
Method Article
تتبع النسب وتحليل التخفي في تطوير قشرة الدماغ باستخدام تحليل الفسيفساء مع علامات مزدوجة (MADM)
In This Article
Summary
يتم تقديم بروتوكول لتنفيذ تتبع النسب والتحليل الجيني الوظيفي للجينات المرشحة على مستوى خلية واحدة باستخدام تحليل الفسيفساء مع علامات مزدوجة (MADM). يوفر تحليل MADM التخفي إطارًا كميًا لقياس السلوك التكاثري ، والإخراج الخلوي ، وعلاقة النسب بين السلف الفردية وخلايا ابنتهم.
Abstract
بدءا من مجموعة محدودة من السلف، والقشرة الدماغية الثدييات تشكل دوائر عصبية وظيفية منظمة للغاية. ومع ذلك، فإن الآليات الخلوية والجزيئية الأساسية التي تنظم انتقالات النسب للخلايا الجذعية العصبية (NSCs) وإنتاج الخلايا العصبية والجليا في الخلايا العصبية النامية لا تزال غير واضحة. وقد تقدمت أساليب تتبع أنماط تقسيم NSC وخريطة نسب الخلايا المرتبطة بالكلونالي بشكل كبير. ومع ذلك ، العديد من تقنيات تتبع النسب المعاصرة تعاني من عدم وجود القرار الخلوية من الخلية السلف مصير ، وهو أمر ضروري لفك أنماط تقسيم الخلية السلف. قدم هو بروتوكول باستخدام تحليل الفسيفساء مع علامات مزدوجة (MADM) لأداء في تحليل clonal الجسم الحي. MADM يتلاعب في الوقت ذاته الخلايا السلف الفردية وتصورات أنماط التقسيم الدقيقة وتطور النسب في قرار خلية واحدة لم يسبق لها مثيل. MADM القائم على أحداث إعادة التركيب بين الكومبوسومات خلال مرحلة G2-X من الانقسام، جنبا إلى جنب مع غير قابل للتكرير زمنيا T2،وتوفير معلومات دقيقة عن تواريخ ميلاد المستنسخين وأنماط تقسيمها. وهكذا، فإن تتبع نسب MADM يوفر قراءات بصرية نوعية وكمية غير مسبوقة لأسلوب انتشار ذريات الخلايا الجذعية على مستوى الخلية الواحدة. كما يسمح MADM بفحص الآليات والمتطلبات الوظيفية للجينات المرشحة في تقدم نسب NSC. هذه الطريقة هي فريدة من نوعها في أن التحليل المقارن للسيطرة و subclones متحولة يمكن أن يؤديها في نفس بيئة الأنسجة في الجسم الحي. هنا ، يتم وصف البروتوكول بالتفصيل ، ويتم عرض نماذج تجريبية لتوظيف MADM للتحليل التخفي وتتبع النسب في قشرة الدماغ النامية. الأهم من ذلك، يمكن تكييف هذا البروتوكول لأداء تحليل MADM clonal في أي المتخصصة الخلية الجذعية مورين، طالما أن سائق T2 CreER موجود.
Introduction
القشرة الدماغية هي بنية منظمة للغاية تتألف من ست طبقات متميزة. القشرة تحتوي على مجموعة متنوعة من أنواع الخلايا بما في ذلك الخلايا العصبية وglia، والتي تتفاعل لتشكيل الدوائر العصبية الوظيفية. معظم, إن لم يكن كل, الخلايا العصبية الإسقاط القشرية excitatory وglia مشتقة من مجموعة مشتركة من الخلايا الجذعية العصبية (NSCs) المعروفة باسم السلفايلي شعاعي (RGPs)1,2,3. يتم اشتقاق RGPs أنفسهم من الخلايا الجذعية العصبية (NESCs) التي تؤلف العصبية الجنينية المبكرة. قبل يوم الجنين 9 (E9) في الفئران، تبدأ NESCs للانتقال إلى RGPs4. RGP تطور النسب يتطلب تنظيما زمنيا ومكانيا دقيقا، وعندما يتم إعاقة هذه العملية، والاضطرابات العصبية الشديدة مثل megalencephaly، microcephaly، lissencephaly، أو العاهات مثل الفصام والتوحد يمكن أن يؤدي5،,6. في E10، تخضع معظم RGPs انقسامات تكاثرية متناظرة، مما يؤدي إلى توسع بركة السلف العصبي4،7. تبدأ RGPs في نهاية المطاف في الانقسام بشكل غير متماثل ، مما ينتج الخلايا العصبية الإسقاط القشرية بطريقة محددة زمنيًا. من خلال موجات متتالية من تولد الخلايا العصبية العصبية, الخلايا العصبية حديثي الولادة تهاجر في لوحة القشرية تشكيل اللعم القشرية مع الخلايا العصبية المولودة في وقت مبكر تحتل طبقات عميقة والخلايا العصبية المولودة في وقت متأخر المقيمين في طبقات سطحية8,,9,,10. لأن الخلايا العصبية الهرمية ذات الصلة كلونالي تهاجر شعاعي في القشرة مع تشتت عرضية قليلا جدا، خلايا ابنة تميل إلى تشكيل عمود أو هيكل مخروط على شكل المشار إليها باسم وحدة شعاعية الخلايا العصبية4,11,12,13. بواسطة E17، توسع عصبي الجنيني كامل في الفئران14. يمكن أيضا إنتاج RGPs الخلايا ependymal وبعض فئات من glia، بما في ذلك الفلكيات و oligodendrocytes1،15،16،17،18،19. إمكانات RGPs لتعطي على حد سواء الخلايا العصبية والخلايا الفلكية ويبدو أن تكون متسقة عبر جميع المناطق القشرية18, مع ما يقرب من 1/6 من RGPs العصبية أيضا إنتاج glia11.
وفي الوقت الراهن، فإن العوامل الوراثية والوراثية اللاجينية التي تنظم التطور الزمني للخلايا الجذعية على طول نسبها غير معروفة في معظمها. قد يكون للأنماط الزمنية للتعبير الجيني تأثير كبير على قرارات النسب في RGPs20،21،22،23،24. كيف لا تعرف هذه العلاقة المتماسكة بإحكام بين النقش الزماني والمكاني إلى التنوع الجزيئي لأنواع الخلايا العصبية البالغة عبر المناطق القشرية. وبالمثل، فإن كيفية تعديل إمكانات الخلايا الجذعية الفردية وإخراجها الخلوي على المستوى الخلوي والجزيئي هو سؤال مهم لم تتم الإجابة عليه. ومن المأمول أن تتناول الدراسات المستقبلية بعض هذه الأسئلة، مما يوسع فهمنا في نهاية المطاف لتشكيل الدائرة القشرية الوظيفية.
يسعى علم الأعصاب التنموي إلى فهم علاقة النسب التي تشترك فيها الخلايا في الدماغ مع بعضها البعض. في البداية، كانت أدوات البحث قليلة جدا المتاحة لهذا، والعديد من الدراسات المبكرة تعتمد على الملاحظات البصرية من أنماط الانقسام في الكائنات الحية الشفافة مثل Caenorhabditis elegans25. شهدت العقود الأخيرة زيادة كبيرة في عدد وتطور التقنيات المتاحة13،26،27،28،29. ظهور CRISPR-Cas9 نظام تحرير الجينوم يسمح لإعادة بناء الاصطناعية للعلاقات نسب الخلية من خلال إدخال الباركود DNA المتطورة27,30. مثالان حديثان على استراتيجيات الباركودينغ تشمل استخدام توجيه الجيش الملكي النيبالي الذي يوجه CRISPR-Cas9 إلى loci الباركود الحمض النووي محددة أو cytidine deaminase تنصهر مع نيكاز Cas9 لاستهداف المناطق تكرار تتخللها الذاتية31,32. توفر هذه التقنيات نهجًا متعددة للغاية من خلال إدخال الباركود الذي يتراكم بشكل تدريجي ومطرد طفرات فريدة مع مرور الوقت. ونُهج تحرير الجينوم ذات قيمة عالية لأنها تسمح بالتحليل بأثر رجعي للعلاقة بين أي خليتين استناداً إلى الميراث المشترك لهذه الباركود. ومع ذلك، من أجل قراءة الباركود في الخلايا الفردية، يجب عادة أن تتعطل الأنسجة، وبالتالي يتم فقدان المعلومات المتعلقة الموقف، مورفولوجيا، وأرقام الخلايا المطلقة من السلف الفردية.
النماذج الملصقات الجمعية الحفاظ على المعلومات المكانية، ومن حيث المبدأ تسمح أيضا للتمييز بين النسخ المترجمة بشكل وثيق أو حتى المتداخلة33,34. لطريقة تتبع النسب لتكون مفيدة يجب أن تسمية النسل الفردية وانسلهم بطريقة متفرقة ولا تمحى. وتجدر الإشارة إلى أن Brainbow35 وقصاصة36،37 نهج استخدام المراسلين متعدد الألوان القائم على Cre المؤتلف الذي يعبر عن مزيج من البروتينات الفلورية من موضع واحد. العدد الواسع من تركيبات الألوان المتزامنة التي يمكن تحقيقها في الجسم الحي تجعل هذه أداة قوية عند تتبع مستنسخات RGP القشرية و34. كما تم تطوير الأنظمة القائمة على Transposon توفير التكامل الجينومي المستقر من transgenes ترميز المراسلين الفلورسنت والسماح النسب تتبع السلف القشرية33,38,39,40,41. نظم transposon القائم على ميزة إضافية في أن المراسل يبني اندماج ثابت في الجينوم، وبالتالي تسمية موثوق بها بالخلايا ابنة ذات الصلة خط. لتتبع أنساب astrocyte على وجه التحديد، وقد تم تطوير عدد من الأساليب التي تنطوي على electroporation من نايغباك transposases بما في ذلك Star Track، والذي يجعل من استخدام مزيج من الانشاءات ترميز البروتينات الفلورية المختلفة40،42. نهج آخر، علامات MAGIC، يقدم ناقلات Brainbow كما transgenes القابلة للناقل. وقد تم استخدام هذا بنجاح لتتبع الجنينية العصبية وprogenitors استروسيت34,43. في الآونة الأخيرة، تم العثور على تحليل الفسيفساء عن طريق تبادل الكاسيت المزدوج بوساطة إعادة الكومبينز (MADR) لتسمية الخلايا المتحولة بشكل ثابت التعبير عن عناصر المعدلة وراثيا من loci الكروموسومات محددة بدقة44. وقد وفرت هذه قوية في تقنيات وضع العلامات الجمعية vivo العديد من الأفكار في ديناميات النسب من الخلايا السلف. ومع ذلك، يتم إجراء هذه التحليلات على أنسجة ثابتة، مما يوفر لقطة من المستنسخات الفردية في مرحلة نمو محددة. من أجل مراقبة التغيرات في ديناميات النسب من استنساخ واحد مع مرور الوقت، المزمنة في تقنيات التصوير الجسم الحي مماثلة لتلك التي أجريت في gyrus دنت الكبار تحتاج إلى تطبيق45.
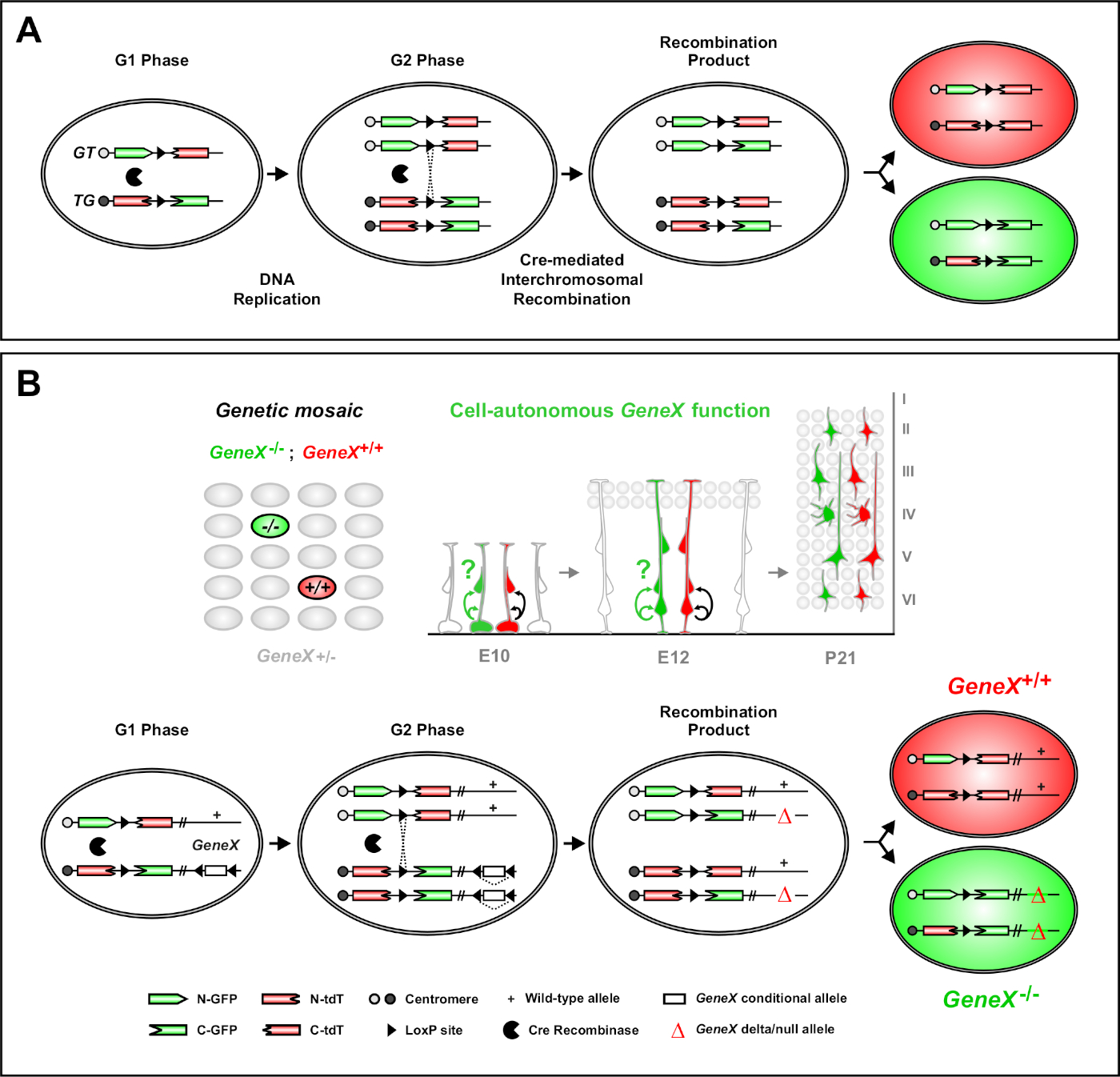
تحليل الفسيفساء مع علامات مزدوجة (MADM) هو قوي مزدوج اللون طريقة وضع العلامات التي تمكن في نسب الجسم الحي تتبع الخلايا السلف الفردية في الفئران46،47. هناك عنصران ضروريان لحدوث أحداث وضع العلامات على MADM: أولاً، يجب أن تكون أشرطة MADM موجهة إلى مكان متطابق على الكروموسومات المتماثلة. تتكون الكاسيتات من جينتين مراسلين من نوع chimeric fluorescent، eGFP (الأخضر، [G]) وطماطم الخافتة جنبا إلى جنب (أحمر، tdT[T]). يحتوي الكاسيت GT على N-terminus eGFP وC-terminus من tdT، مفصولة بـ intron يحتوي على موقع loxP. تم بناء كاسيت TG عكسياً، مع ن-هد من tdT وC-هدفينوس من eGFP. ثانياً، إن التعبير عن إعادة التضمين في الخلية نفسها التي تحتوي على أشرطة MADM المستهدفة أمر ضروري. في غياب Cre، لا تعبر أشرطة الأحرف الشيمية عن eGFP الوظيفية أو tdT لأن تسلسلات الترميز الخاصة بها معطلة. مواقع اللوكسب بمثابة الهدف لإعادة التركيب المشترك بوساطة الكري، مما يؤدي إلى إعادة تشكيل كل من شرائط التعبير في وقت واحد. إذا حدث إعادة التعبئة أثناء مرحلة G2 من دورة الخلية متبوعة بالعزل X (G2-X)، فإن الخلايا اللتين تُرتِب كل منهما عن أحد البروتينات الفلورية. التنظيم الزمني لنشاط CreERT2 باستخدام تاموكسيفين (TM) يوفر معلومات دقيقة عن تاريخ ميلاد مستنسخات MADM وأنماط تقسيم ذريتها (الشكل 1A)29،46،47.
يمكن MADM يحتمل أن التسمية بشكل منهجي استنساخ الفردية مع ارتفاع دقة خلية واحدة في الدماغ الماوس مماثلة لأساليب تقليدية ولكن غير محددة وشاقة مثل Golgi تلطيخ48 أو صبغ ملء49. لأن المروج فقط القيادة CreERT2 يحدد نوع الخلية خصوصية التسميات MADM clonal، يمكن من حيث المبدأ أن تطبق لMMAD تتبع النسب التخفي في جميع أنحاء أي عضو مورين والأنسجة47،50،51،52. في الواقع، وقد استخدمت الدراسات بالفعل MADM للكشف عن علاقات النسب في استنساخ المستمدة من الأنسجة المتنوعة47،50،51،52،53،54،55،56،57،58،59. وقد تم تطبيق ماد النماذج التجريبية لدراسة النسب في الخلايا العصبية الإسقاط القشرية, جليا, والخلايا الجذعية بعد الولادة في تطور القشرة الجديدة7,,11,,12,,46,,60, 61,,6162,,,63,,64,,65. كما تم استخدام MADM لدراسة نسب الخلايا في الجيروسكوبات البالغة ، والمهاد ، وخلايا حبيبات cerebellar ، و interneurons على مستوى التخفي (انظر الجدول 1 للحصول على قائمة كاملة)47،53،54،56،57،66.
ومن السمات الفريدة لماد هي القدرة على ربط وراثيا الطفرات فصاطفة إلى واحد MADM كاسيت، وبالتالي خلق الفسيفساء الوراثية(الشكل 1B والشكل 2). هذا يؤدي إلى نوع البرية خلايا الابنة المسمى مع علامة فلورية واحدة (tdT في الشكل 1B)والأشقاء متحولة homozygous مع الآخر (eGFP في الشكل 1B)في بيئة غير مسمى heterozygous. MADM هي فريدة من نوعها في أن التحليل المقارن للسيطرة وsclones متحولة يمكن أن يؤديها في نفس بيئة الأنسجة في الجسم الحي. في الأصل، تم استهداف أشرطة MADM في موضع Rosa26 47، ولكن تحليل MADM لوظيفة الجينات كان يقتصر على الجينات منفصة إلى موضع. للتغلب (على الأقل جزئيا) هذا القيد وتوسيع إمكانيات التحليلات الجينية التي تستند إلى MADM، تم طرق أشرطة مادم في ما يقرب من تينتروميرس من Chr. 751, Chr. 1146, وChr. 1251. إن استهداف جميع الماوس 19 autosomes مع أشرطة MADM قيد التنفيذ وسيتيح دراسة أي جين تقريبًا في المستقبل ، مما يوفر منصة لا مثيل لها لدراسة علاقات النسب التنموية بالاقتران مع التحليل الجيني الوظيفي.
Protocol
تم مراجعة بروتوكولات الماوس من قبل مرفق أساسي قبل التخصصات المؤسسية (PCF) واللجنة الأخلاقية الداخلية في IST Austria. وقد أجريت جميع عمليات التكاثر والتجريب بموجب ترخيص وافقت عليه وزارة العلوم والبحوث الاتحادية النمساوية وفقا لقوانين الحيوانات النمساوية وقوانين الاتحاد الأوروبي.
1. تربية الفئران التجريبية لتحليل ماد وندور
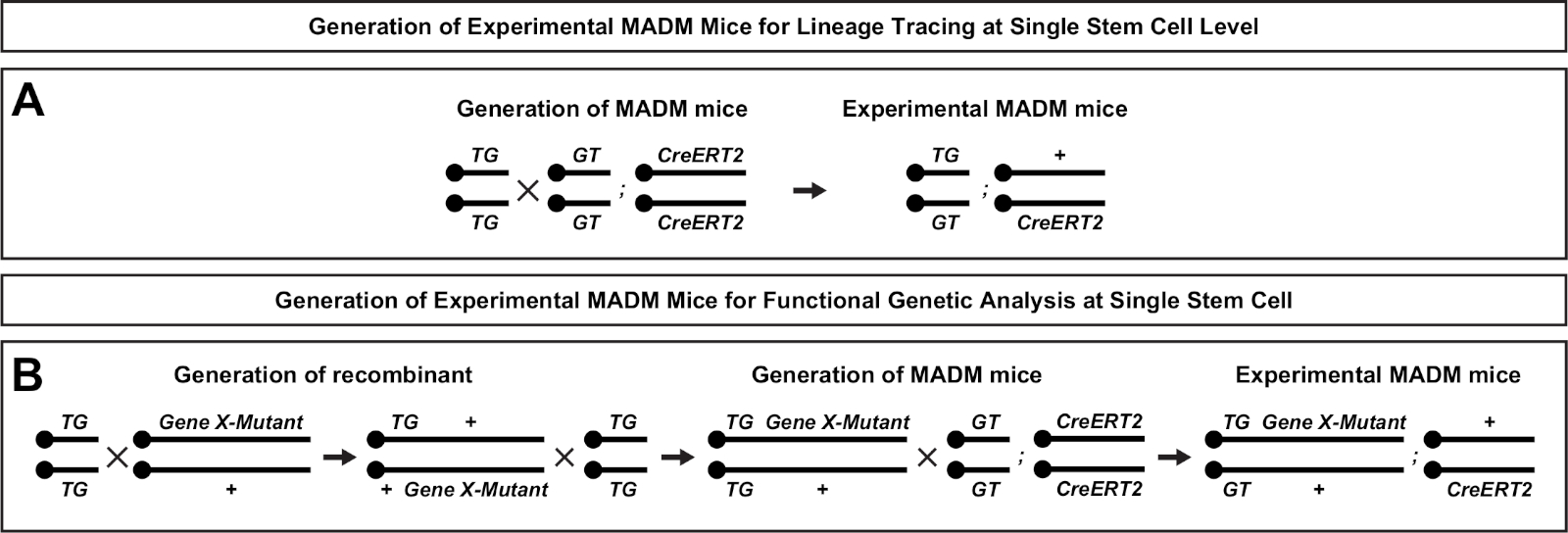
- إعداد التزاوج MADM التجريبية في الوقت المناسب (> P56; CD-1) في وقت متأخر من بعد الظهر (5:00 مساء) والتحقق من المقابس المهبلية صباح (8:00 صباحا). الصباح المكونات هو الحاضر تحسب كـ 0.5 اليوم. انظر الشكل 2 للحصول على نظرة عامة على إعداد التزاوج التجريبي للماوس. تأكد من أن نقاط زمنية لـ TM التعريفي لنشاط CreERT2 وتحليلها مناسبة لمعالجة الأسئلة التجريبية.
ملاحظة: للحصول على مزيد من المعلومات راجع الشكل 3 والنتائج التمثيلية أدناه. - بالنسبة لأخذ العينات بعد الولادة، قم بإعداد سلالات لتوليد الأمهات الحاضنات بالتوازي.
ملاحظة: يجب البدء في هذه االأيام حتى 1-2 قبل إنشاء سلالات تجريبية.
2. TM التعريفي في الفئران MADM
- إعداد 20 ملغ / مل TM حل العمل عن طريق حل في زيت الذرة في أنبوب أجهزة الطرد المركزي المخروطية 15 مل أو 50 مل ووضعه على منصة هزاز ل ~ 4 ح في درجة حرارة الغرفة (RT)، وضمان TM هو حل تماما. متجر حل العمل في 4 درجة مئوية مغطاة رقائق الألومنيوم واستخدامها في غضون 2 أسابيع.
- للحث على أحداث إعادة الدمج MADM، تقديم حقنة واحدة من TM intraperitoneally (IP) باستخدام حقنة 1 مل درنة 1 مل وإبرة 25 G في سد حامل في الوقت المناسب. اعتمادا على مرحلة تكوين العصب القشري، حقن TM بين E10-E15 بجرعة 1-2 ملغ/حامل السد. في وقت مبكر من النقاط الزمنية (أي E10) استخدام الحد الأقصى 1 ملغم / السد الحوامل (25 ملغ / كغ) لمنع المضاعفات أثناء الحمل11. بالنسبة للنقاط الزمنية بين E11-E15 استخدام 2 ملغم / السد الحوامل (50 ملغ / كغ)7.
ملاحظة: بدلا من ذلك، يمكن أن تدار TM مع gavage عن طريق الفم للحمل في وقت متأخر. - بالنسبة لتحليل MADM الزني إلى النقاط الزمنية بعد الولادة، استرد الأجنة الحية في E18-E19 من خلال الولادة القيصرية، ثم قم بتربية الجراء مع الأم الحاضنة.
ملاحظة: اعتمادا على الحالة الصحية للأنثى الحامل، قد لا يكون من الضروري إجراء عملية قيصرية ولكن لا يزال مطلوبا تربية الجراء مع الأم بالتبني لأن الأم الأصلية المعالجة TM قد تواجه مشكلة في المرض. - لاستعادة الأجنة الحية عن طريق الولادة القيصرية أو لاستعادة نقاط زمنية جنينية للتحليل، التضحية بالسد الحامل عن طريق خلع عنق الرحم.
- وضع الحيوان في وضعية supine وتطهير الفراء مع 70٪ الإيثانول. إجراء شق صغير في الجلد في أسفل البطن فوق الرحم باستخدام ملقط الجراحية والمقص. إجراء شق ثان من خلال العضلات وجدار البطن العضلي للكشف عن الصفاق.
- إزالة الرحم عن طريق فصل من الأنسجة المحيطة بها مع مقص. نقل الرحم سليمة على قفاز مملوء بالماء الدافئ لزيادة معدل بقاء الجنين حتى يتم إزالة كل من amnion بشكل فردي.
- استخدم مقصًا وأصابعًا رائعة لفتح جدران الرحم بعناية لإطلاق الأجنة. لا تقطع الحبال السرية قريبة جدا من الجسم لمنع فقدان الدم واسعة النطاق. إذا كان من المقرر استخدام الأجنة للتحليل، انتقل إلى الخطوة 3.9. إذا كان الجراء هي أن تكون برعاية، انتقل إلى الخطوة 2.8.
- إذا كان هناك حاجة إلى الحضانة، قم بتنظيف الجراء قبل نقلها إلى الأم الحاضنة. أثناء تنظيف الجراء، اضغط بلطف على الصدر من وقت لآخر لبدء التنفس. ضع مرة أخرى على قفاز ثان مملوء بالماء الدافئ لتحسين معدل البقاء على قيد الحياة.
ملاحظة: من المهم إزالة بلطف أي أمان و / أو المشيمة المتبقية مع منشفة ورقية. - قبل نقل الجراء إلى الأم بالتبني، إزالة الأم بالتبني من قفص لها، وإزالة الجراء الأصلي، واستبدالها مع الجراء التجريبية. أعيدوا الأم بالتبني إلى قفصها
ملاحظة: راجع مناقشة للحصول على اقتراحات إضافية لتحسين تعزيز معدلات القبول. - إذا كان genotyping مطلوبًا، اجمع خزعات زعابات زعانف اإلقذي أو الذيل بين P6-P8.
ملاحظة: تنفيذ هذه الخطوة فقط إذا التراخيص التجريبية الحيوانية الموافقة على هذه الممارسة.
3. إعداد الأنسجة لالماستنسخات MADM في الدماغ
ملاحظة: بالنسبة للتجارب التي تتضمن أنسجة ما بعد الولادة (≥P4)، انتقل إلى الخطوة 3.1. بالنسبة للنقاط الزمنية الجنينية ولاحقة الولادة المبكرة (P0−P3)، استمر في الخطوة 3.9.
- تخدير MADM التجريبي مع حقن IP من محلول الكيتامين/إكسيلازين/أسيمرامازين (65 ملغ، 13 ملغ، و2 ملغ/كغ من وزن الجسم، على التوالي)، وتأكد من أن الماوس لا يستجيب عن طريق قرص الكف الخلفي.
ملاحظة: يتم استخدام كل من الفئران MADM الذكور والإناث (خلفية CD-1) للتحليل. إذا كان genotyping مطلوبًا ، اجمع خزعات الأذن في هذه المرحلة. - وضع الحيوان تخدير في موقف سوبين على علبة جراحة الضخ وتطهير الفراء مع 70٪ الإيثانول. لبدء الجراحة، قم بعمل شق مع مقص وقطط جراحية بعناية من خلال الطبقة الخارجية للجلد ثم شق ثانٍ عبر طبقة العضلات. رفع غيض من القص وقطع النسيج الضام على الجانبين، مع توخي الحذر أكثر لتجنب قطع الكبد. التجويف الصدري سيكون مرئياً
- قص الحجاب الحاجز ورفع للكشف عن القلب. تقليم بعناية القفص الصدري ودبوس إلى علبة الجراحية لفضح القلب. للجراء، إزالة القفص الصدري تماما.
- أدخل إبرة مع الفوسفات المالحة المخزنة (PBS) في البطين الأيسر السفلي (الأنسجة الأكثر شحوباً). باستخدام مقص القزحية الصغيرة إجراء شق إلى الطرف الخلفي من الأذين الأيمن (الأنسجة الحمراء الداكنة) للدم لاستنزاف.
- أداء perfusion مع برنامج تلفزيوني تليها مباشرة من قبل الطازجة، والجليد الباردة 4٪ شبهformaldehyde (PFA) أعدت في برنامج تلفزيوني. بالنسبة للجراء (P4−P10) استخدم المحاقن لإجراء التسريب. ملء حقنة واحدة مع 10 مل من برنامج تلفزيوني وآخر مع 10 مل من 4٪ PFA. تأكد من إزالة جميع فقاعات الهواء في المحاقن. بالنسبة للحيوانات القديمة، استخدم مضخة مُتعالِقة.
- ابدأ في النفخ مع PBS (10 مل عند 2−4 مل/دقيقة في الجراء؛ 20 مل عند 4−6 مل/دقيقة للبالغين باستخدام مضخة مُتعالِمة). سوف يصبح الكبد واضحة صفراء شاحبة إذا تم وضع الإبرة بشكل صحيح.
- بمجرد اكتمالها، قم بإزالة الإبرة من الجراء وأدخل الإبرة التي تحتوي على PFA في نفس الحفرة. للبالغين، ووقف مضخة ال peristaltic قبل تبادل حل برنامج تلفزيوني مع الجليد الباردة PFA، والتأكد من تجنب فقاعات في امتصاص الأنابيب. استئناف النفخ مع PFA (10 مل في 2−4 مل/ دقيقة في الجراء؛ 30 مل في 4−6 مل/دقيقة للبالغين باستخدام مضخة م peristaltic).
- عندما يكون الضخ كاملاً، قم بقطع رأس الماوس وإزالة الدماغ من خلال تشريح دقيق. نقل الدماغ إلى 4٪ PFA. استخدم على الأقل 5x من أحجام الدماغ (أي 5−10 مل من PFA في أنبوب جهاز طرد مركزي مخروطي 15 مل) واحتضانها بين عشية وضحاها عند 4 درجات مئوية لتثبيت ما بعد الحقن لضمان التثبيت الكامل للأنسجة. متابعة الخطوة 3.10.
- بالنسبة للأنسجة الجنينية والأنسجة اللاحقة للولادة المبكرة (أي P0−P3)، بعد إجراء عملية قيصرية، قطع رأس الأجنة بمقص. إذا كان الجينوتيبينغ مطلوبًا، اجمع ذيل الجنين في هذه المرحلة. تشريح على الفور خارج الدماغ ونقلها إلى لوحة 12 بئر تحتوي على 2-3 مل من 4٪ PFA / جيدا. احتضان بين عشية وضحاها في 4 درجة مئوية ل postfixation.
- في صباح اليوم التالي تبادل PFA مع 10 مل (الكبار) أو 2−3 مل (جنين) من برنامج تلفزيوني وكرر غسل 3x لمدة 15 دقيقة في RT. نقل الأنسجة إلى محلول السكروز 30٪ في الفوسفات العازل (PB) وتخزينها عند 4 درجة مئوية على منصة هزاز حتى تغرق الأنسجة في المحلول.
- تضمين الدماغ في درجة حرارة القطع المثلى (OCT) مركب في قالب تضمين، مع الحرص على توجيه الدماغ لالقطع إما التاجي أو القوس. تجميد عن طريق وضع قالب التضمين على الجليد الجاف حتى تصبح أوكت معتمة تماما (~ 10−15 دقيقة). تخزين الأنسجة في -80 درجة مئوية حتى مزيد من الاستخدام.
4. إعداد أنسجة MADM للكيمياء المناعية
- إرفاق كتلة الأنسجة إلى القرص العينة في cryostat عن طريق تطبيق حلقة من أكتوبر إلى القرص ووضع كتلة مباشرة في أكتوبر عندما يبدأ في التجميد. تأكد من أن الكتلة موجهة بشكل صحيح لطائرة القطع المطلوبة.
ملاحظة: هنا، يتم وصف االقصاء المقطع للتحقيق في استنساخ MADM القشرية بالتفصيل. - تعيين درجة حرارة كتلة في التبريد إلى -20 درجة مئوية ودرجة حرارة النصل إلى -21 درجة مئوية.
- السماح للكتلة الأنسجة لضبط درجة حرارة الغرفة عن طريق تركيب القرص عينة إلى حامل العينة وترك في cryostat لمدة ~ 5 دقيقة قبل بدء تجزئة.
- تقليم كتلة في المقاطع سميكة (45-60 μm) حتى يتم الوصول إلى منطقة الأنسجة من الفائدة.
- بمجرد أن تكون حافة القشرة مرئية بوضوح ، توقف عن القسمة وأقفل النصل. تأكد من أن الشفرة محمية قبل تقليم الكتلة.
- تقليم الزائدة OCT المحيطة الأنسجة مع شفرة، وترك ~ 1-2 مم من أوكت على جميع جوانب الدماغ.
- توجيه المقبل كتلة بحيث يتم توجيه واحدة من الحواف الجانبي من القشرة إلى أسفل والآخر صعودا (أي، وأشار الحافة الأكثر من القشرة إلى اليمين).
- ابدأ بالقطع بسماكة 45 ميكرومتراً للمستنسخات البالغة و30 ميكرومتراً من أجل المستنسخات الجنينية. قم بأداء كل قسم على حدة واستخدم فرشاة صغيرة للحفاظ على المنطقة أسفل السكين نظيفة من أي حطام متبقي أثناء تشذيب الكتلة.
ملاحظة: إذا لم يتم ذلك ثم يقع مقطع قد يكون من الصعب تحديد الترتيب الصحيح من الشرائح. - إذا بدأت المقاطع في التجعيد، تقليم حواف كتلة و / أو ضبط بعناية لوحة الزجاج antiroll.
- بالنسبة لتحليل النسخ الجنيني، قم بتحميل المقاطع مباشرة إلى شريحة متجمدة. يجف على لوحة التدفئة في 37 درجة مئوية قبل الشروع مباشرة في الخطوة 5.6.
ملاحظة: يمكن إضافة عدة مقاطع إلى شريحة واحدة ولكن تأكد من الاحتفاظ بالترتيب التسلسلي. - لجمع المستنسخات البالغة، أعد 24 لوحة بئر تحتوي على 1 مل من PBS/well (عادة، 5−6 لوحات لكل دماغ). بدءا من البئر الأول ، مع ملقط الباردة جمع المقاطع التسلسلية الفردية في برنامج تلفزيوني في ترتيب القطاعات.
ملاحظة: يتم اعتماد طريقة المقطع العائمة للأنسجة البالغة لضمان عدم وجود أقسام ولا تحتوي على أي تجاعيد. - وقف القطاعات بمجرد الوصول إلى نهاية القشرة الجديدة.
- بالنسبة للمستنسخين البالغين، انتقل إلى الأجزاء العائمة المتصاعدة.
ملاحظة: يمكن الاحتفاظ بالقسم في برنامج تلفزيوني عند 4 درجة مئوية حتى 24 ساعة.
5. تصاعد أنسجة البالغين للتصوير
ملاحظة: مطلوب الأدوات التالية: فرشاة الطلاء الصغيرة، طبق بيتري، برنامج تلفزيوني مع 0.5٪ توين (PBS-T)، الشرائح التصاق (جدول المواد)،تصاعد المتوسطة(جدول المواد)،ويغطي، والملقط.
- ملء طبق بيتري مع برنامج تلفزيوني-T.
ملاحظة: يستخدم المنظفات للمساعدة في عملية التركيب. إذا كان التلطيخ لمستضدات إضافية حساسة للمنظفات (أي، بروتينات الجليكو) ضروري، فمن الأفضل لتخطي إضافة توين. - ضع شريحة التصاق في PBS-T بحيث يتم تغطيتها تقريبًا إلى الملصق.
- نقل القسم الأول إلى برنامج تلفزيوني T.
- باستخدام فرشاة طلاء صغيرة، مناورة المقطع على الشريحة وترتيبها للحفاظ على ترتيب القطع. المضي قدما في نفس الطريق مع جميع أقسام أخرى.
- بمجرد أن تكون كل المقاطع في موضعها، ضع الشريحة (~12−16 مقطع/شريحة) في غرفة شرائح داكنة. ارفع الغطاء قليلاً للسماح للقطع بالجفاف بالكامل (~ 10−20 دقيقة)، مع ضمان التزامها في الخطوات اللاحقة.
- إذا كان أداء المناعة الهيستوتكيميا للمستضدات إضافية، انتقل مباشرة إلى القسم 6 أو 7.
ملاحظة: بالنسبة للنقاط الزمنية الجنينية، من الضروري تنفيذ خطوات مناعة لـ GFP وtdT على الأقل (القسم 6). بالنسبة للمستنسخات البالغة، لا يلزم ذلك إلا إذا كان التلطيخ من أجل مستضدات إضافية بالتوازي (القسمان 6 و7). - إعادة هيدرات وغسل أقسام 1x مع 1X PBS لمدة 5 دقائق لإزالة المتبقية PBS-T. تطبيق ~ 1 مل من 4′، 6-diamidino-2-فينيليندول (DAPI) المخفف في 1X PBS (1 ميكروغرام / مل) إلى الشريحة، وضمان تغطية جميع المقاطع واحتضان لمدة 15 دقيقة.
- قم بإزالة DAPI بعناية واغسل 1x مع PBS 1x لمدة 5 دقائق. ختم مع 24 × 60 مم يغطي والسماح الجافة لمدة 3 ساعات على الأقل قبل التصوير.
6. المناعة لGFP وtt فقط
ملاحظة: هذا المقطع ضروري من أجل استنساخ جنينية.
- ضع الشرائح أفقيًا في غرفة حضانة شرائح مرطبة. وضع علامة على حدود الشريحة مع علامة الشمع لتقليل مقدار المخزن المؤقت المطلوب.
- إعادة هيدرات المقاطع مع 1x PBS. لتحسين جودة التلوين، اعمل مع الأنسجة الطازجة.
- أضف 250−400 ميكرولتر من العازلة العازلة (0.5% تريتون X-100، 2−3% مصل الحمير العادي في 1x PBS) لكل شريحة، مما يضمن تغطية جميع الأقسام. احتضان لمدة 1 ساعة.
ملاحظة: يختلف تركيز المنظفات (Triton X-100 أو Tween-20) وفقًا للأجسام المضادة الأولية الإضافية المستخدمة لأن بعض المستضدات أكثر حساسية للمنظفات من غيرها. - إزالة العازلة العازلة وإضافة الأجسام المضادة الأساسية في العازلة حظر إلى الشريحة (300-400 ميكرولتر / شريحة).
ملاحظة: مثال على رد فعل الأجسام المضادة الأولية القياسية لمضادات GFP/anti-tdT (MADM) يمكن استخدام الدجاج المضادة لـ GFP (1:500) والأرانب المضادة لـ RFP (1:500). - احتضان مع الأجسام المضادة الأولية بين عشية وضحاها في 4 درجة مئوية.
ملاحظة: يجب أن تكون الشرائح محتضنة أفقياً تماماً مع المخزن المؤقت الذي يغطي كافة المقاطع. وإلا، قد يؤدي إلى تلطيخ غير متساو أو ضعيف. - تأكد من صباح اليوم التالي أن العازلة حظر مع الأجسام المضادة الأساسية لا تزال تغطي جميع المقاطع على الشريحة. إذا لم يكن كذلك، كرر خطوة الحضانة لمدة 3−4 ساعة في RT.
- إزالة الأجسام المضادة الأولية وغسل 4x مع 1X برنامج تلفزيوني لمدة 10 دقيقة في RT.
- إضافة الأجسام المضادة الثانوية المخففة في العازلة حظر إلى الشريحة (300−400 ميكرولتر / شريحة): اليكسا فلور 488 المضادة للدجاج IgG (1:500) وCy3 المضادة للأرانب IgG (1:500).
- احتضان في RT لمدة 2 ساعة. حافظ على الشرائح المغطاة من الضوء لمنع تبييض الفلوروفوريس.
- إزالة الأجسام المضادة الثانوية وغسل 2x مع 1X PBS لمدة 10 دقيقة.
- احتضان مع DAPI المخفف في برنامج تلفزيوني (1:5,000) لمدة 15 دقيقة.
- غسل أقسام 1x مع 1X برنامج تلفزيوني لمدة 10 دقيقة.
- إزالة برنامج تلفزيوني الزائدة وجافة ل ~ 1−2 دقيقة قبل تضمين في 110 ميكرولتر من متوسطة التركيب.
- ختم مع 24 × 60 مم يغطي والسماح الجافة لمدة 3 ساعة على الأقل قبل التصوير. الصورة تنزلق في غضون 1-2 أسابيع بعد إجراء الكيمياء المناعية لضمان إشارة الأمثل.
7. المناعة لGFP، tdT، ومضاد إضافي
- تنفيذ الخطوات 6.1−6.3.
- إزالة العازلة العازلة وإضافة الأجسام المضادة الأساسية في العازلة حظر إلى الشريحة (300-400 ميكرولتر / شريحة).
NOTE: When staining for three or more antigens (i.e., GFP, tdT, and a protein of interest) and the antibody for the protein of interest was raised in rabbit, it is recommended to use the anti-tdT (goat) primary antibody at a dilution of 1:500. مثال على رد فعل الأجسام المضادة الأولية لثلاثة مستضدات مع تلطيخ TdT بديل يمكن استخدام الدجاج المضادة لـ GFP (1:500) ، والماعز المضادة tdT (1:500) ، والأجسام المضادة ضد البروتين من الفائدة (أي ، أرنب). - تنفيذ الخطوات 6.5−6.7.
- أضف مزيج الأجسام المضادة الثانوي المخفف في عازلة الحجب إلى الشريحة (300−400 ميكرولتر/شريحة): Alexa Fluor 488 Anti-chicken IgG (1:500)، Cy3 المضادة للماعز IgG (1:500)، و Alexa Fluor 647 Anti-Rabbit IgG (1:500).
- تنفيذ الخطوات 6.9−6.14.
8. كونفوكال الحصول على الصور و الكم من استنساخ MADM
- تحديد وتوثيق أقسام الدماغ التي تحتوي على استنساخ ومواقعها في القشرة.
ملاحظة: سوف يختلف عدد المقاطع التي تمتد باستنساخ استناداً إلى عندما تم استحثاث clone برنامج تشغيل T2 CreER ووقت التحليل. يمكن تنفيذ هذه الخطوة إما على المجهر confocal أو مجهر epifluorescence. - باستخدام مجهر مقلوب، ابدأ باختيار وتكوين خطوط الليزر والفلاتر الصحيحة. بالنسبة لأدمغة MADM، حدد DAPI وGFP وtdT (الإثارة: 358 نانومتر، 488 نانومتر، و554 نانومتر، على التوالي؛ ذروة الانبعاثات: 461 نانومتر، 507 نانومتر، و 581 نانومتر، على التوالي). تأكد من ضبط الثقب على وحدة واحدة ذات هواء هواء للحصول على جودة تصوير مثالية.
- بالنسبة للإعدادات المحددة confocal، تستنسخ الصور مع هدف 20x والتكبير 1x. بالنسبة للصور التي سيتم استخدامها في القياسات الكمية، استخدم بكسل سرعة المسح الضوئي بقيمة 1.52−2.06 ميكرومتر (القيم 7−8 في برنامج الحصول على الصور) بدون متوسط. ضبط كثافة الليزر والحصول على الإعدادات لكل قناة حسب الاقتضاء.
ملاحظة: قد تختلف إعدادات سرعة المسح والمتوسط وفقًا لجودة الصورة المطلوبة. - بمجرد أن يتم تحديد استنساخ بوضوح، وترتيب البلاط التصوير لتغطية جميع الأقسام ذات الصلة في استنساخ. ضبط z-المكدس بحيث يتم التقاط جميع الخلايا المسمى MADM في استنساخ مع فاصل زمني من 1.5 μm / z- شريحة المكدس. ضبط المنطقة من البلاط بحيث يتم التقاط عرض كامل من القشرة عند تصوير استنساخ (أي، من سطح بصال إلى اللواهيم.
- صورة استنساخ الفردية التي تمتد أقسام متعددة على التوالي ، وضمان أن أي أقسام دون خلايا داخل استنساخ لا تزال imaged لغرض إعادة الإعمار 3D والتفسير الصحيح للمعلومات الفضائية الخلية.
- تحليل كل قسم يحتوي على خلايا من استنساخ MADM بالتسلسل من الرسترالية إلى نهاية السد من القشرة. تمييز الخلايا العصبية الفردية وglia على أساس مورفولوجيا و / أو وصمة عار. تسجيل المعلومات الموضعية بالتوازي بناءً على حدود الطبقة الخاصة التي يحددها التلطيخ النووي (DAPI).
ملاحظة: انظر الشكل 4 للاطلاع على النتائج التمثيلية للتحليل الجنيني والشكل 5 للاطلاع على النتائج التمثيلية لتحليل الكبار.
9. المسلسل 3D إعادة الإعمار من المستنسخين
ملاحظة: إعادة بناء 3D من المستنسخين الفردية على صور عبر أقسام الدماغ المسلسل مفيد للعرض البصري وكذلك تحليل 3D clonalالهندسة ويمكن القيام بها وفقا للخطوات التالية.
- غرزة وصهر الصور البلاط confocal على أساس المعلمات اكتساب باستخدام برنامج الحصول على الصور. افتح ملف .czi ثم قم بتشغيل أسلوب خياطة ضمن علامة التبويب معالجة في برنامج ZEN (زيس).
- تصدير مكدسات الصور المخيطة كـ planes z فردية بتنسيق TIFF. افتح ملف .czi مخيط ثم قم بتشغيل أسلوب تصدير الصورة ضمن علامة التبويب معالجة. بالنسبة للصور متعددة القنوات، قم بتصدير الصور كـ أحمر/أخضر/أزرق لمعالجة الصور اللاحقة.
- كرر الخطوات 9.1 و 9.2 لكل مقطع الدماغ التسلسلي من استنساخ.
ملاحظة: لإعادة بناء 3D دقيقة، يجب أيضا معالجة جميع أقسام الدماغ داخل استنساخ، بما في ذلك تلك التي لا تحمل علامات الخلايا. - سلسلات الصور الفردية في كومة واحدة في النظام بدءا من معظم أكثر من روسيرال إلى معظم caudal ض الطائرة باستخدام برنامج معالجة الصور مفتوحة المصدر مثل ImageJ / فيجي67،68.
ملاحظة: يجب إزالة أي صور فارغة عند حواف كل قسم من أقسام الدماغ عند هذه النقطة. - إذا لزم الأمر، قم بتصحيح رصة الصورة التي تم الحصول عليها من الخطوة 9.4 لعدم المحاذاة باستخدام مكون مساعد ImageJ يسمى"MultiStackReg"باتباع الخطوات 9.5.1−9.5.5. إذا لم تكن محاذاة الصورة مطلوبة، انتقل إلى الخطوة 9.6.
ملاحظة: يقوم هذا المكون الإضافي بمحاذاة الصورة للقناة بأعلى تباين (عادةً DAPI) ثم يطبق التحويل المسجل على القنوات الأخرى، مما يسمح بمحاذاة صورة موثوقة لكدسات متعددة القنوات. المساعد المساعد يسمى"TurboReg"يجب أن تكون مثبتة مسبقا.- في ImageJ، قم بتثبيت"MultiStackReg"و"TurboReg"الإضافات.
- افتح رصة الصور المستنسخة التي تم الحصول عليها من الخطوة 9.4 لمحاذاتها. تقسيم القنوات إلى DAPI (الأزرق) وGFP (الأخضر) وtdT (أحمر) ضمن خيار اللون ضمن علامة التبويب صورة.
- تشغيل "MultiStackReg" البرنامج المساعد لمحاذاة قناة DAPI من قبل "جامدة الجسم" التحول وحفظ ملف التحول.
- تطبيق ملف التحويل المحفوظة إلى قناتين أخرى باستخدام"MultiStackReg".
- دمج كافة القنوات الثلاثة التي تمت محاذاتها وحفظ المكدس المحاذى.
- لتوجيه النسخ في ImageJ تدوير كومة من الصور استنساخ التي تم الحصول عليها من الخطوة 9.4 (أو الخطوة 9.5.5 بعد المحاذاة) في اتجاه عمودي مع سطح بصال في الأعلى و callosum corpus في أسفل. احصد في المستوى س ص إذا لزم الأمر.
- لكل من العرض النوعي والتحليل الكمي، قم بإنشاء صورة عرض ض أقصى (الخطوة 9.8) أو تنفيذ عرض ثلاثي الأبعاد (الخطوة 9.9) من النسخة.
- في ImageJ، افتح كومة الصور من الخطوة 9.6 وحدد خيار الإسقاط Z مع نوع العرض ماكس كثافة. وهذا سوف يولد صورة من استنساخ كامل المتوقعة على نفس المستوى.
- في ImageJ، افتح كومة الصور من الخطوة 9.6 وحدد وظيفة 3D Project z لإنشاء مرئي ثلاثي الأبعاد للنسخة التي يمكن تدويرها.
ملاحظة: من المهم في هذه الخطوة إدخال الفاصل الزمني شريحة الصحيح مكافئ لسمكة من مكدسات z الفردية أثناء الحصول على الصورة. وينبغي استخدام أداة الاستيفاء لإزالة الفجوات بين الشرائح.
النتائج
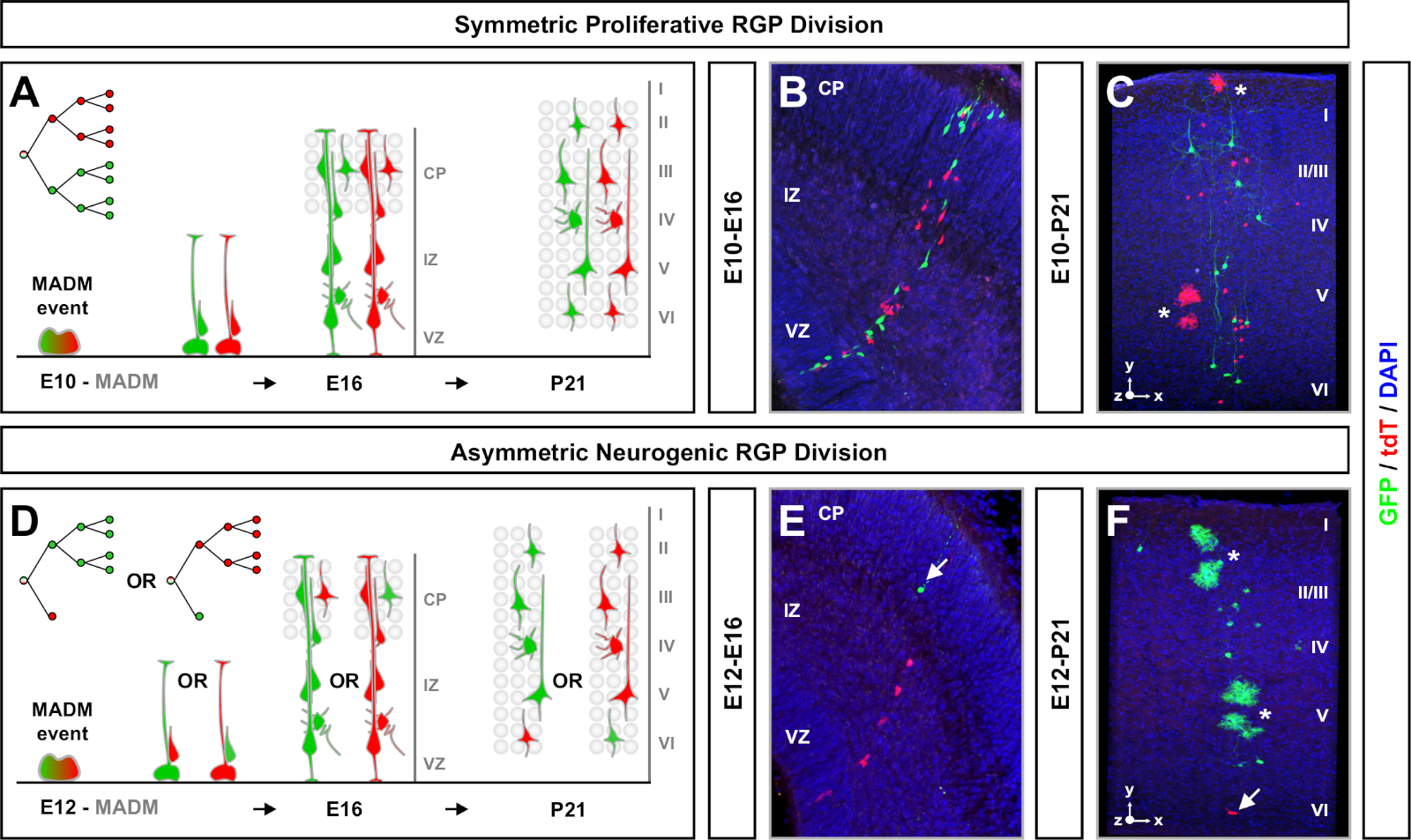
نتائج MADM في إعادة تشكيل البروتينات الفلورية الخضراء والأحمر الوظيفية مع اثنين من الخلايا ابنة كل التعبير عن واحد من اثنين من البروتينات الفلورية على G2-X أحداث الفصل الكروموسوم (الشكل 1أ). لأن الأحداث MADM يؤدي إلى وضع علامات دائمة ومتميزة من اثنين من النسب الهجرت، يمكن إجراء تقييم كمي من نسل خلية ابنة الأخضر والأحمر (subclones). ويمكن تحديد المتغيرات بما في ذلك نمط التقسيم (على سبيل المثال، متماثل مقابل غير متماثل) وإمكانات (مثل عدد النسل) من السلف الأصلي. إن تحديد كل 1 من المستويات الفرعية المسماة بالفلورية مفيد عندما يحدد بأثر رجعي ما إذا كانت الخلية السلف الأصلية تمر بانقسامات تكاثرية متماثلة، أو أقسام عصبية غير متماثلة في وقت تحريض TM. الدراسات السابقة مجمعة Emx1-CreERT2 أو Nestin-CreERT2 المشتقة استنساخ الإسقاط مثير في القشرة إلى فئتين واسعة7,11,46. الأول، الذي يطلق عليه "استنساخ متناظرة التكاثرية"، تتألف في المتوسط من عدد كبير من الخلايا العصبية، مع كل من subclones الأخضر والأحمر تحتوي على أربعة أو أكثر من الخلايا العصبية لكل منهما. المجموعة الثانية ، "استنساخ غير متناظرة" يعرف فئة من المستنسخين حيث "الأقلية" subclone يحتوي على أقل من ثلاث خلايا عصبية و "الأغلبية" subclone ، أربعة أو أكثر11. هذه التعاريف محددة لRGPs القشرية وربما تحتاج إلى إعادة النظر لمناطق الدماغ الأخرى والأنسجة. لكل من فئتي المستنسخات القشرية، سيتم توزيع ذرية في جميع أنحاء طبقات سطحية وعميقة.
عند تصميم الدراسات المُنَسَة في MADM، هناك عدد من الجوانب التي يجب أن تؤخذ في الاعتبار. الوقت الذي يتم فيه حدوث أحداث MADM من قبل إدارة TM هو اعتبار رئيسي (الشكل 3). لالقشرية excitatory إسقاط الخلايا العصبية MADM استنساخ (أي، باستخدام Emx1-CreERT2 أو نستين-كريترT2)في E10، تقريبا جميع RGPs كانت لا تزال تمر الانقسامات المتماثلة11. لذلك ، التقط الحث في E10 مع TM جولات متعددة من تضخيم RGP التكاثري وأسفرت عن استنساخ مع أعداد خلايا عصبية عالية. ومع ذلك، فإن عدد RGPs في E10 كان صغيرًا بشكل عام ، وبالتالي ولدت إدارة TM عددًا قليلًا جدًا من أحداث MADM (في بعض الأحيان أقل من واحد لكل دماغ). تحولت غالبية RGPs من الانقسامات العصبية المتماثلة إلى الانقسامات العصبية غير المتماثلة في جميع أنحاء E12. لاستهداف استنساخ عصبي غير متماثل تماما، كان من الأفضل للحث على E12 أو في وقت لاحق(الشكل 3). كان الوقت بين تحريض TM ومراقبة أحداث إعادة التشزيم MADM في القشرة يميل إلى أن يكون أقل من 24 h. حقن IP كانت الطريقة المفضلة لإدارة TM في المراحل الجنينية لهذه الطريقة لأنها أدت إلى زيادة قابلية التكاثر في الحث الزني. من المهم أيضا للحفاظ على الجرعة TM إلى الحد الأدنى لسببين. أولاً، إذا زاد معدل إعادة التركيب في MADM، فإن احتمال تحفيز المستنسخات المتعددة، وربما المتداخلة، يكون أعلى. ثانياً، إذا تم تسليم الكثير من TM، يمكن ملاحظة معدل متزايد للإجهاض، وإعادة الامتصاص الجنين، وأحجام القمامة الأصغر. وقد لوحظت عمليات الإجهاض في ما يقرب من نصف جميع السدود الحامل عندما تم تسليم حقن TM في E10. انخفض هذا تردد من [إ11] فصاعدا ويقلّل إلى تقريبا 1/3 من حامل سدود إجهاض. للحصول على ملخص لجرعات TM وأوقات الحث ، وكريرT2 السائقين المستخدمة في الدراسات السابقة MADM ، راجع الجدول 1. وقد لوحظ نشاط المراسل في غياب TM مع بعض TM-inducible CreERT2 السائقين69. لم يلاحظ التعبير خارج الرحم أو MADM أحداث إعادة الكومبينيشن في غياب TM مع Emx1-CreERT2 من السائقين نستين-CreERT2. قد يرجع ذلك جزئيًا إلى حقيقة أن الكروموسومات عبر الكومبوسومات التي يتم بوساطة TM تحدث في حوالي 1:1,000 إلى 1:10,000 تردد أقل من إعادة الكومبينيشنات رابطة الدول المستقلة، مما يقلل من احتمال وضع علامات MADM خارج الرحم.
عامل آخر يجب مراعاته عند التخطيط لتجربة تحليل ية منوم هو مدة الدراسة. تغيير طول الوقت بين التعريفي TM وعندما تم تحليل التجربة (A) (نافذة الوقت) يعرض ديناميات الخلايا الجذعية على مر الزمن64. الإطارات الزمنية الجنينية القصيرة (أي TM/E11-A/E13؛ TM/E11−A/E16) التقط ديناميات النشوء العصبي الجنيني(الشكل 4). مقارنة استنساخ من اثنين أو أكثر من النوافذ الوقت يوفر نظرة كمية في عدد الخلايا المنتجة وكيف تختلف توزيع الخلايا العصبية في مراحل مختلفة من تقدم النسب64. لالتقاط كامل إمكانات استنساخ الفردية، فمن الضروري تمديد النافذة الزمنية تحليلها في نقاط زمنية ما بعد الولادة أو الكبار7،11،12. وترد أمثلة على استنساخ النيكورني المستحث في الجنين وتحليلها في الكبار في الشكل 5. لاحظ، يتم الانتهاء في الغالب من تكوين العصب القشري ويزيد من تكوين الدبقية من قبل E17. تقريبا 1/6 العصبية RGP أيضا المضي قدما لتوليد الخلايا الفلكية و / أو oligodendrocytes11.
استنساخ متماثل يحدث عندما RGPs الخضوع لجولات واحدة أو أكثر من شعبة التكاثر11. وكانت استنساخ RGP التي تم الحث عليها بين E10-E12 في المتوسط أكبر في الحجم وقدمت المزيد من الميزات المكانية لتوزيع الخلايا العصبية النهائي(الشكل 4A-C). استغرق استنساخ مع الخلايا العصبية موزعة بالتساوي نسبيا في جميع أنحاء طبقات عميقة وسطحية على شكل "اسطوانة" في حين أن استنساخ مع الخلايا العصبية أكثر تشتتا في طبقات سطحية من طبقات أعمق وضعت شكل "مخروط"11. لالتقاط المعلومات المكانية والمورفولوجية للمستنسخة بشكل كامل ، كان من الضروري إعادة بناء كل استنساخ حسابيًا باستخدام صور متسلسلة. لقياس التشتت الزني، قورن التشتت الجانبي الأقصى (الذي يقاس بجميع الأبعاد) في طبقات سطحية (LII-VI) من استنساخ بتشتت الخلايا العصبية في الطبقات العميقة (LV/LIV). وقدمت هذه النسبة (التوزيع الأعلى: التوزيع الأدنى) قراءات قابلة للقياس الكمي لشكل الاستنساخ الكلي.
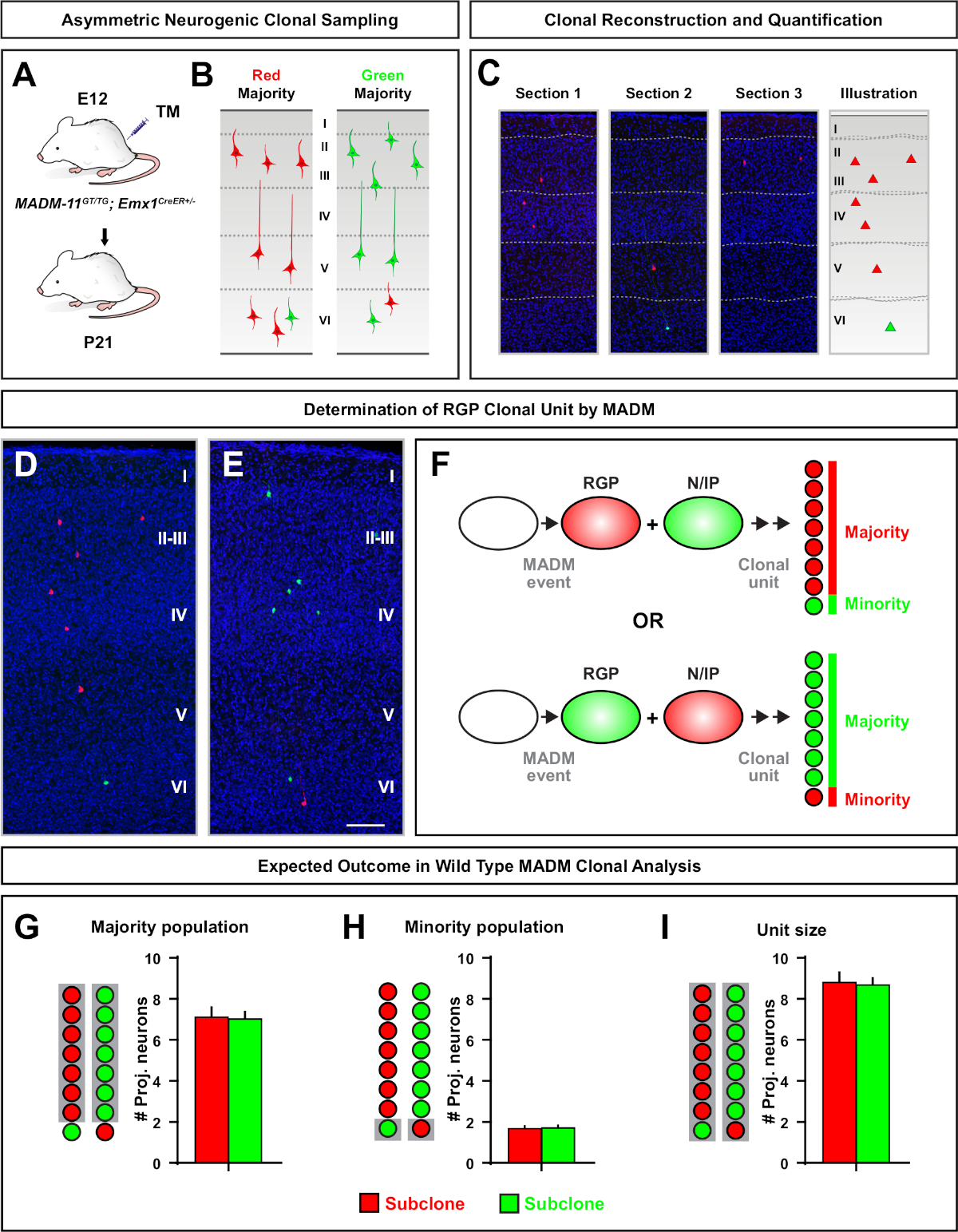
استنساخ غير المتماثلة، حيث كان subclone الأقلية ثلاثة أو أقل، قدمت نظرة ثاقبة في الناتج العصبي من RGP واحد(الشكل 4D-F والشكل 5ألف-F)7،11،12. ويمكن تسمية أغلبية السكان (تحت التكلون الكبير) إما باللون الأحمر أو الأخضر، بمتوسط يبلغ حوالي سبعة خلايا عصبية الإسقاط مثير لكل استنساخ عند تحريض باستخدام Emx1-CreERT2 أو نستين-كريترT2(الشكل 5G)7,11,12. ويمكن تشريح العدد الإجمالي للخلايا في استنساخ MADM عن طريق تحليل توزيع الخلايا العصبية في subclone كبيرة عبر طبقات سطحية وعميقة. وقد تم تصنيف سكان الأقليات (تحت الباطن الصغيرة) باللون المتبادل وكان في المتوسط 1−2 خلية لكل استنساخ(الشكل 5حاء). ويمكن حساب إجمالي "حجم الوحدة"، الذي كان في المتوسط 8-9 من الخلايا العصبية، عن طريق إضافة الثياب الفرعية الصغيرة والكبيرة معاً(الشكل 5I)7،11،12. من المهم أن نلاحظ أنه في حين أن الناتج العصبي من RGPs كان يمكن التنبؤ بها إلى حد كبير، كان هناك درجة من عدم التجانس12،70.
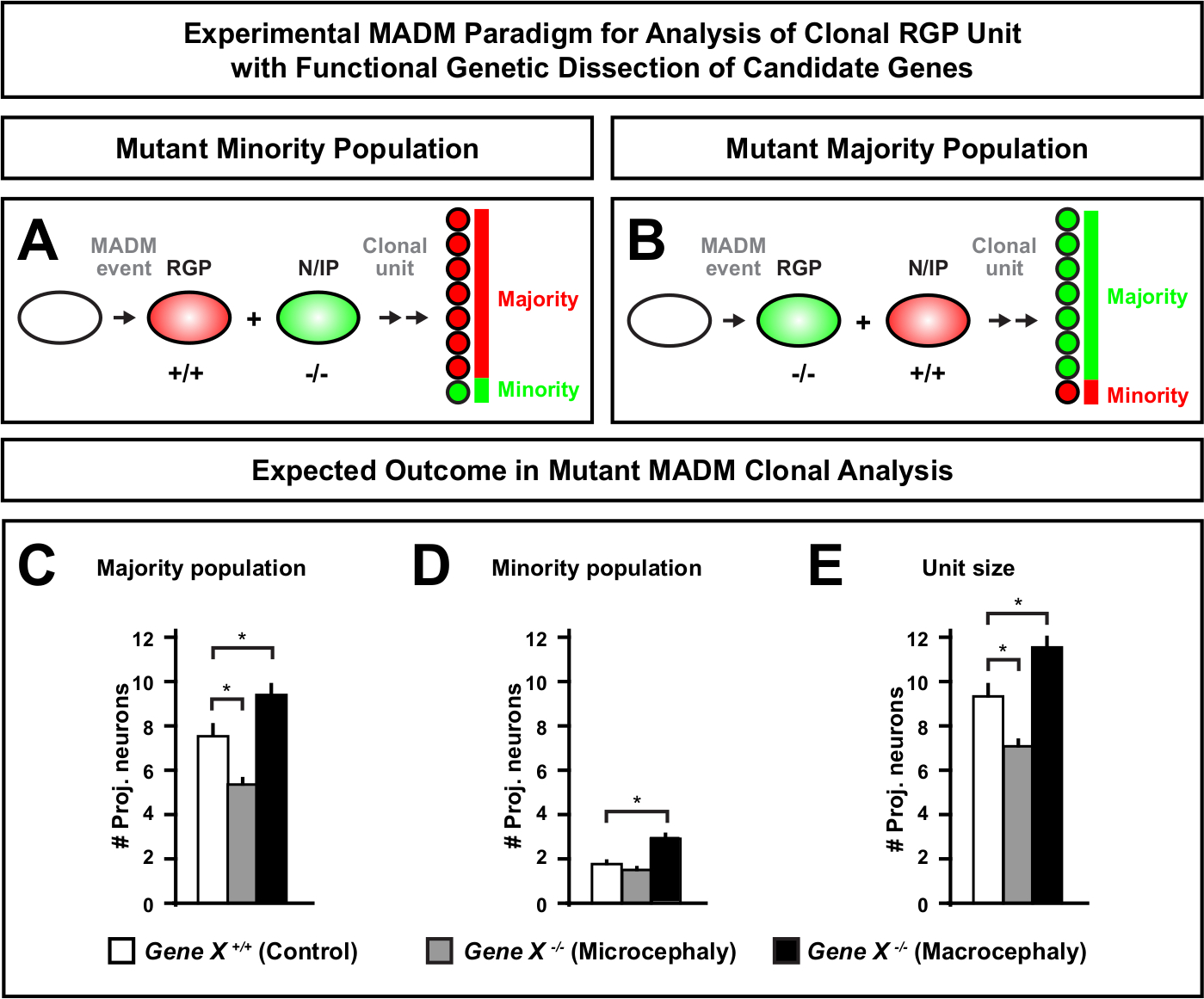
إدخال طفرة في الكاسيت MADM تمكن من توليد الفسيفساء الجينية، وتوفير طريقة فريدة من نوعها لتشريح المنظمين الجزيئية لتطور نسب الخلايا الجذعية. على هذا النحو، يوفر MADM منصة تجريبية لا مثيل لها لدراسة وظيفة الخلية المستقلة للجين (على سبيل المثال، ارتباطها بصغر الرأس أو الرأس الكلي). بمقارنة المستنسخات المستحثة في الفسيفساء الجينية MADM إلى المستنسخات المستحثة في التحكم MADM ، يمكن توليد قراءة كمية عالية للتغيرات في أعداد الخلايا العصبية والتوزيع. وقد حددت الدراسات السابقة المستندة إلى MADM وظيفة الخلايا المستقلة من Otx1 في تكوين الرأس الصغير على المستوى السفلي (انظر الشكل 6A-E للحصول على مثال تمثيلي)11. في دراسة أخرى, أظهر تحليل ماد النحلي أن Ndel1 لا ينظم الخلية بشكل مستقل أرقام الخلايا العصبية الإسقاط, ولكن بدلا من ذلك قدرة الخلايا العصبية حديثي الولادة على الدخول أو الهجرة داخل لوحة القشرية, التي تشكل في وقت لاحق قشرة الكبار46. أظهرت هذه الدراسات الطبيعة الكمية للغاية للتحليل المُدور في MADM في دراسة وظائف الخلايا المستقلة للجينات التي تنظم التنمية القشرية. لا توجد حاليا أمثلة في الأدب باستخدام MADM لدراسة الجينات المتورطة في جاهمة الرأس على المستوى السفلي. ومع ذلك، في الدراسات المستقبلية تحليل الجينات ذات الصلة للسيطرة على حجم القشرية بشكل عام يمكن أن توفر رؤى مرغوبة للغاية على المستوى الجزيئي والخلوي.

الشكل 1: مبدأ MADM لتتبع النسب وتحليل التخفي على مستوى الخلايا الجذعية المفردة. (أ) لإجراء تتبع النسب وتحليل التخفي مع MADM ، يجب وجود مكونين. أولاً، يجب أن تكون أشرطة MADM موجهة إلى بؤر متطابقة على الكروموسومات المتماثلة. تتكون الكاسيتات من جينتين مراسلين من نوع chimeric fluorescent، eGFP (الأخضر، [G]) وطماطم الخافتة جنبا إلى جنب (أحمر، tdT[T]). يحتوي الكاسيت GT على N-terminus eGFP وC-terminus من tdT، مفصولة بـ intron يحتوي على موقع loxP. تم بناء كاسيت TG عكسياً، مع ن-هد من tdT وC-هدفينوس من eGFP. ثانياً، يجب أن يحدث تعبير إعادة الكومبينيز Cre في الخلية التي تحتوي على أشرطة MADM المستهدفة. مواقع اللوكسب بمثابة هدف لإعادة التركيب المشترك بوساطة الكري، مما يؤدي إلى إعادة تشكيل كل من شرائط التعبير في وقت واحد. إذا حدث إعادة الكومبين أثناء مرحلة G2 من دورة الخلية متبوعاً بالعزل X (G2-X)، فإن الخلايا اللتين تُبنِين ستعبران عن أحد البروتينين الفلورسنتين. (ب) مبدأ MADM لتحليل الفسيفساء الجينية على مستوى استنساخ واحد. يمكن إدخال أليل متحولة (طفرات نقطة، حذف، إدراج، أليلات مشروطة محاطة بلوكسب كما هو مبين في الشكل 1B، إلخ) إلى شريط TG-MADM عبر إعادة التركيب الميوائية (انظر الشكل 2 و هيبينماير وآخرون46 للحصول على تفاصيل حول كيفية إدخال أليل متحولة في نظام MADM). إذا كان G2-X كري إعادة الكومبيناسي بوساطة interchromosomal عبر إعادة التشتيل يحدث بين أشرطة MADM فإنه يؤدي إلى خلية متحولة واحد GFP + homozygous(GeneX-/-) للجين من مصلحة واحد tdT + homozygous خلية نوع البرية (GeneX+ / +) في بيئة غير موصومة heterozygous46,47,71. وقد سبق وصف نتائج وضع العلامات البديلة غير المستخدمة في التحليل النيالي (أي الخلايا الصفراء) بالتفصيل11و46,,و47. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 2: مخططات التربية لتوليد فئران MADM التجريبية لتتبع النسب. نظام تربية لتوليد السيطرة MADM (A) وجين X MADM (B) فئران MADM التجريبية لتحليل التخفي. لمزيد من المعلومات حول نماذج التربية MADM انظر بيتي وآخرون7 و هيبينماير وآخرون7،46. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 3: نماذج الدورات الزمنية لتحليل النسب التخفي القائم على MADM. التخطيطي من الإطارات الزمنية التصميم التجريبية. وبالنسبة لنماذج أخذ العينات الطولية، ظلت النقطة الزمنية لاستقراء الاستنساخ ثابتة، وتفاوت طول الفترة الزمنية قبل التحليل. وفي أخذ العينات الفاصلة التدريجية، ظلت نقطة التحليل ثابتة، ولكن وقت الحث يتفاوت. ويمكن استخدام مزيج من نهج واحد أو كلا النهجين تبعاً للأسئلة التي تم تناولها. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 4: تحليل الاس في القشرة الجديدة النامية والبالغة. TM-بوساطة MADM استنساخ التعريفي في التكاثرية بشكل متناظر (TM في E10) (A-C)واعصاب غير متناظرة (TM في E12) (D-F) تقسيم RGPs. يصور هي مستنسخات MADM الفردية في الجسم الحي في النامية (TM/E10−A/E16 و TM/E12−A/E16)(B,E)والبالغة (TM/E10−A/P21 و TM/E12−A/P21)(C,F)في MADM-11GT/TG; نستين-كريسT2+/- (B, E) وMADM-11GT/TG; Emx1-CreERT2+/- (C, F). وكان الناتج الخلايا العصبية مستقلة عن لون subclone والأغلبية الخضراء / الأقلية subclones يمكن مقارنتها بالأغلبية الحمراء / أقلية subclones تحت ظروف السيطرة7،11. تقريبا 1/6 من استنساخ الكبار تحتوي أيضا على استروبيات و / أو oligodendrocytes، المشار إليها من قبل النجمة البيضاء. ويعاد إنتاج اللوحات باء ووا بإذن من هيبنماير وآخرون46 ورولاندز وسيمونز72على التوالي. CP = لوحة كورتيكال. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 5: تحليل MADM التخفي لتحديد كمية الناتج العصبي بوساطة RGP. تحليل الخلايا العصبية مثير (وحدة) الإنتاج من قبل RGPs العصبية الفردية على مستوى التخفي باستخدام MADM7,11. (أ) نموذج تجريبي لتحفيز معظم المستنسخات MADM غير المتماثلة في القشرة النامية. (ب)نتائج استنساخ غير متناظرة ممكن مع الأغلبية subclone المسمى إما الأخضر أو الأحمر (C) تمثيلية أقسام متتالية تمتد استنساخ واحد neurogenic غير متماثل (D, E) 3D صور إعادة الإعمار من ممثل غير متماثل G2-X MADM استنساخ مع غالبية السكان في الأحمر (D) أو الأخضر (E) في MADM -11GT / TG; Emx1-CreERT2+/- مع التعريفي TM في E12 والتحليل في P21. لاحظ كل من الخلايا الخضراء والأحمر المسمى هي نوع البرية. (F) التخطيطي يشير إلى اثنين من المحتمل نتائج استنساخ MADM التجريبية. (زاي)القياس الكمي لحجم أغلبية السكان الناشئة عن تجديد RGPs في مُستنسخات MADM-11. (حاء)القياس الكمي لحجم الأقليات السكانية الناشئة عن تجديد RGPs في مُستنسخات MADM-11. (I) القياس الكمي للحجم الوحدوي من مستنسخات مادومي -11 العصبية غير المتماثلة. يمكن أن تمثل القيم الافتراضية متوسط ± SEM. شريط مقياس = 100 ميكرومتر (D و E). TM = تاموكسيفين. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 6: تحليل MADM التخفي لدراسة الجينات التي تؤدي إلى صغر الرأس وصغر الرأس. افتراضية نتائج تحليل مادوم النحلية عند إجراء تشريح وراثي وظيفي للجينات المرشحة التي تؤدي إلى صغر الرأس أو macrocephaly. لتشريح وظائف الخلية المستقلة من الجينات ذات الاهتمام (جين X) على انتاج الخلايا العصبية ، MADM يتطلب أليل متحولة ليتم إدخالها إلى distals MADM عبر إعادة الدمج meiotic (للحصول على تفاصيل كيفية إدخال أليل متحولة في نظام MADM انظر أيضا الشكل 2، Hippenmeyer وآخرون46، وLaukoter وآخرون46،73). (أ، ب) التخطيطي يشير إلى نموذج MADM التجريبي للتحليل الوظيفي لوحدات RGP التخفي. يمكن أن تشكل فرعية متحولة إما الأقلية (A) أو الأغلبية (ب) السكان. (C-E) افتراضية نتائج تحليل ناسق MADM عند تحديد كمي السيطرة MADM (أشرطة بيضاء), جين-X MADM microcephaly (الحانات الرمادية) وجين-X MADM الحانات السوداء macrocephaly) استنساخ غير المتماثلة. (ج)القياس الكمي لحجم أغلبية السكان. (د)القياس الكمي لحجم الأقليات السكانية. (E) القياس الكمي للحجم الوحدوي من استنساخ العصبية غير المتماثلة. يمكن أن تمثل القيم الافتراضية يعني ± SEM. S = سيناريو افتراضي حيث يمكن أن يصل الاختلاف في رقم الخلية subclone إلى أهمية، نسبة إلى عنصر التحكم. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
الجدول 1: دراسات MADM التخفي في الأدبيات. ملخص الدراسات في الأدب التي تحتوي على تجارب نسب باطني MADM، بما في ذلك برنامج التشغيل CreERT2 المستخدم، جرعة TM، ووقت الحقن. الرجاء النقر هنا لعرض هذا الجدول (انقر بزر الماوس الأيمن للتحميل).
Discussion
يتم وصف طريقة لاستخدام MADM لتتبع نسب الخلايا من RGPs الفردية في الجسم الحي في القشرة الجديدة النامية. عند دمجها مع TM-غير قابل للتكرير T2، يمكن توقيت أحداث MADM بدقة ، مما يوفر قراءات بصرية نوعية وكمية للغاية لأنماط انقسام الخلايا الجذعية على مستوى الخلية الواحدة. عن طريق المعايرة جرعة من TM تسليمها، في حالة مثالية يمكن الحصول على في المتوسط من استنساخ واحد أقل من واحد في نصف الكرة القشرية، وتوفير فصل المكانية كافية للتمييز بشكل لا لبس فيه استنساخ الفردية. من خلال الحفاظ على سلامة الأنسجة، وهذا الأسلوب يلتقط أيضا المعلومات الأساسية بشأن الموقف، مورفولوجيا، وأرقام الخلايا المطلقة. MADM كاسيت على Chr. 117,11,12,46,56,57, على Chr.7 51, وMM الأصلي في Rosa2647,53,وقد استخدمت59 في دراسات تحليل clonal MADM. عالية الدقة من الخلايا الفردية يوفر نظرة غير مسبوقة في كل من مورفولوجيا والعلاقة التخفي من الخلايا ابنة ويسمح التصوير الحي للخلايا الجذعية المتكاثرة والمستنسخات الناشئة46,52.
العملية القيصرية وتعزيز الجراء لتحليل المستنسخات في النقاط الزمنية بعد الولادة هي خطوة ضرورية وحاسمة في البروتوكول. اعتمادا على الحالة الصحية للسد الحوامل المعالجة TM، قد لا يكون من الضروري إجراء عملية قيصرية. ومع ذلك ، لا يزال من الضروري تربية الجراء مع أم بالتبني ، لأن الأم المعالجة TM قد تواجه مشكلة في المرضعات. لم تلاحظ أي اختلافات في الحاجة إلى تعزيز مع مختلف السائقين CreERT2. يتم الاحتفاظ بخطوط MADM والأمهات الحاضنات على خلفية CD-1 ذاتية. إذا لم تكن العملية القيصرية ضرورية، يمكن إعادة استخدام السد الحامل المعالج TM المستخدم لتوليد الجراء التجريبية للتكاثر التجريبي الإضافي وفقًا لمبادئ 3R (لاحظ أنه لا يمكن القيام بذلك إلا إذا وافقت تراخيص التجارب على الحيوانات على هذه الممارسة). ويمكن استخدام الأمهات الحاضنات لرعاية الجراء في غضون يومين بعد الولادة، ولكن لوحظت معدلات نجاح أعلى عندما تلد الأمهات الحاضنات في نفس اليوم الذي تلد فيه الفئران التجريبية التي يجب تعزيزها. ولذلك، من المهم إعداد التزاوج في الوقت المناسب للأمهات الحاضنات بالتوازي مع التزاوج التجريبي في الخطوة 1-1. ويمكن للحفاظ على عدد مماثل من القمامة مثل القمامة الأم الحاضنة الأصلية تحسين معدل البقاء على قيد الحياة من الجراء برعاية، وبالتالي إزالة بعض إلى كل من القمامة الأصلية قد يكون من الضروري. وتشمل الخطوات الإضافية التي قد تحسن من تعزيز فرك قفازات المجرب مع القمامة والغذاء (لإزالة رائحة القفازات)؛ فرك الجراء بلطف بعد العملية القيصرية مع شظايا من القمامة الأم الحاضنة متسخة وعش; ووضع الجراء على اتصال وثيق مع الجراء الأم بالتبني قبل وضعها في قفص الماوس بالتبني.
كما هو الحال في أساليب تتبع النسب الأخرى المستندة إلى المراسلين ، يجب أخذ الاعتبار الدقيق عند اختيار برنامج تشغيل CreERT2 الأمثل للتجارب المنعزلة MADM. أولاً، يجب على المروج المستخدم التعبير عن إعادة التركيب الزمني والمكاني في السكان السلف الذين ينهمون. العثور على هذا المروج يمكن أن يكون تحديا، لأن بعض المروجين قد تغير أنماط التعبير أو تصبح صامتة في مراحل مختلفة من التنمية. لتحسين نوع الخلية خصوصية متعددة موقع محدد recombinases ، كل مدفوعة من قبل المروجين منفصلة ، وقد استخدمت. عندما يتم التعبير عن واحد أو كليهما في نفس الخلية، وهذا التسمية الخلية وذريتها مع مراسل الفلورسنت74،75،76،77. باختصار، من المهم اختيار برنامج تشغيل CreER T2 الذي هو محدد إلى عدد السلف التي يتم تحليلها.
الخطوة الأكثر أهمية في هذا الأسلوب هو تعريف استنساخ لأنه يجب أن تكون كافة الخلايا مشتقة بشكل لا لبس فيه من حدث إعادة تجميع واحد (الخطوة 8.1). المعايرة من تركيز TM يضمن أقل من مجموعة واحدة من الخلايا الحمراء / الخضراء في نصف الكرة الدماغي ويزيد من احتمال تحليل استنساخ واحد (الخطوة 2.2)7،11. وينبغي التخلص من المستنسخات في حالة حدوث مجموعات مجاورة من الخلايا داخل 500 ميكرومتر من استنساخ الفائدة. لذلك، من المهم فحص عدة مقاطع قبل وبعد ظهور استنساخ للتأكد من أن هناك أية أحداث إعادة اشوب إضافية في مكان قريب. بسبب ضعف إشارة الفلوروفوروريس، من الضروري إجراء الكيمياء المناعية لـ eGFP وtdT في المستنسخات الجنينية (انظر القسم 6). ويوصى بذلك فقط في المستنسخات البالغة إذا كانت المستضدات الإضافية ستكون كولايغن. عند استنساخ التصوير ، من المهم التقاط العرض الكامل للقشرة حيث يوجد الاستنساخ (أي ، من سطح بيجال إلى الكلوسوم ؛ انظر الخطوة 8.4) لعدم تفويت أي خلايا. وهذا أيضا يسهل محاذاة الصورة أثناء معالجة الصور (القسم 9). يتطلب القسم 8 من البروتوكول مجهرًا مقلوبًا ولكن يمكن تكييفه اعتمادًا على إعداد المجهر المتاح. يمكن استخدام المجهر Epifluorescence ، ولكن ينصح بالمجهر confocal لأن هذا يؤدي إلى انخفاض في التلوث الضوئي من خارج الطائرة التركيز. من المهم أيضا أن يتم تعديل كثافة الليزر ومكسب بحيث يمكن تحديد الخلايا الخضراء والأحمر والأصفر بشكل لا لبس فيه. بغض النظر عن الإعداد، فمن المستحسن استخدام هدف لا يقل عن 20x لضمان الفصل المكاني الكامل للخلايا في وضع وثيق. بالإضافة إلى تسجيل العمق القشري لجميع الخلايا (الخطوة 8.6)، يجب تحديد المناطق القشرية التي توجد فيها المستنسخات باستخدام أطلس الدماغ مثل أطلس العقل ألين أو خرائط تنسيقية مجسمة أخرى. وينبغي أيضا أن يتم اعتماد نموذج تسمية الملف للتأكد من صور استنساخ يمكن التعرف عليها بسهولة. ويمكن تضمين المعلومات التالية في تسمية الملف: معرف صورة فريدة من نوعها، تم التقاط صورة التاريخ، النمط الجيني للحيوان، عمر الحث، عمر التحليل، رقم الصورة بالنسبة لبقية الصور من نفس استنساخ.
إدخال من الطفرة لكسر واحد MADM يسمح بشكل مميز لتوليد الفسيفساء الوراثية71 ويسمح تشريح المنظمين الجزيئية من النسب وتنوع نوع الخلية في المستوى التخفي7,11,46,62. لتوليد فسيفساء وراثية مع MADM، يجب أن تكون مرتبطة meiotically الكاسيت إلى نفس الكروموسوم الجين من الفائدة (انظر الشكل 2 لنظام التربية). وهذا يحد من التحليل الدونالي الحالي مع MADM إلى الجينات الموجودة على Chr.7 51, Chr. 1146, Chr. 1251, وChr. 6 distals إلى 47 12. 47 وسوف تستخدم الدراسات المستقبلية أشرطة MADM التي تستهدف أي كروموسوم، مما يسمح بتحليل الفسيفساء لجميع جينات جينوم الماوس تقريبا على المستوى السفلي.
وأخيرا، لا يقتصر MADM على تحليل الخلايا السلف في القشرة الجديدة النامية. ويمكن أن تستفيد دراسة العديد من محاريب الخلايا الجذعية من القدرة على حل الترتيبات الزمانية للزشاء للخلايا ذات الصلة بالكلون. من خلال تطبيق MADM على مناطق أخرى من الدماغ، حالات المرض (على سبيل المثال، السرطان)، أو في الأنسجة الأخرى47،,50،,51،,52،,53،,,55،,56،,57،,58،59، كشفت الدراسات علاقات النسب في استنساخ مشتقة من فئات متنوعة من السلف والخلايا الجذعية (انظر الجدول 1 للحصول على القائمة الحالية للدراسات المُزخرِجة).55 آخر تطبيق مهم في المستقبل من MADM هو الجمع بين ذلك مع المراسلين وظيفية أو دون الخلوية إضافية، والتي من شأنها أن تزيد من درجة المعلومات التي يمكن الحصول عليها من المستنسخين.
Disclosures
ليس لدى أصحاب البلاغ ما يكشفون عنه.
Acknowledgements
نشكر جميع أعضاء مختبر Hippenmeyer على المناقشة ، ومرفق التصوير البيولوجي ، ومرفق علوم الحياة ، ومرفق ما قبل السريرية في IST Austria للحصول على الدعم التقني. وقد دعم هذا العمل الصناديق المؤسسية IST Austria؛ تلقى برنامج Lise-Meitner (M 2416) دعما من صندوق العلوم النمساوي (FWF). تلقت N.A الدعم من صندوق العلوم النمساوي (FWF) Firnberg-Programm (T 1031)؛ تلقت GC الدعم من برنامج الاتحاد الأوروبي للبحوث والابتكار في أفق 2020 بموجب اتفاقية منحة ماري سكودودوسكا - كوري رقم 754411 كزميلة ما بعد الدكتوراه في ISTplus؛ تلقى أ. هـ. الدعم من زمالة الدكتوراه في الأكاديمية النمساوية للعلوم. وقد تم دعم هذه الدراسة أيضا من قبل مجلس البحوث الأوروبي (ERC) في إطار برنامج الاتحاد الأوروبي للبحث والابتكار أفق 2020 (اتفاقية منحة رقم 725780 LinPro) إلى S.H.
Materials
| Name | Company | Catalog Number | Comments |
| 1 mL tuberculin syringe (Omnifix Luer Lock) | Braun | 9204512N | |
| 1,4-diazabicyclooctane (DABCO) | Roth | 0718.2 | |
| 10 mL Syringe (Omnifix Luer Lock) | Braun | 8508429N | |
| 15 mL conical centrifuge | Sarstedt | 65.554.502 | |
| 24 multi-well dishes | Roth/Greiner Bio-one | CE56.1 | |
| 27- gauge x 3/4 needle (Sterican) | Braun | 16010256E | |
| Corn oil | Sigma | C8267-500ML | |
| Coverslips (24 x 60 mm #1) | Thermo Fisher Scientific (Menzel) | 15747592 | |
| Cryostat Cryostar NX70 | Thermo Fisher Scientific | 957000H | |
| Dako Pen (Wax marker) | Agilent | S200230-2 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Invitrogen | D1306 | |
| Disposable microtome blade (MX35 Ultra) | Thermo Fisher Scientific | 705830 | |
| Fine Forceps (Dumont #5) | Fine Science Tools (FST) | 11254-20 | |
| Glass anti-roll plate | Histocom | M 449980 | |
| Glycerol | Sigma | G5516 | |
| LSM 800 Confocal | Zeiss | ||
| Mounting medium | 25 mg/mL DAPCO, 6 g Glycerol, 2.4 g Mowiol 4-88, 6 mL dH2O, 12 mL 0.2 M Tris-HCl (pH 8.5) | ||
| Mowiol 4-88 | Roth | 0713.2 | |
| Normal donkey serum | Innovative Research | IGDNSER100ML | |
| Paraformaldehyde | Sigma | 441244-1KG | |
| Peristaltic pump 323E/D 400RPM | Watson-Marlow | 036.3124.00A | |
| Sucrose | Sigma | S8501-5KG | |
| Superfrost plus glass slides | Thermo Fisher Scientific | J1800AMNT | |
| Tamoxifen | Sigma | T5648 | |
| Tissue Embedding mold T-12 (22mm square) | Polysciences Inc. | 18986-1 | |
| Tissue-Tek O.C.T | Sakura | 4583 | |
| Triton X-100 | Sigma | T8787-250ML | |
| Trizma hydrochloride | Sigma | 93363 | |
| Tween-20 | Sigma | P9416-100ML | |
| Software and Plugins: | |||
| Fiji | 1.52p | Fiji | |
| MultiStackReg | 1.45 | Download link | |
| TurboReg | EPFL Bioimaging | ||
| Zen Blue | 2.6 | Zeiss | |
| Experimental Models: Organisms/Strains: | |||
| Mouse: Emx1-CreER | The Jackson Laboratory | JAX:027784 | |
| Mouse: MADM-11-GT | The Jackson Laboratory | JAX:013749 | |
| Mouse: MADM-11-TG | The Jackson Laboratory | JAX:013751 | |
| Primary antibodies: | |||
| Chicken anti-GFP 1:500 | Aves Labs | GFP-1020 | |
| Goat anti-tdTomato 1:500 | Sicgen Antibodies | AB8181-200 | |
| Rabbit anti-RFP 1:500 | MBL | PM005 | |
| Secondary antibodies: | |||
| Donkey Anti-Chicken Alexa Fluor 488 1:500 | Jackson Immuno Research | 715-475-150 | |
| Donkey Anti-Goat Cy3 1:500 | Jackson Immuno Research | 705-165-147 | |
| Donkey Anti-Rabbit Cy3 1:500 | Jackson Immuno Research | 711-165-152 |
References
- Malatesta, P., et al. Neuronal or glial progeny: regional differences in radial glia fate. Neuron. 37 (5), 751-764 (2003).
- Miyata, T., Kawaguchi, A., Okano, H., Ogawa, M. Asymmetric inheritance of radial glial fibers by cortical neurons. Neuron. 31 (5), 727-741 (2001).
- Noctor, S. C., Flint, A. C., Weissman, T. A., Dammerman, R. S., Kriegstein, A. R. Neurons derived from radial glial cells establish radial units in neocortex. Nature. 409 (6821), 714-720 (2001).
- Taverna, E., Götz, M., Huttner, W. B. The Cell Biology of Neurogenesis: Toward an Understanding of the Development and Evolution of the Neocortex. Annual Review of Cell and Developmental Biology. 30 (1), 465-502 (2014).
- Desikan, R. S., Barkovich, A. J. Malformations of cortical development. Annals of Neurology. 80 (6), 797-810 (2016).
- Gao, R., Penzes, P. Common mechanisms of excitatory and inhibitory imbalance in schizophrenia and autism spectrum disorders. Current Molecular Medicine. 15 (2), 146-167 (2015).
- Beattie, R., et al. Mosaic Analysis with Double Markers Reveals Distinct Sequential Functions of Lgl1 in Neural Stem Cells. Neuron. 94 (3), 517-533 (2017).
- Hippenmeyer, S. Molecular pathways controlling the sequential steps of cortical projection neuron migration. Advances in Experimental Medicine and Biology. 800, 1-24 (2014).
- Lodato, S., Arlotta, P. Generating neuronal diversity in the mammalian cerebral cortex. Annual Review of Cell and Developmental Biology. 31, 699-720 (2015).
- Hansen, A. H., Duellberg, C., Mieck, C., Loose, M., Hippenmeyer, S. Cell Polarity in Cerebral Cortex Development-Cellular Architecture Shaped by Biochemical Networks. Frontiers in Cellular Neuroscience. 11, 176 (2017).
- Gao, P., et al. Deterministic progenitor behavior and unitary production of neurons in the neocortex. Cell. 159 (4), 775-788 (2014).
- Llorca, A., et al. A stochastic framework of neurogenesis underlies the assembly of neocortical cytoarchitecture. eLife. 8, e51381 (2019).
- Ma, J., Shen, Z., Yu, Y. C., Shi, S. H. Neural lineage tracing in the mammalian brain. Current Opinion in Neurobiology. 50, 7-16 (2018).
- Caviness, V., Takahashi, T., Nowakowski, R. Numbers, time and neocortical neuronogenesis: a general developmental and evolutionary model. Trends in Neurosciences. 18 (9), 379-383 (1995).
- Schmechel, D. E., Rakic, P. A Golgi study of radial glial cells in developing monkey telencephalon: morphogenesis and transformation into astrocytes. Anatomy and Embryology. 156 (2), 115-152 (1979).
- Kessaris, N., et al. Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nature Neuroscience. 9 (2), 173-179 (2006).
- Magavi, S., Friedmann, D., Banks, G., Stolfi, A., Lois, C. Coincident generation of pyramidal neurons and protoplasmic astrocytes in neocortical columns. The Journal of Neuroscience. 32 (14), 4762-4772 (2012).
- Anthony, T. E., Klein, C., Fishell, G., Heintz, N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 41 (6), 881-890 (2004).
- Voigt, T. Development of glial cells in the cerebral wall of ferrets: direct tracing of their transformation from radial glia into astrocytes. The Journal of Comparative Neurology. 289 (1), 74-88 (1989).
- Amberg, N., Laukoter, S., Hippenmeyer, S. Epigenetic cues modulating the generation of cell-type diversity in the cerebral cortex. Journal of Neurochemistry. 149 (1), 12-26 (2019).
- Beattie, R., Hippenmeyer, S. Mechanisms of Radial Glia Progenitor Cell Lineage Progression. FEBS letters. 591 (24), 3993-4008 (2017).
- Telley, L., et al. Temporal patterning of apical progenitors and their daughter neurons in the developing neocortex. Science. 364 (6440), eaav2522 (2019).
- Oberst, P., et al. Temporal plasticity of apical progenitors in the developing mouse neocortex. Nature. 573 (7774), 370-374 (2019).
- Telley, L., et al. Sequential transcriptional waves direct the differentiation of newborn neurons in the mouse neocortex. Science. 351 (6280), 1443 (2016).
- Deppe, U., et al. Cell lineages of the embryo of the nematode Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America. 75 (1), 376-380 (1978).
- Woodworth, M. B., Girskis, K. M., Walsh, C. A. Building a lineage from single cells: genetic techniques for cell lineage tracking. Nature Reviews Genetics. 18 (4), 230-244 (2017).
- Masuyama, N., Mori, H., Yachie, N. DNA barcodes evolve for high-resolution cell lineage tracing. Current Opinion in Chemical Biology. 52, 63-71 (2019).
- Legue, E., Joyner, A. L. Chapter Ten-Genetic Fate Mapping Using Site-Specific Recombinases. Methods in Enzymology. 477, 153-181 (2010).
- Postiglione, M. P., Hippenmeyer, S. Monitoring neurogenesis in the cerebral cortex: an update. Future Neurology. 9 (3), 323-340 (2014).
- Espinosa-Medina, I., Garcia-Marques, J., Cepko, C., Lee, T. High-throughput dense reconstruction of cell lineages. Open Biology. 9 (12), 190229 (2019).
- Hwang, B., et al. Lineage tracing using a Cas9-deaminase barcoding system targeting endogenous L1 elements. Nature Communications. 10 (1), 1234 (2019).
- Kalhor, R., et al. Developmental barcoding of whole mouse via homing CRISPR. Science. 361 (6405), eaat9804 (2018).
- García-Moreno, F., Vasistha, N. A., Begbie, J., Molnár, Z. CLoNe is a new method to target single progenitors and study their progeny in mouse and chick. Development. 141 (7), 1589-1598 (2014).
- Loulier, K., et al. Multiplex Cell and Lineage Tracking with Combinatorial Labels. Neuron. 81 (3), 505-520 (2014).
- Livet, J., et al. Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature. 450 (7166), 56-62 (2007).
- Snippert, H. J., et al. Intestinal Crypt Homeostasis Results from Neutral Competition between Symmetrically Dividing Lgr5 Stem Cells. Cell. 143 (1), 134-144 (2010).
- Amitai-Lange, A., et al. A method for lineage tracing of corneal cells using multi-color fluorescent reporter mice. Journal of Visualized Experiments. (106), e53370 (2015).
- Vasistha, N. A., et al. Cortical and Clonal Contribution of Tbr2 Expressing Progenitors in the Developing Mouse Brain. Cerebral Cortex. 25 (10), 3290-3302 (2015).
- Chen, F., LoTurco, J. A method for stable transgenesis of radial glia lineage in rat neocortex by piggyBac mediated transposition. Journal of Neuroscience Methods. 207 (2), 172-180 (2012).
- Siddiqi, F., et al. Fate mapping by piggyBac transposase reveals that neocortical GLAST+ progenitors generate more astrocytes than Nestin+ progenitors in rat neocortex. Cerebral Cortex. 24 (2), 508-520 (2014).
- Yoshida, A., et al. Simultaneous expression of different transgenes in neurons and glia by combining in utero electroporation with the Tol2 transposon-mediated gene transfer system. Genes to Cells. 15 (5), 501-512 (2010).
- García-Marqués, J., López-Mascaraque, L. Clonal Identity Determines Astrocyte Cortical Heterogeneity. Cerebral Cortex. 23 (6), 1463-1472 (2013).
- Clavreul, S., et al. Cortical astrocytes develop in a plastic manner at both clonal and cellular levels. Nature Communications. 10 (1), 4884 (2019).
- Kim, G. B., et al. Rapid Generation of Somatic Mouse Mosaics with Locus-Specific, Stably Integrated Transgenic Elements. Cell. 179 (1), 251-267 (2019).
- Pilz, G. A., et al. Live imaging of neurogenesis in the adult mouse hippocampus. Science. 359 (6376), 658 (2018).
- Hippenmeyer, S., et al. Genetic Mosaic Dissection of Lis1 and Ndel1 in Neuronal Migration. Neuron. 68 (4), 695-709 (2010).
- Zong, H., Espinosa, J. S., Su, H. H., Muzumdar, M. D., Luo, L. Mosaic analysis with double markers in mice. Cell. 121 (3), 479-492 (2005).
- Ramón y Cajal, S. . Histologie du système nerveux de l'homme et des vertébrés. , (1911).
- Cowan, W. M. The emergence of modern neuroanatomy and developmental neurobiology. Neuron. 20 (3), 413-426 (1998).
- Ali, S. R., et al. Existing cardiomyocytes generate cardiomyocytes at a low rate after birth in mice. Proceedings of the National Academy of Sciences of the United States of America. 111 (24), 8850-8855 (2014).
- Hippenmeyer, S., Johnson, R. L., Luo, L. Mosaic analysis with double markers reveals cell-type-specific paternal growth dominance. Cell Reports. 3 (3), 960-967 (2013).
- Riccio, P., Cebrian, C., Zong, H., Hippenmeyer, S., Costantini, F. Ret and Etv4 promote directed movements of progenitor cells during renal branching morphogenesis. PLoS Biology. 14 (2), e1002382 (2016).
- Bonaguidi, M. A., et al. In vivo clonal analysis reveals self-renewing and multipotent adult neural stem cell characteristics. Cell. 145 (7), 1142-1155 (2011).
- Mayer, C., et al. Clonally Related Forebrain Interneurons Disperse Broadly across Both Functional Areas and Structural Boundaries. Neuron. 87 (5), 989-998 (2015).
- Muzumdar, M. D., et al. Clonal dynamics following p53 loss of heterozygosity in Kras-driven cancers. Nature Communications. 7, 12685 (2016).
- Shi, W., et al. Ontogenetic establishment of order-specific nuclear organization in the mammalian thalamus. Nature Neuroscience. 20, 516 (2017).
- Wong, S. Z. H., et al. In vivo clonal analysis reveals spatiotemporal regulation of thalamic nucleogenesis. PLoS Biology. 16 (4), e2005211 (2018).
- Xu, H. T., et al. Distinct Lineage-Dependent Structural and Functional Organization of the Hippocampus. Cell. 157 (7), 1552-1564 (2014).
- Brennand, K., Huangfu, D., Melton, D. All β Cells Contribute Equally to Islet Growth and Maintenance. PLoS Biology. 5 (7), e163 (2007).
- Ortiz-Alvarez, G., et al. Adult neural stem cells and multiciliated ependymal cells share a common lineage regulated by the geminin family members. Neuron. 102 (1), 159-172 (2019).
- Kaplan, E. S., Ramos-Laguna, K. A., Mihalas, A. B., Daza, R. A. M., Hevner, R. F. Neocortical Sox9+ radial glia generate glutamatergic neurons for all layers, but lack discernible evidence of early laminar fate restriction. Neural Development. 12 (1), 14 (2017).
- Lv, X., et al. TBR2 coordinates neurogenesis expansion and precise microcircuit organization via Protocadherin 19 in the mammalian cortex. Nature Communications. 10 (1), 3946 (2019).
- Mihalas, A. B., Hevner, R. F. Clonal analysis reveals laminar fate multipotency and daughter cell apoptosis of mouse cortical intermediate progenitors. Development. 145 (17), dev164335 (2018).
- Picco, N., et al. A mathematical insight into cell labelling experiments for clonal analysis. Journal of Anatomy. 235 (3), 687-696 (2019).
- Johnson, C. A., Ghashghaei, H. T. Sp2 regulates late neurogenic but not early expansive divisions of neural stem cells underlying population growth in the mouse cortex. Development. , (2020).
- Espinosa, J. S., Luo, L. Timing Neurogenesis and Differentiation: Insights from Quantitative Clonal Analyses of Cerebellar Granule Cells. The Journal of Neuroscience. 28 (10), 2301 (2008).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Liu, Y., et al. Tamoxifen-independent recombination in the RIP-CreER mouse. PLoS One. 5 (10), e13533 (2010).
- Klingler, E., Jabaudon, D. Do progenitors play dice?. eLife. 9, e54042 (2020).
- Hippenmeyer, S. Dissection of gene function at clonal level using mosaic analysis with double markers. Frontiers in Biology. 8 (6), 557-568 (2013).
- Rulands, S., Simons, B. D. Tracing cellular dynamics in tissue development, maintenance and disease. Current Opinion in Cell Biology. 43, 38-45 (2016).
- Laukoter, S., et al. Imprinted Cdkn1c genomic locus cell-autonomously promotes cell survival in cerebral cortex development. Nature Communications. 11 (1), 195 (2020).
- Daigle, T. L., et al. A Suite of Transgenic Driver and Reporter Mouse Lines with Enhanced Brain-Cell-Type Targeting and Functionality. Cell. 174 (2), 465-480 (2018).
- He, M., et al. Strategies and Tools for Combinatorial Targeting of GABAergic Neurons in Mouse Cerebral Cortex. Neuron. 91 (6), 1228-1243 (2016).
- Yamamoto, M., et al. A multifunctional reporter mouse line for Cre- and FLP-dependent lineage analysis. Genesis. 47 (2), 107-114 (2009).
- Plummer, N. W., et al. Expanding the power of recombinase-based labeling to uncover cellular diversity. Development. 142 (24), 4385 (2015).
- Imayoshi, I., Ohtsuka, T., Metzger, D., Chambon, P., Kageyama, R. Temporal regulation of Cre recombinase activity in neural stem cells. Genesis. 44 (5), 233-238 (2006).
- Sasaki, S., et al. Complete loss of Ndel1 results in neuronal migration defects and early embryonic lethality. Molecular and Cellular Biology. 25 (17), 7812-7827 (2005).
- Acampora, D., et al. Epilepsy and brain abnormalities in mice lacking the Otx1 gene. Nature Genetics. 14 (2), 218-222 (1996).
- Soeda, T., et al. Sox9-expressing precursors are the cellular origin of the cruciate ligament of the knee joint and the limb tendons. Genesis. 48 (11), 635-644 (2010).
- Klezovitch, O., Fernandez, T. E., Tapscott, S. J., Vasioukhin, V. Loss of cell polarity causes severe brain dysplasia in Lgl1 knockout mice. Genes & Development. 18 (5), 559-571 (2004).
- Pimeisl, I. M., et al. Generation and characterization of a tamoxifen-inducible EomesCreER mouse line. Genesis. 51 (10), 725-733 (2013).
- Nakagawa, N., et al. Memo1-Mediated Tiling of Radial Glial Cells Facilitates Cerebral Cortical Development. Neuron. 103 (5), 836-852 (2019).
- Nowotschin, S., et al. The T-box transcription factor Eomesodermin is essential for AVE induction in the mouse embryo. Genes & Development. 27 (9), 997-1002 (2013).
- Balordi, F., Fishell, G. Mosaic removal of hedgehog signaling in the adult SVZ reveals that the residual wild-type stem cells have a limited capacity for self-renewal. Journal of Neuroscience. 27 (52), 14248-14259 (2007).
- Liang, H., et al. Neural development is dependent on the function of specificity protein 2 in cell cycle progression. Development. 140 (3), 552-561 (2013).
- Guo, C., Yang, W., Lobe, C. G. A Cre recombinase transgene with mosaic, widespread tamoxifen-inducible action. Genesis. 32 (1), 8-18 (2002).
- Ahn, S., Joyner, A. L. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature. 437 (7060), 894-897 (2005).
- Storm, R., et al. The bHLH transcription factor Olig3 marks the dorsal neuroepithelium of the hindbrain and is essential for the development of brainstem nuclei. Development. 136 (2), 295-305 (2009).
- Koundakjian, E. J., Appler, J. L., Goodrich, L. V. Auditory neurons make stereotyped wiring decisions before maturation of their targets. Journal of Neuroscience. 27 (51), 14078-14088 (2007).
- Dor, Y., Brown, J., Martinez, O. I., Melton, D. A. Adult pancreatic β-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 429 (6987), 41-46 (2004).
- Sohal, D. S., et al. Temporally regulated and tissue-specific gene manipulations in the adult and embryonic heart using a tamoxifen-inducible Cre protein. Circulation Research. 89 (1), 20-25 (2001).
- Ventura, A., et al. Restoration of p53 function leads to tumour regression in vivo. Nature. 445 (7128), 661-665 (2007).
- Johnson, L., et al. Somatic activation of the K-ras oncogene causes early onset lung cancer in mice. Nature. 410 (6832), 1111-1116 (2001).
- Tasic, B., et al. Extensions of MADM (mosaic analysis with double markers) in mice. PLoS One. 7 (3), e33332 (2012).
- Yu, J., Carroll, T. J., McMahon, A. P. Sonic hedgehog regulates proliferation and differentiation of mesenchymal cells in the mouse metanephric kidney. Development. 129 (22), 5301-5312 (2002).
- Zhao, H., et al. Role of fibroblast growth factor receptors 1 and 2 in the ureteric bud. Developmental Biology. 276 (2), 403-415 (2004).
- Schuchardt, A., D'Agati, V., Larsson-Blomberg, L., Costantini, F., Pachnis, V. Defects in the kidney and enteric nervous system of mice lacking the tyrosine kinase receptor Ret. Nature. 367 (6461), 380-383 (1994).
- Livet, J., et al. ETS gene Pea3 controls the central position and terminal arborization of specific motor neuron pools. Neuron. 35 (5), 877-892 (2002).
- Volz, K. S., et al. Pericytes are progenitors for coronary artery smooth muscle. eLife. 4, e10036 (2015).
- Cai, C. L., et al. A myocardial lineage derives from Tbx18 epicardial cells. Nature. 454 (7200), 104-108 (2008).
- Lasrado, R., et al. Lineage-dependent spatial and functional organization of the mammalian enteric nervous system. Science. 356 (6339), 722-726 (2017).
- Matsuoka, T., et al. Neural crest origins of the neck and shoulder. Nature. 436 (7049), 347-355 (2005).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved