсивности (рисунок 3 и рисунок 4), поля клеточного смещения (рисунок 1C и рисунок 2C) и поля клеточного тяги (рисунок 1D и рисунок 2D) ). Результаты показывают, что (1) в течение первых 10 ч распространения клеток на субстратах, которые имеют физиологически значимую механическую жесткость13,14,15,16,17,18, соотношение YAP N/C отдельных B2B-клеток показывает более заметные зависящие от времени вариации и флуктуации по сравнению с одиночными клетками PC9 (рисунок 5 и рисунок 6). ); и (2) раковые клетки PC9 генерируют заметную тягу в своих околоядерных областях (рисунок 7). Интегрированная система и методологии, описанные в этом протоколе, выходят за рамки конкретных типов клеток и оптогенетических молекул. Исследователи могут применять протоколы для настройки своих конкретных экспериментов по опросу живых клеток и выяснения многогранной динамики передачи сигналов в контексте физиологии и патологии клеток.
протокол
1. Генерация стабильной CRISPR/Cas9-отредактированной клеточной линии рака легких человека (PC9) и бронхиальной эпителиальной клеточной линии человека (Beas2B), которые эндогенно экспрессируют белок YAP с меткой mNeonGreen21-10/11
- Выполняют полимеразную цепную реакцию (ПЦР) для амплификации последовательности ДНК, кодирующей 11-ю цепь флуоресцентного белка, mNeonGreen2, используя высокоточную ДНК-полимеразу (см. Таблицу материалов).
- Вбивание амплифицированной последовательности ДНК в геномный локус YAP клеточных линий PC9 и B2B с помощью системы редактирования генов CRISPR-Cas9.
ПРИМЕЧАНИЕ: Эта последовательность ДНК дополняет нити 1-10 mNeonGreen2 для испускания флуоресценции. Карта геномных последовательностей YAP-mNeonGreen21-10/11 показана на дополнительном рисунке S1. Карта содержит помеченные геномные, донорские и mNeonGreen2 последовательности. - Проверьте экспрессию mNeonGreen2, разработанную CRISPR/Cas9, с помощью эпифлуоресцентного микроскопа (см. Таблицу материалов). Поскольку mNeonGreen2 помечается YAP всякий раз, когда клетки экспрессируют YAP в контексте своей собственной регуляторной сети генов, проверьте наличие интенсивности флуоресценции в обеих клетках, спроектированных CRISPR / Cas9, и сравните ее с таковой в родительских клетках (контроль).
ПРИМЕЧАНИЕ: Чтобы следовать этому протоколу, используйте (1) лазер 488 нм (47,5 мВт/мм2) для возбуждения, (2) 40-кратный объектив (числовая апертура (NA) = 0,95) и полосовой эмиссионный фильтр (ET525/50 нм) для измерения флуоресценции и (3) программное обеспечение ImageJ для измерения, количественной оценки и сравнения интенсивностей флуоресценции. - Подтвердить правильную интеграцию mNeonGreen211 путем извлечения геномной ДНК из клеточных линий, отредактированных CRISPR/Cas9; выполнять ПЦР с помощью праймеров, фланкирующих геномную вставку и секвенирование для подтверждения введения в правильных геномных локусах19,20.
- Сбивайте mNeonGreen211 с помощью системы редактирования генов CRISPR/Cas9 и проверяйте снижение интенсивности флуоресценции в клетках с помощью тех же систем микроскопа и параметров визуализации, описанных на шаге 1.3.
ПРИМЕЧАНИЕ: Этот шаг подтверждает правильное интегрирование mNeonGreen211 путем сравнения интенсивностей флуоресценции. В качестве контроля используются клетки, спроектированные CRISPR/Cas9, без нокдауна и родительские клетки. - Соберите клетки с помеченным интересующим белком с помощью флуоресцентно-активированной сортировки клеток (FACS).
- Чтобы подготовить клетки к сортировке FACS, попробуйте их и повторно суспендируйте в фосфатно-буферном физиологическом растворе (PBS).
- Соберите клетки с флуоресценцией mNeonGreen2 выше фонового уровня родительских клеточных линий в два обогащающих раунда сортировки FACS.
ПРИМЕЧАНИЕ: Сроки генерации crispr/Cas9-отредактированных клеточных линий, описанных здесь, составляют порядка 1-2 месяцев. Все клеточные линии становятся общедоступными по запросу, чтобы другие исследовательские лаборатории могли воспроизвести результаты.
2. Обслуживание pc9 и B2B ячеек
- Поддерживать обе клеточные линии в увлажненных тканевых инкубаторах с 5% CO2 при 37 °C.
- Культивирование 106 эндогенно меченых клеток PC9 и Beas2B в колбах объемом 75 см2 с 12 мл среды RPMI-1640 с добавлением 10% фетальной бычьей сыворотки и 100 мкг/мл пенициллина-стрептомицина. Субкультура обеих клеточных линий, когда слияние клеток достигает ~80%.
- Тестируйте обе клеточные линии на микоплазму каждые 3 месяца, используя набор для обнаружения микоплазмы, строго следуя всем рекомендуемым производителем протоколам.
- Храните клеточные линии в морозильной камере при температуре -80 °C.
- Используйте клеточные линии, которые <20 проходов от оттепели для всех экспериментов.
3. Настройка аппаратно-программной среды
- Настройка аппаратной среды эксперимента
- Подключите конфокальный контроллер и инвертированный микроскоп к компьютеру (см. Таблицу материалов).
- Установите программную платформу (Таблица материалов).
- Включите конфокальный контроллер и перевернутый микроскоп. Затем запустите Элементы.
- Откройте панели управления конфокального, лазерного и инвертированного микроскопа в Elements. Затем проверьте, правильно ли функционируют три панели, проверив движение моторизованной ступени, переключение целей микроскопа и пространственное сканирование лазерных линий.
- Настройка программной среды AMFIP
- Установите на компьютер IntelliJ, Java Development Kit 14.0, μManager версии 2.0 gamma и Fiji ImageJ.
- Откройте проект AMFIP, загруженный с GitHub (ссылка: https://github.com/njheadshotz/AMFIP) в IntelliJ.
- Нажмите на настройки | | компилятора Обработчики аннотаций и установите флажок Включить обработку аннотаций.
- Нажмите на Структуру проекта | Артефакты и создание JAR-файла. Задайте для выходного каталога значение mmplugins в каталоге μManager .
- Нажмите на Структуру проекта | Библиотеки и добавляйте mmplugins и плагины в каталог μManager .
- Нажмите добавить конфигурацию в раскрывающемся меню Выполнить и создайте приложение.
- Введите ij. ImageJ в класс Main.
- Введите -Xmx3000m -Dforce.annotation.index=true в параметр виртуальной машине.
- Задайте для каталога μManager значение Каталог Work.
- Нажмите кнопку Выполнить , чтобы активировать μManager с помощью плагина AMFIP.
- Подключите μManager к инвертированному микроскопу.
- Добавьте адаптивный драйвер инвертированного микроскопа21 в каталог μManager .
- Открыть μManager. Нажмите на устройства | Мастер настройки оборудования и создайте новую конфигурацию.
- Добавьте драйвер Ti2 в раздел Доступные устройства.
- Выберите все периферийные устройства и сохраните новый файл конфигурации.
- Перезапустите μManager и выберите файл конфигурации на шаге 3.2.4 в конфигурации запуска Micro-Manager.
4. Приготовление геля
- Обработать стеклянную крышку 3-аминопропилтриметоксисиланом в течение 7 мин при комнатной температуре (24 °C).
- Используйте деионизированную (DI) воду, чтобы промыть стеклянную крышку и высушить крышку в течение 20 минут при 160 °C.
- Обработайте стеклянную крышку 0,5% глутаровым альдегидом в течение 30 минут и промойте водой DI.
- Смешайте раствор акриламида, раствор N,N'-метиленбисакламида (бис) и флуоресцентные шарики, суспендированные в 10 мМ HEPES-буферного физиологического раствора. Используют 10% (мас./об.) раствор персульфата аммония и N,N,N',N'-тетраметилэтилендиамин (TEMED) в качестве инициаторов полимеризации. Изменять процентное содержание каждого компонента для достижения желаемой механической жесткости гидрогелей полиакриламида (ПАА) в соответствии с установленными протоколами, описанными ранее13,14.
ПРИМЕЧАНИЕ: В настоящем протоколе 2 кПа геля: акриламид = 12,5% и бис-акриламид = 6,5%; 5 кПа гель: акриламид = 12,5% и бис-акриламид = 21,5%; и гель 40 кПа: акриламид = 12,5% и бис-акриламид = 31,5%. Все перечисленные % являются процентом объема. - Через 35 мин дважды (по 5 мин) очистите стеклянную крышку от затвердевшего гидрогеля PAA и промыть гидрогель 50 мМ hePES-буферным физиологическим раствором дважды (по 5 мин каждый раз).
- Обработать поверхность гидрогеля гидразин-гидратным раствором в течение 6 ч.
- Промыть гидрогель уксусной кислотой в течение 30 мин. Удалите уксусную кислоту и промойте PBS в течение 30 минут.
- Окисляют раствор фибронектина (50 мкг/мл в PBS) периодатом натрия в течение 30 мин.
- Поверхность гидрогеля покрыть окисленным раствором фибронектина и подождать 35 мин.
- Добавьте PBS, чтобы погрузить гидрогель и хранить при температуре 4 °C. Покройте все чашки Петри, содержащие гидрогели, алюминиевой фольгой, чтобы избежать воздействия света на гидрогели.
5. Клеточная культура
ПРИМЕЧАНИЕ: Выполняют культивирование клеток с использованием асептической техники.
- Свяжите стеклянные крышки с гидрогелями PAA с чашкой Петри со стеклянным дном 35 мм, чтобы избежать физического дрейфа гелей во время процессов посева клеток и визуализации.
- Используя стерилизованный чистый пинцет, поднимите крышку (с гидрогелем PAA сверху) из чашки Петри, содержащей приготовленные гели.
- Используйте сухую салфетку, чтобы очистить капли воды на нижней поверхности стеклянной крышки.
- Используйте стерилизованный пинцет для удержания стеклянной крышки.
- Поместите небольшие капли (1-5 мкл) цианоакрилатного клея в два диагональных угла на нижней поверхности.
- Используйте стерилизованные салфетки, чтобы удалить лишний клей.
- Используйте стерилизованный пинцет для замены крышки в чашке Петри со стеклянным дном. Слегка прижмите углы крышки, чтобы капли клея полностью соприкасались с поверхностью чашки Петри.
- Поместите крышку обратно на чашку Петри, чтобы свести к минимуму испарение PBS в гидрогелях PAA. Подождите 3 мин, чтобы клей затвердел и высох в чашке Петри.
- Наполните чашку Петри 4 мл PBS.
- Повторите вышеуказанные шаги 5.1.1-5.1.8 для остальных образцов гидрогеля ЧУК в чашках Петри, используемых для визуализации.
- Используйте 75% этанол для стерилизации наружной поверхности всех чашек Петри и переноса их в кабинет биобезопасности культуры тканей. Включите ультрафиолетовый свет на 5 минут и стерилизуйте образцы.
- Посейте клетки на верхнюю поверхность геля.
- Выключите ультрафиолетовый свет. Выньте колбу (содержащую ячейки B2B/PC9) из инкубатора с температурой 37 °C в шкаф биобезопасности. Используйте пипетку, подключенную к вакуумному насосу, чтобы аспирировать всю питательную среду и добавить 5 мл PBS для промывки колбы.
- Добавьте 2 мл 0,05% трипсина, чтобы отделить клетки от нижней части колбы.
- Поместите колбу в инкубатор. Подождите 5 мин.
- Перенесите колбу в шкаф биобезопасности. Добавьте 8 мл свежей питательной среды в колбу и пипетку вверх и вниз несколько раз, чтобы подвешивать клетки однородно.
- Переложите все 10 мл клеточной суспензии в трубку 15 мл и центрифугу при 300 × г в течение 5 мин.
- Проверьте ячейку гранулы в нижней части трубки. Медленно наклоните трубку горизонтально и используйте аспирационную пипетку, чтобы удалить всю культуральную среду из трубки, не касаясь гранулы клетки. Затем добавьте 8 мл свежей питательной среды и пипетку вверх и вниз несколько раз, пока все клетки не смешатся однородно со средой.
- Нанесите 100 мкл клеточной суспензии (150 клеток/мкл) на поверхность геля и подождите 5 минут. Затем медленно добавить 4 мл свежей питательной среды в чашку Петри; избегайте добавления свежей среды непосредственно на гель.
- Поместите чашку Петри в инкубатор при температуре 37 °C. Подождите, чтобы позволить клеткам прикрепиться к поверхности геля (B2B: 0,5-1 ч; ПК9: 4-5 ч).
6. Визуализация клеток
ПРИМЕЧАНИЕ: AMFIP обеспечивает автоматическую, многоканальную и долгосрочную визуализацию путем координации с различными аппаратными и программными системами: (1) AMFIP манипулирует μManager для автоматического перемещения моторизованной ступени микроскопа Ti2-E в несколько полей зрения (FOV) и получения изображений яркого поля через монохромную камеру (Таблица материалов); и (2) AMFIP активирует несколько файлов макросов внутри Elements с помощью настраиваемого Java-скрипта для выполнения автоматических операций для конфокальной z-стековой визуализации и переключения различных лазерных каналов (405 нм и 488 нм).
- Настройка среды для долгосрочной визуализации.
- Поместите камеру окружающей среды на моторизованную ступень перевернутого микроскопа. Установите скорость потока CO2 на 160 мл/мин и отрегулируйте температуру камеры (сверху: 44 °C; ванна: 42 °C; ступень: 40 °C). Затем добавьте 40 мл очищенной воды в ванну камеры.
- Выньте чашку Петри со стеклянным дном с клетками из инкубатора и поместите ее в камеру окружающей среды.
- Включите конфокальный контроллер и перевернутый микроскоп. Переключите световой контур вправо и наблюдайте за присоединением ячеек с помощью μManager. Если к гелю прикреплено достаточное количество клеток, перенесите чашку Петри обратно в инкубатор. Если к гелю прикреплено недостаточно клеток, продолжайте инкубацию клеток еще 30 минут для B2B и 60 минут для клеток PC9.
- Отрежьте два небольших кусочка скотча и наклейте их на камеру вокруг круглого отверстия. Далее нанесите немного клеевого клея на ленту (только на тот участок, который покроет чашка Петри).
- Достаньте чашку Петри из инкубатора. Затем медленно поместите чашку Петри в камеру и дайте дну чашки соприкоснуться с клеем.
- Прижмите крышку чашки Петри в течение 1 мин, чтобы клей полностью соприкоснулся с чашкой Петри и затвердел. Затем осторожно надавите на чашку Петри горизонтально, чтобы подтвердить, что чашка Петри неподвижна в камере.
- Закройте крышку камеры.
- Задайте параметры получения изображения для визуализации яркого поля.
- Откройте IntelliJ и задайте параметр T1 (например, 120 с) в строке 93 файла Elements_script.java. Убедитесь, что это значение больше времени выполнения макроса в Elements, используемого для конфокальной визуализации одного поля зрения (FOV). Нажмите кнопку Выполнить , чтобы запустить проект AMFIP IntelliJ.
- Нажмите кнопку Live и Multi-D Acq. на главном интерфейсе μManager. Затем переключите световой путь инвертированного микроскопа вправо для визуализации яркого поля, переключитесь на объектив 10x и откройте светодиодный (LED) свет (источник света для визуализации яркого поля; интенсивность: 5%).
- Нажмите на световой контур, объектив микроскопа и кнопку светодиодной лампы в панели Elements Ti2 или вручную нажмите соответствующие кнопки на микроскопе.
- Отрегулируйте джойстик XY и ручку Z-плоскости, чтобы найти правильное положение и плоскость фокусировки геля на чашке Петри. Используйте 10-кратный объектив, чтобы найти соответствующие FOV нескольких одиночных клеток, прикрепленных к гелю.
- Установите флажок Несколько позиций (XY) в окне Многомерное получение . Нажмите кнопку «Редактировать список позиций...» и обратите внимание на всплывающее окно «Список позиций рабочей области ». Затем измените цель на 40x, увеличьте интенсивность светодиодного света до 15%, повторно отрегулируйте XY-моторизованный этап, чтобы найти FOV, и запишите координаты, нажав кнопку Mark в окне Список позиций сцены .
- Запишите 67 желаемых FOV. Нажмите кнопку Сохранить как... в окне Список позиций рабочей области , чтобы записать координаты. Введите T1 (параметр, например, 120 с, определенный на шаге 6.2.1) во временной интервал получения изображения до T1 в разделе «Временные точки» в окне «Многомерное получение ».
- Настройка получения изображений для 2D-YAP и изображений бусин.
- Откройте Элементы, измените траекторию света вправо для конфокального изображения и выключите светодиодный свет. Затем нажмите кнопку «Удалить блокировку» и включите лазерный канал FITC (для визуализации YAP), установив флажок FITC .
- Отрегулируйте скорость сканирования до 1 кадра на 2 с, нажав кнопку 1/2 и поверните ручку Z-плоскости, чтобы быстро найти Z-положение прикрепленных ячеек. Запишите нижний и верхний пределы для Z-стека.
- Щелкните Макрос на верхней ленте, выберите Редактор макросов в раскрывающемся меню Макрос и введите значения из шага 6.3.2 в файл макроса.
- Включите лазерный канал 4',6-диамидино-2-фенилиндол (DAPI) (для визуализации шариков), установив флажок DAPI , чтобы найти и записать сфокусированное Z-положение бусин. Перейдите в редактор макросов и введите записанные значения в файл макроса.
- Поставьте задачу перемещения моторизованной ступени с помощью AMFIP.
- Перейдите в μManager и нажмите на Плагины | Автоматизация для открытия графического интерфейса пользователя (GUI) AMFIP. Нажмите кнопки «Добавить точку» или «Удалить точку», чтобы получить точное количество выбранных FOV. Введите записанные координаты FOV в панель координат.
- Определите общее время эксперимента в текстовом поле Общее время эксперимента .
- Нажмите кнопку «Дополнительная конфигурация времени» и определите временной интервал T2 (например, 30 минут) перемещения моторизованной ступени к каждому FOV.
- Разверните размер окна Elements и перетащите графический интерфейс AMFIP в правую часть экрана, чтобы графический интерфейс не мешал автоматическим операциям курсора.
- Нажмите на кнопку Enter . После завершения первого макроса нажмите кнопку «Получить!» в окне «Многомерное получение ».
- Растворите клетки после получения изображения.
- После завершения долгосрочной визуализации остановите задачу AMFIP, нажав кнопку Пауза в окне плагина автоматизации и кнопку Стоп в окне Многомерное получение .
- Откройте Elements и задайте Z-стек изображений, нажав кнопки Top и Bottom в окне ND Acquisition (установите Z-диапазон больше, чем Z-диапазон бусин). Переключите световой путь вправо и откройте светодиодный индикатор (интенсивность: 15%).
- Медленно и осторожно снимите крышки камеры и чашки Петри. Между тем, следите за видом яркого поля для любого дрейфа FOV.
- Используя пластиковую пипетку, чтобы взять 0,5 мл раствора додецилсульфата натрия (SDS), осторожно подержите пластиковую пипетку немного выше питательной среды в чашке Петри и добавьте 1-2 капли раствора SDS в культуральную среду.
- Как только ячейки в виде яркого поля растворятся, переключите световой контур влево, закройте светодиодный индикатор, нажмите кнопку «Удалить блокировку ».
- Запустите образ Z-стека. Сохраните стек изображений и назовите его Reference_N (N — порядковый номер каждого FOV).
- Нажмите кнопку «Несколько позиций» (XY) в окне «Многомерное получение ». Затем выберите следующий FOV и нажмите кнопку Go to, чтобы переместить моторизованный этап на второй FOV.
- Повторите шаг 6.5.7 для каждого FOV.
7. Измерение соотношения YAP N/C
- Выполните анализ изображений для измерения соотношения YAP N/C с помощью программного обеспечения Fiji ImageJ (рисунок 4).
- Открыть Фиджи ImageJ. Импортируйте стек изображений яркого поля для всех FOV, полученных μManager.
- Откройте раскрывающееся меню Изображение и выберите Стеки | Инструменты | Хранитель срезов. Затем экспортируйте стек изображений яркого поля для каждого FOV.
- Импортируйте флуоресцентное изображение канала FITC и наложите на него изображение яркого поля для того же FOV. Для этого выберите флуоресцентное изображение и выберите Наложение | Добавить изображение... (Изображение для добавления: изображение яркого поля; Расположение X и Y зависит от размера изображения яркого поля, полученного различными камерами; Непрозрачность: 60-70).
- Откройте раскрывающееся меню Анализ и выберите Установить измерения.... Выберите Область; Интегральная плотность и среднее значение серого.
- Нажмите на кнопку «Выбор от руки » на главном интерфейсе ImageJ.
- Нарисуйте контур тела клетки и желаемого ядра. Затем нажмите « Анализировать | Измерьте или нажмите кнопку M на клавиатуре.
- Обратите внимание на всплывающее окно Результаты . Обратите внимание, что значения в столбце Площадь представляют площадь выбранной области (мкм2), а значения в столбце IntDen представляют интенсивность флуоресценции выбранной области.
- Рассчитайте соотношение YAP N/C, используя следующие формулы (1), (2) и (3):
 (1)
(1) (2)
(2) (3)
(3)
Где Inuc и Icel представляют относительную интенсивность ядра и тела клетки, а Anuc и Acel представляют площадь ядра и тела клетки. R — коэффициент YAP N/C. - Сохраните контуры для будущего расчета силы тяги диполя и периклеточного/периядерного смещения. Для этого нажмите на кнопку Анализ | Инструменты | Сохранить координаты XY...
8. Измерение тягового поля
- Применяйте тракционную микроскопию через плагины Fiji ImageJ22,23.
- Открыть Фиджи ImageJ.
- Импортируйте стек изображений бусин для FOV.
- Выберите фрагмент, который показывает наиболее четкое распределение бусин и извлеките его, нажав на Изображения | Стеки | Инструменты | Хранитель срезов.
- Импортируйте стек изображений ссылки для того же FOV.
- Выберите фрагмент с той же яркостью и контрастностью, что и фрагмент на шаге 8.1.3. Затем извлеките его в качестве эталонного изображения.
- Выберите изображения | Стеки | Инструменты | Объедините, чтобы объединить два фрагмента из шагов 8.1.3 и 8.1.5 (выберите ссылочное изображение в качестве первого фрагмента).
- Выберите плагины | | сопоставления шаблонов Выравнивание фрагментов в стеке или плагинах | Стабилизатор изображения для выравнивания двух фрагментов.
- Выберите | изображений Стеки | Стек в изображения. Затем выберите | изображений таблицы подстановки | Зеленый , чтобы преобразовать цвет первого фрагмента в зеленый, и выберите Изображение | таблицы подстановки | Красный , чтобы преобразовать цвет второго фрагмента в красный.
- Выберите | изображений Цвет | Объедините каналы , чтобы объединить два изображения.
- Перекройте изображение с изображением яркого поля из того же поля и используйте это перекрывающееся изображение для наблюдения за смещением бусин.
- Выберите плагины | PIV | итеративный PIV(Базовый).... Установите размер окна опроса равным 128/256; 64/128; 32/64 (не менее четырех бусин на окно допроса). Установите порог корреляции равным 0,6.
- Нажмите OK. После завершения вычисления сохраните текстовый файл с необработанными данными о перемещении бисера в обычную папку, созданную пользователем.
- Выберите плагины | | FTTC FTTC и выберите текстовый файл на шаге 8.1.9.
- Введите размер пикселя (мкм), модуль Юнга геля (Паскаль), а также ширину и высоту графика на основе эксперимента и изображения бусин.
- Нажмите OK , чтобы автоматически сохранить текстовый файл, содержащий необработанные данные о тяговой силе, в том же каталоге, что и текстовый файл на шаге 8.1.12.
- Используйте графическое программное обеспечение (Таблица материалов) для построения поля тяги с одинаковым масштабом для нескольких ячеек (рисунок 1B, C и рисунок 2B, C).
- Вставьте текстовый файл, содержащий необработанные данные тяги, в электронную таблицу.
- Создайте новый лист, введите координаты Y тяги в первую строку (расположите от высоких значений до низких значений) и координаты X в первый столбец (расположите от низкого к высокому).
- Введите значение тяги к каждой координате из необработанных данных.
- Сохраните лист на шаге 8.2.2 в виде *.csv файла.
- Открытое происхождение.
- Нажмите на файл | Откройте и импортируйте файл *.csv на шаге 8.2.4. Выделите все ячейки и нажмите на Plot | Контур| Контур - Цветная заливка.
- В окне Построение: plotvm выберите Y в столбцах, чтобы автоматически задать значения Y в первой строке, а значения X — в первый столбец. Затем назовите заголовок и нажмите OK.
- В появившемся окне графика дважды щелкните тепловую карту.
- Нажмите на «Уровни » в окне «Цветовая карта/Контуры ». Затем измените уровень шкалы на разумный диапазон (0300 в этом анализе) и нажмите OK.
- Нажмите на Строки, снимите флажок Показывать только на основных уровнях и установите флажок Скрыть все. Затем нажмите OK.
- Щелкните график правой кнопкой мыши и выберите Экспорт графиков.... Сохраните изображение по указанному пути.
- Используйте MATLAB для расчета тяги дипольных клеток.
- Сохраните текстовый файл необработанных данных тяги (из шага 8.1.12) и файл координат области границы ячейки (ROI) (из шага 7.1.9) в той же папке, определенной на шаге 8.1.12. Перенесите все файлы MATLAB, которые находятся в пакете AMFIP, в эту папку.
- Откройте MATLAB. Откройте папку, определенную на шаге 8.1.12, и откройте файл функции расчета дипольной тяги absdipole.m перенесен в эту папку на шаге 8.3.1.
- Прочитайте два текстовых/csv-файла на шаге 8.3.1 в рабочем пространстве MATLAB и назначьте матрицу двум переменным (например, traction и roi).
- Запустите функцию absdiple (traction,roi).
ПРИМЕЧАНИЕ: Первым столбцом выхода является сила дипольного тяги в nN (нано-Ньютон). Вторая колонка выхода представляет собой угол силы тяги диполя относительно горизонтальной оси.
Результаты
Отчетливое распределение и динамика YAP в CRISPR/Cas9-инженерном раке PC9 и нормальных клетках B2B во время распространения клеток
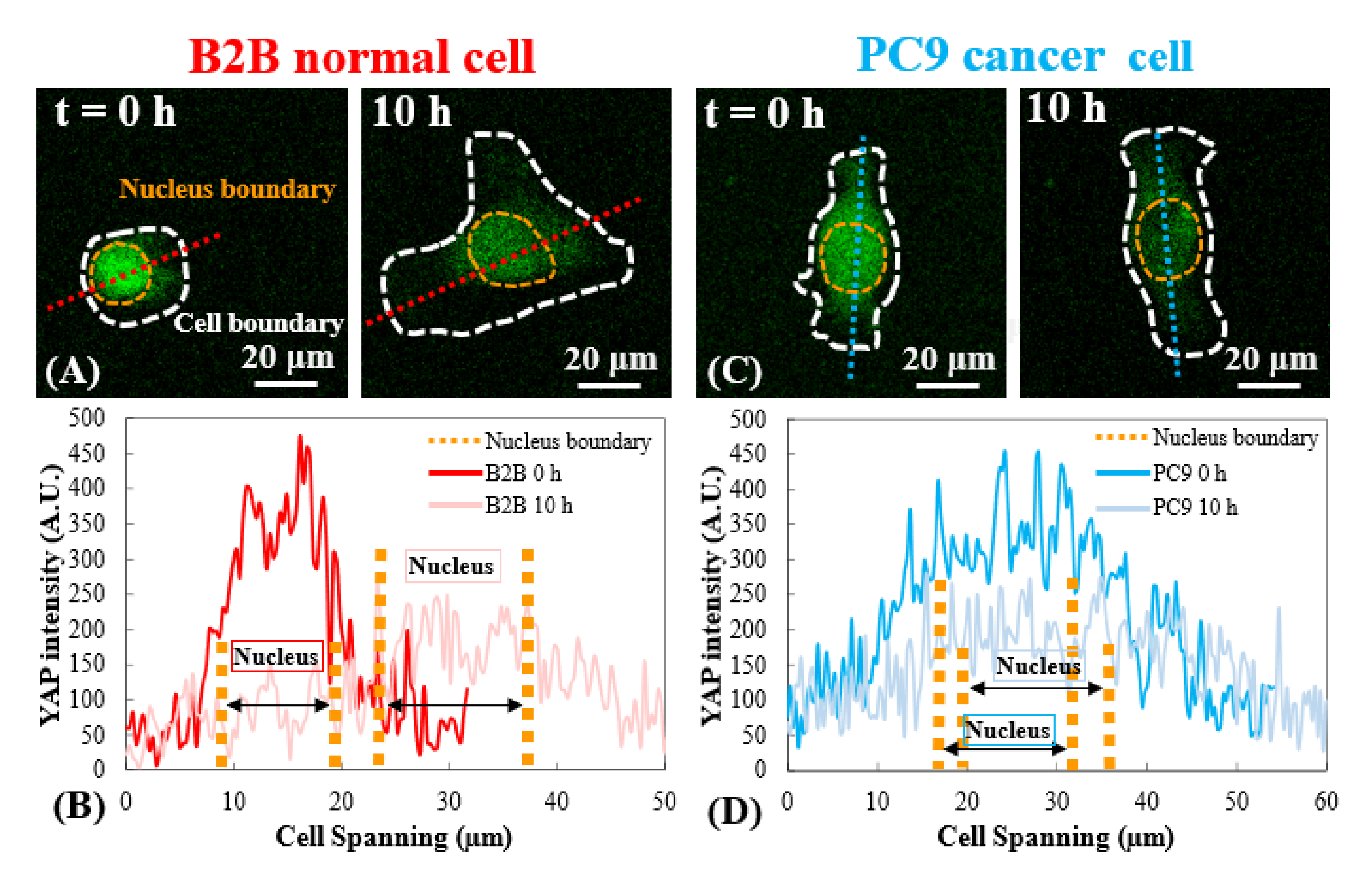
Репрезентативные флуоресцентные изображения распределения YAP в одиночных ячейках B2B и PC9 на гелях PAA 2, 5, 40 кПа и стеклянной крышке показаны на рисунках 1A и 2A. Ядерная локализация YAP в B2B-клетках увеличивалась с увеличением жесткости субстрата (рисунок 1A), тогда как клетки PC9 показали аналогичную концентрацию YAP в ядре и цитоплазме на субстратах различной жесткости (рисунок 2A). Репрезентативные флуоресцентные изображения распределения YAP в одиночных, распространяющихся ячейках B2B и PC9 на гидрогелевой подложке 5 кПа (от 0-го ч до 10-го ч после прикрепленных к подложкам клеток) показаны на Фиг.1В и Фиг.2В соответственно. Ячейка B2B монотонно увеличивала площадь распространения с течением времени вместе с уменьшением коэффициента YAP N/C (рисунок 1B), в то время как ячейка PC9 сохраняла сравнительно неизменную площадь распространения ячейки, ориентацию и соотношение YAP N/C на протяжении всего 10-часового процесса распространения (рисунок 2B). В течение 10-часовой продолжительности раннего распространения репрезентативная ячейка B2B конститутивно деформировала поверхность подложки и применяла развивающуюся во времени тягу клеток по всей площади ячейки (рисунок 1C и рисунок 1D).
Напротив, репрезентативная ячейка PC9 развивала смещение и тягу только на двух концах тела клетки, и ее тяга уменьшалась через 7,5 ч (рисунок 2C и рисунок 2D). Дополнительные покадровые изображения и измерения тяги клеток B2B и PC9 на ранней стадии распространения представлены на дополнительном рисунке S2 и дополнительном рисунке S3. Наблюдались и другие режимы динамики ячеек PC9 (рисунок 6). Параллельно с этими различными характеристиками распространения ячейки B2B и PC9 показали различное распределение и динамику YAP (рисунок 3). На геле 5 кПа YAP в клетках B2B концентрировались в ядре в 0-м ч и становились более однородно распределенными по телу клетки в 10 ч . Однако клетки PC9 показали более однородное распределение YAP в ядре и цитоплазме на протяжении всех 10 ч процесса распространения. Для количественного анализа активности и транслокации YAP в клетках B2B и PC9 соотношение YAP N/C было рассчитано с использованием алгоритма, описанного на рисунке 4.
Для дальнейшего исследования отчетливой динамики YAP сравнивали временные изменения в соотношении YAP N/C, площади клетки/ядра и тяге нескольких одиночных B2B-клеток (n = 10) и клеток PC9 (n = 5) (рисунок 5). Установлено, что среднее yaP N/C соотношение B2B клеток снизилось с 2,54 ± 0,22 до 1,79 ± 0,21 (n = 10; p = 0,0022**; Рисунок 5А), в то время как среднее отношение YAP N/C клеток PC9 изменилось с 1,92 ± 0,26 до 1,57 ± 0,07 (n = 5; p = 0,187 (незначительно (ns)); Рисунок 5А). Среднее дипольное тяговое усилие B2B-клеток изменилось с 256,17 ± 123,69 нН до 287,44 ± 99,79 нН (p = 0,7593 (нс); Рисунок 5B). Средняя дипольная тяга клеток PC9 изменилась с 141,19 ± 33,62 нН до 168,52 ± 73,01 нН (p = 0,7137 (нс); Рисунок 5B). Средняя площадь распространения клеток B2B увеличилась с 613,89 ± 102,43 мкм2 до 942,51 ± 226,71 мкм2 (p = 0,0512 (ns); Рисунок 5С).
Средняя площадь распространения клеток PC9 изменилась с 495,78 ± 97,04 мкм2 до 563,95 ± 89,92 мкм2 (p = 0,5804 (ns); Рисунок 5С). Средняя площадь распространения ядра B2B-клеток увеличилась с 181,55 ± 36,18 мкм2 до 239,38 ± 43,12 мкм2 (p = 0,1217 (ns); Рисунок 5D) а средняя площадь распространения ядра клеток PC9 изменилась со 133,31 ± 30,05 мкм2 до 151,93 ± 22,49 мкм2 (p = 0,5944 (ns); Рисунок 5D). Эти результаты свидетельствуют о том, что (1) B2B-клетки демонстрируют конститутивно зависящее от жесткости субстрата отношение YAP N/C; (2) тяга B2B-клеток выше, чем у клеток PC9; и (3) в отличие от B2B-клеток, клетки PC9 демонстрируют ограниченное увеличение площади клеток и изменения в соотношении YAP N/C в течение 10-часового процесса распространения.
Корреляция распределения и динамики YAP с состояниями миграции B2B-клеток
Сравнивали коэффициент YAP N/C и дипольное тяговое усилие всех клеток B2B (n=10) и PC9 (n=5) в зависимости от площади распространения клеток и площади распространения ядра. Коэффициент YAP N/C и дипольное вытяжение клеток PC9 не были четко коррелированы с их диапазонами площади распространения мелких клеток и ядер (рисунок 6). Напротив, коэффициент YAP N/C и дипольное тяговое сцепление B2B-клеток, по-видимому, следуют двум различным тенденциям (рисунок 6A и рисунок 6C), предполагая, что в этом эксперименте могут сосуществовать две группы B2B-клеток. В первой группе коэффициент YAP N/C и дипольная тяга увеличиваются вместе с увеличением площади распространения клеток и достигают своих максимумов при ~ 1000 мкм2 (рисунок 6C и рисунок 6D, обозначенные желтой пунктирной линией). Во второй группе коэффициент YAP N/C и дипольное тяговое усиливание увеличиваются медленнее с увеличением площади распространения клеток и поддерживают почти постоянные значения, когда площадь распространения клеток продолжает увеличиваться (рисунок 6C,D, обозначенный зеленой пунктирной линией).
Раковые клетки PC9 генерируют тяги в околоядерных областях
Одиночные, распространяющиеся клетки PC9 вытесняют субстраты в околоядерных областях, начиная с 6-го ч культуры (рисунок 7С). Чтобы визуализировать периядерное смещение, вызванное тягой клеток, мы перекрыли изображения флуоресцентных шариков, сделанные до (красный) и после (зеленый) удаления клеток из субстратов (см. раздел протокола для деталей). Бусины, которые не имеют никакого смещения, будут казаться желтыми на перекрывающихся изображениях, то есть добавлением красного и зеленого цветов. Напротив, бусины, которые смещены из положения покоя из-за тяги клеток, будут иметь разделенные зеленый и красный цвета.
Примечательно, что как в клетках PC9 (рисунок 7C, D), так и в клетках B2B (рисунок 7E) смещение шариков наблюдалось в цитоплазме и внутри ядра, в дополнение к тем, которые находятся на границе клетки. Чтобы выделить околоядерное смещение, уравнение Буссинеска из теории линейной упругости используется для прогнозирования 2D-теоретического смещения, генерируемого гипотетической дипольной силой на границе ячейки (черная пунктирная линия на рисунке 7B)24. Сравнивая эту теоретическую кривую с реальным смещением субстрата, измеренным вдоль той же оси (белая пунктирная линия на рисунке 7D), было обнаружено, что реальные смещения внутри ядра в 1,5-8 раз превышают теоретическое значение (рисунок 7B), что указывает на существование силы тяги в околоядерных областях.

Рисунок 1: Изменения экспрессии/распределения YAP, поля смещения субстрата и поля тяги нормальной ячейки B2B на субстратах различной жесткости и во время раннего распространения. (A) Экспрессия YAP клетки B2B, засеянной на 2, 5 и 40 кПа гелях PAA и стеклянном покровном листе через 60 ч от начального прикрепления клеточной подложки. (B) Клетку B2B засыпали на гель PAA 5 кПа и визуализировали в течение 10 ч после первоначального прикрепления клеточного субстрата. Экспрессия YAP представлена интенсивностью зеленой флуоресценции. Примечание: Интенсивность YAP внутри ядра постепенно уменьшается, но со временем остается выше, чем в цитоплазме. Цветовые полосы обозначают уровни выражения YAP (зеленый = высокое выражение; черный = низкое выражение) в (A) и (B). (C) Деформация субстрата (перекрывающаяся изображением яркого поля) в месте расположения ячейки представлена полем смещения в каждой точке времени. Направление смещения и величина показаны направлением стрелки и цветом соответственно. Смещение становится больше на концах тела клетки B2B по мере увеличения площади распространения клеток. Цветовая шкала указывает на величину смещения (малиновый = высокая величина; черный = низкая величина). D) Поле тяги (перекрывающееся изображением яркого поля), рассчитанное на основе поля смещения. Тяга концентрируется на границе B2B-клеток. Белые и желтые пунктирные контуры очерчивают границы клетки и ядра соответственно. Цветовая шкала указывает на величину тяги (малиновый = высокая величина; черный = низкая величина). Шкала стержней = 20 мкм. Сокращения: YAP = Yes-ассоциированный белок; PAA = полиакриламид. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Изменения в экспрессии/распределении YAP, поле смещения субстрата и поле тяги раковой клетки PC9 на субстратах различной жесткости и во время раннего распространения. (A) Экспрессия YAP клетки PC9, посеянной на гелях PAA 2, 5 и 40 кПа и стеклянной крышке через 65 ч от первоначального прикрепления клеточной подложки. (B) Клетку PC9 засыпали на гель PAA 5 кПа и визуализировали в течение 10 ч после первоначального прикрепления клеточной подложки. Экспрессия YAP представлена интенсивностью зеленой флуоресценции. Примечание: Интенсивность YAP плато от 1,5 ч и далее. Цветовые полосы указывают уровни выражения YAP (зеленый = высокое выражение; черный = низкое выражение) в (A) и (B). (C) Деформация субстрата (перекрывающаяся с изображением яркого поля) в месте расположения ячейки представлена флуоресцентным полем смещения шарика в каждой точке времени. Направление смещения и величина показаны направлением стрелки и цветом соответственно. Поле смещения, вызванное клетками PC9, меньше, чем поле, вызванное ячейкой B2B. В течение 10-часового процесса распространения площадь клеток PC9 остается почти постоянной. Цветовая шкала указывает на величину смещения (малиновый = высокая величина; черный = низкая величина). D) Поле тяги (перекрывающееся с изображением яркого поля), рассчитанное по полю смещения. Тяга, генерируемая этой репрезентативной клеткой PC9, постепенно уменьшается с 6-го ч до 10-го ч. Белые и желтые пунктирные контуры очерчивают границы клетки и ядра соответственно. Цветовая шкала указывает на величину тяги (малиновый = высокая величина; черный = низкая величина). Шкала стержней = 20 мкм. Сокращения: YAP = Yes-ассоциированный белок; PAA = полиакриламид. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Распределение YAP в клетках B2B и PC9 на ранней стадии распространения. (A) Интенсивность YAP клетки B2B измеряется вдоль назначенной красной оси в 0-м и 10-м ч. (B) В 0-м ч интенсивность YAP показывает драматические различия концентраций между ядром и цитоплазмой. Через 10 ч интенсивность YAP становится более однородной по всему телу клетки. (C) Интенсивность YAP клетки PC9 измеряется вдоль назначенной синей оси в 0-м и 10-м ч. (D) В 0-м ч интенсивность YAP в ядре кажется выше, чем в цитоплазме, хотя разница не так заметна, как в клетках B2B. На 10-м ч интенсивность YAP в ядре все еще кажется немного выше, чем в цитоплазме, с тенденцией вариаций, аналогичной той, что в 0-м ч. Шкала стержней = 20 мкм (A, C). Аббревиатура: YAP = Yes-ассоциированный белок. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4: Измерение соотношения YAP N/C. (1) Примените Fiji ImageJ, чтобы нарисовать контур ядра и измерить его 2D-проецируемую область Анук. (2) Измерьте интенсивность флуоресценции внутри ядра Inuc. (3) Нарисуйте контур тела клетки и измерьте ее проецируемую площадь Acel. (4) Измерьте интенсивность флуоресценции внутри ячейки Icel. (5) Рассчитать плотность ядра YAP Dnuc, плотность цитоплазмы YAP Dcyto и их соотношение R: Dnuc=Inuc/Anuc; Dcyto=(Icel-Inuc)/(Acel-Anuc); R=Dnuc/Dcyto. Цветовая шкала обозначает уровни выражения YAP (зеленый = высокое выражение; черный = низкое выражение). Шкала бар = 20 мкм. Сокращения: YAP = Yes-ассоциированный белок; N = ядро; C = цитоплазма. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5: Отчетливая экспрессия YAP, морфология клеток / ядер и клеточное вытяжение при раке PC9 и нормальных клетках B2B во время распространения клеток. (A) Изменение соотношения YAP N/C в течение первых 10 ч одноклеточного распространения. Среднее отношение YAP N/C ячеек B2B (красная колонка; n = 10) изменилось с 2,54 ± 0,22 до 1,79 ± 0,21 (n = 10; p = 0,0022**), в то время как среднее отношение YAP N/C ячеек PC9 (синий столбец; n = 5) изменилось с 1,92 ± 0,26 до 1,57 ± 0,07 (p = 0,187 (ns)). (B) Среднее сцепление диполей в зависимости от времени. Среднее дипольное тяговое стяжение B2B-клеток изменилось с 256,17 ± 123,69 нН до 287,44 ± 99,79 нН (p = 0,7593 (нс)), а среднее дипольное стяжение клеток PC9 изменилось с 141,19 ± 33,62 нН до 168,52 ± 73,01 нН (p = 0,7137 (нс)). (C) Средняя площадь ячейки в зависимости от времени. Средняя площадь распространения клеток B2B увеличилась с 613,89 ± 102,43 мкм2 до 942,51 ± 226,71 мкм2 (p = 0,0512 (ns)), а средняя площадь распространения клеток PC9 изменилась с 495,78 ± 97,04 мкм2 до 563,95 ± 89,92 мкм2 (p = 0,5804 (ns)). D) средняя площадь ядра в зависимости от времени. Средняя площадь распространения ядра B2B-клеток увеличилась с 181,55 ± 36,18 мкм2 до 239,38 ± 43,12 мкм2 (p = 0,1217 (ns)), а средняя площадь распространения ядра клеток PC9 изменилась с 133,31 ± 30,05 мкм2 до 151,93 ± 22,49 мкм2 (p = 0,5944 (ns)). Сокращения: YAP = Yes-ассоциированный белок; N = ядро; C = цитоплазма; ns = незначимый. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 6: Коэффициент YAP N/C и сила дипольного тяги в зависимости от площади распространения клетки и ядра. Коэффициент YAP N/C и дипольное тяговое стяжение B2B-клеток (n=10) и PC9-клеток (n=5) рассчитываются от 6-го ч до 10-го ч после присоединения к подложке. (A) YaP N/C отношение как функция области распространения клеток. Коэффициенты YAP N/C клеток B2B варьируются от 1,16 до 2,53, в то время как yap N/C соотношения клеток PC9 варьируются от 1,27 до 1,88. Площадь распространения клеток B2B варьируется от 391,94 мкм2 до 1986,40 мкм2. Площадь распространения клеток PC9 колеблется от 284,46 мкм2 до 830,12 мкм2. (B) YaP N/C соотношение в зависимости от площади распространения ядра. Площадь распространения ядра B2B-клеток варьируется от 107,09 мкм2 до 514,28 мкм2. Площадь распространения ядра клеток PC9 колеблется от 58,03 мкм2 до 259,65 мкм2. Дипольное вытяжение B2B-клеток в зависимости от области распространения клеток (C) и области распространения ядра (D). Распространяющиеся и немигрирующие B2B-клетки демонстрируют более высокую тягу (от 47,50 нН до 1051,48 нН) с более низкой площадью клеток и ядер. При распространении и миграции B2B-клетки демонстрируют более низкое тяговое усилие (от 105,80 нН до 310,28 нН) с большими диапазонами площади клеток и ядер. Сокращения: YAP = Yes-ассоциированный белок; N = ядро; C = цитоплазма. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 7: Периядерное смещение в нормальных B2B и раковых клетках PC9. (A) Схематическая диаграмма бокового вида смещения околоядерных и периячеек, измеренных от смещения шарика в подложке. (B) Смещение подложки под ячейкой PC9 измеряется вдоль оси ячейки (белая пунктирная линия в 7D). Теоретическое смещение, создаваемое дипольной силой на границе ячейки, показано уравнением Буссинеска (черная пунктирная кривая). (C) и (D) Перекрывающиеся флуоресцентные изображения шариков с (красными) и без (зелеными) ячейками для ячеек PC9 через 6 ч после присоединения (вид сверху). Желтые (точное перекрытие красного и зеленого цветов) бусины указывают на отсутствие смещения. Разделенные зеленые и красные бусины (заостренные желтыми стрелками) представляют собой периядерное смещение. Желтые стрелки указывают на эти сжатые периядровые пятна, расположенные на периферии ядра. (E) Периядерное смещение, создаваемое ячейкой B2B через 1,5 ч после прикрепления ячейки-субстрата. Шкала стержней = 10 мкм (C–E). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительный рисунок S1: Карта геномных последовательностей YAP-mNeonGreen21-10/11. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S2: Изменения в экспрессии/распределении YAP, поле смещения субстрата и поле тяги нормальных клеток B2B во время раннего распространения. (A, D, G, J, M) Клетку B2B засыпали на гель PAA 5 кПа и визуализировали в течение 10 ч после первоначального прикрепления клетки-подложки. Экспрессия YAP представлена интенсивностью зеленой флуоресценции. Примечание: Интенсивность YAP внутри ядра постепенно уменьшается, но со временем остается выше, чем в цитоплазме. Цветовые полосы обозначают уровни выражения YAP (зеленый = высокое выражение; черный = низкое выражение) в (A, D, G, J, M). (B, E, H, K, N) Деформация подложки (перекрывающаяся с изображением яркого поля) в месте расположения ячейки представлена полем смещения в каждой точке времени. Направление смещения и величина показаны направлением стрелки и цветом соответственно. Смещение становится больше на периферии тела клетки B2B по мере увеличения площади распространения клеток. Цветовые полосы обозначают величину смещения (малиновый = высокая величина; черный = низкая величина) в (B, E, H, K, N). (С, Ф, И, Л, О) Поле тяги (перекрывающееся с изображением яркого поля), рассчитанное из поля смещения с помощью traction Force Microscopy. Тяга концентрируется на периферии B2B-клеток. Цветовые полосы обозначают величину тяги (малиновый = высокая величина; черный = низкая величина) в (C, F, I, L, O). Шкала стержней = 20 мкм. Сокращения: YAP = Yes-ассоциированный белок; PAA = полиакриламид. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S3: Изменения в экспрессии/распределении YAP, поле смещения субстрата и поле тяги раковых клеток PC9 во время раннего распространения. (A, D, G, J) Клетку PC9 засевали на гель PAA 5 кПа и визуализировали в течение 10 ч после первоначального прикрепления клетки-субстрата. Экспрессия YAP представлена интенсивностью зеленой флуоресценции. Примечание: Интенсивность YAP внутри ядра постепенно уменьшается, но остается аналогичной или немного ниже, чем в цитоплазме с течением времени. Цветовые полосы обозначают уровни выражения YAP (зеленый = высокое выражение; черный = низкое выражение) в (A, D, G, J). (В, Е, Х, К) Деформация подложки (перекрывающаяся с изображением яркого поля) в месте расположения ячейки представлена полем смещения в каждой точке времени. Направление смещения и величина показаны направлением стрелки и цветом соответственно. Смещение становится больше на периферии тела клетки PC9 по мере увеличения площади распространения клеток. Цветовые полосы обозначают величину смещения (малиновый = высокая величина; черный = низкая величина) в (B, E, H, K). (С, Ф, И, Л) Поле тяги (перекрывающееся с изображением яркого поля), вычисляемое из поля смещения. Тяга концентрируется на периферии клеток PC9. Цветовые полосы обозначают величину тяги (малиновый = высокая величина; черный = низкая величина) в (C, F, I, L). Шкала стержней = 20 мкм. Сокращения: YAP = Yes-ассоциированный белок; PAA = полиакриламид. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
Процесс визуализации (этап 6.3) имеет решающее значение для обеспечения того, чтобы флуоресцентные изображения имели достаточно хорошее качество для получения достоверных результатов количественной оценки. Изображения z-стека флуоресцентного белка или бусин должны иметь z-диапазон, который достаточно велик, чтобы включать изображения в фокусе для всех Z-позиций, которые охватывает образец. Другим важным этапом является сбор эталонных изображений флуоресцентных шариков после растворения клеток (шаг 6.5). Поскольку эталонные изображения должны быть сделаны в тех же положениях на этапе 6.3, не следует индуцировать относительное смещение между чашкой Петри, камерой окружающей среды и микроскопом. Исследователи, выполняющие этап растворения, должны быть осторожны, чтобы снять крышку чашки Петри и убедиться, что нанесенное механическое возмущение недостаточно велико, чтобы изменить местоположение чашки в камере окружающей среды.
Ниже приведены решения для устранения некоторых ошибок, которые могут возникнуть во время экспериментов. Если макрос не активирован после нажатия клавиши Ввод на шаге 6.4, скорее всего, это связано с тем, что левая нижняя область экрана занята окном, не являющимся элементом. В таком случае левая нижняя часть окна должна быть очищена, чтобы макросы можно было активировать в Elements. Другая распространенная ошибка заключается в том, что изображения яркого поля кажутся черными. Эта проблема вызвана недостаточным временным интервалом между получением флуоресцентных и ярко-полевых изображений. Небольшие задержки в подсчете времени флуоресцентной визуализации могут накапливаться с течением времени и вызывать значительные задержки и мешать визуализации яркого поля. Одним из решений является регулировка продолжительности одного цикла визуализации всех положений, чтобы она была меньше (не равна) временному интервалу между началом последовательных движений. Эта операция обновляет подсчет времени и устраняет кумулятивную ошибку в начале каждого цикла обработки изображений.
Эта полностью оптическая технология опроса поддерживает (1) широкий спектр аппаратного и программного обеспечения, включая, но не ограничиваясь Nikon, (2) различные типы проверенных гидрогелевых систем, включая желатин, PEG, Matrigel и коллаген I гели, и (3) программируемую настройку на основе различных потребностей исследователей. Однако, если какая-либо из функций управления нижнего уровня недоступна из коммерческого микроскопа, настройка функций с помощью AMFIP становится сложной задачей. Другим ограничением этого метода является пространственный дрейф образца как в плоскости XY, так и в плоскости фокусировки (Z). Хотя это ограничение может быть преодолено во время постобработки изображений, важно улучшить функцию автофокусировки для коррекции дрейфа образцов в реальном времени. Это улучшение увеличит пропускную способность процесса визуализации и уменьшит потенциальную ошибку, вызванную дрейфом во время экспериментов.
Механотрансдукторы, такие как YAP, могут служить новыми терапевтическими мишенями для разработки перспективных методов лечения рака25,26,27. Новые данные свидетельствуют о том, что YAP способствует пролиферации и инвазии раковых клеток. Индуцированная механикой транслокация YAP из цитоплазмы в ядро активирует транскрипцию генов, связанных с миграцией клеток, пролиферацией, инвазией и апоптозом, что приводит к аберрантному поведению клеток28,29,30,31. Эта работа была направлена на изучение потенциальной корреляции соотношения YAP N / C и клеточной механики в двух типичных линиях рака легких человека и нормальных клеточных линиях. В течение 10-часового периода распространения клеток клетки PC9 демонстрируют аналогичные концентрации YAP в ядре и цитоплазме (рисунок 3D и рисунок 5A). Клетки B2B показывают более высокую концентрацию YAP в ядре, чем в цитоплазме (рисунок 3C и рисунок 5A). Эта связь, обнаруженная на ранней стадии распространения, отличается от большинства опубликованных результатов, которые сравнивают концентрацию YAP в ядре между нормальными и раковыми клетками. Хотя это не обязательно на ранней стадии распространения, большинство опубликованных результатов показывают, что YAP более сконцентрирован в ядре раковых клеток, чем в ядре нормальных клеток27,28. Только в одном исследовании рака молочной железы сообщалось об исключении32, которое показывает, что YAP более сконцентрирован в цитоплазме, что согласуется с нашими текущими наблюдениями, сделанными в клетках рака легких PC9. Насколько известно авторам, эта работа является первой, которая показала более низкое соотношение YAP N / C в клеточной линии рака легких человека. Авторы предполагают, что причина стабильного соотношения YAP N/C в клетках PC9 может быть связана с низкой вариацией в области распространения клетки / ядра и вытяжением в клетках PC9 на ранней стадии распространения. Продолжается вскрытие основополагающих молекулярных механизмов низкого соотношения YAP N/C в клетках PC9 и B2B.
В течение первых 10 ч распространения эти две клеточные линии показывают четкую связь между соотношением YAP N/C, тягой клеток и площадью распространения (рисунок 5). Для B2B-клеток более высокое соотношение YAP N/C коррелирует с более высокой площадью распространения клеток и ядер (рисунок 6A,B), что согласуется с представленными данными других нормальных клеток33. Интересно, что, хотя тенденция развития этой связи обычно обнаруживается во всех зарегистрированных B2B-клетках, обнаруживаются две разные степени (высокая и низкая) этой связи. Клетки B2B, которые распространяются и мигрируют одновременно, демонстрируют более низкое вытяжение и более высокую площадь распространения клеток и ядер с более высоким коэффициентом YAP N / C (2,05 ± 0,32). Для B2B-клеток, которые распространяются и остаются в том же месте, они показывают более высокую тягу и более низкую площадь распространения клеток и ядер с более низким соотношением YAP N / C (1,74 ± 0,21). Эти две степени взаимосвязей демонстрируются в раздвоенных разрозненных группах данных (рисунок 6C,D). Как сообщается в литературе, стационарные нормальные клетки, такие как эмбриональные фибробласты NIH 3T3, имеют более высокую тягу, чем мигрирующие клетки34. Данные, представленные в этой статье, свидетельствуют о том, что распространяющиеся и немигрирующие B2B-клетки применяли более высокую тягу, чем распространяющиеся и мигрирующие B2B-клетки, вероятно, предполагая, что для стабилизации немигрирующих клеток на субстрате требуется высокое сцепление.
Кроме того, эти данные показывают, что стационарные нормальные B2B-клетки генерируют более высокую периядерную силу, тогда как предыдущие исследования, проведенные другими исследователями, сообщали только о более высокой тяге клеток, генерируемой на периферии стационарных клеток34,35,36,37. Авторы считают, что разница во внутренней тенденции миграции в экспериментах может привести к этим противоречивым результатам. В опубликованных экспериментах микроструктурирование квадратной формы использовалось для ограничения распространения одиночных клеток и ингибирования миграции; имели ли клетки тенденцию к миграции, неизвестно. Поскольку мигрирующие клетки часто демонстрируют высокую тяговую силу на периферии клеток38, вполне вероятно, что клетки с тенденцией к миграции будут по-прежнему поддерживать высокое тяговое усилие периферии, даже если их миграция ограничена. В этом настоящем исследовании стационарные клетки не ограничены каким-либо микрошаблоном, но не мигрируют, что указывает на то, что клетки, как правило, сохраняют свое немигрирующее состояние. Другая возможность заключается в том, что форма клеток, определенная микрошаблоном, может влиять на распределение очаговых спаек и сил тяги39. Результаты этого исследования были получены без какого-либо ограничивающего микроструктурирования и представляют собой распределение сил стационарных клеток в их первоначальной форме.
Насколько известно авторам, только в одной публикации на сегодняшний день конкретно сообщалось об обнаружении периядерных сил в нормальных клетках (эмбриональных фибробластах мышей), потенциально вызванных актиновым колпачком, охватывающим ядро40. Транслокация цитоплазмы в ядро YAP коррелирует с увеличением периядерной силы40. Тщательный поиск соответствующей литературы не привел ни к одной публикации, в которой сообщалось бы о периядерной силе или актиновом колпачке в раковых клетках. Косвенное исследование раковых клеток меланомы показало, что актиновый край (другая периядерная актиновая организация, расположенная вокруг, но не покрывающая ядро) снижает скорость миграции клеток41, косвенно предполагая существование периядерной силы. Однако прямых экспериментальных данных не сообщается. В этом исследовании авторы обнаружили, что как PC9, так и B2B-клетки показывают периядерное смещение и тягу. Механизмы генерации периядерных сил и их последствия остаются спорными. Сообщалось, что в нормальных клетках актиновый колпачок играет роль в регулировании морфологии ядра и организации хроматина42, передавая механические сигналы от фокальных спаек в ядро через линкеры нуклеоскелета и цитоскелета (LINC) complex43 и регулируя миграцию клеток44. Lamin A/C связан с образованием и разрушением актиновой капсулы40,41,42,43,44. Однако в докладе, в котором утверждалось, что актиновый колпачок генерирует периядерную силу, не рассматривалась потенциальная роль актина rim40. В раковых клетках сверхэкспрессия Ламина А облегчает образование актинового обода и ограничивает миграцию раковых клеток. Сверхэкспрессия Ламина В уменьшает образование актиновых ободков и способствует миграции. Периядерные силы могут быть вовлечены в этот процесс из-за существования периядерной актиновой организации и эффекта Ламина А. Однако результаты этого исследования не показали каких-либо доказательств измеренных периядерных сил или поведения актинового колпачка. Таким образом, открытие периядерных сил в клетках PC9 в этом настоящем исследовании является первым отчетом, показывающим периядерные силы и смещения в клетках рака легких. В настоящее время авторы исследуют молекулярные механизмы и функции периядерных сил в клетках PC9 и B2B, спроектированных CRISPR / Cas9.
Помимо полностью оптического механобиологического опроса, который демонстрируется в этой статье, интегрированная многофункциональная система может быть применена для оптического зондирования множества других важных физиологических и патобиологических сигналов в живых системах. Например, лаборатория авторов недавно установила несколько стабильно трансдуцированных линий раковых клеток человека, которые совместно экспрессируют три светочувствительных мембранных белка: индикатор напряжения мембраны QuasAr2 (возбуждение: 640 нм; излучение: 660 нм-740 нм), деполяризатор мембранного напряжения CheRiff (возбуждение: 488 нм) и гиперполяризатор мембранного напряжения eNpHR3 (возбуждение: 590 нм). Эти три функциональных белка могут быть активированы спектрально-ортогональными лазерными линиями без перекрестных помех, что обеспечивает полностью оптическую двустороннюю сигнальную связь (считывание и управление) мембранной электрофизиологии. Используя интегрированную оптоэлектронную систему и ручной патч-зажим, авторы подтвердили полностью оптический контроль и считывание напряжения мембраны (Vm) в одиночных раковых клетках человека и многоклеточных сфероидах опухолей. Полностью оптический электрофизиологический опрос открывает возможность для детальных исследований ранее недоступного биоэлектричества в раковых клетках, что может помочь продвинуть биологию опухоли с новой оси.
Раскрытие информации
Нет никаких конфликтов интересов, о которых можно было бы заявить.
Благодарности
Этот проект финансово поддерживается премией Cancer Pilot Award от UF Health Cancer Center (X. T. и D. S.) и Gatorade Award Start-up Package (X. T.). Авторы искренне ценят интеллектуальные дискуссии и техническую поддержку со стороны д-ра Джонатана Лихта (UFHCC), д-ра Рольфа Ренне (UFHCC), д-ра Джи-Хён Ли (биостатистика, UF), д-ра Хью Фана (MAE, UF), д-ра Уоррена Диксона (MAE, UF), д-ра Гату Субхаша (MAE, UF), д-ра Марка Шеплака (MAE & ECE, UF), д-ра Малисы Сарнтиноранонт (MAE, UF), д-ра Скотта Бэнкса (MAE, UF), д-р Мэтью Траум (MAE, UF), д-р Дэвид Хан (Университет Аризоны), д-р Вэйхонг Ван (Oracle Corporation), д-р Юхуа Тан (Гонконгский политехнический университет) и группа поддержки Nikon (д-р Хосе Серрано-Велес, Ларри Кордон и Джон Экман). Авторы глубоко благодарны за щедрую и эффективную поддержку со стороны всех членов исследовательских лабораторий Tang' s, Siemann и Guan, а также всех сотрудников MAE & ECE & Physics & Radiation Oncology Departments, UF.
Материалы
| Name | Company | Catalog Number | Comments |
| (3-Aminopropyl)triethoxysilane | Sigma-aldrich | 440140 | |
| 0.05 % Trypsin | Corning | 25-051-CI | |
| 75 cm2 flask | Corning | 430641U | |
| 8 Benchtop Centrifuge | Thermo | 75007210 | |
| A1R confocal system | Nikon | HD25 | |
| Acetic acid | Sigma-aldrich | 695092 | glacial, ACS reagent, ≥99.7% |
| BEAS-2B (B2B) cells | Sigma-aldrich | 95102433 | human epithelial cells from lung tissue |
| Carboxylate-Modified Microspheres | Invitrogen | F8797 | |
| Culture medium (RPMI-1640) | Gibco | 11875093 | |
| Desktop Computer | Dell | 2018 | with Windows 10 operating system |
| Environmental chamber TIZB | Tokai Hit | TIZB | |
| Fetal bovine serum (FBS) | Gibco | 26140 | |
| Fibronectin Human Protein, Plasma | Gibco | 33016015 | |
| Fiji ImageJ | National Institutes of Health and the Laboratory for Optical and Computational Instrumentation | 1.53k | |
| Glass-bottom petri dish | MatTek | P35G-1.5-14-C | |
| HEPES buffered saline | Sigma-aldrich | 51558 | |
| Hydrazine hydrate solution | Sigma-aldrich | 53847 | |
| IntelliJ IDEA | JetBrains | 2020 | Java development platform |
| Java Development Kit | Oracle | 14.0 | |
| Kimwipe | Kimtech Science | 3066-05 | |
| MATLAB | MathWorks | 2020b | |
| Monochrome Camera | FLIR | BFS-U3-70S7M-C | |
| MycoAlert Mycoplasma Detection Kit | Lonza | LT07-218 | |
| N,N′-Methylenebisacrylamide solution | Sigma-aldrich | M1533 | |
| NIS-Elements software platform | Nikon | 4.50 | software platform |
| Origin | OriginLab | OriginPro 2017 (Learning Edition) | data analysis and graphing software |
| Penicillin-streptomycin | Gibco | 15140122 | |
| PC9 cells | Sigma-aldrich | 90071810 | human adenocarcinoma cells from lung tissue |
| Phosphate buffered saline (PBS) | Gibco | 10010023 | |
| Phusion High-Fidelity DNA Polymerase | New England Biolabs | F-553S | high-fidelity DNA polymerase |
| Scotch tape | Scotch | adhesive tape | |
| Sodium dodecyl sulfate solution | Sigma-aldrich | 05030 | |
| Super glue | Gorilla | cyanoacrylate glue | |
| Ti2-E inverted microscope | Nikon | MEA54000 | |
| TI2-S-SE-E Motorized Stage with Encoder | Nikon | MEC56120 | |
| μManager | version 2.0 gamma | open source microscopy software (https://micro-manager.org/) |
Ссылки
- Werley, C., Boccardo, S., Rigamonti, A., Hansson, E., Cohen, A. Multiplexed optical sensors in arrayed islands of cells for multimodal recordings of cellular physiology. Nature Communications. 11 (1), 3881 (2020).
- Yang, B., et al. Epi-illumination SPIM for volumetric imaging with high spatial-temporal resolution. Nature Methods. 16 (6), 501-504 (2019).
- Saraswathibhatla, A., Galles, E. E., Notbohm, J. Spatiotemporal force and motion in collective cell migration. Scientific Data. 7 (1), 197 (2020).
- Saraswathibhatla, A., Henkes, S., Galles, E. E., Sknepnek, R., Notbohm, J. Coordinated tractions control the size of a collectively moving pack in a cell monolayer. Extreme Mechanics Letters. 48, 101438 (2021).
- Wang, W., Kim, C. K., Ting, A. Y. Molecular tools for imaging and recording neuronal activity. Nature Chemical Biology. 15 (2), 101-110 (2019).
- Eliceiri, K. W., et al. Biological imaging software tools. Nature Methods. 9 (7), 697-710 (2012).
- Carpenter, A. E., Kamentsky, L., Eliceiri, K. W. A call for bioimaging software usability. Nature Methods. 9 (7), 666-670 (2012).
- Skylaki, S., Hilsenbeck, O., Schroeder, T. Challenges in long-term imaging and quantification of single-cell dynamics. Nature Biotechnology. 34 (11), 1137-1144 (2016).
- Lin, M. Z., Schnitzer, M. J. Genetically encoded indicators of neuronal activity. Nature Neuroscience. 19 (9), 1142-1153 (2016).
- Luo, Q., et al. Automatic multi-functional integration program (AMFIP) towards all-optical mechanobiology interrogation. bioRxiv. , (2021).
- Edelstein, A., Amodaj, N., Hoover, K., Vale, R., Stuurman, N. Computer control of microscopes using manager. Current Protocols in Molecular Biology. 92 (1), 14-20 (2010).
- Tulpule, A., et al. Kinase-mediated RAS signaling via membraneless cytoplasmic protein granules. Cell. 184 (10), 2649-2664 (2021).
- Tang, X., Tofangchi, A., Anand, S. V., Saif, T. A. A novel cell traction force microscopy to study multi-cellular system. PLOS Computational Biology. 10 (6), 1003631 (2014).
- Tang, X., et al. Mechanical force affects expression of an in vitro metastasis-like phenotype in HCT-8 cells. Biophysical Journal. 99 (8), 2460-2469 (2010).
- Guimarães, C. F., Gasperini, L., Marques, A. P., Reis, R. L. The stiffness of living tissues and its implications for tissue engineering. Nature Reviews Materials. 5, 351-370 (2020).
- Phelps, E. A., et al. Maleimide cross-linked bioactive PEG hydrogel exhibits improved reaction kinetics and cross-linking for cell encapsulation and in situ delivery. Advanced Materials. 24 (1), 64-70 (2012).
- Bajaj, P., Tang, X., Saif, T. A., Bashir, R. Stiffness of the substrate influences the phenotype of embryonic chicken cardiac myocytes. Journal of Biomedical Materials Research. Part A. 95 (4), 1261-1269 (2010).
- Temples, M. N., Adjei, I. M., Nimocks, P. M., Djeu, J., Sharma, B. Engineered three-dimensional tumor models to study natural killer cell suppression. ACS Biomaterials Science & Engineering. 6 (7), 4179-4199 (2020).
- Feng, S., et al. Improved split fluorescent proteins for endogenous protein labeling. Nature Communications. 8, 370 (2017).
- Guan, J., Liu, H., Shi, X., Feng, S., Huang, B. Tracking multiple genomic elements using correlative CRISPR imaging and sequential DNA FISH. Biophysical Journal. 112 (6), 1077-1084 (2017).
- . Micro-Manager Available from: https://micro-manager.org/wiki/NikonTi2 (2021)
- Schindelin, J., et al. Fiji: An open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Martiel, J. L., et al. Measurement of cell traction forces with ImageJ. Methods in Cell Biology. 125, 269-287 (2015).
- Okumurai, I. A. On the generalization of Cerruti's problem in an elastic half-space. Doboku Gakkai Ronbunshu. 1995, 1-10 (1995).
- Piccolo, S., Dupont, S., Cordenonsi, M. The biology of YAP/TAZ: hippo signaling and beyond. Physiological Reviews. 94 (4), 1287-1312 (2014).
- Hong, W. W., Guan, K. L. The YAP and TAZ transcription co-activators: Key downstream effectors of the mammalian Hippo pathway. Seminars in Cell and Developmental Biology. 23 (7), 785-793 (2012).
- Zanconato, F., Cordenonsi, M., Piccolo, S. YAP/TAZ at the roots of cancer. Cancer Cell. 29 (6), 783-803 (2016).
- Wang, Y., et al. Overexpression of yes-associated protein contributes to progression and poor prognosis of non-small-cell lung cancer. Cancer Science. 101 (5), 1279-1285 (2010).
- Li, H., et al. Inhibition of YAP suppresses CML cell proliferation and enhances efficacy of imatinib in vitro and in vivo. Journal of Experimental & Clinical Cancer Research. 35 (1), 134 (2016).
- Tang, X., et al. A mechanically-induced colon cancer cell population shows increased metastatic potential. Molecular Cancer. 13, 131 (2014).
- Panciera, T., Azzolin, L., Cordenonsi, M., Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nature Reviews Molecular Cell Biology. 18 (12), 758-770 (2017).
- Yuan, M., et al. Yes-associated protein (YAP) functions as a tumor suppressor in breast. Cell Death and Differentiation. 15 (11), 1752-1759 (2008).
- Koushki, N., et al. Lamin A redistribution mediated by nuclear deformation determines dynamic localization of YAP. bioRxiv. , (2020).
- Chang, S. S., Rape, A. D., Wong, S. A., Guo, W. H., Wang, Y. L. Migration regulates cellular mechanical states. Molecular Biology of the Cell. 30 (26), 3104-3111 (2019).
- Lee, J., Abdeen, A. A., Tang, X., Saif, T. A., Kilian, K. A. Geometric guidance of integrin mediated traction stress during stem cell differentiation. Biomaterials. 69, 174-183 (2015).
- Lee, J., Abdeen, A., Tang, X., Saif, T. A., Kilian, K. A. Matrix directed adipogenesis and neurogenesis of mesenchymal stem cells derived from adipose tissue and bone marrow. Acta Biomaterialia. 42, 46-55 (2016).
- Tang, X., Bajaj, P., Bashir, R., Saif, T. A. How far cardiac cells can see each other mechanically. Soft Matter. 7 (13), 6151-6158 (2011).
- Dembo, M., Wang, Y. L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophysical Journal. 76 (4), 2307-2316 (1999).
- Rape, A., Guo, W. H., Wang, Y. L. The regulation of traction force in relation to cell shape and focal adhesions. Biomaterials. 32 (8), 2043-2051 (2011).
- Shiu, J. Y., Aires, L., Lin, Z., Vogel, V. Nanopillar force measurements reveal actin-cap-mediated YAP mechanotransduction. Nature Cell Biology. 20 (3), 262-271 (2018).
- Fracchia, A., Asraf, T., Salmon-Divon, M., Gerlitz, G. Increased lamin B1 levels promote cell migration by altering perinuclear actin organization. Cells. 9 (10), 2161 (2020).
- Ramdas, N. M., Shivashankar, G. V. Cytoskeletal control of nuclear morphology and chromatin o1rganization. Journal of Molecular Biology. 427 (3), 695-706 (2015).
- Khatau, S. B., et al. A perinuclear actin cap regulates nuclear shape. Proceedings of the National Academy of Sciences of the United States of America. 106 (45), 19017-19022 (2009).
- Kim, D. H., Cho, S., Wirtz, D. Tight coupling between nucleus and cell migration through the perinuclear actin cap. Journal of Cell Science. 127 (11), 2528-2541 (2014).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены