
Method Article
Tracciamento del lignaggio e analisi clonale nello sviluppo della Corteccia cerebrale utilizzando l'analisi mosaica con doppi marcatori (MADM)
In questo articolo
Riepilogo
Viene presentato un protocollo per eseguire il tracciamento del lignaggio e l'analisi genetica funzionale dei geni candidati a livello di singola cellula utilizzando l'analisi del mosaico con doppi marcatori (MADM). L'analisi clonale MADM fornisce un quadro quantitativo per misurare il comportamento proliferativo, l'output cellulare e la relazione di lignaggio dei singoli progenitori e delle loro cellule figlie.
Abstract
Partendo da un pool limitato di progenitori, la corteccia cerebrale dei mammiferi forma circuiti neurali funzionali altamente organizzati. Tuttavia, i meccanismi cellulari e molecolari sottostanti che regolano le transizioni di lignaggio delle cellule staminali neurali (NSC) e l'eventuale produzione di neuroni e glia nello sviluppo di neuroepitelio rimane poco chiaro. I metodi per tracciare i modelli di divisione NSC e mappare il lignaggio delle celle correlate clonalmente sono avanzati notevolmente. Tuttavia, molte tecniche di tracciamento del lignaggio contemporaneo soffrono della mancanza di risoluzione cellulare del destino delle cellule progenie, che è essenziale per decifrare i modelli di divisione delle cellule progenitori. Presentato è un protocollo che utilizza l'analisi a mosaico con doppi marcatori (MADM) per eseguire l'analisi clonale in vivo. MADM manipola concomitante singole cellule progenitrici e visualizza precisi modelli di divisione e progressione del lignaggio a una risoluzione cellulare singola senza precedenti. Gli eventi di ricombinazione intercromosomica basati su MADM durante la fase G2-X della mitosi, insieme a CreERT2,temporaneamente inducibili, forniscono informazioni esatte sulle date di nascita dei cloni e sui loro modelli di divisione. Pertanto, il tracciamento del lignaggio MADM fornisce letture ottiche qualitative e quantitative senza precedenti della modalità di proliferazione dei progenitori di cellule staminali a livello di singola cellula. MADM consente anche di esaminare i meccanismi e i requisiti funzionali dei geni candidati nella progressione del lignaggio NSC. Questo metodo è unico in quanto l'analisi comparativa del controllo e dei subcloni mutanti può essere eseguita nello stesso ambiente tissutale in vivo. Qui, il protocollo è descritto in dettaglio, e paradigmi sperimentali per impiegare MADM per l'analisi clonale e traccia di lignaggio nella corteccia cerebrale in via di sviluppo sono dimostrati. È importante sottolineare che questo protocollo può essere adattato per eseguire l'analisi clonale MADM in qualsiasi nicchia di cellule staminali murine, purché sia presente il driver CreERT2.
Introduzione
La corteccia cerebrale è una struttura altamente organizzata composta da sei strati distinti. La corteccia contiene una vasta gamma di tipi di cellule tra cui neuroni e glia, che interagiscono per formare circuiti neurali funzionali. La maggior parte, se non tutti, i neuroni di proiezione eccitatoria corticale e glia sono derivati da un pool comune di cellule staminali neurali (NSC) noto come i progenitori gliali radiali (RGP)1,2,3. Gli XML stessi sono derivati da cellule staminali neuroepiteliali (NESC) che compongono il neuroetag embrionale precoce. Entro il giorno embrionale 9 (E9) nei topi, i NESC iniziano a passare in RGP4. La progressione del lignaggio RGP richiede una precisa regolazione temporale e spaziale, e quando questo processo è ostacolato, gravi disturbi neurologici come megalenfalia, microcefalia, lissencephaly, o menomazioni come la schizofrenia e l'autismo possono risultare5,6. All'E10, la maggior parte degli RGP subiscono divisioni proliferanti simmetriche, con conseguente espansione del pool di progenitrici neurali4,7. Gli RGP alla fine iniziano a dividersi asimmetricamente, producendo neuroni di proiezione corticale in modo definito temporalmente. Attraverso ondate consecutive di neurogenesi, i neuroni neonati migrano nella piastra corticale formando laminari corticali con neuroni nati precoci che occupano strati profondi e neuroni nati tardi che risiedono negli strati superficiali8,9,10. Poiché i neuroni piramidali correlati clonalmente migrano radialmente nella corteccia con una piccola dispersione tangenziale, le cellule figlie tendono a formare una struttura a forma di colonna o cono, indicata come unità radiale neuronale4,11,1212,13. Entro e17, l'espansione neurogenica embrionale è completa nei topi14. Gli RGP possono anche produrre cellule ependymal e alcune classi di glia, tra cui astrociti e oligodendrociti1,15,16,17,18,19. Il potenziale degli RGP di dare origine sia ai neuroni che agli astrociti sembra essere coerente in tutte le regioni corticali18, con circa 1/6 di RGP neurogenici che producono anche glia11.
Attualmente, i fattori genetici ed epigenetici che regolano la progressione temporale di una cellula staminale lungo il suo lignaggio sono per lo più sconosciuti. I modelli temporali di espressione genica possono avere un impatto sostanziale sulle decisioni di lignaggio nei RGP20,21,22,23,24. Non è noto come questa relazione strettamente lavorata tra modelli temporali e spaziali porti alla diversità molecolare dei tipi neuronali adulti nelle aree corticali. Allo stesso modo, il modo in cui il potenziale delle singole cellule staminali e la sua produzione cellulare sono modulati a livello cellulare e molecolare è un'importante domanda senza risposta. Si spera che studi futuri affrontino alcune di queste domande, ampliando in ultima analisi la nostra comprensione della formazione funzionale dei circuiti corticali.
La neurobiologia dello sviluppo cerca di capire il rapporto di lignaggio che le cellule nel cervello condividono tra loro. Inizialmente, pochissimi strumenti di ricerca erano disponibili per questo, e molti primi studi si basavano su osservazioni visive di modelli di divisione in organismi trasparenti come Caenorhabditis elegans25. Gli ultimi decenni hanno visto un drammatico aumento del numero e della raffinatezza delle tecniche disponibili13,26,27,28,29. L'emergere del sistema di editing genomico CRISPR-Cas9 consente la ricostruzione sintetica delle relazioni di lignaggio cellulare introducendo codici a barre in evoluzione del DNA27,30. Due esempi recenti di strategie di codifica a barre includono l'uso di RNA guida homing che dirige CRISPR-Cas9 a specifici loci codice a barre DNA o un deaminasi civogliao fuso con nickase Cas9 per indirizzare regioni di ripetizione interspersed endogene31,32. Queste tecnologie forniscono approcci altamente multiplexed attraverso l'introduzione di codici a barre che accumulano progressivamente e stabilmente mutazioni uniche nel tempo. Gli approcci di editing genomico sono molto preziosi perché consentono un'analisi retroattiva della relazione tra due celle qualsiasi in base all'ereditarietà condivisa di questi codici a barre. Tuttavia, al fine di leggere i codici a barre in singole cellule, il tessuto di solito deve essere interrotto, e quindi le informazioni riguardanti la posizione, morfologia, e numeri di cella assoluti da un singolo progenitore viene perso.
I paradigmi di etichettatura combinatori contengono informazioni spaziali e, in linea di principio, consentono anche la distinzione tra cloni strettamente localizzati o addirittura sovrapposti33,34. Affinché un metodo di tracciamento del lignaggio sia informativo, deve etichettare i singoli progenitori e la loro progenie in modo scarso e indelebile. In particolare, il Brainbow35 e Confetti36,37 approcci utilizzano stocaststici multicolore Cre ricombinante a base di reporter che esprimono una combinazione di proteine fluorescenti da un unico locus. L'ampio numero di combinazioni di colori simultanei che possono essere raggiunte in vivo lo rendono uno strumento potente per tracciare cloni corticali RGP e astrociti34. Sono stati sviluppati anche sistemi basati su trasposoni che forniscono un'integrazione genomica stabile dei transgeni che codificano reporter fluorescenti e permettendo traccia lignaggio dei progenitori corticali sono stati sviluppati33,38,39,40,41. I sistemi basati su trasposoni hanno un ulteriore vantaggio in quanto il reporter costruisce un'integrazione stabilmente nel genoma, e quindi etichettare in modo affidabile cellule figlie correlate. Per tracciare specificamente le linee di astrociti, sono stati sviluppati una serie di metodi che coinvolgono l'elettroporazione delle trasposta piggyBac tra cui Star Track, che fa uso di una combinazione di costrutti che codificano diverse proteine fluorescenti40,42. Un altro approccio, MAGIC markers, introduce i vettori Brainbow come transgeni trasponibili. Questo è stato utilizzato con successo per tracciare progenitori neurali e astrociti embrionali34,43. Recentemente, è stata trovata un'analisi a mosaico da parte di un doppio scambio di cassette mediato ricombinante (MADR) per etichettare stabilmente le cellule mutanti che esprimono elementi transgenici da loci cromosomici precisamente definiti44. Queste potenti tecniche di etichettatura combinatoria in vivo hanno fornito numerose informazioni sulla dinamica del lignaggio delle cellule progenitrici. Tuttavia, queste analisi vengono eseguite su tessuto fisso, fornendo un'istantanea di singoli cloni in una fase di sviluppo definita. Per osservare i cambiamenti nella dinamica del lignaggio dei singoli cloni nel tempo, è necessario applicare metodi di imaging in vivo cronici simili a quelli eseguiti nel giro dentato adulto45.
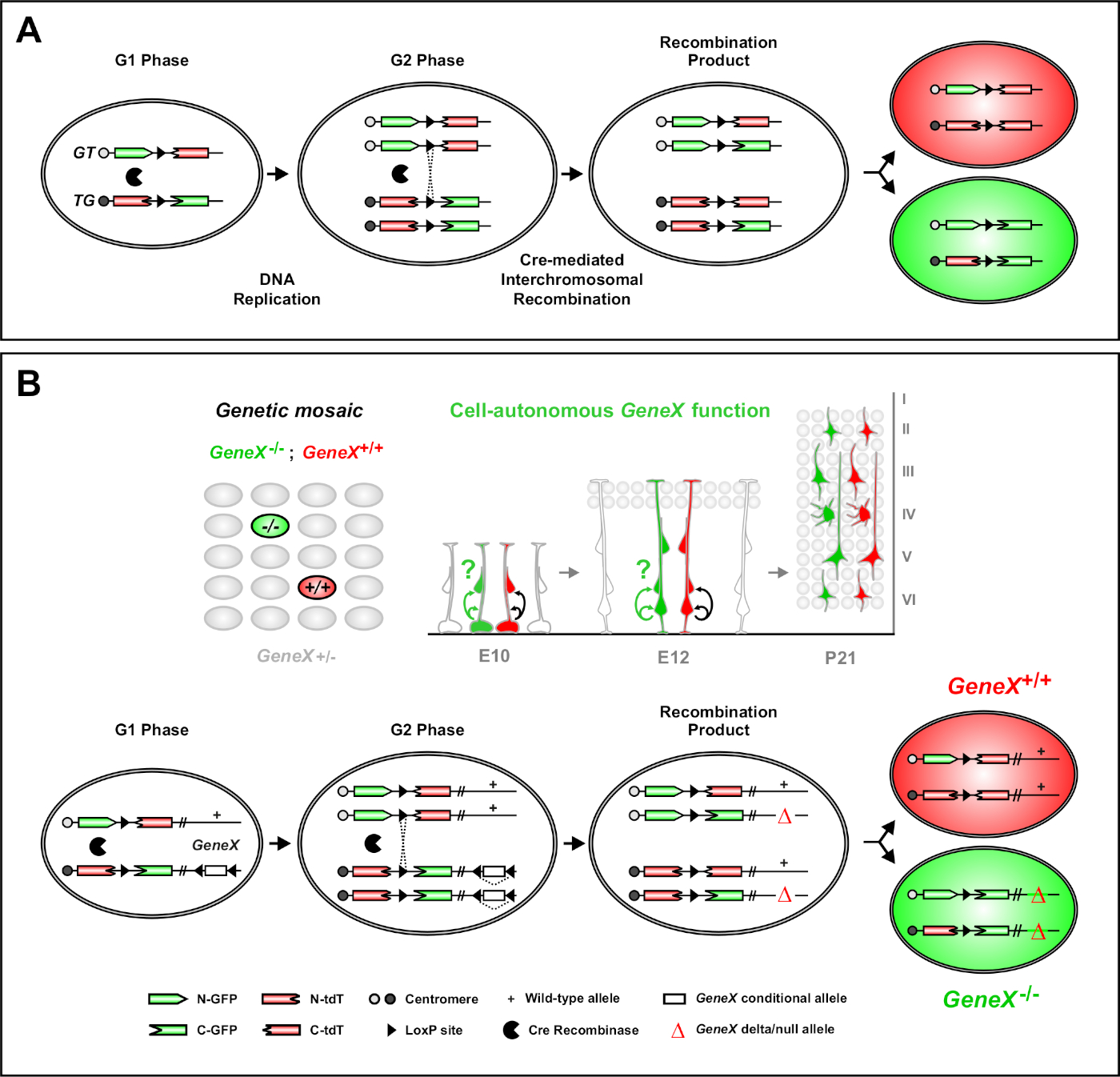
L'analisi mosaica con doppio marcatore (MADM) è un potente metodo di etichettatura a doppio colore che consente la tracciatura del lignaggio in vivo delle singole cellule progenitrici nei topi46,47. Due componenti sono necessari per gli eventi di etichettatura MADM: in primo luogo, le cassette MADM devono essere mirate a loci identici sui cromosomi omologhi. Le cassette sono costituite da due geni di reporter fluorescenti chimerici, eGFP (verde, [G]) e dimero tandem Pomodoro (rosso, tdT[T]). La cassetta GT contiene il capoN di eGFP e il capo-C del tdT, separati da un intron contenente un sito loxP. La cassetta TG è costruita inversamente, con il capoN del tdT e il capo-C di eGFP. In secondo luogo, l'espressione di Cre ricombinarsi nella stessa cella contenente le cassette MADM mirate è essenziale. In assenza di Cre, le cassette chimesiche non esprimono eGFP funzionale o tdT perché le loro sequenze di codifica sono interrotte. I siti loxP fungono da bersaglio per la ricombinazione intercromosomica mediata da Cre, con conseguente ricostituzione simultanea di entrambe le cassette di espressione. Se la ricombinazione avviene durante la fase G2 del ciclo cellulare seguita dalla segregazione X (G2-X), le due cellule figlie esprimeranno ciascuna una delle due proteine fluorescenti. La regolazione temporale dell'attività CreERT2 mediante tamoxifene (TM) fornisce informazioni precise sulla data di nascita dei cloni MADM e sui modelli di divisione della loro progenie (Figura 1A)29,46,47.
MADM può potenzialmente etichettare sistematicamente singoli cloni con alta risoluzione cellulare singola nel cervello del topo simile ai metodi tradizionali ma non specifici e laboriosi come golgi colorazione48 o colorante riempimento49. Poiché solo il promotore di guida CreERT2 determina la specificità del tipo di cella dell'etichettatura CLONale MADM, MADM può in linea di principio essere applicato per la tracciatura del lignaggio clonale in qualsiasi organo murino e tessuto47,50,51,52. Infatti, gli studi hanno già utilizzato MADM per rivelare le relazioni di lignaggio in cloni derivati da tessuti diversi47,50,51,52,53,54,55,56,57,58,59. I paradigmi sperimentali MADM sono stati applicati per studiare il lignaggio nei neuroni di proiezione corticale, glia e cellule staminali postnatali nella neocortecciainvia di sviluppo 7,11,12,46,60,61,62,63,64,65. MADM è stato utilizzato anche per studiare il lignaggio cellulare nel giro dentato adulto, talamo, cellule granule cerebellari, e interneuroni a livello clonale (vedi tabella 1 per un elenco completo)47,53,54,56,5757,66.
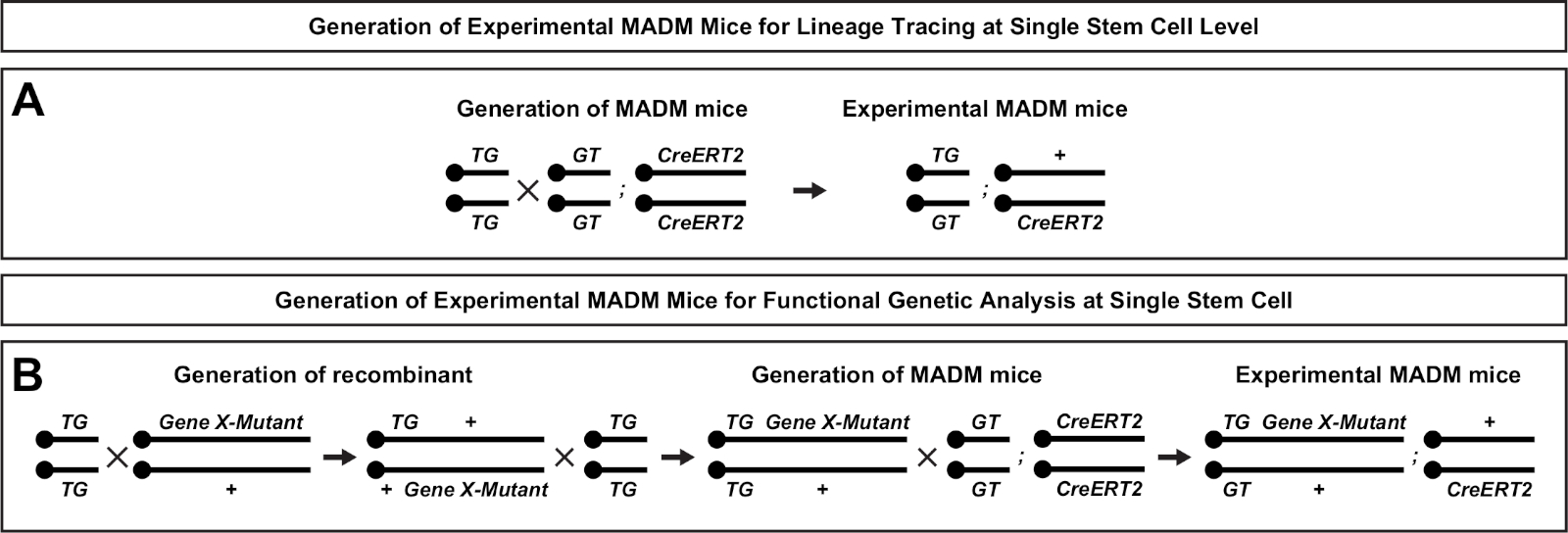
Una caratteristica unica di MADM è la capacità di collegare geneticamente le mutazioni distale a una cassetta MADM, creando così un mosaico genetico (Figura 1B e Figura 2). Questo si traduce in cellule figlie di tipo selvaggio etichettate con un marcatore fluorescente (tdT nella Figura 1B) e fratelli mutanti omoygous con l'altro (eGFP in Figura 1B) in un ambiente eterozigoso senza etichetta. IL MADM è unico in quanto l'analisi comparativa del controllo e dei subcloni mutanti può essere eseguita nello stesso ambiente tissutale in vivo. Originariamente, le cassette MADM erano mirate al locus Rosa26 47, ma l'analisi MADM della funzione genica era limitata ai geni disslocanti al locus. Per superare (almeno in parte) questa limitazione ed espandere le possibilità di analisi genica basata su MADM, le cassette MADM sono state bussate vicino ai centrimeri di Chr. 751, Chr. 1146e Chr. 1251. È in corso di mira tutti i 19 autosomes per topi con cassette MADM che consentirà di studiare praticamente qualsiasi gene in futuro, fornendo una piattaforma senza precedenti per lo studio delle relazioni di lignaggio dello sviluppo in combinazione con l'analisi genetica funzionale.
Protocollo
I protocolli del mouse sono stati esaminati dalla struttura centrale preclinica istituzionale (PCF) e dal comitato etico interno dell'IST Austria. Tutti gli allevamento e le sperimentazioni sono stati effettuati in base a una licenza approvata dal Ministero federale austriaco della scienza e della ricerca in conformità con le leggi austriache e dell'UE sugli animali.
1. Allevamento di topi sperimentali per l'analisi clonale MADM
- Impostare accoppiamenti MADM sperimentali a tempo (>P56; CD-1) nel tardo pomeriggio (5:00 PM) e verificare la presenza di tappi vaginali la mattina procedinte (8:00 AM). La mattina in cui la spina è presente conta come giorno 0.5. Vedere Figura 2 per una panoramica della configurazione di accoppiamento del mouse sperimentale. Assicurarsi che i tempi per l'introduzione di attività e analisi di CreERT2 siano appropriati per affrontare questioni sperimentali.
NOTA: Per ulteriori informazioni, fare riferimento alla figura 3 e ai risultati rappresentativi riportati di seguito. - Per il campionamento postnatale, impostare allevamenti per generare madri affidatarie in parallelo.
NOTA: Questi dovrebbero essere avviati fino a 1/2 giorni prima di impostare allevamenti sperimentali.
2. Induzione TM nei topi MADM
- Preparare una soluzione di lavoro da 20 mg/mL TM sciogliendola nell'olio di mais in un tubo di centrifuga conica da 15 mL o 50 mL e posizionandola su una piattaforma a dondolo per 4 h a temperatura ambiente (RT), assicurandoche che la TM sia completamente dissolta. Conservare la soluzione di lavoro a 4 gradi centigradi ricoperta di lamina di alluminio e utilizzarla entro 2 settimane.
- Per indurre eventi di ricombinazione MADM, effettuare una singola iniezione di TM intraperitonealmente (IP) utilizzando una siringa di tubercolina da 1 mL e un ago da 25 G in una diga incinta a tempo. A seconda dello stadio della neurogenesi corticale, iniettare la TM tra E10 e 15 ad una dose di diga da 1 mg/gravidanza. Per i tempi precoci (cioè E10) utilizzare un massimo di 1 mg/diga incinta (25 mg/kg) per prevenire complicazioni durante la gravidanza11. Per i tempi tra E11-E15 utilizzare 2 mg/diga incinta (50 mg/kg)7.
NOTA: In alternativa, TM può essere somministrato con un gavage orale per le gravidanze tardive. - Per l'analisi clonale MADM ai tempi postnatali, recupera gli embrioni vivi all'E18-E19 attraverso la sezione cesareo, e poi alleva i cuccioli con una madre affidataria.
NOTA: A seconda dello stato di salute della femmina incinta, potrebbe non essere necessario eseguire una sezione cesareo, ma allevare cuccioli con una madre adottiva è ancora necessario perché la madre originale trattata in TM potrebbe avere problemi ad allattarsi. - Per recuperare gli embrioni vivi per sezione cesareo o per recuperare i tempi embrionali per l'analisi, sacrificare la diga incinta per lussazione cervicale.
- Mettere l'animale in posizione supina e disinfettare la pelliccia con il 70% di etanolo. Fare una piccola incisione nella pelle nell'addome inferiore sopra l'utero utilizzando pinze chirurgiche e forbici. Fare una seconda incisione attraverso i muscoli e la parete muscolare addominale per rivelare il peritoneo.
- Rimuovere l'utero separando i tessuti circostanti con le forbici. Trasferire l'utero intatto su un guanto riempito con acqua tiepida per aumentare il tasso di sopravvivenza dell'embrione fino a quando ogni viene rimosso singolarmente dall'amnione.
- Utilizzare forbici e dita con punta fine per aprire con cura le pareti uterine per rilasciare gli embrioni. Non tagliare i cordoni ombelicali troppo vicini al corpo per prevenire la perdita di sangue estensiva. Se gli embrioni devono essere utilizzati per l'analisi, procedere al passaggio 3.9. Se i cuccioli devono essere affidati, procedere al passaggio 2.8.
- Se è necessario l'affidamento, pulire i cuccioli prima di trasferirli alla madre affidataria. Durante la pulizia dei cuccioli, premere delicatamente il torace di tanto in tanto per iniziare la respirazione. Rimettere su un secondo guanto riempito con acqua tiepida per migliorare il tasso di sopravvivenza.
NOTA: È importante rimuovere delicatamente l'amnione rimanente e/o la placenta con un tovagliolo di carta. - Prima di trasferire i cuccioli alla madre affidataria, rimuovi la madre affidataria dalla sua gabbia, rimuovi i cuccioli originali e sostituiscili con i cuccioli sperimentali. Rirestituisci la madre affidataria alla sua gabbia.
NOTA: vedere la discussione per ulteriori suggerimenti per migliorare i tassi di accettazione della promozione. - Se è necessaria la genotipizzazione, raccogliere le biopsie di p6-P8.
NOTA: eseguire questo passaggio solo se le licenze sperimentali per animali approvano questa procedura.
3. Preparazione dei tessuti per cloni MADM nel cervello
NOTA: per gli esperimenti che includono il tessuto postnatale (P4), procedere al passaggio 3.1. Per i tempi embrionali e i primi postnatali (P0p3), continuare con il passaggio 3.9.
- Anestesizzare l'animale sperimentale MADM con un'iniezione IP di una soluzione di ketamina/xylazina/acepromazina (65 mg, 13 mg e 2 mg/kg di peso corporeo, rispettivamente) e confermare che il topo non risponde pizzicando la zampa posteriore.
NOTA: sia i topi MADM maschi che i topi maschi (sfondo CD-1) vengono utilizzati per l'analisi. Se è necessaria la genotipizzazione, raccogliere le biopsie dell'orecchio a questo punto. - Posizionare l'animale anestesizzato in posizione supina sul vassoio per la chirurgia perfusione e disinfettare la pelliccia con il 70% di etanolo. Per iniziare l'intervento chirurgico, fare con attenzione un'incisione con forbici e cimici chirurgiche attraverso lo strato esterno della pelle e quindi una seconda incisione attraverso lo strato muscolare. Sollevare la punta dello sterno e tagliare il tessuto connettivo sui lati, prendendo particolare cautela per evitare di tagliare il fegato. La cavità toracica sarà visibile.
- Snip il diaframma e sollevare per rivelare il cuore. Tagliare con attenzione la gabbia toracica e fissare il vassoio chirurgico per esporre il cuore. Per i cuccioli, rimuovere completamente la gabbia toracica.
- Inserire un ago con salina tampone fosfato (PBS) nel ventricolo inferiore sinistro (tessuto più pallido). Utilizzando piccole forbici da iride fare un'incisione all'estremità posteriore dell'atrio destro (tessuto rosso scuro) per il sangue di drenare.
- Eseguire la perfusione con PBS seguito immediatamente da fresco, ghiacciato 4% paraformaldeide (PFA) preparato in PBS. Per i cuccioli di P4-P10 utilizzare le siringhe per eseguire la perfusione. Riempire una siringa con 10 mL di PBS e un'altra con 10 mL del 4% Di PFA. Assicurarsi che tutte le bolle d'aria nelle siringhe siano state rimosse. Per gli animali più anziani, utilizzare una pompa peristaltica.
- Iniziare a perfinare con PBS (10 mL a 2,4 mL/min in cuccioli; 20 mL a 4-6 mL/min per gli adulti che utilizzano una pompa peristale). Il fegato diventerà chiaro e giallo pallido se l'ago è posizionato correttamente.
- Una volta completato, rimuovere l'ago dai cuccioli e inserire l'ago contenente PFA nello stesso foro. Per gli adulti, interrompere la pompa peristaltica prima di scambiare la soluzione PBS con PFA ghiacciato, assicurandosi di evitare bolle nel tubo di assorbimento. Riprendere a perfinare con il PFA (10 mL a 2,4 mL/min in cuccioli; 30 mL a 4-6 mL/min per gli adulti che utilizzano una pompa peristaltica).
- Quando la perfusione è completa, decapitare il topo e rimuovere il cervello attraverso un'attenta dissezione. Trasferire il cervello al 4% PFA. Utilizzare almeno 5 volte i volumi cerebrali (cioè 5,10 mL di PFA in un tubo di centrifuga conica da 15 mL) e incubare durante la notte a 4 gradi centigradi per la fissazione postperfusione per garantire la completa fissazione del tessuto. Continuare con il passaggio 3.10.
- Per il tessuto embrionale e il tessuto postnatale precoce (cioè P0-P3), dopo aver eseguito una sezione cesarea, decapitare gli embrioni con le forbici. Se è necessaria la genotipizzazione, raccogliere la coda dell'embrione a questo punto. Sezionare immediatamente il cervello e trasferire a una piastra di 12 pozzetti contenente 2-3 mL di 4% PFA/pozzo. Incubare pernottamento a 4 gradi centigradi per la post-fissazione.
- Cambio del mattino successivo PFA con 10 mL (adulto) o 2-3 mL (embrione) di PBS e ripetere il lavaggio 3x per 15 min a RT. Trasferire il tessuto al 30% soluzione di saccarosio (PB) e conservare a 4 gradi centigradi su una piattaforma a dondolo fino a quando il tessuto affonda nella soluzione.
- Incorporare il cervello in composto di temperatura di taglio ottimale (OCT) in uno stampo di incorporamento, avendo cura di orientare il cervello per sezionamento coronale o sagittale. Congelare posizionando lo stampo che incorpora sul ghiaccio secco fino a quando lo OCT non diventa completamente opaco ( . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Conservare il tessuto a -80 gradi centigradi fino a un ulteriore utilizzo.
4. Preparazione del tessuto MADM per l'immunohistochimica
- Collegare il blocco di tessuto al disco campione nel criostato applicando un anello dello Strumento di personalizzazione di Office sul disco e posizionando il blocco direttamente nello Strumento di personalizzazione di Office quando inizia a congelarsi. Assicurarsi che il blocco sia orientato correttamente per il piano di taglio desiderato.
NOTA: Qui, la sezionamento coronale per studiare i cloni corticali MADM è descritta in dettaglio. - Impostare la temperatura del blocco nel criostato su -20 gradi centigradi e la temperatura della lama a -21 gradi centigradi.
- Lasciare che il blocco di tessuto si adatti alla temperatura della camera montando il disco del provino al supporto del campione e lasciare in criostat per 5 min prima di iniziare la sezionamento.
- Tagliare il blocco in sezioni spesse (45-60 m) fino a raggiungere la regione tissutale di interesse.
- Una volta che il bordo della corteccia è chiaramente visibile, interrompere il sezionamento e bloccare la lama. Assicurarsi che la lama sia schermata prima di tagliare il blocco.
- Tagliare lo strumento di produzione dello strumento di produzione dello strumento Office in eccesso che circonda il tessuto con una lama, lasciando 1 x 2 mm di OCT su tutti i lati del cervello.
- Poi orientare il blocco in modo che uno dei bordi laterali della corteccia sia orientato verso il basso e l'altro verso l'alto (cioè, il bordo più rostrale della corteccia è puntato a destra).
- Iniziare la sezionamento con uno spessore di 45 m per i cloni adulti e di 30 m per i cloni embrionali. Eseguire ogni sezione singolarmente e utilizzare un piccolo pennello per mantenere l'area sotto il coltello pulita da eventuali detriti lasciati durante il taglio del blocco.
NOTA: Se questo non viene fatto e una sezione cade, potrebbe essere difficile determinare l'ordine corretto delle sezioni. - Se le sezioni iniziano ad arricciarsi, tagliare i bordi del blocco e/o regolare con attenzione la piastra antiroll di vetro.
- Per l'analisi dei cloni embrionali, montare le sezioni direttamente su uno scivolo smerigliato. Asciugare su una piastra di riscaldamento a 37 gradi centigradi prima di procedere direttamente al passaggio 5.6.
NOTA: È possibile aggiungere più sezioni a una diapositiva, ma assicurarsi che l'ordine sequenziale venga mantenuto. - Per raccogliere cloni adulti, prepara 24 lastre di pozzo contenenti 1 mL di PBS/pozzo (tipicamente, piastre di 5/6 per cervello). A partire dal primo pozzo, con le pinze fredde raccogliere singole sezioni seriali in PBS nell'ordine di sezionamento.
NOTA: Il metodo di sezione galleggiante viene adottato per il tessuto adulto per garantire che nessuna sezione venga persa e che le sezioni montate non contengano rughe. - Interrompere la sezionamento una volta raggiunta la fine della neocorteccia.
- Per i cloni per adulti, procedere al montaggio di sezioni galleggianti.
NOTA: Le sezioni possono essere conservate in PBS a 4 gradi centigradi per un massimo di 24 h.
5. Montaggio di tessuto adulto per l'imaging
NOTA: sono necessari i seguenti strumenti: pennello piccolo, parabola Petri, PBS con 0,5% Tween (PBS-T), diapositive di adesione (Tabella dei materiali), supporto di montaggio ( Tabella deimateriali), coverlips e pinze.
- Riempire un piatto Petri con PBS-T.
NOTA: Detergent viene utilizzato per facilitare il processo di montaggio. Se è necessario colorare per antigeni aggiuntivi sensibili ai detergenti (ad esempio, glicoproteine), è meglio saltare l'aggiunta di Interpol. - Inserire una diapositiva di adesione nel PBS-T in modo che sia quasi coperta fino all'etichetta.
- Trasferire la prima sezione nel PBS-T.
- Utilizzando un piccolo pennello, manovrare la sezione sul vetrino e disporla per preservare l'ordine di taglio. Procedere nello stesso modo con tutte le altre sezioni.
- Una volta che tutte le sezioni sono in posizione, posizionate la diapositiva (12-16 sezioni/diapositiva) in una camera di scorrimento scura. Sollevare leggermente il coperchio per consentire alle sezioni di asciugarsi completamente (10 o 20 min), assicurandosi che rimangano adere nei passi successivi.
- Se si esegue l'immunohistochimica per antigeni aggiuntivi, procedere direttamente alla sezione 6 o 7.
NOTA: Per i tempi embrionali, è necessario eseguire passaggi immunostaining per almeno GFP e tdT (sezione 6). Per i cloni adulti, questo è necessario solo se la colorazione per gli antigeni aggiuntivi in parallelo (sezioni 6 e 7). - Reidratare e lavare le sezioni 1x con 1x PBS per 5 min per rimuovere il PBS-T residuo. Applicare sulla diapositiva 1 mL di 4,6-diamidino-2-phenylindole (DAPI) diluito in 1 x PBS (1 g/mL), assicurando che tutte le sezioni siano coperte e incubate per 15 min.
- Rimuovere con cura il DAPI e lavare 1x con 1x PBS per 5 min. Rimuovere il PBS in eccesso e asciugare per 1/2 min prima di incorporarlo in 110 gradi di supporto di montaggio. Sigillare con un coperchio di 24 x 60 mm e lasciare asciugare per almeno 3 h prima dell'imaging.
6. Immunostaining solo per GFP e tdT
NOTA: Questa sezione è necessaria per i cloni embrionali.
- Posizionare i vetrini orizzontalmente in una camera di incubazione dei vetrini umidificata. Contrassegnare i contorni della diapositiva con un indicatore di cera per ridurre al minimo la quantità di buffer necessaria.
- Reidratare le sezioni con 1x PBS. Per migliorare la qualità della colorazione, lavorare con tessuto appena sezionato.
- Aggiungere in 1x PBX il numero di buffer di blocco (0,5% Triton X-100, siero d'asino normale del 2-3% in 1 x PBS) per diapositiva, assicurandosi che tutte le sezioni siano coperte. Incubare per 1 h.
NOTA: La concentrazione di detergente (Triton X-100 o Tween-20) varia a seconda degli anticorpi primari aggiuntivi utilizzati perché alcuni antigeni sono più sensibili ai detergenti rispetto ad altri. - Rimuovere il buffer di blocco e aggiungere gli anticorpi primari nel buffer di blocco alla diapositiva (300-400 l/diapositiva).
NOTA: un esempio di reazione anticorpale primaria standard per anti-GFP/anti-tdT (MADM) potrebbe utilizzare pollo anti-GFP (1:500) e coniglio anti-RFP (1:500). - Incubare con anticorpi primari durante la notte a 4 gradi centigradi.
NOTA: I vetrini devono essere incubati perfettamente orizzontalmente con tampone che copre tutte le sezioni. In caso contrario, può verificarsi una colorazione irregolare o scarsa. - Verificare la mattina successiva che il buffer di blocco con anticorpi primari copre ancora tutte le sezioni del vetrino. In caso contrario, ripetere la fase di incubazione per 3/4 h a RT.
- Rimuovere gli anticorpi primari e lavare 4x con 1x PBS per 10 min a RT.
- Aggiungere gli anticorpi secondari diluiti nel buffer di blocco per scivolare (300-400 -L/diapositiva): Alexa Fluor 488 anti-pollo IgG (1:500) e Cy3 anti-coniglio Ig (1:500).
- Incubare a RT per 2 h. Tenere i vetrini coperti dalla luce per evitare lo sbiancamento dei fluorofori.
- Rimuovere gli anticorpi secondari e lavare 2x con 1x PBS per 10 min.
- Incubazione con DAPI diluita in PBS (1:5,000) per 15 min.
- Lavare le sezioni 1x con 1x PBS per 10 min.
- Rimuovere il PBS in eccesso e asciugare per 1/2 min prima dell'incorporamento in 110 gradi l di supporto di montaggio.
- Sigillare con coperchio 24 x 60 mm e lasciare asciugare per almeno 3 h prima dell'imaging. Le diapositive dell'immagine entro 1o2 settimane dall'esecuzione dell'immunoistochimica per garantire un segnale ottimale.
7. Immunostaining per GFP, tdT e antigeni aggiuntivi
- Eseguire i passaggi 6.1/6.3.
- Rimuovere il buffer di blocco e aggiungere gli anticorpi primari nel buffer di blocco alla diapositiva (300-400 l/diapositiva).
NOTA: Quando la colorazione per tre o più antigeni (ad esempio, GFP, tdT e una proteina di interesse) e l'anticorpo per la proteina di interesse è stato sollevato nel coniglio, si consiglia di utilizzare l'anticorpo primario anti-tdT (capra) con una diluizione di 1:500. Un esempio di reazione anticorpale primaria per tre antigeni con colorazione tdT alternativa potrebbe utilizzare pollo anti-GFP (1:500), capra anti-tdT (1:500) e anticorpi contro la proteina di interesse (cioè coniglio). - Eseguire i passaggi 6.5-6.7.
- Aggiungere un mix di anticorpi secondario diluito nel buffer di blocco per scivolare (300-400 l/diapositiva): Alexa Fluor 488 anti-pollo IgG (1:500), Cy3 anti-capra IgG (1:500) e Alexa Fluor 647 anti-coniglio IgG (1:500).
- Eseguire i passaggi 6.9-6.14.
8. Acquisizione e quantificazione di immagini confocali di cloni MADM
- Identificare e documentare le sezioni cerebrali contenenti cloni e le relative posizioni nella corteccia.
NOTA: il numero di sezioni di un intervallo di cloni varia a seconda di quando il clone è stato indotto, del driver CreERT2 e del tempo di analisi. Questo passaggio può essere eseguito sia su un microscopio confocale che su un microscopio a epifluorescenza. - Utilizzando un microscopio confocale invertito, iniziare selezionando e configurando le linee laser e i filtri corretti. Per i cervelli MADM, selezionare DAPI, GFP e tdT (eccitazione: 358 nm, 488 nm e 554 nm, rispettivamente; picco di emissione: 461 nm, 507 nm e 581 nm, rispettivamente). Assicurarsi che il foro stenopeico sia impostato su 1 unità ariosa per una qualità di imaging ottimale.
- Per impostazioni specifiche confocali, cloni di immagini con un obiettivo 20x e zoom 1x. Per le immagini che verranno utilizzate nelle quantificazioni, utilizzare un valore di afconsiderazione pixel della velocità di scansione di 1,52 s 2,06 s (valori 7-8 nel software di acquisizione delle immagini) senza una media. Regolare l'intensità del laser e le impostazioni di guadagno per ogni canale in base alle esigenze.
NOTA: a seconda della qualità dell'immagine richiesta, le impostazioni per la velocità di scansione e la media possono variare. - Una volta identificato chiaramente il clone, disporre i riquadri di imaging in modo da coprire tutte le sezioni rilevanti del clone. Regolare lo z-stack in modo che tutte le celle etichettate MADM nel clone vengano acquisite con un intervallo di 1,5 sm/z-stack slice. Regolare l'area affiancata in modo che l'intera larghezza della corteccia venga acquisita durante l'imaging del clone (ad esempio, dalla superficie del pial al corpo calloso).
- Immagine singoli cloni che si estendono su più sezioni consecutivamente, assicurando che tutte le sezioni senza celle all'interno di un clone siano ancora immagini allo scopo di ricostruire 3D e una corretta interpretazione delle informazioni spaziali delle celle.
- Analizzare ogni sezione contenente le cellule di un clone MADM in sequenza dal rostral all'estremità caudale della corteccia. Distinguere i singoli neuroni e glia in base alla loro morfologia e/o colorazione marcatore. Registrare le informazioni posizionali in parallelo in base ai rispettivi limiti dei livelli definiti dalla colorazione nucleare (DAPI).
NOTA: Vedere la figura 4 per i risultati rappresentativi per l'analisi embrionale e la figura 5 per i risultati rappresentativi per l'analisi degli adulti.
9. Ricostruzione 3D seriale dei cloni
NOTA: La ricostruzione 3D dei singoli cloni visualizzati su sezioni cerebrali seriali è utile per la visualizzazione visiva e l'analisi delle architetture clonali 3D e può essere eseguita secondo i seguenti passaggi.
- Stitch e fondere immagini confocali piastrellate in base ai parametri di acquisizione utilizzando il software di acquisizione delle immagini. Aprire il file .czi e quindi eseguire il metodo di cucitura nella scheda Elaborazione nel software .ens.
- Esportare pile di immagini cucite come singoli piani z in formato TIFF. Aprire il file .czi ecucito, quindi eseguire il metodo di esportazione delle immagini nella scheda Elaborazione. Per le immagini multicanale, esportare come immagini rosse/verdi/blu per la successiva elaborazione delle immagini.
- Ripetere i passaggi 9.1 e 9.2 per ogni sezione del cervello seriale di un clone.
NOTA: per una ricostruzione 3D accurata, tutte le sezioni cerebrali all'interno di un clone, comprese quelle senza cellule etichettate, devono essere elaborate. - Concatena le singole immagini in un'unica pila per poter passare dal piano z più saturdale a quello più caudale utilizzando un software di elaborazione delle immagini open source come ImageJ/Fiji67,68.
NOTA: Tutte le immagini vuote ai bordi di ogni sezione del cervello devono essere rimosse a questo punto. - Se necessario, correggere lo stack di immagini ottenuto dal passaggio 9.4 per l'errata allineamento utilizzando un plug-in ImageJ denominato "MultiStackReg" seguendo i passaggi 9.5.1.9.5.5. Se l'allineamento dell'immagine non è necessario, procedere al passaggio 9.6.
NOTA: Questo plugin esegue l'allineamento dell'immagine del canale con il massimo contrasto (di solito DAPI) e quindi applica la trasformazione registrata agli altri canali, consentendo così l'allineamento affidabile dell'immagine degli stack multicanale. Un plugin ausiliario chiamato "TurboReg" deve essere preinstallato.- In ImageJ, installare i plugin "MultiStackReg" e "TurboReg".
- Aprire la pila di immagini di immagini clone ottenute dal passaggio 9.4 da allineare. Suddividere i canali in DAPI (blu), GFP (verde) e tdT (rosso) all'interno dell'opzione Colore nella scheda Immagine.
- Eseguire ilplug-in " MultiStackReg" per allineare il canale DAPI mediante la trasformazione "Corpo rigido" e salvare il file di trasformazione.
- Applicare il file di trasformazione salvato agli altri due canali utilizzando "MultiStackReg".
- Unisci tutti e tre i canali allineati e salva la pila allineata.
- Per orientare il clone nella pila di rotazione ImageJ di immagini clone ottenute dal passo 9.4 (o passo 9.5.5 dopo l'allineamento) in un orientamento verticale con la superficie del pial in alto e il corpo calloso in basso. Se necessario, ritagliare il piano xy.
- Sia per la presentazione qualitativa che per l'analisi quantitativa, generare un'immagine di proiezione z massima (passaggio 9.8) o eseguire il rendering 3D (passaggio 9.9) del clone.
- In ImageJ, aprite la pila di immagini dal punto 9.6 e selezionate l'opzione di proiezione z con il tipo di proiezione Intensitàmassima . Questo genererà un'immagine dell'intero clone proiettato sullo stesso piano.
- In ImageJ, aprire la pila di immagini dal passaggio 9.6 e selezionare la funzione z progetto 3D per generare una visualizzazione 3D del clone che può essere ruotata.
NOTA: in questo passaggio è importante immettere l'intervallo di sezione corretto equivalente allo spessore dei singoli z-stack durante l'acquisizione dell'immagine. Lo strumento di interpolazione deve essere utilizzato per rimuovere gli spazi tra le sezioni.
Risultati
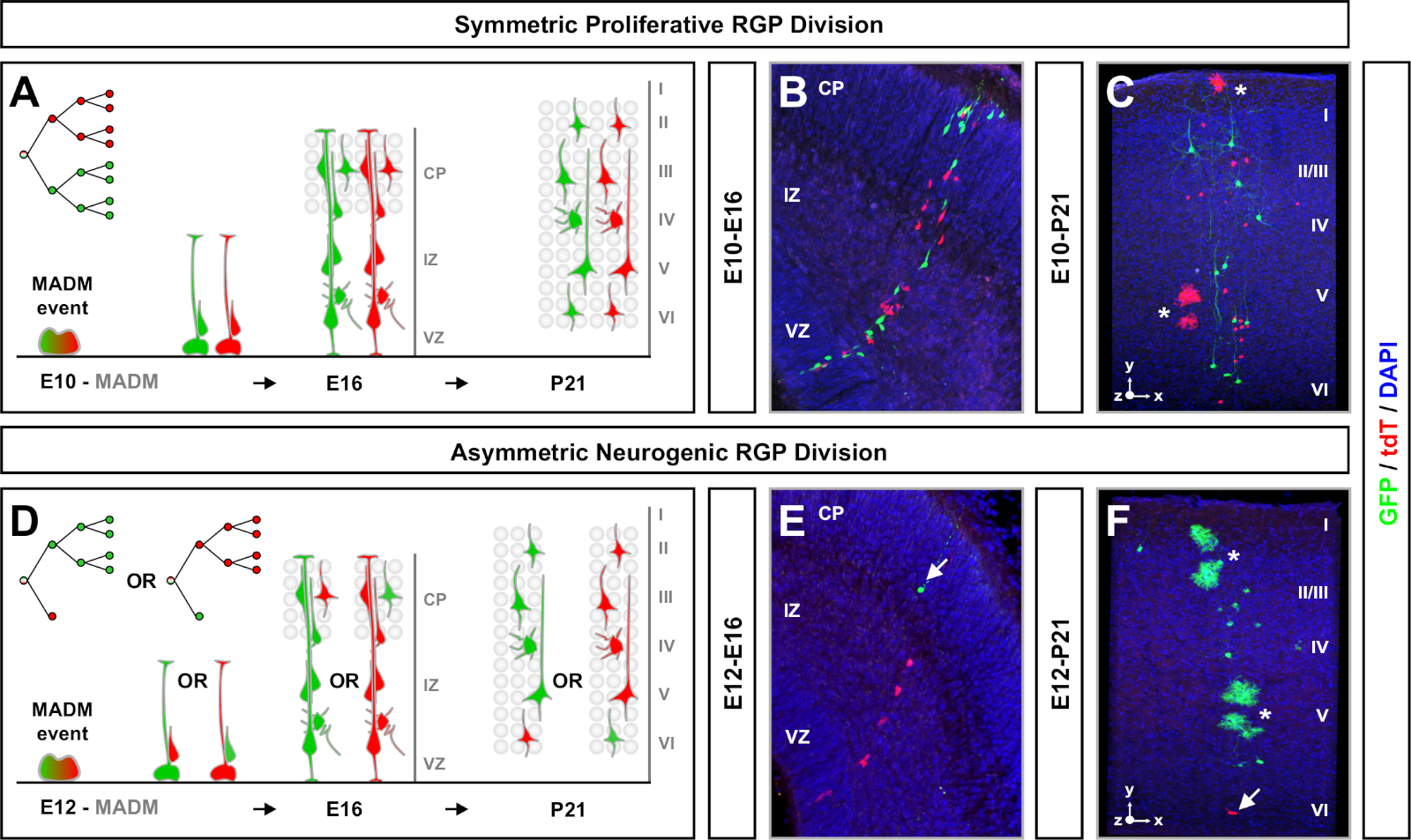
MADM si traduce nella ricostituzione di proteine fluorescenti verdi e rosse funzionali con due cellule figlie ciascuna che esprimono una delle due proteine fluorescenti in caso di eventi di segregazione cromosomica G2-X (Figura 1A). Poiché gli eventi MADM si traducono in un'etichettatura permanente e distinta dei due lignaggi discendenti, è possibile eseguire una valutazione quantificabile dei lignaggi di cellule verdi e rosse (subcloni). È possibile determinare le variabili, tra cui il modello di divisione (ad esempio, simmetrico o asimmetrico) e il potenziale (ad esempio, il numero di progenie) del progenitore originale. Quantificare ogni subclone fluorescente è informativo quando si determina in modo retroattivo se la cellula progenitore originale è sottoposta a divisioni proliferanti simmetriche o divisioni neurogeniche asimmetriche al momento dell'induzione di TM. Studi precedenti hanno raggruppato Emx1-CreERT2 o Nestin-CreERT2 derivati cloni di proiezione eccitatori nella corteccia in due ampie classi7,11,46. I primi, ricorsi "cloni proliferativi simmetrici", sono composti in media da un numero considerevole di neuroni, con subclone sia verdi che rossi contenenti quattro o più neuroni ciascuno. Il secondo gruppo, "cloni asimmetrici" definisce una classe di cloni in cui il subclone "minoranza" contiene meno di tre neuroni e il subclone "maggioranza", quattro o più11. Queste definizioni sono specifiche per GLI RG corticale e potrebbero aver bisogno di essere rivisitate per altre regioni e tessuti del cervello. Per entrambe le classi di cloni corticali, la progenie sarà distribuita in tutti gli strati superficiali e profondi.
Quando si progettano studi clonali MADM ci sono una serie di aspetti che devono essere presi in considerazione. Il momento in cui gli eventi MADM vengono indotti dalla somministrazione di TM è una considerazione chiave (Figura 3). Per i cloni MADM del neurone della proiezione eccitatoria corticale (ad esempio, utilizzando Emx1-CreERT2 o Nestin-CreERT2) a E10, quasi tutti gli RGP erano ancora in fase di divisione simmetrica11. Pertanto, l'induzione a E10 con TM ha catturato più cicli di amplificazione RGP proliferante e ha portato a cloni con un alto numero di neuroni. Tuttavia, il numero di RGP all'E10 era generalmente ridotto e quindi la somministrazione di TM generava pochissimi eventi MADM (a volte meno di uno per cervello). La maggior parte degli RGP è passata da divisioni neurogeniche simmetriche a asimmetriche a circa E12. Per colpire cloni neurogenici strettamente asimmetrici, era meglio indurre a E12 o versioni successive (Figura 3). Il tempo che intercorre tra l'induzione della TM e l'osservazione degli eventi di ricombinazione MADM nella corteccia tendeva ad essere inferiore a 24 h. Le iniezioni di IP erano il metodo preferito per la somministrazione di TM nelle fasi embrionali di questo metodo perché ha portato a una maggiore riproducibilità nell'induzione clonale. È anche importante mantenere la dose di TM al minimo per due motivi. In primo luogo, se il tasso di ricombinazione MADM aumenta, la probabilità di indurre più cloni, forse sovrapposti, è maggiore. In secondo luogo, se viene erogata troppa TM, si può osservare un aumento del tasso di aborto, la riassorbimento degli embrioni e le dimensioni più piccole della lettiera. Sono stati osservati aborti in circa la metà di tutte le dighe in gravidanza quando sono state somministrate iniezioni di TM all'E10. Questa frequenza è diminuita da E11 in poi e è diminuita a circa 1/3 dell'interruzione delle dighe incinte. Per una sintesi delle dosi di TM, dei tempi di induzione e dei driver CreERT2 utilizzati negli studi MADM precedenti, fare riferimento alla Tabella 1. L'attività di reporter in assenza di TM è stata osservata con alcuni driver CreERT2 tm-inducible69. L'espressione ectopica o gli eventi di ricombinazione MADM in assenza di TM non sono stati osservati con l'Emx1-CreERT2 dei driver Nestin-CreERT2. Ciò può essere in parte dovuto al fatto che le trans-ricombinazioni cromosomiche mediate da TM si verificano a circa 1:1.000-1:10.000 frequenza inferiore rispetto alle cis-recombinations, riducendo la probabilità di etichettatura MADM ectopica.
Un altro fattore da considerare quando si pianifica un esperimento di analisi clonale MADM è la durata dello studio. Variando la durata dell'induzione di TM e quando l'esperimento è stato analizzato (A) (finestra temporale) viene visualizzata la dinamica delle cellule staminali nel tempo64. Brevi intervalli di tempo embrionali (ad es. TM/E11-A/E13; La TM/E11-A/E16) ha catturato le dinamiche della neurogenesi embrionale (Figura 4). Il confronto di cloni di due o più finestre temporali fornisce una comprensione quantitativa del numero di cellule prodotte e di come la distribuzione dei neuroni varia nelle diverse fasi della progressione del lignaggio64. Per cogliere l'intero potenziale dei singoli cloni, è necessario estendere l'intervallo di tempo analizzato in timepoint postnatali o per adulti7,11,12. Esempi di cloni neocorticali indotti nell'embrione e analizzati nell'adulto sono mostrati nella Figura 5. Da nota, la neurogenesi corticale è per lo più completata e la gliogenesi aumenta di E17. Circa 1/6 RGP neurogenico anche procedere a generare astrociti e/o oligodendrociti11.
I cloni simmetrici si verificano quando gli RGP subiscono uno o più cicli di divisione prolifera11. I cloni RGP indotti tra E10 e12 erano in media di dimensioni maggiori e fornivano più caratteristiche spaziali della distribuzione finale dei neuroni (Figura 4A-C). I cloni con neuroni relativamente equamente distribuiti in strati profondi e superficiali hanno assunto una forma "cilindro", mentre i cloni con neuroni più dispersi negli strati superficiali rispetto agli strati più profondi hanno sviluppato una forma "cono"11. Per catturare completamente le informazioni spaziali e morfologiche di un clone, è stato necessario ricostruire computazionalmente ogni clone utilizzando immagini sequenziali. Per misurare la dispersione clonale, la dispersione laterale massima (misurata in tutte le dimensioni) in strati superficiali (LII-VI) di un clone è stata confrontata con la dispersione del neurone negli strati profondi (LV/LIV). Questo rapporto (distribuzione superiore:distribuzione inferiore) ha fornito una lettura quantificabile della forma complessiva del clone.
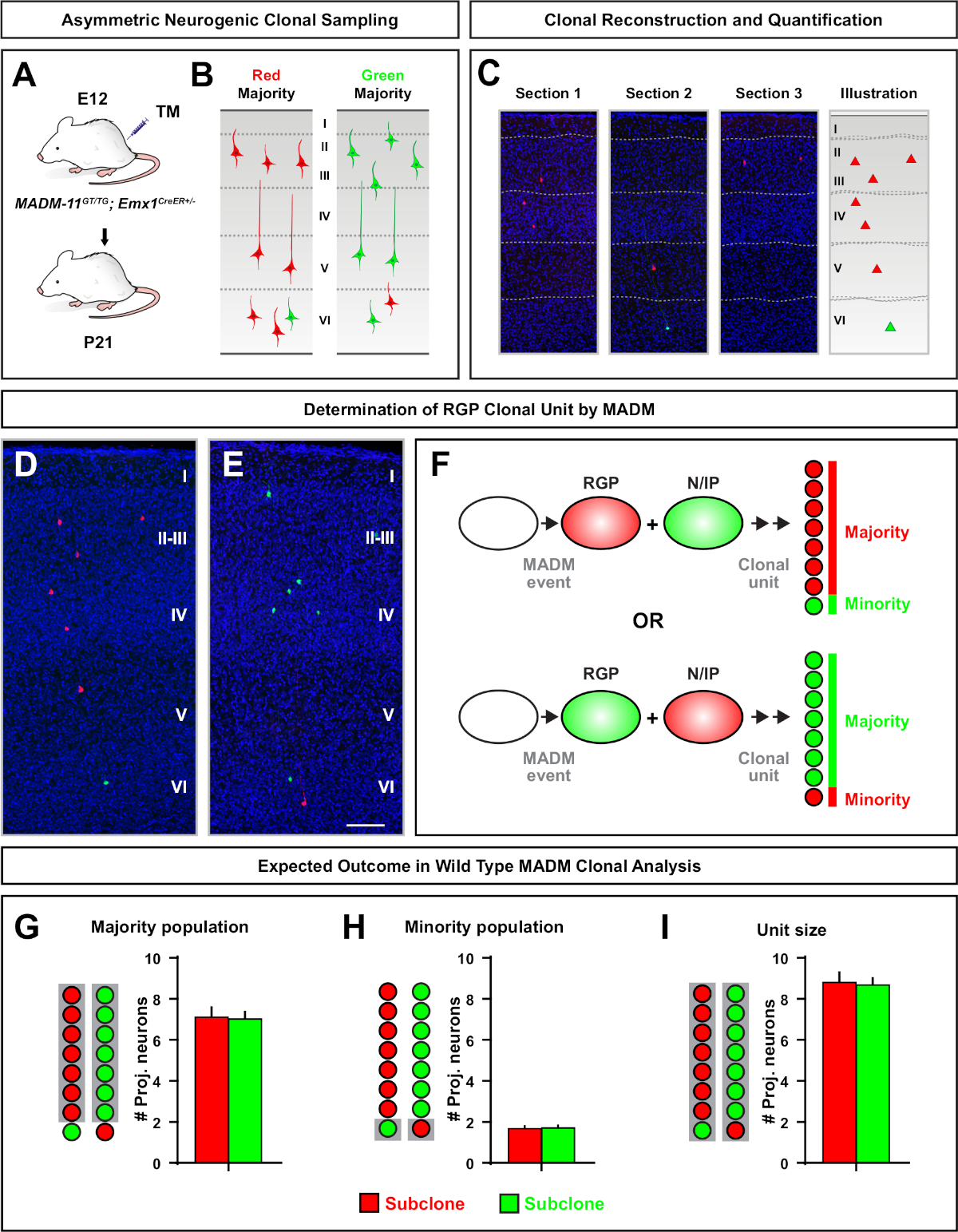
I cloni asimmetrici, in cui il sottoclone di minoranza era tre o meno, fornivano informazioni sull'output neuronale di un singolo RGP (Figura 4D-F e Figura 5A-F)7,11,12. La maggioranza della popolazione (grande subclone) potrebbe essere etichettata in rosso o verde, con una media di circa sette neuroni di proiezione eccitatori per clone se indotta utilizzando un Emx1-CreERT2 o Nestin-CreERT2(Figura 5G)7,11,12. Il numero totale di cellule in un clone DI MADM potrebbe essere ulteriormente sezionato analizzando la distribuzione dei neuroni nel grande subclone attraverso strati superficiali e profondi. La popolazione minoritaria (piccolo subclone) era etichettata con il colore reciproco ed era in media 1/2 cellule per clone (Figura 5H). La "dimensione unitaria" totale, che era in media di 8-9 neuroni, potrebbe essere calcolata sommando i subclone piccoli e grandi insieme (Figura 5I)7,11,12. È importante notare che mentre l'uscita neuronale degli RGP era altamente prevedibile, c'era un grado di eterogeneità clonale12,70.
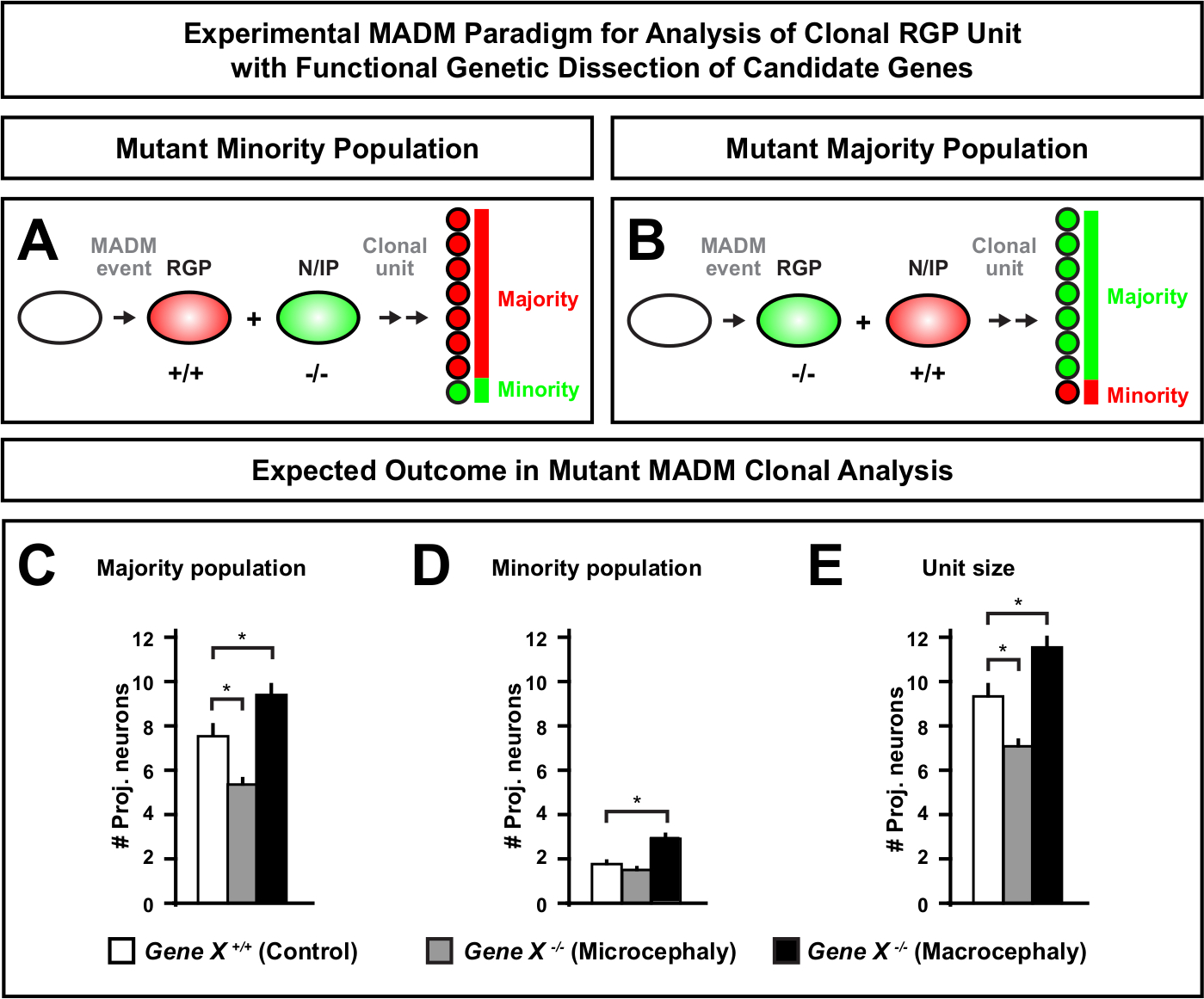
L'introduzione di una mutazione distale nella cassetta MADM consente la generazione di mosaici genetici, fornendo un metodo unico per sezionare i regolatori molecolari della progressione del lignaggio delle cellule staminali. Come tale, il MADM fornisce una piattaforma sperimentale senza precedenti per studiare la funzione cellulare-autonoma di un gene (ad esempio, la sua associazione alla microcefalia o alla macrocefalia). Confrontando i cloni indotti in un mosaico genetico MADM con i cloni indotti in un MADM di controllo, è possibile generare una lettura altamente quantitativa dei cambiamenti nei numeri dei neuroni e nella distribuzione. Precedenti studi basati su MADM hanno quantificato la funzione cellulare-autonoma di Otx1 nella formazione di microcefalia a livello clonale (vedere la figura 6A-E per un esempio rappresentativo)11. In un altro studio, l'analisi clonale MADM ha dimostrato che Ndel1 non regola autonomamente i numeri dei neuroni di proiezione, ma invece la capacità dei neuroni neonati di entrare o migrare all'interno della placca corticale, che in seguito forma la corteccia adulta46. Questi studi hanno dimostrato la natura altamente quantitativa dell'analisi clonale MADM nello studio delle funzioni cellule-autonome dei geni che regolano lo sviluppo corticale. Attualmente non ci sono esempi in letteratura che utilizzino il MADM per studiare i geni implicati nella macrocefalia a livello clonale. Tuttavia, in studi futuri l'analisi dei geni rilevanti per il controllo delle dimensioni corticali in generale può fornire informazioni altamente desiderabili a livello molecolare e cellulare.

Figura 1: Il principio MADM per la tracciatura del lignaggio e l'analisi clonale a livello di singola cellula staminale. (A) Per eseguire l'analisi del lignaggio e l'analisi clonale con MADM, devono essere presenti due componenti. In primo luogo, le cassette MADM devono essere destinate a loci identici sui cromosomi omologhi. Le cassette sono costituite da due geni di reporter fluorescenti chimerici, eGFP (verde, [G]) e dimero tandem Pomodoro (rosso, tdT[T]). La cassetta GT contiene il capoN di eGFP e il capo-C del tdT, separati da un intron contenente un sito loxP. La cassetta TG è costruita inversamente, con il capoN del tdT e il capo-C di eGFP. In secondo luogo, l'espressione di Cre ricombinase deve avvenire nella cella contenente le cassette MADM mirate. I siti loxP fungono da bersaglio per la ricombinazione intercromosomica mediata da Cre, con conseguente ricostituzione simultanea di entrambe le cassette di espressione. Se la ricombinazione avviene durante la fase G2 del ciclo cellulare seguita dalla segregazione X (G2-X), le due cellule figlie esprimeranno una delle due proteine fluorescenti. (B) Principio MADM per l'analisi genetica del mosaico a livello di singolo clone. Gli alleli mutanti (mutazioni puntiformi, delezioni, inserimenti, alleli condizionali con fianco a loxP come raffigurato nella Figura 1B,ecc.) possono essere introdotti distale nella cassetta TG-MADM tramite ricombinazione meiotica (vedi Figura 2 e Hippenmeyer et al.46 per i dettagli su come introdurre alleli mutanti nel sistema MADM). Se si verifica una trans-ricombinazione intercromromosomica mediata da G2-X Cre tra le cassette MADM, si traduce in una cellula mutante omozina GFP (GeneX-/-) per il gene di interesse e una cella di tipo selvaggio omozino tdT (GeneX)in un ambiente eterozigo senza etichettaggio46,477171. I risultati alternativi dell'etichettatura non utilizzati nell'analisi clonale (cioè le celle gialle) sono stati descritti in precedenza nei dettagli11,46,47. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Schemi di allevamento per la generazione di topi MADM sperimentali per il tracciamento del lignaggio. Schema di allevamento per la generazione di controllo MADM (A) e Gene X MADM (B) mouse sperimentali MADM per l'analisi clonale. Per ulteriori informazioni sui paradigmi di allevamento MADM, vedere Beattie et al.7 e Hippenmeyer et al.7,46. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: paradigmi del percorso temporale per l'analisi del lignaggio clonale basata su MADM. Schematici delle finestre temporali di progettazione sperimentale. Per i paradigmi di campionamento longitudinale, il punto temporale dell'induzione dei cloni è rimasto costante e la lunghezza del tempo prima che l'analisi variasse. Nel campionamento a intervalli progressivi, il timepoint dell'analisi è rimasto costante, ma il tempo di induzione variava. Una combinazione di uno o entrambi gli approcci può essere utilizzata a seconda delle domande affrontate. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Analisi clonale MADM nella neocorteccia in via di sviluppo e adulta. Induzione di cloni MADM mediati da TM in simmetricamente proliferazione (TM a E10) (A-C) e asimmetricamente neurogenica (TM at E12) (D-F) che divide RGP. Sono raffigurati singoli cloni MADM in vivo nello sviluppo (TM/E10-A/E16 e TM/E12-A/E16)eadulti (TM/E10-A/P21 e TM/E12-A/P21) (C,F) in MADM-11GT/TG; Nestin-CreERT2/- (B,E) e MADM-11GT/TG; Emx1-CreERT2/- (C,F). L'output dei neuroni era indipendente dal colore dei subclone e i subcloni della maggioranza/minoranza verde potevano essere confrontati con i subcloni della maggioranza/minoranza rossa nelle condizioni di controllo7,11. Circa 1/6 di cloni adulti contenevano anche astrociti e/o oligodendrociti, indicati da asterischi bianchi. I pannelli B e F sono riprodotti con il permesso rispettivamente di Hippenmeyer et al.46 e Rulands e Simons72. CP - Piastra corticale. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Analisi clonale MADM per quantificare l'output neuronale mediato da RGP. Analisi della produzione di neuroni eccitatori (unità) da singoli RGP neurogenici a livello clonale utilizzando MADM7,11. (A) Paradigma sperimentale per indurre cloni MADM per lo più asimmetrici nella corteccia in via di sviluppo. (B) Possibili esiti di cloni asimmetrici con la maggior parte dei subclone etichettati in verde o rosso (C) Sezioni consecutive rappresentative che abbracciano un singolo clone asimmetrico neurogenico (D,E) immagini di ricostruzione 3D di cloni di MADM g2-X asimmetrici rappresentativi con popolazione maggioritaria in rosso (D) o verde (E) in MADM-11GT/TG; Emx1-CreERT2/- con induzione TM all'E12 e analisi a P21. Si noti che le celle con etichetta verde e rosso sono di tipo selvaggio. (F)Schema che indica i due possibili risultati sperimentali del clone MADM. (G) Quantificazione delle dimensioni della maggioranza della popolazione derivante dal rinnovo degli AMP nei cloni MADM-11. HH) Quantificazione delle dimensioni della popolazione minoritaria derivante dal rinnovo degli RGP nei cloni MADM-11. (I) Quantificazione delle dimensioni unitarie dei cloni neurogenici simmetrici MADM-11. I valori ipotetici potrebbero rappresentare la barra della scala media , ovvero 100 m (D ed E). TM - Tamoxifene. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 6: Analisi clonale DEL MADM per studiare i geni che portano alla microcefalia e alla macrocefalia. Risultati ipotetici dell'analisi clonale MADM durante l'esecuzione della dissezione genetica funzionale dei geni candidati che portano alla microcefalia o alla macrocefalia. Per sezionare le funzioni cellulari-autonome di un gene di interesse (Gene X) sulla produzione neuronale, MADM richiede alleali mutanti da introdurre distale alle cassette MADM tramite ricombinazione meiotica (per i dettagli come introdurre alleli mutanti nel sistema MADM vedi anche Figura 2, Hippenmeyer et al.46e Laukoter et al.46,73). (A,B) Schematico che indica il paradigma sperimentale MADM per l'analisi funzionale delle unità RGP clonali. Il sottoclone mutante può formare la popolazione minoritaria (A) o maggioranza (B). (C-E) Ipotetici risultati dell'analisi clonale MADM durante la quantificazione del controllo MADM (barre bianche), microcefalia Microcefalia Gene-X MADM (barre grigie) e Gene-X MADM macrocefali di barre nere). (C) Quantificazione delle dimensioni della maggioranza della popolazione. (D) Quantificazione delle dimensioni della popolazione minoritaria. (E) Quantificazione delle dimensioni unitarie dei cloni neurogenici asimmetrici. I valori ipotetici potrebbero rappresentare lo scenario medio (SEM. S ) in cui la differenza nel numero di cella dei subclone potrebbe raggiungere un significato, rispetto al controllo. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Tabella 1: studi clonali MADM in letteratura. Riassunto degli studi nella letteratura contenente esperimenti di lignaggio clonale MADM, tra cui il driver CreERT2 utilizzato, la dose di TM e il tempo di iniezione. Fare clic qui per visualizzare questa tabella (fare clic con il pulsante destro del mouse per scaricare).
Discussione
Viene descritto un metodo per utilizzare MADM per monitorare il lignaggio cellulare dei singoli RG in vivo nella neocorteccia in via di sviluppo. In combinazione con CreERT2,gli eventi MADM possono essere cronometrati con precisione, fornendo una lettura visiva altamente qualitativa e quantitativa dei modelli di divisione delle cellule staminali a livello di singola cellula. Titando la dose di TM consegnata, in una situazione ideale si può ottenere una media di meno di un clone per emisfero corticale, fornendo un'adeguata separazione spaziale per distinguere inmodo i singoli cloni. Mantenendo l'integrità dei tessuti, questo metodo acquisisce anche le informazioni essenziali per quanto riguarda la posizione, la morfologia e i numeri di cellule assolute. Cassette MADM su Chr. 117,11,12,46,56,57,su Chr. 751, e il MADM originale a Rosa2647,53,59 sono stati utilizzati negli studi di analisi clonale MADM. L'alta risoluzione delle singole cellule fornisce una visione senza precedenti sia della morfologia che del rapporto clonale delle cellule figlie e permette l'imaging dal vivo delle cellule staminali proliferanti e dei cloni emergenti46,52.
La sezione cesarea e la promozione dei cuccioli per l'analisi dei cloni ai tempi postnatali è un passo necessario e critico nel protocollo. A seconda dello stato di salute della diga incinta trattata con TM, potrebbe non essere necessario eseguire un taglio cesareo. Tuttavia, è ancora necessario allevare i cuccioli con una madre affidataria, perché la madre trattata in TM può avere problemi ad allattare. Non sono state osservate differenze nella necessità di affidarsi a diversi conducenti CreERT2. Sia le linee MADM che le madri affidatarie sono mantenute su uno sfondo di CD-1. Se il taglio cesareo non è necessario, la diga incinta trattata con TM utilizzata per generare cuccioli sperimentali può essere riutilizzata per ulteriori allevamenti sperimentali secondo i principi del 3R (si noti che questo può essere fatto solo se le licenze sperimentali animali approvano questa pratica). Le madri affidatarie possono essere utilizzate per l'affidamento dei cuccioli entro 2 giorni dal parto, ma sono stati osservati tassi di successo più elevati quando le madri affidatarie partoriscono lo stesso giorno dei topi sperimentali che devono essere promossi. Pertanto, è importante impostare accoppiamenti a tempo per le madri affidatarie in parallelo agli accoppiamenti sperimentali nel passaggio 1.1. Mantenere un numero simile a quello della lettiera della madre affidataria originale può migliorare il tasso di sopravvivenza dei cuccioli affidati, e quindi potrebbe essere necessaria la rimozione di alcuni di tutti i rifiuti originali. Ulteriori passaggi che possono migliorare l'adottazione includono lo sfregamento dei guanti dello sperimentatore con lettiera e cibo (per rimuovere il profumo dei guanti); strofinando delicatamente i cuccioli dopo la sezione cesarea con frammenti della lettiera sporca e del nido della madre adottiva; e il posizionamento dei cuccioli a stretto contatto con i cuccioli della madre affidataria prima del loro posizionamento nella gabbia dei topi adottivi.
Come in altri metodi di tracciamento del lignaggio basato su reporter, è necessario prestare attenzione quando si sceglie il driver CreERT2 ottimale per esperimenti clonali MADM. In primo luogo, il promotore utilizzato deve esprimere il ricombinarsi sia temporale che spaziale nella popolazione progenitrice di interesse. Trovare questo promotore può essere difficile, perché alcuni promotori possono cambiare i modelli di espressione o diventare silenziati in diverse fasi di sviluppo. Per migliorare la specificità del tipo di cellula sono stati utilizzati più ricombinasi specifici del sito, ciascuno guidato da promotori separati. Quando una o entrambe le ricombinazioni sono espresse nella stessa cella, questa etichetta la cella e la sua progenie con un reporter fluorescente74,75,76,77. In sintesi, è importante scegliere un driver CreERT2 specifico per la popolazione di progenitori analizzati.
Il passaggio più critico in questo metodo è l'identificazione di un clone, perché tutte le celle devono essere derivate senza ambiguità da un singolo evento di ricombinazione (passaggio 8.1). La titolazione della concentrazione di TM garantisce meno di un cluster di cellule rosse/verdi per emisfero cerebrale e massimizza la probabilità di analizzare un singolo clone (passaggio 2.2)7,11. I cloni devono essere scartati se i gruppi vicini di cellule si verificano entro 500 m dal clone di interesse. Pertanto, è importante esaminare diverse sezioni prima e dopo la comparsa di un clone per assicurarsi che non vi siano ulteriori eventi di ricombinazione nelle vicinanze. A causa del segnale più debole dei fluorofori, è necessario eseguire l'immunohistochimica per eGFP e tdT nei cloni embrionali (vedi sezione 6). Questo è raccomandato solo nei cloni adulti se gli antigeni aggiuntivi saranno coetichettati. Quando si immaginano i cloni, è importante catturare l'intera larghezza della corteccia in cui si trova il clone (cioè dalla superficie del pial al corpo calloso; vedi il punto 8.4) per non perdere alcuna cella. Ciò facilita anche l'allineamento dell'immagine durante l'elaborazione dell'immagine (sezione 9). La sezione 8 del protocollo richiede un microscopio confocale invertito, ma può essere adattata a seconda della configurazione del microscopio disponibile. La microscopia a epifluorescenza può essere utilizzata, ma la microscopia confocale è consigliata perché questo porta a una diminuzione della contaminazione della luce dall'esterno del piano di messa a fuoco. È inoltre importante che l'intensità e il guadagno del laser siano regolati in modo che le celle verdi, rosse e gialle possano essere identificate in modo inequivocabile. Indipendentemente dalla configurazione, si consiglia di utilizzare un obiettivo di almeno 20x per garantire la completa separazione spaziale delle celle strettamente posizionate. Oltre a registrare la profondità corticale di tutte le cellule (passaggio 8.6), le regioni corticali in cui si trovano i cloni devono essere identificate utilizzando un atlante cerebrale come l'atlante del cervello di Allen o altre mappe delle coordinate stereotassiche. Dovrebbe inoltre essere adottato un paradigma di denominazione dei file per assicurarsi che le immagini clonate siano facilmente identificabili. Le seguenti informazioni potrebbero essere incluse nella denominazione del file: ID immagine univoco, immagine data è stata scattata, genotipo di animale, età di induzione, età di analisi, numero di immagine in relazione al resto delle immagini dallo stesso clone.
L'introduzione di una mutazione distale a una cassetta MADM permette distintamente la generazione di mosaici genetici71 e permette la dissezione dei regolatori molecolari di lignaggio e diversità del tipo di cellula al livello clonale7,11,46,62. Per generare un mosaico genetico con MADM, le cassette MADM devono essere meioticamente collegate allo stesso cromosoma del gene di interesse (cfr. figura 2 per lo schema di allevamento). Questo limita l'attuale analisi clonale con MADM ai geni situati su Chr. 751, Chr. 1146, Chr. 1251e Chr. 6 distale al locus Rosa26 47. Studi futuri utilizzeranno cassette MADM mirate a qualsiasi cromosoma, permettendo l'analisi del mosaico di praticamente tutti i geni del genoma del topo a livello clonale.
Infine, il MADM non si limita all'analisi delle cellule progenitrici nella neocorteccia in via di sviluppo. Lo studio di molte nicchie di cellule staminali potrebbe beneficiare della capacità di risolvere le disposizioni spatiotemporali di cellule clonate correlate. Applicando MADM ad altre regioni del cervello, le condizioni di malattia (ad esempio, il cancro), o in altri tessuti47,50,51,52,53,54,55,56,57,58,59, gli studi hanno rivelato relazioni di lignaggio in cloni derivati da diverse classi di progenie e cellule staminali (vedere la tabella 1 per l'elenco corrente degli studi cloni MADM).50 Un'altra interessante applicazione futura di MADM è quella di combinarlo con ulteriori reporter funzionali o subcellulari, che aumenterebbero il grado di informazioni che possono essere acquisite dai cloni.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Ringraziamo tutti i membri del laboratorio Hippenmeyer per la discussione, lo strumento di bioimaging, la struttura di scienze biologiche e lo strumento pre-clinico di IST Austria per il supporto tecnico. Questo lavoro è stato sostenuto dai fondi istituzionali IST Austria; R.B. ha ricevuto il sostegno del programma lise-Meitner del Fondo scientifico austriaco (FWF) (M 2416); N.A ha ricevuto il sostegno del Fondo scientifico austriaco (FWF) Firnberg-Programm (T 1031); Il programma di ricerca e innovazione orizzonte 2020 dell'Unione europea ha ricevuto il sostegno del programma di ricerca e innovazione Orizzonte 2020 dell'Unione europea nell'ambito dell'accordo di sovvenzione Marie Skaodowska-Curie n. 754411 in qualità di borsista post-dottorato ISTplus; A.H. ha ricevuto il sostegno di un DOC (Dottorato dell'Accademia austriaca delle scienze). Questo studio è stato sostenuto anche dal Consiglio europeo della ricerca (CER) nell'ambito del programma di ricerca e innovazione Orizzonte 2020 dell'Unione europea (accordo di sovvenzione n. 725780 LinPro) a S.H.
Materiali
| Name | Company | Catalog Number | Comments |
| 1 mL tuberculin syringe (Omnifix Luer Lock) | Braun | 9204512N | |
| 1,4-diazabicyclooctane (DABCO) | Roth | 0718.2 | |
| 10 mL Syringe (Omnifix Luer Lock) | Braun | 8508429N | |
| 15 mL conical centrifuge | Sarstedt | 65.554.502 | |
| 24 multi-well dishes | Roth/Greiner Bio-one | CE56.1 | |
| 27- gauge x 3/4 needle (Sterican) | Braun | 16010256E | |
| Corn oil | Sigma | C8267-500ML | |
| Coverslips (24 x 60 mm #1) | Thermo Fisher Scientific (Menzel) | 15747592 | |
| Cryostat Cryostar NX70 | Thermo Fisher Scientific | 957000H | |
| Dako Pen (Wax marker) | Agilent | S200230-2 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Invitrogen | D1306 | |
| Disposable microtome blade (MX35 Ultra) | Thermo Fisher Scientific | 705830 | |
| Fine Forceps (Dumont #5) | Fine Science Tools (FST) | 11254-20 | |
| Glass anti-roll plate | Histocom | M 449980 | |
| Glycerol | Sigma | G5516 | |
| LSM 800 Confocal | Zeiss | ||
| Mounting medium | 25 mg/mL DAPCO, 6 g Glycerol, 2.4 g Mowiol 4-88, 6 mL dH2O, 12 mL 0.2 M Tris-HCl (pH 8.5) | ||
| Mowiol 4-88 | Roth | 0713.2 | |
| Normal donkey serum | Innovative Research | IGDNSER100ML | |
| Paraformaldehyde | Sigma | 441244-1KG | |
| Peristaltic pump 323E/D 400RPM | Watson-Marlow | 036.3124.00A | |
| Sucrose | Sigma | S8501-5KG | |
| Superfrost plus glass slides | Thermo Fisher Scientific | J1800AMNT | |
| Tamoxifen | Sigma | T5648 | |
| Tissue Embedding mold T-12 (22mm square) | Polysciences Inc. | 18986-1 | |
| Tissue-Tek O.C.T | Sakura | 4583 | |
| Triton X-100 | Sigma | T8787-250ML | |
| Trizma hydrochloride | Sigma | 93363 | |
| Tween-20 | Sigma | P9416-100ML | |
| Software and Plugins: | |||
| Fiji | 1.52p | Fiji | |
| MultiStackReg | 1.45 | Download link | |
| TurboReg | EPFL Bioimaging | ||
| Zen Blue | 2.6 | Zeiss | |
| Experimental Models: Organisms/Strains: | |||
| Mouse: Emx1-CreER | The Jackson Laboratory | JAX:027784 | |
| Mouse: MADM-11-GT | The Jackson Laboratory | JAX:013749 | |
| Mouse: MADM-11-TG | The Jackson Laboratory | JAX:013751 | |
| Primary antibodies: | |||
| Chicken anti-GFP 1:500 | Aves Labs | GFP-1020 | |
| Goat anti-tdTomato 1:500 | Sicgen Antibodies | AB8181-200 | |
| Rabbit anti-RFP 1:500 | MBL | PM005 | |
| Secondary antibodies: | |||
| Donkey Anti-Chicken Alexa Fluor 488 1:500 | Jackson Immuno Research | 715-475-150 | |
| Donkey Anti-Goat Cy3 1:500 | Jackson Immuno Research | 705-165-147 | |
| Donkey Anti-Rabbit Cy3 1:500 | Jackson Immuno Research | 711-165-152 |
Riferimenti
- Malatesta, P., et al. Neuronal or glial progeny: regional differences in radial glia fate. Neuron. 37 (5), 751-764 (2003).
- Miyata, T., Kawaguchi, A., Okano, H., Ogawa, M. Asymmetric inheritance of radial glial fibers by cortical neurons. Neuron. 31 (5), 727-741 (2001).
- Noctor, S. C., Flint, A. C., Weissman, T. A., Dammerman, R. S., Kriegstein, A. R. Neurons derived from radial glial cells establish radial units in neocortex. Nature. 409 (6821), 714-720 (2001).
- Taverna, E., Götz, M., Huttner, W. B. The Cell Biology of Neurogenesis: Toward an Understanding of the Development and Evolution of the Neocortex. Annual Review of Cell and Developmental Biology. 30 (1), 465-502 (2014).
- Desikan, R. S., Barkovich, A. J. Malformations of cortical development. Annals of Neurology. 80 (6), 797-810 (2016).
- Gao, R., Penzes, P. Common mechanisms of excitatory and inhibitory imbalance in schizophrenia and autism spectrum disorders. Current Molecular Medicine. 15 (2), 146-167 (2015).
- Beattie, R., et al. Mosaic Analysis with Double Markers Reveals Distinct Sequential Functions of Lgl1 in Neural Stem Cells. Neuron. 94 (3), 517-533 (2017).
- Hippenmeyer, S. Molecular pathways controlling the sequential steps of cortical projection neuron migration. Advances in Experimental Medicine and Biology. 800, 1-24 (2014).
- Lodato, S., Arlotta, P. Generating neuronal diversity in the mammalian cerebral cortex. Annual Review of Cell and Developmental Biology. 31, 699-720 (2015).
- Hansen, A. H., Duellberg, C., Mieck, C., Loose, M., Hippenmeyer, S. Cell Polarity in Cerebral Cortex Development-Cellular Architecture Shaped by Biochemical Networks. Frontiers in Cellular Neuroscience. 11, 176 (2017).
- Gao, P., et al. Deterministic progenitor behavior and unitary production of neurons in the neocortex. Cell. 159 (4), 775-788 (2014).
- Llorca, A., et al. A stochastic framework of neurogenesis underlies the assembly of neocortical cytoarchitecture. eLife. 8, e51381 (2019).
- Ma, J., Shen, Z., Yu, Y. C., Shi, S. H. Neural lineage tracing in the mammalian brain. Current Opinion in Neurobiology. 50, 7-16 (2018).
- Caviness, V., Takahashi, T., Nowakowski, R. Numbers, time and neocortical neuronogenesis: a general developmental and evolutionary model. Trends in Neurosciences. 18 (9), 379-383 (1995).
- Schmechel, D. E., Rakic, P. A Golgi study of radial glial cells in developing monkey telencephalon: morphogenesis and transformation into astrocytes. Anatomy and Embryology. 156 (2), 115-152 (1979).
- Kessaris, N., et al. Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nature Neuroscience. 9 (2), 173-179 (2006).
- Magavi, S., Friedmann, D., Banks, G., Stolfi, A., Lois, C. Coincident generation of pyramidal neurons and protoplasmic astrocytes in neocortical columns. The Journal of Neuroscience. 32 (14), 4762-4772 (2012).
- Anthony, T. E., Klein, C., Fishell, G., Heintz, N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 41 (6), 881-890 (2004).
- Voigt, T. Development of glial cells in the cerebral wall of ferrets: direct tracing of their transformation from radial glia into astrocytes. The Journal of Comparative Neurology. 289 (1), 74-88 (1989).
- Amberg, N., Laukoter, S., Hippenmeyer, S. Epigenetic cues modulating the generation of cell-type diversity in the cerebral cortex. Journal of Neurochemistry. 149 (1), 12-26 (2019).
- Beattie, R., Hippenmeyer, S. Mechanisms of Radial Glia Progenitor Cell Lineage Progression. FEBS letters. 591 (24), 3993-4008 (2017).
- Telley, L., et al. Temporal patterning of apical progenitors and their daughter neurons in the developing neocortex. Science. 364 (6440), eaav2522 (2019).
- Oberst, P., et al. Temporal plasticity of apical progenitors in the developing mouse neocortex. Nature. 573 (7774), 370-374 (2019).
- Telley, L., et al. Sequential transcriptional waves direct the differentiation of newborn neurons in the mouse neocortex. Science. 351 (6280), 1443 (2016).
- Deppe, U., et al. Cell lineages of the embryo of the nematode Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America. 75 (1), 376-380 (1978).
- Woodworth, M. B., Girskis, K. M., Walsh, C. A. Building a lineage from single cells: genetic techniques for cell lineage tracking. Nature Reviews Genetics. 18 (4), 230-244 (2017).
- Masuyama, N., Mori, H., Yachie, N. DNA barcodes evolve for high-resolution cell lineage tracing. Current Opinion in Chemical Biology. 52, 63-71 (2019).
- Legue, E., Joyner, A. L. Chapter Ten-Genetic Fate Mapping Using Site-Specific Recombinases. Methods in Enzymology. 477, 153-181 (2010).
- Postiglione, M. P., Hippenmeyer, S. Monitoring neurogenesis in the cerebral cortex: an update. Future Neurology. 9 (3), 323-340 (2014).
- Espinosa-Medina, I., Garcia-Marques, J., Cepko, C., Lee, T. High-throughput dense reconstruction of cell lineages. Open Biology. 9 (12), 190229 (2019).
- Hwang, B., et al. Lineage tracing using a Cas9-deaminase barcoding system targeting endogenous L1 elements. Nature Communications. 10 (1), 1234 (2019).
- Kalhor, R., et al. Developmental barcoding of whole mouse via homing CRISPR. Science. 361 (6405), eaat9804 (2018).
- García-Moreno, F., Vasistha, N. A., Begbie, J., Molnár, Z. CLoNe is a new method to target single progenitors and study their progeny in mouse and chick. Development. 141 (7), 1589-1598 (2014).
- Loulier, K., et al. Multiplex Cell and Lineage Tracking with Combinatorial Labels. Neuron. 81 (3), 505-520 (2014).
- Livet, J., et al. Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature. 450 (7166), 56-62 (2007).
- Snippert, H. J., et al. Intestinal Crypt Homeostasis Results from Neutral Competition between Symmetrically Dividing Lgr5 Stem Cells. Cell. 143 (1), 134-144 (2010).
- Amitai-Lange, A., et al. A method for lineage tracing of corneal cells using multi-color fluorescent reporter mice. Journal of Visualized Experiments. (106), e53370 (2015).
- Vasistha, N. A., et al. Cortical and Clonal Contribution of Tbr2 Expressing Progenitors in the Developing Mouse Brain. Cerebral Cortex. 25 (10), 3290-3302 (2015).
- Chen, F., LoTurco, J. A method for stable transgenesis of radial glia lineage in rat neocortex by piggyBac mediated transposition. Journal of Neuroscience Methods. 207 (2), 172-180 (2012).
- Siddiqi, F., et al. Fate mapping by piggyBac transposase reveals that neocortical GLAST+ progenitors generate more astrocytes than Nestin+ progenitors in rat neocortex. Cerebral Cortex. 24 (2), 508-520 (2014).
- Yoshida, A., et al. Simultaneous expression of different transgenes in neurons and glia by combining in utero electroporation with the Tol2 transposon-mediated gene transfer system. Genes to Cells. 15 (5), 501-512 (2010).
- García-Marqués, J., López-Mascaraque, L. Clonal Identity Determines Astrocyte Cortical Heterogeneity. Cerebral Cortex. 23 (6), 1463-1472 (2013).
- Clavreul, S., et al. Cortical astrocytes develop in a plastic manner at both clonal and cellular levels. Nature Communications. 10 (1), 4884 (2019).
- Kim, G. B., et al. Rapid Generation of Somatic Mouse Mosaics with Locus-Specific, Stably Integrated Transgenic Elements. Cell. 179 (1), 251-267 (2019).
- Pilz, G. A., et al. Live imaging of neurogenesis in the adult mouse hippocampus. Science. 359 (6376), 658 (2018).
- Hippenmeyer, S., et al. Genetic Mosaic Dissection of Lis1 and Ndel1 in Neuronal Migration. Neuron. 68 (4), 695-709 (2010).
- Zong, H., Espinosa, J. S., Su, H. H., Muzumdar, M. D., Luo, L. Mosaic analysis with double markers in mice. Cell. 121 (3), 479-492 (2005).
- Ramón y Cajal, S. . Histologie du système nerveux de l'homme et des vertébrés. , (1911).
- Cowan, W. M. The emergence of modern neuroanatomy and developmental neurobiology. Neuron. 20 (3), 413-426 (1998).
- Ali, S. R., et al. Existing cardiomyocytes generate cardiomyocytes at a low rate after birth in mice. Proceedings of the National Academy of Sciences of the United States of America. 111 (24), 8850-8855 (2014).
- Hippenmeyer, S., Johnson, R. L., Luo, L. Mosaic analysis with double markers reveals cell-type-specific paternal growth dominance. Cell Reports. 3 (3), 960-967 (2013).
- Riccio, P., Cebrian, C., Zong, H., Hippenmeyer, S., Costantini, F. Ret and Etv4 promote directed movements of progenitor cells during renal branching morphogenesis. PLoS Biology. 14 (2), e1002382 (2016).
- Bonaguidi, M. A., et al. In vivo clonal analysis reveals self-renewing and multipotent adult neural stem cell characteristics. Cell. 145 (7), 1142-1155 (2011).
- Mayer, C., et al. Clonally Related Forebrain Interneurons Disperse Broadly across Both Functional Areas and Structural Boundaries. Neuron. 87 (5), 989-998 (2015).
- Muzumdar, M. D., et al. Clonal dynamics following p53 loss of heterozygosity in Kras-driven cancers. Nature Communications. 7, 12685 (2016).
- Shi, W., et al. Ontogenetic establishment of order-specific nuclear organization in the mammalian thalamus. Nature Neuroscience. 20, 516 (2017).
- Wong, S. Z. H., et al. In vivo clonal analysis reveals spatiotemporal regulation of thalamic nucleogenesis. PLoS Biology. 16 (4), e2005211 (2018).
- Xu, H. T., et al. Distinct Lineage-Dependent Structural and Functional Organization of the Hippocampus. Cell. 157 (7), 1552-1564 (2014).
- Brennand, K., Huangfu, D., Melton, D. All β Cells Contribute Equally to Islet Growth and Maintenance. PLoS Biology. 5 (7), e163 (2007).
- Ortiz-Alvarez, G., et al. Adult neural stem cells and multiciliated ependymal cells share a common lineage regulated by the geminin family members. Neuron. 102 (1), 159-172 (2019).
- Kaplan, E. S., Ramos-Laguna, K. A., Mihalas, A. B., Daza, R. A. M., Hevner, R. F. Neocortical Sox9+ radial glia generate glutamatergic neurons for all layers, but lack discernible evidence of early laminar fate restriction. Neural Development. 12 (1), 14 (2017).
- Lv, X., et al. TBR2 coordinates neurogenesis expansion and precise microcircuit organization via Protocadherin 19 in the mammalian cortex. Nature Communications. 10 (1), 3946 (2019).
- Mihalas, A. B., Hevner, R. F. Clonal analysis reveals laminar fate multipotency and daughter cell apoptosis of mouse cortical intermediate progenitors. Development. 145 (17), dev164335 (2018).
- Picco, N., et al. A mathematical insight into cell labelling experiments for clonal analysis. Journal of Anatomy. 235 (3), 687-696 (2019).
- Johnson, C. A., Ghashghaei, H. T. Sp2 regulates late neurogenic but not early expansive divisions of neural stem cells underlying population growth in the mouse cortex. Development. , (2020).
- Espinosa, J. S., Luo, L. Timing Neurogenesis and Differentiation: Insights from Quantitative Clonal Analyses of Cerebellar Granule Cells. The Journal of Neuroscience. 28 (10), 2301 (2008).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Liu, Y., et al. Tamoxifen-independent recombination in the RIP-CreER mouse. PLoS One. 5 (10), e13533 (2010).
- Klingler, E., Jabaudon, D. Do progenitors play dice?. eLife. 9, e54042 (2020).
- Hippenmeyer, S. Dissection of gene function at clonal level using mosaic analysis with double markers. Frontiers in Biology. 8 (6), 557-568 (2013).
- Rulands, S., Simons, B. D. Tracing cellular dynamics in tissue development, maintenance and disease. Current Opinion in Cell Biology. 43, 38-45 (2016).
- Laukoter, S., et al. Imprinted Cdkn1c genomic locus cell-autonomously promotes cell survival in cerebral cortex development. Nature Communications. 11 (1), 195 (2020).
- Daigle, T. L., et al. A Suite of Transgenic Driver and Reporter Mouse Lines with Enhanced Brain-Cell-Type Targeting and Functionality. Cell. 174 (2), 465-480 (2018).
- He, M., et al. Strategies and Tools for Combinatorial Targeting of GABAergic Neurons in Mouse Cerebral Cortex. Neuron. 91 (6), 1228-1243 (2016).
- Yamamoto, M., et al. A multifunctional reporter mouse line for Cre- and FLP-dependent lineage analysis. Genesis. 47 (2), 107-114 (2009).
- Plummer, N. W., et al. Expanding the power of recombinase-based labeling to uncover cellular diversity. Development. 142 (24), 4385 (2015).
- Imayoshi, I., Ohtsuka, T., Metzger, D., Chambon, P., Kageyama, R. Temporal regulation of Cre recombinase activity in neural stem cells. Genesis. 44 (5), 233-238 (2006).
- Sasaki, S., et al. Complete loss of Ndel1 results in neuronal migration defects and early embryonic lethality. Molecular and Cellular Biology. 25 (17), 7812-7827 (2005).
- Acampora, D., et al. Epilepsy and brain abnormalities in mice lacking the Otx1 gene. Nature Genetics. 14 (2), 218-222 (1996).
- Soeda, T., et al. Sox9-expressing precursors are the cellular origin of the cruciate ligament of the knee joint and the limb tendons. Genesis. 48 (11), 635-644 (2010).
- Klezovitch, O., Fernandez, T. E., Tapscott, S. J., Vasioukhin, V. Loss of cell polarity causes severe brain dysplasia in Lgl1 knockout mice. Genes & Development. 18 (5), 559-571 (2004).
- Pimeisl, I. M., et al. Generation and characterization of a tamoxifen-inducible EomesCreER mouse line. Genesis. 51 (10), 725-733 (2013).
- Nakagawa, N., et al. Memo1-Mediated Tiling of Radial Glial Cells Facilitates Cerebral Cortical Development. Neuron. 103 (5), 836-852 (2019).
- Nowotschin, S., et al. The T-box transcription factor Eomesodermin is essential for AVE induction in the mouse embryo. Genes & Development. 27 (9), 997-1002 (2013).
- Balordi, F., Fishell, G. Mosaic removal of hedgehog signaling in the adult SVZ reveals that the residual wild-type stem cells have a limited capacity for self-renewal. Journal of Neuroscience. 27 (52), 14248-14259 (2007).
- Liang, H., et al. Neural development is dependent on the function of specificity protein 2 in cell cycle progression. Development. 140 (3), 552-561 (2013).
- Guo, C., Yang, W., Lobe, C. G. A Cre recombinase transgene with mosaic, widespread tamoxifen-inducible action. Genesis. 32 (1), 8-18 (2002).
- Ahn, S., Joyner, A. L. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature. 437 (7060), 894-897 (2005).
- Storm, R., et al. The bHLH transcription factor Olig3 marks the dorsal neuroepithelium of the hindbrain and is essential for the development of brainstem nuclei. Development. 136 (2), 295-305 (2009).
- Koundakjian, E. J., Appler, J. L., Goodrich, L. V. Auditory neurons make stereotyped wiring decisions before maturation of their targets. Journal of Neuroscience. 27 (51), 14078-14088 (2007).
- Dor, Y., Brown, J., Martinez, O. I., Melton, D. A. Adult pancreatic β-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 429 (6987), 41-46 (2004).
- Sohal, D. S., et al. Temporally regulated and tissue-specific gene manipulations in the adult and embryonic heart using a tamoxifen-inducible Cre protein. Circulation Research. 89 (1), 20-25 (2001).
- Ventura, A., et al. Restoration of p53 function leads to tumour regression in vivo. Nature. 445 (7128), 661-665 (2007).
- Johnson, L., et al. Somatic activation of the K-ras oncogene causes early onset lung cancer in mice. Nature. 410 (6832), 1111-1116 (2001).
- Tasic, B., et al. Extensions of MADM (mosaic analysis with double markers) in mice. PLoS One. 7 (3), e33332 (2012).
- Yu, J., Carroll, T. J., McMahon, A. P. Sonic hedgehog regulates proliferation and differentiation of mesenchymal cells in the mouse metanephric kidney. Development. 129 (22), 5301-5312 (2002).
- Zhao, H., et al. Role of fibroblast growth factor receptors 1 and 2 in the ureteric bud. Developmental Biology. 276 (2), 403-415 (2004).
- Schuchardt, A., D'Agati, V., Larsson-Blomberg, L., Costantini, F., Pachnis, V. Defects in the kidney and enteric nervous system of mice lacking the tyrosine kinase receptor Ret. Nature. 367 (6461), 380-383 (1994).
- Livet, J., et al. ETS gene Pea3 controls the central position and terminal arborization of specific motor neuron pools. Neuron. 35 (5), 877-892 (2002).
- Volz, K. S., et al. Pericytes are progenitors for coronary artery smooth muscle. eLife. 4, e10036 (2015).
- Cai, C. L., et al. A myocardial lineage derives from Tbx18 epicardial cells. Nature. 454 (7200), 104-108 (2008).
- Lasrado, R., et al. Lineage-dependent spatial and functional organization of the mammalian enteric nervous system. Science. 356 (6339), 722-726 (2017).
- Matsuoka, T., et al. Neural crest origins of the neck and shoulder. Nature. 436 (7049), 347-355 (2005).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati