
Method Article
Отслеживание линий и клональный анализ при разработке коры головного мозга с использованием мозаичного анализа с двойными маркерами (MADM)
В этой статье
Резюме
Представлен протокол для отслеживания линий и функционального генетического анализа генов-кандидатов на уровне одной клетки с использованием мозаичного анализа с двойными маркерами (MADM). Клональный анализ MADM обеспечивает количественную основу для измерения пролиферативного поведения, клеточного выхода и родовых отношений отдельных прародителей и их клеток дочери.
Аннотация
Начиная с ограниченного пула прародителей, кора головного мозга млекопитающих образует высокоорганизованные функциональные нейронные цепи. Тем не менее, основные клеточные и молекулярные механизмы, регулирующие линейные переходы нервных стволовых клеток (НСЦ) и возможное производство нейронов и глии в развивающемся нейроэпителии, остаются неясными. Методы отслеживания шаблонов деления NSC и картирования линии клонально связанных клеток значительно продвинулись вперед. Тем не менее, многие современные методы отслеживания линий страдают от отсутствия клеточного разрешения судьбы клеток потомства, что имеет важное значение для расшифровки моделей деления клеток-предшественников. Представлен протокол с использованием мозаичного анализа с двойными маркерами (MADM) для выполнения клонального анализа in vivo. MADM одновременно манипулирует отдельными клетками-предшественниками и визуализирует точные модели деления и прогрессии линии при беспрецедентном разрешении одной клетки. МАДМ основе межхромомономных рекомбинации событий во время G2-X фазы митоза, вместе с временно неуступчивой CreERT2, предоставить точную информацию о датах рождения клонов и их разделения моделей. Таким образом, отслеживание линии MADM обеспечивает беспрецедентные качественные и количественные оптические считывание режима пролиферации прародителей стволовых клеток на уровне одной клетки. MADM также позволяет изучить механизмы и функциональные требования генов-кандидатов в прогрессировании линии NSC. Этот метод уникален тем, что сравнительный анализ контроля и мутантных субклонов может быть выполнен в той же среде ткани in vivo. Здесь протокол подробно описан, и продемонстрированы экспериментальные парадигмы для использования MADM для клонального анализа и отслеживания линий в развивающейся коре головного мозга. Важно отметить, что этот протокол может быть адаптирован для выполнения клонального анализа MADM в любой нише стволовой клетки, до тех пор, пока присутствует драйвер CreERT2.
Введение
Кора головного мозга представляет собой высокоорганизованную структуру, состоящую из шести различных слоев. Кора содержит широкий спектр типов клеток, включая нейроны и глии, которые взаимодействуют, чтобы сформировать функциональные нейронные цепи. Большинство, если не все, корковые возбудители проекции нейронов и глии являются производными от общего пула нервных стволовых клеток (NSCs), известных как радиальные глиальные прародители (RGPs)1,2,3. RGPs сами являются производными от нейроэпителиальных стволовых клеток (NESCs), составляющих ранние эмбриональные нейроэпителий. К эмбриональному дню 9 (E9) у мышей, РЭЦ начинают переход в RGPs4. RGP прогрессирование линии требует точного височного и пространственного регулирования, и когда этот процесс препятствует, тяжелые неврологические расстройства, такие как мегаленцефалия, микроцефалия, лиссенцефалия, или нарушения, такие как шизофрения и аутизм может привести5,6. На E10, большинство RGPs проходят симметричное пролиферативное деление, в результате чего расширение нейронного пула прародителя4,7. RGPs в конечном итоге начинают делиться асимметрично, производя корковые нейроны проекции в временно определенным образом. Через последовательные волны нейрогенеза, новорожденные нейроны мигрируют в корковой пластины формирования корковых ламина с раннего рождения нейронов, занимающих глубокие слои и позднего рождениянейронов,проживающих в поверхностных слоях8,9,10. Потому что клонально связанных пирамидальных нейронов мигрируют радиально в кору с очень мало касательной дисперсии, дочь клетки, как правило, образуют колонку или конус формы структуры называют нейрональной радиальной единицы4,11,12,13. По E17, эмбриональное нейрогенное расширение завершено у мышей14. RGPs может также производить эпендимальные клетки и некоторые классы глии, в том числе астроцитов и олигодендроцитов1,15,16,17,18,19., Потенциал RGPs, чтобы привести к как нейроны и астроциты, как представляется, являются последовательными во всех корковых регионах18, примерно с 1/6 нейрогенных RGPs также производит glia11.
В настоящее время генетические и эпигенетические факторы, регулирующие височное прогрессирование стволовой клетки вдоль ее линии, в основном неизвестны. Временные модели экспрессии генов могут оказать существенное влияние на решения линий в RGPs20,,21,,22,,23,,24. Как это плотно вязать отношения между височной и пространственной узором приводит к молекулярному разнообразию взрослых нейронных типов через корковые области не известно. Аналогичным образом, как индивидуальный потенциал стволовых клеток и ее клеточный выход модулируется на клеточном и молекулярном уровне является важным вопросом без ответа. Будущие исследования, мы надеемся, адрес некоторые из этих вопросов, в конечном итоге расширение нашего понимания функциональных корковых формации цепи.
Развитие нейробиологии стремится понять связь линии, что клетки в головном мозге доля друг с другом. Первоначально, очень немногие инструменты исследования были имеющиеся для этого, и много предыдущих изучений полагались на визуально замечаниях картин разделения в прозрачных организмах such as Caenorhabditis elegans25. В последние десятилетия наблюдается резкое увеличение числа и сложности методов, доступных13,26,27,28,29. Появление системы редактирования генома CRISPR-Cas9 позволяет проводить синтетическую реконструкцию отношений линии клеток путем введения эволюционировавщих штрих-кодов ДНК27,,30. Два последних примера стратегии штрих-кодирования включают в себя использование самонаведения руководство РНК, которая направляет CRISPR-Cas9 к конкретным ДНК штрих-код локус или цитидин деаминазы сливается с никас Cas9 для целевой эндогенных перемежаются31,32. Эти технологии обеспечивают высоко мультиплексированные подходы путем введения штрих-кодов, которые постепенно и стабильно накапливают уникальные мутации с течением времени. Подходы к редактированию генома очень ценны, поскольку они позволяют ретроактивно анализировать взаимосвязь между любыми двумя клетками на основе общего наследования этих штрих-кодов. Однако, для того, чтобы читать штрих-коды в отдельных клетках, ткани, как правило, должны быть нарушены, и поэтому информация о положении, морфологии, и абсолютные цифры клеток от отдельного прародителя теряется.
Комбинированные парадигмы маркировки сохраняют пространственную информацию и в принципе также позволяют проводить различие между тесно локализованными или даже перекрывающимися клонами33,,34. Для метода отслеживания линии, чтобы быть информативным он должен маркировать отдельных прародителей и их потомство в редких и неизгладимый образом. Примечательно, что Brainbow35 и Confetti36,37 подходов использовать стохастический многоцветный Cre рекомбиназы основе репортеров, которые выражают сочетание флуоресцентных белков из одного локуса. Обширное количество одновременных цветовых комбинаций, которые могут быть достигнуты in vivo сделать это мощный инструмент при отслеживании корковых клонов RGP и астроцитов34. Системы на основе Транспосона, обеспечивающие стабильную геномную интеграцию трансгенов, кодирующих флуоресцентные репортеры и позволяющие отслеживать линии корковых прародителей, также были разработаны33,,38,,39,,40,41. Системы на основе Транспосона имеют дополнительное преимущество в том, что репортер конструкции стабильно интегрироваться в геном, и тем самым надежно этикетки линейно родственных клеток дочери. Для отслеживания линий астроцитов в частности, был разработан ряд методов, которые включают электропорации piggyBac транспозазы, включая Star Track, который использует комбинацию конструкций кодирования различных флуоресцентных белков40,42. Другой подход, МАРКЕРы MAGIC,вводит векторы Brainbow как транспонируемые трансгены. Это было успешно использовано для отслеживания эмбриональных нейронных и астроцитов прародителей34,43. Недавно, мозаичный анализ двойной рекомбинациозно-опосредованной кассеты обмена (MADR) было установлено, чтобы стабильно этикетки мутантных клеток, выражающих трансгенные элементы из точно определенных хромосомных локусов44. Эти мощные в vivo комбинаторные методы маркировки предоставили многочисленные идеи в динамику линии клеток-предшественников. Тем не менее, эти анализы выполняются на фиксированной ткани, обеспечивая снимок отдельных клонов на определенном этапе развития. Для того, чтобы наблюдать изменения в динамике линии одиночных клонов с течением времени, хронические методы визуализации in vivo, аналогичные тем, которые выполняются во взрослой зубной извилине, должны быть применены45.
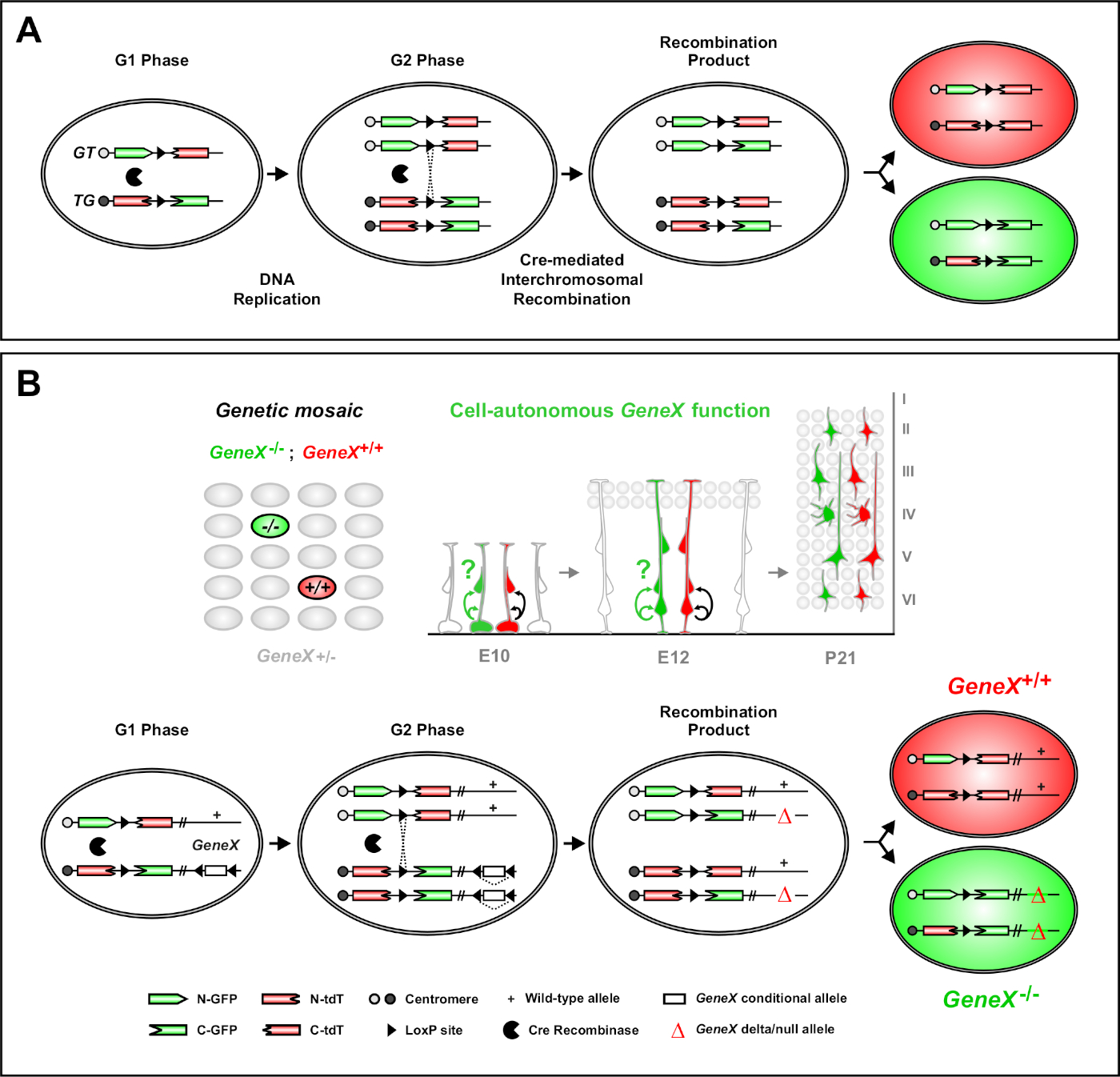
Мозаичный анализ с двойными маркерами (MADM) является мощным методом двойной окраски маркировки, который позволяет in vivo отслеживания линии отдельных клеток-предшественников у мышей46,47. Для возникновения событий маркировки MADM необходимы два компонента: во-первых, кассеты MADM должны быть ориентированы на идентичные локусы на гомологичные хромосомы. Кассеты состоят из двух химерных флуоресцентных генов репортера, eGFP (зеленый, «G») и тандем dimer Tomato (красный, tdT'T). Кассета GT содержит N-термин eGFP и C-термин tdT, разделенный интроном, содержащим локсП сайт. Кассета TG построена обратно, с N-термином tdT и C-термином eGFP. Во-вторых, выражение рекомбинации Cre в той же клетке, содержащей целевые кассеты MADM имеет важное значение. В отсутствие Cre,химерные кассеты не выражают функциональные eGFP или tdT, потому что их кодировки последовательности нарушаются. Сайты loxP служат мишенью для кре-опосредованной межхромомной рекомбинации, что приводит к восстановлению обеих выразительных кассет одновременно. Если рекомбинация происходит во время фазы G2 клеточного цикла с последующей сегрегацией X (G2-X), то две клетки дочери будут выражать один из двух флуоресцентных белков. Временное регулирование активности CreERT2 с использованием тамоксифена (ТМ) предоставляет точную информацию о дате рождения клонов MADM и моделях деления их потомства(Рисунок 1A)29,46,47.
MADM может потенциально систематически маркировать отдельных клонов с высоким разрешением одной клетки в мозге мыши похож на традиционные, но неспецифические и трудоемкие методы, как Golgi окрашивания48 или красителя заполнения49. Потому что только промоутер вождения CreERT2 определяет тип клеток специфичность клональной маркировки MADM, MADM в принципе может быть применен для отслеживания клональной линии всей любой орган и ткань47,50,51,52. Действительно, исследования уже использовали MADM выявить отношения линии в клонах, полученных из различных тканей47,50,51,52,53,54,55,56,57,58,59.50 Экспериментальные парадигмы MADM были применены для изучения линии в корковых проекционных нейронах, глии и постнатальных стволовых клетках в развивающихся неокортексах7,,11,,12,,46,,60,,61,,62,,63,,64,,65. MADM также был использован для изучения линии клеток во взрослом зубчатой извилине, таламусе, мозжечковых гранулах и интернейронах на клональном уровне (см. таблицу 1 для полного списка)47,53,54,56,57,66.
Уникальной особенностью MADM является способность генетически связать мутации дистальной к одной кассете MADM, создавая тем самым генетическую мозаику(рисунок 1B и рисунок 2). Это приводит к диким клеткам дочери типа помечены одним флуоресцентным маркером (tdT на рисунке 1B) и гомозиготных мутантов братьев и сестер с другими (eGFP на рисунке 1B) в немаркированной гетрозиготной среде. MADM уникален тем, что сравнительный анализ контроля и мутантных субклионов может быть выполнен в той же среде ткани in vivo. Первоначально кассеты MADM были нацелены на Locus47 Rosa26, но анализ функции гена MADM ограничивался генами, дисталлированными к локусу. Чтобы преодолеть (по крайней мере частично) это ограничение и расширить возможности для АНАЛИЗА генов на основе MADM, кассеты MADM были сбиты вблизи центрометров Chr. 751, Chr. 1146, и Chr. 1251. Ориентация на все 19 автосом мыши с кассетами MADM продолжается и позволит практически любой ген, который будет изучен в будущем, обеспечивая беспрецедентную платформу для изучения отношений линии развития в сочетании с функциональным генетическим анализом.
протокол
Протоколы мыши были рассмотрены институциональным доклиническим основным предприятием (PCF) и внутренним этическим комитетом в IST Austria. Все разведения и эксперименты проводились по лицензии, одобренной Австрийским федеральным министерством науки и исследований в соответствии с законодательством Австрии и ЕС о животных.
1. Разведение экспериментальных мышей для клонального анализа MADM
- Настройка приурочен экспериментальных MADM спаривания (Nogt;P56; CD-1) во второй половине дня (5:00 вечера) и проверить на вагинальные пробки продолжения утром (8:00 утра). Утром штепсельная вилка присутствует рассчитывает как день 0.5. Смотрите рисунок 2 для обзора экспериментальной установки спаривания мыши. Убедитесь, что временные точки для индукции TM деятельности и анализа CreERT2 подходят для решения экспериментальных вопросов.
ПРИМЕЧАНИЕ: Для получения дополнительной информации относятся к рисунку 3 и репрезентативные результаты ниже. - Для послеродовой выборки, создать разведения для создания приемных матерей параллельно.
ПРИМЕЧАНИЕ: Они должны быть начаты до 1-2 дней до создания экспериментальных разведения.
2. TM индукции в MADM мышей
- Приготовьте рабочий раствор 20 мг/mL TM, растворив его в кукурузном масле в конической трубке конической центрифуги 15 мл или 50 мл и поместив ее на платформу качания на 4 ч при комнатной температуре (RT), гарантируя, что ТМ полностью растворяется. Храните рабочее решение при 4 градусов по Цельсию, покрытое алюминиевой фольгой и используйте в течение 2 недель.
- Чтобы вызвать события рекомбинации MADM, доставить одну инъекцию ТМ интраперитонально (IP) с помощью 1 mL tuberculin шприц и 25 G иглы в приурочен беременной плотины. В зависимости от стадии кортикального нейрогенеза, вводить ТМ между Е10-Е15 в дозе 1-2 мг/беременной плотины. Для ранних точек времени (т.е. E10) использовать максимум 1 мг / беременных плотины (25 мг/кг) для предотвращения осложнений во время беременности11. Для временных точек между E11-E15 использовать 2 мг / беременных плотины (50 мг/кг)7.
ПРИМЕЧАНИЕ: Кроме того, TM может быть введен с устной gavage для поздней беременности. - Для клонального анализа MADM до постнатальных временных точек, восстановить живые эмбрионы на E18'E19 через кесарево сечение, а затем растить щенков с приемной матерью.
ПРИМЕЧАНИЕ: В зависимости от состояния здоровья беременной женщины, это не может быть необходимо для выполнения кесарева сечения, но повышение щенков с приемной матерью по-прежнему требуется, потому что оригинальный TM-лечение матери могут возникнуть проблемы с кормления. - Чтобы восстановить живые эмбрионы путем кесарева сечения или получить эмбриональные временные точки для анализа, пожертвовать беременной плотины путем вывиха шейки матки.
- Поместите животное в положение на спине и дезинфицировать мех с 70% этанола. Сделайте небольшой разрез в коже в нижней части живота над маткой, используя хирургические щипцы и ножницы. Сделайте второй разрез через мышцы и брюшную мышечную стенку, чтобы выявить брюшную полость.
- Удалите матку, отделив от окружающих тканей ножницами. Передача нетронутой матки на перчатку, наполненную теплой водой, чтобы увеличить выживаемость эмбриона, пока каждый из них удаляется из амниона индивидуально.
- Используйте тонкие наконечником ножницы и пальцы, чтобы тщательно открыть стенки матки, чтобы освободить эмбрионов. Не перерезайте пуповины слишком близко к телу, чтобы предотвратить обширную кровопотерю. Если эмбрионы должны быть использованы для анализа, переходите к шагу 3.9. Если щенки должны быть воспитаны, перейти к шагу 2.8.
- Если требуется воспитание, очистите щенков, прежде чем передать их приемной матери. Во время чистки щенков, осторожно нажмите на грудь время от времени, чтобы начать дыхание. Поместите обратно на вторую перчатку, наполненную теплой водой, чтобы улучшить выживаемость.
ПРИМЕЧАНИЕ: Важно аккуратно удалить все оставшиеся амнион и / или плаценты с бумажным полотенцем. - Перед передачей щенков приемной матери, удалить приемную мать из клетки, удалить оригинальные щенки, и заменить экспериментальных щенков. Верните приемную мать в клетку.
ПРИМЕЧАНИЕ: Смотрите обсуждение дополнительных предложений по улучшению повышения уровня приема. - Если требуется генотипирование, собирайте биопсию ног или хвоста между P6-P8.
ПРИМЕЧАНИЕ: Выполните этот шаг только в том случае, если экспериментальные лицензии на животных одобрят эту практику.
3. Ткань подготовки для клонов MADM в головном мозге
ПРИМЕЧАНИЕ: Для экспериментов, которые включают в себя послеродовую ткань (P4), перейти к шагу 3.1. Для эмбриональных временных точек и ранних послеродовых (P0-P3), продолжают шаг 3.9.
- Обезболивать экспериментальное животное MADM с инъекцией IP кетамина/ксилазина/ацепромазина (65 мг, 13 мг и 2 мг/кг массы тела, соответственно) и подтверждайте, что мышь не реагирует, щипая заднюю лапу.
ПРИМЕЧАНИЕ: Как мужчины, так и женщины MADM мышей (CD-1 фон) используются для анализа. Если требуется генотипирование, собирайте биопсию уха в этой точке. - Поместите обезболивающее животное в положение на спине на подносе перфузии и дезинфицировать мех 70% этанолом. Чтобы начать операцию, тщательно сделать разрез с ножницами и хирургическими щипцами через внешний слой кожи, а затем второй разрез через мышечный слой. Поднимите кончик грудины и вырежьте соединительной ткани по бокам, принимая дополнительную осторожность, чтобы избежать резки печени. Грудная полость будет видна.
- Snip диафрагмы и поднять, чтобы раскрыть сердце. Тщательно обрезать грудную клетку и булавку к хирургическому подносу, чтобы разоблачить сердце. Для щенков, удалить грудную клетку полностью.
- Вставьте иглу с фосфатом буферизированного солевого раствора (PBS) в нижний левый желудочек (бледная ткань). Используя небольшие ножницы радужной оболочки сделать разрез на задний конец правого предсердия (темно-красная ткань) для крови, чтобы слить.
- Выполните перфузию с PBS следуют сразу же свежеприготовленные, ледяной 4% параформальдегид (PFA), приготовленные в PBS. Для щенков (P4-P10) используйте шприцы для выполнения перфузии. Заполните один шприц 10 мл PBS и другой с 10 мл 4% PFA. Убедитесь, что все пузырьки воздуха в шприцах были удалены. Для пожилых животных, используйте перистальтический насос.
- Начните перфузию с PBS (10 мл при 2-4 м/мин у щенков; 20 мл на 4-6 мл/мин для взрослых с помощью перистальтического насоса). Печень станет ясной и бледно-желтой, если игла расположена правильно.
- После завершения, удалить иглу из щенков и вставить иглу, содержащую PFA в том же отверстие. Для взрослых, остановить перистальтический насос перед обменом решения PBS с ледяной PFA, убедившись, чтобы избежать пузырьков в поглощении трубки. Возобновление перфузии с PFA (10 мл при 2-4 м/мин у щенков; 30 мЛ на 4-6 мл/мин для взрослых с помощью перистальтического насоса).
- Когда перфузия завершена, обезглавить мышь и удалить мозг через тщательное вскрытие. Передача мозга на 4% PFA. Используйте по крайней мере 5x объемы мозга (т.е. 5-10 мЛ PFA в конической трубке центрифуги 15 мЛ) и инкубировать на ночь при 4 градусах Цельсия для постперфузионной фиксации, чтобы обеспечить полную фиксацию ткани. Продолжайте шаг 3.10.
- Для эмбриональной ткани и ранней послеродовой ткани (т.е. P0-P3), после выполнения кесарева сечения, обезглавить эмбрионы ножницами. Если требуется генотипирование, соберите хвост эмбриона в этой точке. Немедленно вскрыть мозг и передать в 12 хорошо пластины, содержащие 2'3 мL 4% PFA/ well. Инкубировать на ночь при 4 градусов по Цельсию для постфиксации.
- На следующее утро обмен PFA с 10 мл (взрослый) или 2'3 mL (эмбрион) PBS и повторить мыть 3x в течение 15 мин на RT. Передача ткани до 30% раствора сахарозы в фосфат буфер (PB) и хранить при 4 градусов по Цельсию на платформе качания, пока ткань не погружается в раствор.
- Встраивание мозга в оптимальную температуру резки (OCT) соединение в встраиваемой формы, заботясь, чтобы сориентировать мозг для короналовых или сагиттальных секций. Заморозить, разместив встраиваемую форму на сухой лед, пока OCT не станет полностью непрозрачным (от 10 до 15 мин). Храните ткани при -80 градусов по Цельсию до дальнейшего использования.
4. Приготовление ткани МАДМ для иммуногистохимии
- Прикрепите блок ткани к диску образца в криостате, применив кольцо OCT к диску и поместив блок прямо в OCT, когда он начинает замерзать. Убедитесь, что блок правильно ориентирован на нужную плоскость резки.
ПРИМЕЧАНИЕ: Здесь, корональный раздел для исследования корковых клонов MADM описывается подробно. - Установите температуру блока в криостате до -20 градусов по Цельсию, а температура лезвия до -21 градусов по Цельсию.
- Разрешить блок тканей, чтобы приспособиться к температуре камеры путем монтажа образца диска на держатель образца и оставить в криостате в течение 5 минут до начала сечения.
- Обрезать блок толстыми секциями (45–60 мкм) до тех пор, пока не будет достигнута интересующая ткань.
- Как только край коры хорошо виден, прекратите секции и заблокируйте лезвие. Убедитесь, что лезвие защищено перед обрезкой блока.
- Обрезать избыток OCT окружающих ткани с лезвием, оставляя 1-2 мм OCT со всех сторон мозга.
- Далее ориентируйте блок так, чтобы один из боковых краев коры ориентировался вниз, а другой вверх (т.е. наиболее ростальный край коры указывается вправо).
- Начните секцию толщиной 45 мкм для взрослых клонов и 30 мкм для эмбриональных клонов. Выполните каждый раздел индивидуально и использовать небольшую кисть, чтобы держать область ниже ножа чистой любого мусора слева при обрезке блока.
ПРИМЕЧАНИЕ: Если это не сделано и раздел падает, это может быть трудно определить правильный порядок срезов. - Если секции начинают скручиваться, обрезать края блока и / или тщательно настроить стеклянную пластину антиролла.
- Для анализа эмбрионального клона, смонтировать разделы непосредственно на матовый слайд. Высушите на отопительной пластине при 37 градусов по Цельсию, прежде чем приступить непосредственно к шагу 5.6.
ПРИМЕЧАНИЕ: Несколько разделов могут быть добавлены к одному слайду, но убедитесь, что последовательный порядок поддерживается. - Для сбора клонов для взрослых, подготовить 24 хорошо пластин, содержащих 1 мл PBS / хорошо (как правило, 5-6 пластин на мозг). Начиная с первой скважины, с холодными щипцами собирают отдельные серийные секции в PBS в порядке секций.
ПРИМЕЧАНИЕ: Метод плавающей секции принят для взрослых тканей, чтобы гарантировать, что никакие разделы не пропущены и что установленные разделы не содержат морщин. - Прекратите секцию после того, как будет достигнут конец неокортекса.
- Для взрослых клонов, приступить к монтажу плавающих секций.
ПРИМЕЧАНИЕ: Разделы могут храниться в PBS при 4 градусах Цельсия до 24 ч.
5. Установка ткани взрослого для визуализации
ПРИМЕЧАНИЕ: Необходимы следующие инструменты: небольшая кисть краски, чашка Петри, PBS с 0,5% Tween (PBS-T), слайдыслипа (Таблица материалов),монтажная среда(Таблица материалов),крышки и щипцы.
- Заполните чашку Петри PBS-T.
ПРИМЕЧАНИЕ: Двимт используется для оказания помощи в процессе монтажа. Если необходимо окрашивание дополнительных антигенов, чувствительных к моющим средствам (т.е. гликопротеинам), то лучше пропустить добавление Tween. - Поместите слайд склеиния в PBS-T так, чтобы он был почти прикрыт до этикетки.
- Перенести первый раздел в PBS-T.
- Используя небольшую кисть краски, маневрируйте секцию на горку и организуйте ее, чтобы сохранить порядок резки. Продолжить таким же образом со всеми другими разделами.
- После того, как все разделы находятся в положении, поместите слайд (разделы/слайды) в темную слайд-камеру. Слегка поднимите крышку, чтобы разделы полностью высохли (10–20 мин), гарантируя, что они остаются прилипшими в последующих шагах.
- При выполнении иммуногистохимии для дополнительных антигенов, переходить непосредственно к разделу 6 или 7.
ПРИМЕЧАНИЕ: Для эмбриональных временных факторов необходимо выполнять иммуностиминговые шаги по крайней мере для GFP и tdT (раздел 6). Для взрослых клонов, это необходимо только, если окрашивание для дополнительных антигенов параллельно (разделы 6 и 7). - Регидратировать и мыть разделы 1x с 1x PBS в течение 5 минут, чтобы удалить остаточные PBS-T. Нанесите на слайд 1 мл 4,6-диамидино-2-фенилиндола (DAPI), разбавленного в 1x PBS (1 мкг/мл), гарантируя, что все секции покрыты и инкубируются в течение 15 минут.
- Тщательно удалите DAPI и вымойте 1x PBS в течение 5 минут. Удалите излишки PBS и высушите в течение 1-2 минут, прежде чем встраивать в 110 мл монтажной среды. Печать с 24 х 60 мм крышки и дайте высохнуть, по крайней мере 3 ч до изображения.
6. Иммуносвейтинг только для GFP и tdT
ПРИМЕЧАНИЕ: Этот раздел необходим для эмбриональных клонов.
- Поместите слайды горизонтально в увлажненную камеру инкубации слайда. Отметьте границы слайдов с восковым маркером, чтобы свести к минимуму требуемое количество буфера.
- Регидратировать секции с 1x PBS. Для улучшения качества окрашивания работайте со свежесрезанной тканью.
- Добавьте 250-400 мл блокирующего буфера (0,5% Triton X-100, 2-3% нормальную сыворотку осла в 1x PBS) на слайд, гарантируя, что все разделы покрыты. Инкубировать за 1 ч.
ПРИМЕЧАНИЕ: Концентрация моющего средства (Triton X-100 или Tween-20) будет варьироваться в зависимости от дополнительных первичных антител, используемых, потому что некоторые антигены более чувствительны к моющим средствам, чем другие. - Удалите блокирующий буфер и добавьте первичные антитела в блокирующий буфер к слайду (300–400 мл/слайд).
ПРИМЕЧАНИЕ: Пример стандартной первичной реакции антител для анти-GFP/anti-tdT (MADM) может использовать куриные анти-GFP (1:500) и кролика анти-RFP (1:500). - Инкубировать первичными антителами на ночь при 4 градусах Цельсия.
ПРИМЕЧАНИЕ: Слайды должны быть точно инкубированы идеально горизонтально с буфером, охватывающим все разделы. В противном случае, неравномерное или плохое окрашивание может привести. - Подтвердите на следующее утро, что блокирующий буфер с первичными антителами по-прежнему охватывает все разделы на слайде. Если нет, повторите инкубационный шаг на 3х4 ч на RT.
- Удалить первичные антитела и мыть 4x с 1x PBS в течение 10 минут на RT.
- Добавьте вторичные антитела, разбавленные в блокирующий буфер, чтобы скользить (300–400 мл/слайд): Alexa Fluor 488 anti-chicken IgG (1:500) и Cy3 anti-rabbit IgG (1:500).
- Инкубировать на RT в течение 2 ч. Держите слайды покрыты от света, чтобы предотвратить отбеливание фторфоров.
- Удалите вторичные антитела и вымойте 2x с 1x PBS в течение 10 мин.
- Инкубировать DAPI разбавленным в PBS (1:5,000) в течение 15 мин.
- Вымойте разделы 1x с 1x PBS в течение 10 мин.
- Удалите избыток PBS и высушите в течение 1–2 минут, прежде чем встраивать в 110 мл монтажной среды.
- Печать с 24 х 60 мм крышки и дайте высохнуть, по крайней мере 3 ч до изображения. Изображение скользит в течение 1-2 недель после выполнения иммуногистохимии для обеспечения оптимального сигнала.
7. Иммуносхейтинг для GFP, tdT и дополнительных антигенов
- Выполняйте шаги 6.1–6.3.
- Удалите блокирующий буфер и добавьте первичные антитела в блокирующий буфер к слайду (300–400 мл/слайд).
ПРИМЕЧАНИЕ: При окрашивании трех или более антигенов (т.е. GFP, tdT и интересующего белка) и антитела к интересующему белку были подняты у кролика, рекомендуется использовать антитtт (козу) первичного антитела при разбавлении 1:500. Пример первичной реакции антител для 3 антигенов с альтернативным окрашиванием tdT смог использовать цыпленок anti-GFP (1:500), anti-tdT козы (1.500), и антитела против протеина интереса (т.е., кролика). - Выполните шаги 6.5–6.7.
- Добавьте вторичную смесь антител, разбавленную в блокирующий буфер, чтобы сползти (300–400 мл/слайд): Alexa Fluor 488 anti-chicken IgG (1:500), Cy3 anti-goat IgG (1:500) и Alexa Fluor 647 anti-rabbit IgG (1:500).
- Выполните шаги 6.9–6.14.
8. Confocal приобретение изображений и количественная оценка клонов MADM
- Определите и документируйте разделы мозга, содержащие клоны и их расположение в коре головного мозга.
ПРИМЕЧАНИЕ: Количество секций, которые охватывает клон, будет варьироваться в зависимости от того, когда был индуцирован клон, драйвер CreERT2 и время анализа. Этот шаг может быть выполнен либо на конфокальном микроскопе или микроскопе эпифлюоресценции. - Используя перевернутый конфокальный микроскоп, начните с выбора и настройки правильных лазерных линий и фильтров. Для мозгов MADM выберите DAPI, GFP и tdT (возбуждение: 358 нм, 488 нм и 554 нм соответственно; пиковый уровень выброса: 461 нм, 507 нм и 581 нм соответственно). Убедитесь, что пинхол установлен на 1 воздушный блок для оптимального качества изображения.
- Для конфокальных специфических настроек, клоны изображений с 20x объективным и 1x зумом. Для изображений, которые будут использоваться в количественной оценке, используйте значение пикселя скорости сканирования в размере 1,52–2,06 с (значения 7–8 в программном обеспечении для приобретения изображений) без усреднения. Отрегулируйте интенсивность лазера и получите настройки для каждого канала по мере необходимости.
ПРИМЕЧАНИЕ: В зависимости от требуемого качества изображения, настройки скорости сканирования и усреднения могут варьироваться. - После того, как клон четко определен, организовать визуализации плитки, чтобы покрыть все соответствующие разделы в клоне. Отрегулируйте z-стек так, чтобы все ячейки, обозначенные MADM в клоне, захватывались с интервалом 1,5 мкм/z-стек. Отрегулируйте черепичную область так, чтобы вся ширина коры захватилась при визуализации клона (т.е. от пиальной поверхности до мозоли корпуса).
- Изображение отдельных клонов, охватывающих несколько разделов последовательно, гарантируя, что любые разделы без клеток в клоне по-прежнему изображены с целью 3D-реконструкции и правильной интерпретации пространственной информации клеток.
- Проанализируйте каждый раздел, содержащий клетки клона MADM последовательно от рострала до хвостового конца коры. Различать отдельные нейроны и глии на основе их морфологии и / или маркер окрашивания. Запись позиционной информации параллельно на основе соответствующих границ слоя, определенных ядерным окрашиванием (DAPI).
ПРИМЕЧАНИЕ: Смотрите рисунок 4 для репрезентативных результатов для эмбрионального анализа и рисунок 5 для репрезентативных результатов для анализа взрослых.
9. Серийная 3D реконструкция клонов
ПРИМЕЧАНИЕ: 3D-реконструкция отдельных клонов, изображенных на серийных участках мозга, полезна для визуального отображения, а также для анализа 3D клональных архитектур и может выполняться в соответствии со следующими шагами.
- Стежок и предохранитель confocal черепичные изображения на основе параметров приобретения с использованием программного обеспечения для приобретения изображений. Откройте файл .czi, а затем запустите метод сшивания под вкладкой Обработки в программном обеспечении ЗЕН (Зейсс).
- Экспорт сшитые стеки изображений в виде индивидуальных z-самолетов в формате TIFF. Откройте сшитый файл .czi, а затем запустите метод экспорта изображений под вкладкой Обработки. Для многоканальных изображений экспортируем как красные/зеленые/синие изображения для последующей обработки изображений.
- Повторите шаги 9.1 и 9.2 для каждого серийного раздела мозга клона.
ПРИМЕЧАНИЕ: Для точной 3D-реконструкции, все участки мозга в клоне, в том числе те, без помеченных клеток, также должны быть обработаны. - Concatenate отдельных изображений в один стек для того, начиная от самых ростральных до самых caudal z-плоскост с использованием программного обеспечения для обработки изображений с открытым исходным кодом, таких как ImageJ / Фиджи67,68.
ПРИМЕЧАНИЕ: Любые пустые изображения по краям каждого раздела мозга должны быть удалены в этой точке. - При необходимости исправьте стек изображения, полученный с шага 9.4 для несогласованности с помощью плагина ImageJ под названием"MultiStackReg",следуя шагам 9.5.1'9.5. Если выравнивание изображения не требуется, перейдите к шагу 9.6.
ПРИМЕЧАНИЕ: Этот плагин выполняет выравнивание изображения канала с наивысшим контрастом (обычно DAPI), а затем применяет записанное преобразование к другим каналам, тем самым позволяя надежное выравнивание изображения многоканальных стеков. Вспомогательный плагин под названием"TurboReg"должен быть предустановлен.- В ImageJ установите плагины"MultiStackReg"иTurboReg.
- Откройте стопку изображений изображений, полученных от шага 9.4, чтобы выровняться. Разделите каналы на DAPI (синий), GFP (зеленый) и tdT (красный) в варианте Color под вкладкой Изображения.
- Выполнить "MultiStackReg" плагин, чтобы выровнять канал DAPI по "Rigid Body" преобразование и сохранить файл преобразования.
- Примените сохраненный файл преобразования к двум другим каналам, используя «MultiStackReg».
- Слияние всех трех выровненных каналов и сохранить выровненный стек.
- Чтобы сориентировать клон в ImageJ, поверните стопку изображений клонов, полученных из шага 9.4 (или шаг 9.5.5 после выравнивания) в вертикальной ориентации с pial поверхности сверху и корпусом мозоли в нижней части. Урожай в плоскости Кси, если это необходимо.
- Для качественной презентации и количественного анализа создайте максимальное изображение z-проекции (шаг 9.8) или выполните 3D-рендеринг (шаг 9.9) клона.
- В ImageJ откройте стек изображения с шага 9.6 и выберите опцию «проекция» с интенсивностью «макс» типа проекции. Это позволит создать изображение всего клона, проецируемого на ту же плоскость.
- В ImageJ откройте стек изображения с шага 9.6 и выберите 3D-проект z-функции для создания 3D визуализации клона, который может быть повернут.
ПРИМЕЧАНИЕ: На этом этапе важно ввести правильный интервал среза, эквивалентный толщине отдельных z-стеков во время приобретения изображения. Инструмент интерполяции следует использовать для удаления зазоров между ломтиками.
Результаты
MADM приводит к восстановлению функциональных зеленых и красных флуоресцентных белков с двумя клетками дочери, каждая из которых выражает один из двух флуоресцентных белков на G2-X хромосомы сегрегации событий (Рисунок 1). Поскольку события MADM приводят к постоянной и четкой маркировке двух потомков линий, может быть выполнена поддающаяся количественной оценке зеленых и красных линий клеток дочери (субклионов). Можно определить переменные, включая шаблон деления (например, симметричную и асимметричную) и потенциал (например, количество потомства) первоначального прародителя. Количественная оценка каждого флуоресцентно помеченного подклона является информативной при ретроактивном определении того, проходит ли оригинальная клетка-предшественник симметричные пролиферативные деления, или асимметричные нейрогенные деления во время индукции ТМ. Предыдущие исследования сгруппированы Emx1-CreERT2 или Nestin-CreERT2 производные возбуждающие клоны проекции в коре головного мозга в два широких классов7,11,46. Первый, называется "симметричными пролиферативными клонами", состоит в среднем из значительного числа нейронов, с зелеными и красными субклонами, содержащими четыре или более нейронов каждый. Вторая группа, "асимметричные клоны" определяет класс клонов, где "меньшинство" субклон содержит менее трех нейронов и "большинство" подклон, четыре или более11. Эти определения специфичны для корковых RGPs и, возможно, потребуется пересмотреть для других областей мозга и тканей. Для обоих классов корковых клонов потомство будет распределено по поверхностным и глубоким слоям.
При разработке клональных исследований MADM необходимо учитывать ряд аспектов. Время, когда события MADM индуцируются администрацией ТМ, является ключевым соображением(рисунок 3). Для корковых возбудимых проекционных нейронов MADM клонов (т.е. с помощью Emx1-CreERT2 или Nestin-CreERT2) на E10, почти все RGPs все еще проходят симметричные деления11. Таким образом, индукция на E10 с TM захватили несколько раундов пролиферативной RGP усиления и привело к клонов с высоким числом нейронов. Тем не менее, количество RGPs на E10, как правило, невелики и, таким образом, администрация ТМ генерируется очень мало событий MADM (иногда меньше, чем один на мозг). Большинство RGP перешли от симметричных к асимметричным нейрогенных делений вокруг E12. Для целевой строго асимметричных нейрогенных клонов, было лучше, чтобы вызвать на E12 или позже (Рисунок 3). Время между индукцией ТМ и наблюдением событий рекомбинации MADM в коре головного мозга, как правило, составляет менее 24 ч. Инъекции IP были предпочтительным методом для администрирования ТМ на эмбриональных стадиях для этого метода, поскольку это привело к большей воспроизводимости в клональной индукции. Важно также, чтобы сохранить дозу ТМ до минимума по двум причинам. Во-первых, если скорость рекомбинации MADM увеличивается, вероятность индуцирования нескольких, возможно перекрывающихся клонов выше. Во-вторых, если будет доставлено слишком много ТМ, может наблюдаться повышенный уровень абортов, реабсорбции эмбрионов и меньших размеров помета. Аборты примерно в половине всех беременных плотин наблюдались, когда ТМ инъекции были доставлены на E10. Эта частота снизилась с Е11 и уменьшилась примерно до 1/3 беременных плотин, прерывающих беременность. Для резюме доз ТМ, индукции раз, и CreERT2 драйверы, используемые в предыдущих исследованиях MADM, относятся к таблице 1. Репортер деятельности в отсутствие ТМ наблюдался с некоторыми TM-индуцируемых CreERT2 драйверов69. Эктопическое выражение или события рекомбинации MADM в отсутствие ТМ не наблюдалось с Помощью Emx1-CreERT2 драйверов Nestin-CreERT2. Это может быть частично связано с тем, что TM-опосредованные хромосомные транс-рекомбинации происходят примерно на 1:1,000 до 1:10,000 более низкой частоты, чем цис-рекомбинации, уменьшая вероятность эктопической маркировки MADM.
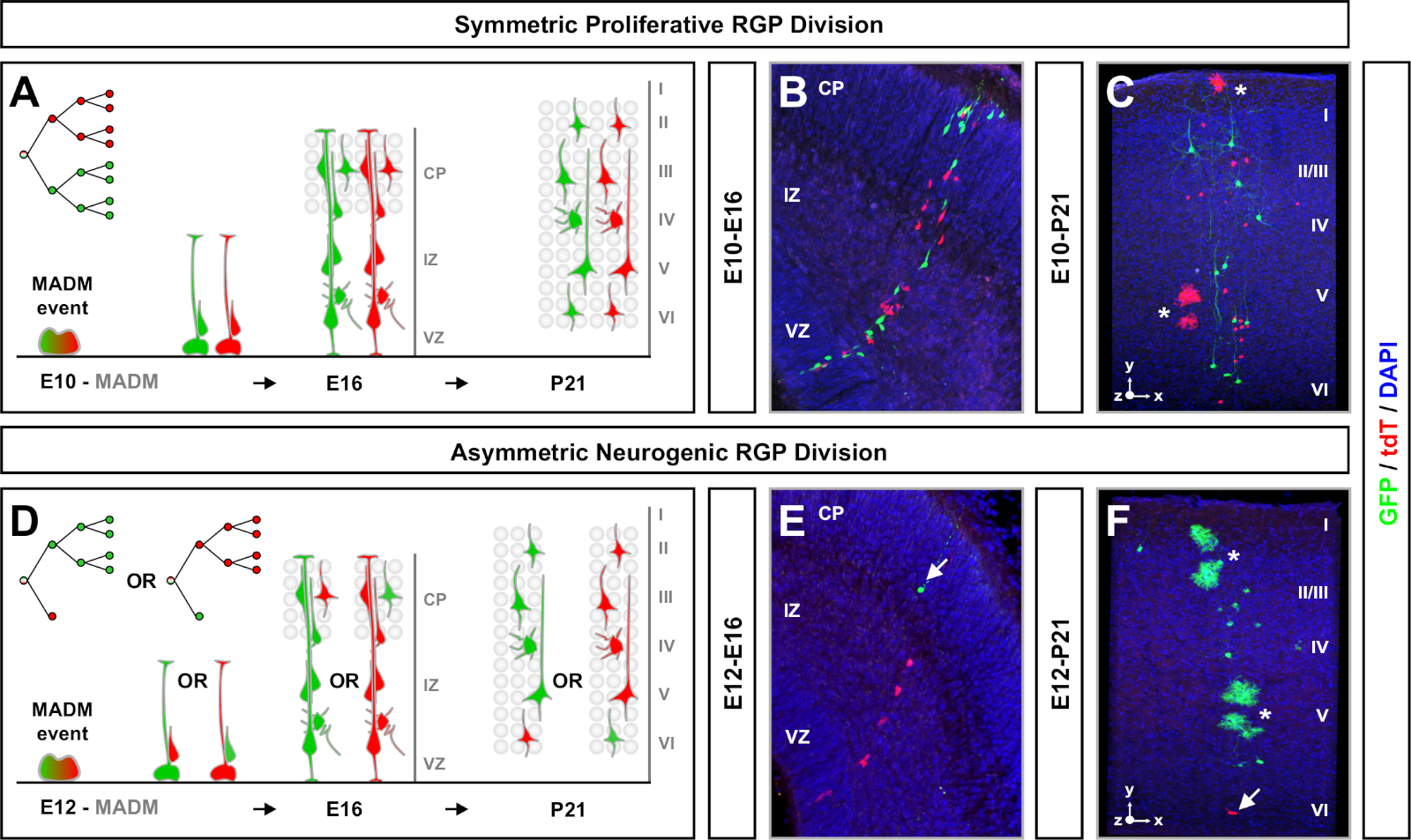
Еще одним фактором, который следует учитывать при планировании эксперимента по клональному анализу MADM, является продолжительность исследования. Изменение продолжительности времени между индукцией ТМ и когда эксперимент был проанализирован (A) (временное окно) отображает динамику стволовых клеток с течениемвремени 64. Короткие эмбриональные временные окна (т.е. ТМ/Е11-А/Е13; ТМ/Е11-А/Е16) запечатлел динамику эмбрионального нейрогенеза(рисунок 4). Сравнение клонов из двух или более временных окон дает количественное представление о количестве производимых клеток и о том, как распределение нейронов меняется на разных стадиях прогрессирования линии64. Чтобы зафиксировать весь потенциал отдельных клонов, необходимо продлить время, анализируемое в постнатальные или взрослые временные точки7,,11,,12. Примеры неокортикальных клонов, индуцированных в эмбрионе и проанализированных у взрослых, показаны на рисунке 5. Следует отметить, что кортикальный нейрогенез в основном завершен и глиогенез увеличивается на Е17. Приблизительно 1/6 нейрогенных RGP также приступить к генерации астроцитов и / или олигодендроцитов11.
Симметрические клоны возникают, когда RGPs проходят один или несколько раундов пролиферативного разделения11. Клоны RGP, индуцированные между E10-E12, были в среднем больше по размеру и обеспечивали более пространственные особенности окончательного распределения нейронов(рисунок 4A-C). Клоны с нейронами относительно равномерно распределены по глубоким и поверхностным слоям, приобрели «цилиндрическую» форму, в то время как клоны с нейронами более рассеяны поверхностными слоями, чем более глубокие слои, выработали «конусную» форму11. Чтобы полностью зафиксировать пространственную и морфологическую информацию клона, необходимо было вычислить каждый клон с помощью последовательных изображений. Для измерения клональной дисперсии максимальная боковая дисперсия (измеренная во всех измерениях) в поверхностных слоях клона была сопоставлена с дисперсией нейронов в глубоких слоях (LV/LIV). Это соотношение (распределение верхнее: распределение ниже) обеспечивает количественное считывание общей формы клона.
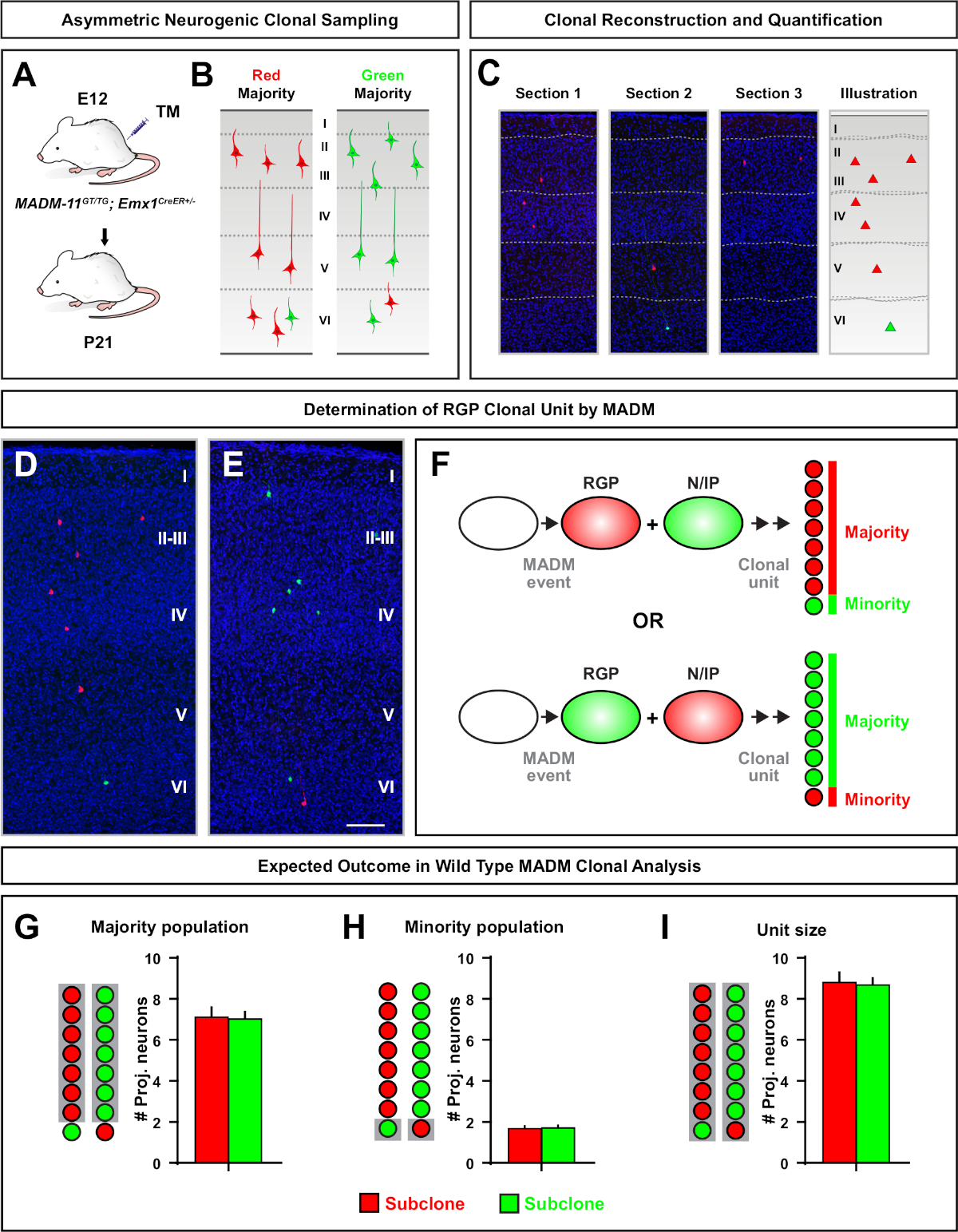
Асимметричные клоны, где меньшинство подклон было три или меньше, при условии, понимание нейронов выход одного RGP (Рисунок 4D-F и Рисунок 5A-F)7,11,12. Большинство населения (большой субклон) может быть помечены либо красным или зеленым, в среднем около семи возбуждающих нейронов проекции на клон при индуцированной с помощью Emx1-CreERT2 или Nestin-CreERT2(Рисунок 5G)7,11,12. Общее количество клеток в клоне MADM может быть дополнительно вскрыто путем анализа распределения нейронов в большом подклоне по поверхностным и глубоким слоям. Меньшинство (небольшой субклон) был помечен взаимным цветом и был в среднем 1'2 клеток на клона(рисунок 5H). Общий "размер единицы", который был в среднем 8'9 нейронов, может быть рассчитана путем добавления малых и больших подклонов вместе (Рисунок 5I)7,11,12. Важно отметить, что в то время как нейрональный выход RGPs был весьма предсказуемым, была степень клональной неоднородности12,70.
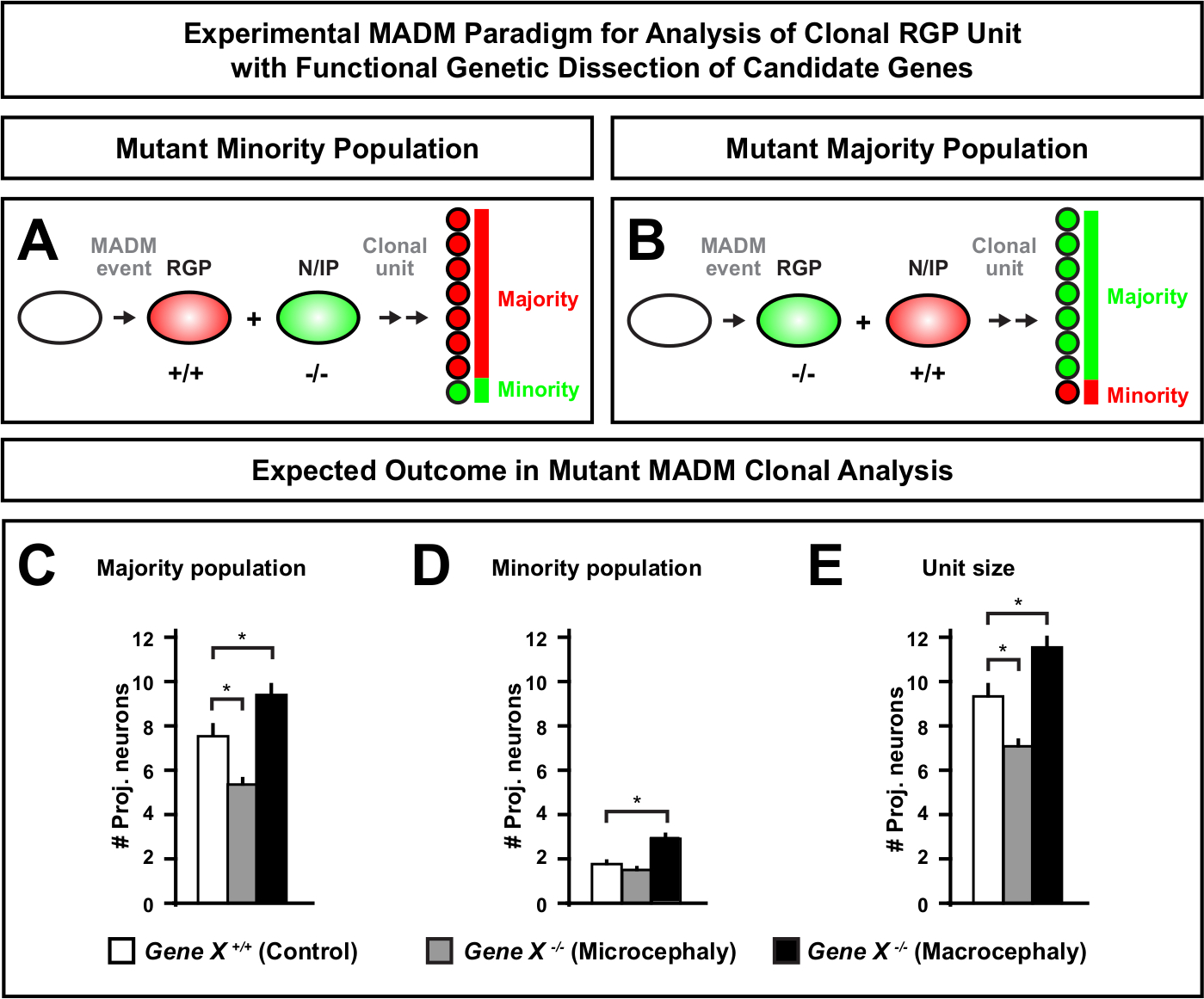
Введение мутации дистальной к кассете MADM позволяет создать генетическую мозаику, обеспечивая уникальный метод для рассечения молекулярных регуляторов прогрессирования линии стволовых клеток. Таким образом, MADM предоставляет беспрецедентную экспериментальную платформу для изучения клеточно-автономной функции гена (например, его связь с микроцефалией или макроцефалией). Сравнивая клоны, индуцированные в генетической мозаике MADM, с клонами, индуцированными в контрольном МАДМ, может быть сгенерировано высококоличественное считывание изменений в номерах и распределении нейронов. Предыдущие исследования на основе МАДМ количественно клеточно-автономной функции Otx1 в формировании микроцефалии на клональном уровне (см. рисунок 6A-E для репрезентативного примера)11. В другом исследовании, MADM клональный анализ показал, что Ndel1 не клеточно-автономно регулировать проекционные нейронные номера, но вместо этого способность новорожденных нейронов вводить или мигрировать в корковой пластине, которая позже образует взрослую кору46. Эти исследования продемонстрировали высококоличественный характер клонального анализа MADM при изучении клеточно-автономных функций генов, регулирующих корковое развитие. В настоящее время в литературе нет примеров, использующих MADM для изучения генов, вовлеченных в макроцефалию на клональном уровне. Однако в будущих исследованиях анализ генов, имеющих отношение к контролю размеров коры головногоса, в целом может обеспечить весьма желательные идеи на молекулярном и клеточном уровне.

Рисунок 1: Принцип MADM для отслеживания линий и клонального анализа на уровне одной стволовой клетки. (A) Для выполнения отслеживания линий и клонального анализа с MADM должны присутствовать два компонента. Во-первых, кассеты MADM должны быть ориентированы на идентичные локусы на гомологичные хромосомы. Кассеты состоят из двух химерных флуоресцентных генов репортера, eGFP (зеленый, «G») и тандем dimer Tomato (красный, tdT'T). Кассета GT содержит N-термин eGFP и C-термин tdT, разделенный интроном, содержащим локсП сайт. Кассета TG построена обратно, с N-термином tdT и C-термином eGFP. Во-вторых, выражение рекомбинации Cre должно происходить в ячейке, содержащей целевые кассеты MADM. Локссы loxP служат мишенью для кре-опосредованной межхромомной рекомбинации, что приводит к восстановлению обеих кассет выражения одновременно. Если рекомбинация происходит во время фазы G2 клеточного цикла с последующей сегрегацией X (G2-X), две клетки дочери выражают один из двух флуоресцентных белков. (B)принцип MADM для анализа генетической мозаики на одном уровне клона. Мутантные аллели (точечные мутации, удаления, вставки, loxP-фланговые условные аллели, изображенные на рисунке 1Bи т.д.) могут быть введены дисталлические к кассете TG-MADM через мейотическую рекомбинацию (см. рисунок 2 и Hippenmeyer et al.46 для подробной информации о том, как ввести мутантные аллели в систему MADM). Если G2-X Cre рекомбиназы-опосредованной межхромосомальной транс-рекомбинации происходит между кассетами MADM это приводит к одному GFP гомозиготной мутантной клетке(GeneX-/-) для гена интереса и46,47,одного tdT гомозиготной клетки дикого типа(GeneX. .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .71 Альтернативные результаты маркировки, не используемые в клональном анализе (т.е. желтые клетки), были ранее подробно описаны11,,46,,47. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}

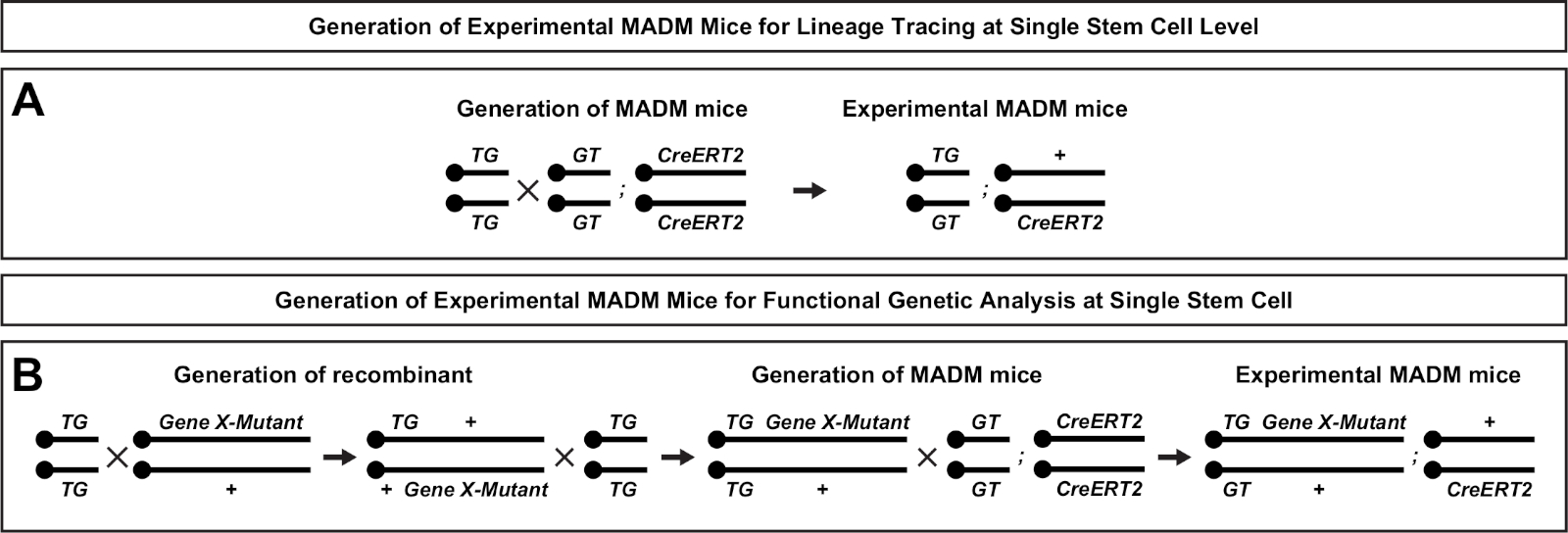
Рисунок 2: Схемы разведения для генерации экспериментальных мышей MADM для отслеживания линий. Схема разведения для генерации контроля MADM(A) и Gene X MADM(B) экспериментальных мышей MADM для клонального анализа. Для получения дополнительной информации о парадигмах разведения MADM см. Beattie et al.7 и Hippenmeyer et al.7,46. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}

Рисунок 3: Парадигмы временных курсов для анализа клональной линии на основе MADM. Схема экспериментального дизайна временных окон. Для продольных парадигм выборки временной точки индукции клонов оставалась неизменной, а продолжительность времени, прежде чем анализ был разнообразным. В прогрессивной интервальной выборке временные точки анализа оставались неизменными, однако время индукции менялось. Комбинация одного или обоих подходов может быть использована в зависимости от рассмотренных вопросов. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}

Рисунок 4: клональный анализ MADM в развивающемся и взрослом неокортексе. TM-опосредованный MADM клон индукции в симметрично пролиферативной (TM на E10) (A-C) и асимметрично нейрогенных (TM на E12) (D-F) деление RGPs. Изображены отдельные клоны MADM in vivo в развивающихся (TM/E10-A/E16 и TM/E12-A/E16) (B,E)и взрослые (TM/E10'A/P21 и TM/E12-A/P21)(C,F)в MADM-11GT/TG; Нестин-КреЕРТ2/- (B,E)и MADM-11GT/TG; Emx1-CreERT2 "/- (C,F). Нейрон выход был независим от цвета подклона и зеленого большинства / меньшинства subclones можно сравнить с красным большинством / меньшинства subclones под контролем условиях7,11. Приблизительно 1/6 взрослых клонов также содержали астроциты и/или олигодендроциты, обозначенные белыми звездочками. Панели B и F воспроизводятся с разрешения Hippenmeyer et al.46 и Rulands и Simons72, соответственно. CP и кортиковая пластина. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}

Рисунок 5: клональный анализ MADM для количественной оценки вывода нейронов RGP-опосредованного. Анализ возбуждающих нейронов (единиц) производства отдельными нейрогенными RGPs на клональном уровне с помощью MADM7,11. ( A )Экспериментальнаяпарадигма для индуцирования в основном асимметричных клонов MADM в развивающейся коре. ( B) Возможные асимметричные результаты клона с большинством субклон помечены либо зеленый или красный (C) Представитель последовательных разделов, охватывающих один нейрогенный асимметричный клон (D, E) 3D-реконструкции изображения репрезентативных G2-X MADM клонов с большинством населения в красном(D) или зеленый (E) в MADM-11GT /TG; Emx1-CreERT2/- с индукцией ТМ на E12 и анализом на P21. Обратите внимание, как зеленые, так и красные помеченные клетки дикого типа. (F) Схема с указанием двух возможных экспериментальных исходов клона MADM. (G) Количественная оценка размера большинства населения, возникшая в результате обновления РГП в клонах MADM-11. (H) Количественная оценка численности меньшинства, возникшая в результате обновления РГП в клонах МАДМ-11. (I) Количественная оценка унитарного размера асимметричных нейрогенных клонов MADM-11. Гипотетические значения могут представлять собой означает - SEM. Шкала бар No 100 мкм(D и E). ТМ и Тамоксифен. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}

Рисунок 6: клональный анализ MADM для изучения генов, которые приводят к микроцефалии и макроцефалии. Гипотетический результат клонального анализа MADM при выполнении функционального генетического вскрытия генов-кандидатов, которые приводят к микроцефалии или макроцефалии. Чтобы вскрыть клеточно-автономные функции гена интереса(Gene X) на выходе нейронов, MADM требует мутантных аллелей, которые будут введены дистальной к кассетам MADM через мейотическую рекомбинацию (подробнее отом,как ввести мутантные аллели в систему MADM, смотрите также Рисунок 2,Hippenmeyer и др.46, и Laukoter et al.46,73). (A,B) Схема, показывающая экспериментальную парадигму MADM для функционального анализа клональных единиц RGP. Мутант subclone может либо сформировать меньшинство (A) или большинство(B) населения. (C-E) Гипотетические результаты клонального анализа MADM при количественной оценке контроля MADM (белые бары), микроцефалии Gene-X MADM (серые полосы) и Gene-X MADM макроцефалии черных полосы) асимметричных клонов. (C) Количественная оценка численности большинства населения. (D) Количественная оценка численности меньшинства. (E) Количественная оценка унитарного размера асимметричных нейрогенных клонов. Гипотетические значения могут представлять собой означает SEM. S - Гипотетический сценарий, при котором разница в количестве клеток subclone может достигать значения по сравнению с контролем. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}
Таблица 1: MADM клональные исследования в литературе. Резюме исследований в литературе, содержащей ЭКСПЕРИМЕНТы по клональной линии MADM, включая использованный драйвер CreERT2, дозу ТМ и время инъекций. Пожалуйста, нажмите здесь, чтобы просмотреть эту таблицу (Право нажмите, чтобы загрузить).
Обсуждение
Описан метод использования MADM для отслеживания клеточной линии отдельных RGPs in vivo в развивающемся неокортексе. В сочетании с TM-индуцируемым CreERT2,события MADM могут быть точно приурочены, обеспечивая высоко качественное и количественное визуальное считывание моделей деления стволовых клеток на уровне одной клетки. По титрая дозы TM доставлены, в идеальной ситуации в среднем менее одного клона на корковое полушарие может быть получено, обеспечивая адекватное пространственное разделение однозначно различать отдельных клонов. Поддерживая целостность тканей, этот метод также фиксирует важную информацию о положении, морфологии и абсолютных числах клеток. Кассеты MADM на Chr. 117,11,12,46,56,57,на Chr. 751, и оригинальный MADM на Rosa2647,53,,59 были использованы в исследованиях анализа клонала MADM. Высокое разрешение отдельных клеток обеспечивает беспрецедентное понимание как морфологии и клональной связи клеток дочери и позволяет живое изображение размножающихся стволовых клеток и новых клонов46,52.
Кесарево сечение и воспитание щенков для анализа клонов в послеродовой временной точки является необходимым и важным шагом в протоколе. В зависимости от состояния здоровья беременной плотины, обработанной ТМ, может быть необязательно проводить кесарево сечение. Тем не менее, повышение щенков с приемной матерью по-прежнему требуется, потому что TM-обработанные матери могут возникнуть проблемы с кормления. Не было отмечено никаких различий в необходимости создания различных драйверов CreERT2. Обе линии MADM и приемные матери поддерживаются на выкрытом фоне CD-1. Если кесарево сечение не является необходимым, ТМ-обработанных беременных плотины, используемые для создания экспериментальных щенков могут быть повторно использованы для дополнительных экспериментальных разведения в соответствии с принципами 3R (обратите внимание, что это может быть сделано только в том случае, если животных экспериментальных лицензий одобрить эту практику). Приемные матери могут быть использованы для воспитания щенков в течение 2 дней после родов, но более высокие показатели успеха наблюдались, когда приемные матери рожают в тот же день, как экспериментальные мыши, которые должны быть воспитаны. Поэтому важно установить приуроченные спаривания для приемных матерей параллельно с экспериментальными спариваниями в шаге 1.1. Поддержание аналогичного номера помета, как помет первоначальной приемной матери может улучшить выживаемость воспитательных щенков, и, следовательно, удаление некоторых из всех оригинальных помет может быть необходимо. Дополнительные шаги, которые могут улучшить воспитание включает в себя потирая перчатки экспериментатора с мусором и пищей (чтобы удалить запах перчаток); потирая щенков нежно после кесарева сечения с фрагментами загрязненного помета и гнезда приемной матери; и размещение щенков в тесном контакте с щенками приемной матери до их размещения в клетке для приемных мышей.
Как и в других методах отслеживания линий, основанных на репортерах, при выборе оптимального драйвера CreERT2 для клональных экспериментов MADM необходимо тщательно учитывать. Во-первых, промоутер используется должны выразить рекомбинации как временно, так и пространственно в популяции прародителей интересов. Поиск этого промоутера может быть сложной задачей, потому что некоторые промоутеры могут изменить шаблоны выражения или замолчать на разных стадиях развития. Для улучшения специфичности типа клеток были использованы несколько рекомбиназу, каждая из которых была создана отдельными промоутерами. Когда одна или обе рекомбины выражены в одной клетке, это маркирует клетку и ее потомство с флуоресцентным репортером74,,75,,76,77. Таким образом, важно выбрать драйвер CreERT2, который специфичен для популяции анализируемых прародителей.
Наиболее важным шагом в этом методе является идентификация клона, поскольку все клетки должны быть получены однозначно из одного события рекомбинации (шаг 8.1). Титрация концентрации ТМ обеспечивает менее одного скопления красных/зеленых клеток на полушарие мозга и максимизирует вероятность анализа одного клона (шаг 2.2)7,11. Клоны должны быть отброшены, если соседние кластеры ячеек происходят в пределах 500 мкм от интересующего клона. Поэтому важно изучить несколько разделов до и после появления клона, чтобы убедиться, что поблизости нет дополнительных событий рекомбинации. Из-за более слабого сигнала фторфоров, необходимо выполнять иммуногистохимию для eGFP и tdT в эмбриональных клонах (см. раздел 6). Это рекомендуется только для взрослых клонов, если дополнительные антигены будут колебелированные. При клонах изображений важно захватить всю ширину коры, где находится клон (т.е. от пиальной поверхности до мозоли корпуса; см. шаг 8.4), чтобы не пропустить ни одной клетки. Это также облегчает выравнивание изображения во время обработки изображений (раздел 9). Раздел 8 протокола требует перевернутого конфокального микроскопа, но может быть адаптирован в зависимости от доступной установки микроскопа. Эпифлуоресценция микроскопия может быть использована, но конфокальная микроскопия рекомендуется, потому что это приводит к снижению светового загрязнения из-за пределов фокус-плоскости. Важно также, чтобы интенсивность и увеличение лазера корректировались таким образом, чтобы можно было однозначно определить зеленые, красные и желтые клетки. Независимо от настройки, рекомендуется использовать цель, по крайней мере 20x, чтобы обеспечить полное пространственное разделение близко расположенных клеток. В дополнение к записи корковой глубины всех клеток (шаг 8.6), корковые области, где находятся клоны, должны быть идентифицированы с помощью атласа мозга, такого как атлас мозга Аллена или других стереотакксических карт координат. Парадигма именования файлов также должна быть принята, чтобы убедиться, что изображения клонов легко идентифицировать. Следующая информация может быть включена в файл именования: уникальный идентификатор изображения, дата изображение было принято, генотип животного, возраст индукции, возраст анализа, номер изображения по отношению к остальным изображениям из того же клона.
Введение мутации дистальной к одной кассете MADM отчетливо позволяет генерации генетических мозаик71 и позволяет вскрытие молекулярных регуляторов линии и типа клеток разнообразия на уровне клональногоуровня 7,11,46,62. Чтобы создать генетическую мозаику с MADM, кассеты MADM должны быть мейотически связаны с той же хромосомой, что и ген, представляющий интерес (см. рисунок 2 для схемы размножения). Это ограничивает текущий клональный анализ с MADM генами, расположенными на Chr. 751, Chr. 1146, Chr. 1251, и Chr. 6 дистальный к Rosa26 локус47. В будущих исследованиях будут использованы кассеты MADM, ориентированные на любую хромосому, что позволит провести мозаичный анализ практически всех генов генома мыши на уровне клона.
Наконец, MADM не ограничивается анализом клеток-предшественников в развивающемся неокортексе. Изучение многих ниш стволовых клеток может извлечь выгоду из способности решать пространственно-временной договоренности клонально связанных клеток. Применяя МАДМ к другим регионам мозга, заболеваемости (например, рак), или в других тканях47,50,5151,52,53,54,55,56,57,58,59,исследования показали, линии отношения в клонов, полученных из различных классов прагенитора и стволовых клеток (см. таблицу 1 для текущего списка MADM исследований). Еще одно интересное будущее приложение MADM заключается в том, чтобы объединить его с дополнительными функциональными или субклеточными репортерами, что увеличит степень информации, которая может быть получена от клонов.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Мы благодарим всех членов лаборатории Hippenmeyer за обсуждение, Биоизмовывания фонда, фонда науки о жизни, и доклинических средств в IST Австрии за техническую поддержку. Эта работа была поддержана институциональными фондами IST Austria; Р.Б. получил поддержку от Австрийского научного фонда (FWF) Лиз-Мейтнер программы (M 2416); N.A получил поддержку от Австрийского научного фонда (FWF) Firnberg-Programm (T 1031); ГК получила поддержку от исследовательской и инновационной программы Европейского союза «Горизонт 2020» в рамках соглашения о предоставлении гранта «Мария Склодовская-Кюри» No 754411 в качестве аспиранта ISTplus; А.Х. получил поддержку от ЗAW DOC (докторская стипендия Австрийской академии наук). Это исследование было также поддержано Европейским исследовательским советом (ERC) в рамках исследовательской и инновационной программы Европейского союза Horizon 2020 (соглашение о предоставлении грантов No 725780 LinPro) S.H.
Материалы
| Name | Company | Catalog Number | Comments |
| 1 mL tuberculin syringe (Omnifix Luer Lock) | Braun | 9204512N | |
| 1,4-diazabicyclooctane (DABCO) | Roth | 0718.2 | |
| 10 mL Syringe (Omnifix Luer Lock) | Braun | 8508429N | |
| 15 mL conical centrifuge | Sarstedt | 65.554.502 | |
| 24 multi-well dishes | Roth/Greiner Bio-one | CE56.1 | |
| 27- gauge x 3/4 needle (Sterican) | Braun | 16010256E | |
| Corn oil | Sigma | C8267-500ML | |
| Coverslips (24 x 60 mm #1) | Thermo Fisher Scientific (Menzel) | 15747592 | |
| Cryostat Cryostar NX70 | Thermo Fisher Scientific | 957000H | |
| Dako Pen (Wax marker) | Agilent | S200230-2 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Invitrogen | D1306 | |
| Disposable microtome blade (MX35 Ultra) | Thermo Fisher Scientific | 705830 | |
| Fine Forceps (Dumont #5) | Fine Science Tools (FST) | 11254-20 | |
| Glass anti-roll plate | Histocom | M 449980 | |
| Glycerol | Sigma | G5516 | |
| LSM 800 Confocal | Zeiss | ||
| Mounting medium | 25 mg/mL DAPCO, 6 g Glycerol, 2.4 g Mowiol 4-88, 6 mL dH2O, 12 mL 0.2 M Tris-HCl (pH 8.5) | ||
| Mowiol 4-88 | Roth | 0713.2 | |
| Normal donkey serum | Innovative Research | IGDNSER100ML | |
| Paraformaldehyde | Sigma | 441244-1KG | |
| Peristaltic pump 323E/D 400RPM | Watson-Marlow | 036.3124.00A | |
| Sucrose | Sigma | S8501-5KG | |
| Superfrost plus glass slides | Thermo Fisher Scientific | J1800AMNT | |
| Tamoxifen | Sigma | T5648 | |
| Tissue Embedding mold T-12 (22mm square) | Polysciences Inc. | 18986-1 | |
| Tissue-Tek O.C.T | Sakura | 4583 | |
| Triton X-100 | Sigma | T8787-250ML | |
| Trizma hydrochloride | Sigma | 93363 | |
| Tween-20 | Sigma | P9416-100ML | |
| Software and Plugins: | |||
| Fiji | 1.52p | Fiji | |
| MultiStackReg | 1.45 | Download link | |
| TurboReg | EPFL Bioimaging | ||
| Zen Blue | 2.6 | Zeiss | |
| Experimental Models: Organisms/Strains: | |||
| Mouse: Emx1-CreER | The Jackson Laboratory | JAX:027784 | |
| Mouse: MADM-11-GT | The Jackson Laboratory | JAX:013749 | |
| Mouse: MADM-11-TG | The Jackson Laboratory | JAX:013751 | |
| Primary antibodies: | |||
| Chicken anti-GFP 1:500 | Aves Labs | GFP-1020 | |
| Goat anti-tdTomato 1:500 | Sicgen Antibodies | AB8181-200 | |
| Rabbit anti-RFP 1:500 | MBL | PM005 | |
| Secondary antibodies: | |||
| Donkey Anti-Chicken Alexa Fluor 488 1:500 | Jackson Immuno Research | 715-475-150 | |
| Donkey Anti-Goat Cy3 1:500 | Jackson Immuno Research | 705-165-147 | |
| Donkey Anti-Rabbit Cy3 1:500 | Jackson Immuno Research | 711-165-152 |
Ссылки
- Malatesta, P., et al. Neuronal or glial progeny: regional differences in radial glia fate. Neuron. 37 (5), 751-764 (2003).
- Miyata, T., Kawaguchi, A., Okano, H., Ogawa, M. Asymmetric inheritance of radial glial fibers by cortical neurons. Neuron. 31 (5), 727-741 (2001).
- Noctor, S. C., Flint, A. C., Weissman, T. A., Dammerman, R. S., Kriegstein, A. R. Neurons derived from radial glial cells establish radial units in neocortex. Nature. 409 (6821), 714-720 (2001).
- Taverna, E., Götz, M., Huttner, W. B. The Cell Biology of Neurogenesis: Toward an Understanding of the Development and Evolution of the Neocortex. Annual Review of Cell and Developmental Biology. 30 (1), 465-502 (2014).
- Desikan, R. S., Barkovich, A. J. Malformations of cortical development. Annals of Neurology. 80 (6), 797-810 (2016).
- Gao, R., Penzes, P. Common mechanisms of excitatory and inhibitory imbalance in schizophrenia and autism spectrum disorders. Current Molecular Medicine. 15 (2), 146-167 (2015).
- Beattie, R., et al. Mosaic Analysis with Double Markers Reveals Distinct Sequential Functions of Lgl1 in Neural Stem Cells. Neuron. 94 (3), 517-533 (2017).
- Hippenmeyer, S. Molecular pathways controlling the sequential steps of cortical projection neuron migration. Advances in Experimental Medicine and Biology. 800, 1-24 (2014).
- Lodato, S., Arlotta, P. Generating neuronal diversity in the mammalian cerebral cortex. Annual Review of Cell and Developmental Biology. 31, 699-720 (2015).
- Hansen, A. H., Duellberg, C., Mieck, C., Loose, M., Hippenmeyer, S. Cell Polarity in Cerebral Cortex Development-Cellular Architecture Shaped by Biochemical Networks. Frontiers in Cellular Neuroscience. 11, 176 (2017).
- Gao, P., et al. Deterministic progenitor behavior and unitary production of neurons in the neocortex. Cell. 159 (4), 775-788 (2014).
- Llorca, A., et al. A stochastic framework of neurogenesis underlies the assembly of neocortical cytoarchitecture. eLife. 8, e51381 (2019).
- Ma, J., Shen, Z., Yu, Y. C., Shi, S. H. Neural lineage tracing in the mammalian brain. Current Opinion in Neurobiology. 50, 7-16 (2018).
- Caviness, V., Takahashi, T., Nowakowski, R. Numbers, time and neocortical neuronogenesis: a general developmental and evolutionary model. Trends in Neurosciences. 18 (9), 379-383 (1995).
- Schmechel, D. E., Rakic, P. A Golgi study of radial glial cells in developing monkey telencephalon: morphogenesis and transformation into astrocytes. Anatomy and Embryology. 156 (2), 115-152 (1979).
- Kessaris, N., et al. Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nature Neuroscience. 9 (2), 173-179 (2006).
- Magavi, S., Friedmann, D., Banks, G., Stolfi, A., Lois, C. Coincident generation of pyramidal neurons and protoplasmic astrocytes in neocortical columns. The Journal of Neuroscience. 32 (14), 4762-4772 (2012).
- Anthony, T. E., Klein, C., Fishell, G., Heintz, N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 41 (6), 881-890 (2004).
- Voigt, T. Development of glial cells in the cerebral wall of ferrets: direct tracing of their transformation from radial glia into astrocytes. The Journal of Comparative Neurology. 289 (1), 74-88 (1989).
- Amberg, N., Laukoter, S., Hippenmeyer, S. Epigenetic cues modulating the generation of cell-type diversity in the cerebral cortex. Journal of Neurochemistry. 149 (1), 12-26 (2019).
- Beattie, R., Hippenmeyer, S. Mechanisms of Radial Glia Progenitor Cell Lineage Progression. FEBS letters. 591 (24), 3993-4008 (2017).
- Telley, L., et al. Temporal patterning of apical progenitors and their daughter neurons in the developing neocortex. Science. 364 (6440), eaav2522 (2019).
- Oberst, P., et al. Temporal plasticity of apical progenitors in the developing mouse neocortex. Nature. 573 (7774), 370-374 (2019).
- Telley, L., et al. Sequential transcriptional waves direct the differentiation of newborn neurons in the mouse neocortex. Science. 351 (6280), 1443 (2016).
- Deppe, U., et al. Cell lineages of the embryo of the nematode Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America. 75 (1), 376-380 (1978).
- Woodworth, M. B., Girskis, K. M., Walsh, C. A. Building a lineage from single cells: genetic techniques for cell lineage tracking. Nature Reviews Genetics. 18 (4), 230-244 (2017).
- Masuyama, N., Mori, H., Yachie, N. DNA barcodes evolve for high-resolution cell lineage tracing. Current Opinion in Chemical Biology. 52, 63-71 (2019).
- Legue, E., Joyner, A. L. Chapter Ten-Genetic Fate Mapping Using Site-Specific Recombinases. Methods in Enzymology. 477, 153-181 (2010).
- Postiglione, M. P., Hippenmeyer, S. Monitoring neurogenesis in the cerebral cortex: an update. Future Neurology. 9 (3), 323-340 (2014).
- Espinosa-Medina, I., Garcia-Marques, J., Cepko, C., Lee, T. High-throughput dense reconstruction of cell lineages. Open Biology. 9 (12), 190229 (2019).
- Hwang, B., et al. Lineage tracing using a Cas9-deaminase barcoding system targeting endogenous L1 elements. Nature Communications. 10 (1), 1234 (2019).
- Kalhor, R., et al. Developmental barcoding of whole mouse via homing CRISPR. Science. 361 (6405), eaat9804 (2018).
- García-Moreno, F., Vasistha, N. A., Begbie, J., Molnár, Z. CLoNe is a new method to target single progenitors and study their progeny in mouse and chick. Development. 141 (7), 1589-1598 (2014).
- Loulier, K., et al. Multiplex Cell and Lineage Tracking with Combinatorial Labels. Neuron. 81 (3), 505-520 (2014).
- Livet, J., et al. Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature. 450 (7166), 56-62 (2007).
- Snippert, H. J., et al. Intestinal Crypt Homeostasis Results from Neutral Competition between Symmetrically Dividing Lgr5 Stem Cells. Cell. 143 (1), 134-144 (2010).
- Amitai-Lange, A., et al. A method for lineage tracing of corneal cells using multi-color fluorescent reporter mice. Journal of Visualized Experiments. (106), e53370 (2015).
- Vasistha, N. A., et al. Cortical and Clonal Contribution of Tbr2 Expressing Progenitors in the Developing Mouse Brain. Cerebral Cortex. 25 (10), 3290-3302 (2015).
- Chen, F., LoTurco, J. A method for stable transgenesis of radial glia lineage in rat neocortex by piggyBac mediated transposition. Journal of Neuroscience Methods. 207 (2), 172-180 (2012).
- Siddiqi, F., et al. Fate mapping by piggyBac transposase reveals that neocortical GLAST+ progenitors generate more astrocytes than Nestin+ progenitors in rat neocortex. Cerebral Cortex. 24 (2), 508-520 (2014).
- Yoshida, A., et al. Simultaneous expression of different transgenes in neurons and glia by combining in utero electroporation with the Tol2 transposon-mediated gene transfer system. Genes to Cells. 15 (5), 501-512 (2010).
- García-Marqués, J., López-Mascaraque, L. Clonal Identity Determines Astrocyte Cortical Heterogeneity. Cerebral Cortex. 23 (6), 1463-1472 (2013).
- Clavreul, S., et al. Cortical astrocytes develop in a plastic manner at both clonal and cellular levels. Nature Communications. 10 (1), 4884 (2019).
- Kim, G. B., et al. Rapid Generation of Somatic Mouse Mosaics with Locus-Specific, Stably Integrated Transgenic Elements. Cell. 179 (1), 251-267 (2019).
- Pilz, G. A., et al. Live imaging of neurogenesis in the adult mouse hippocampus. Science. 359 (6376), 658 (2018).
- Hippenmeyer, S., et al. Genetic Mosaic Dissection of Lis1 and Ndel1 in Neuronal Migration. Neuron. 68 (4), 695-709 (2010).
- Zong, H., Espinosa, J. S., Su, H. H., Muzumdar, M. D., Luo, L. Mosaic analysis with double markers in mice. Cell. 121 (3), 479-492 (2005).
- Ramón y Cajal, S. . Histologie du système nerveux de l'homme et des vertébrés. , (1911).
- Cowan, W. M. The emergence of modern neuroanatomy and developmental neurobiology. Neuron. 20 (3), 413-426 (1998).
- Ali, S. R., et al. Existing cardiomyocytes generate cardiomyocytes at a low rate after birth in mice. Proceedings of the National Academy of Sciences of the United States of America. 111 (24), 8850-8855 (2014).
- Hippenmeyer, S., Johnson, R. L., Luo, L. Mosaic analysis with double markers reveals cell-type-specific paternal growth dominance. Cell Reports. 3 (3), 960-967 (2013).
- Riccio, P., Cebrian, C., Zong, H., Hippenmeyer, S., Costantini, F. Ret and Etv4 promote directed movements of progenitor cells during renal branching morphogenesis. PLoS Biology. 14 (2), e1002382 (2016).
- Bonaguidi, M. A., et al. In vivo clonal analysis reveals self-renewing and multipotent adult neural stem cell characteristics. Cell. 145 (7), 1142-1155 (2011).
- Mayer, C., et al. Clonally Related Forebrain Interneurons Disperse Broadly across Both Functional Areas and Structural Boundaries. Neuron. 87 (5), 989-998 (2015).
- Muzumdar, M. D., et al. Clonal dynamics following p53 loss of heterozygosity in Kras-driven cancers. Nature Communications. 7, 12685 (2016).
- Shi, W., et al. Ontogenetic establishment of order-specific nuclear organization in the mammalian thalamus. Nature Neuroscience. 20, 516 (2017).
- Wong, S. Z. H., et al. In vivo clonal analysis reveals spatiotemporal regulation of thalamic nucleogenesis. PLoS Biology. 16 (4), e2005211 (2018).
- Xu, H. T., et al. Distinct Lineage-Dependent Structural and Functional Organization of the Hippocampus. Cell. 157 (7), 1552-1564 (2014).
- Brennand, K., Huangfu, D., Melton, D. All β Cells Contribute Equally to Islet Growth and Maintenance. PLoS Biology. 5 (7), e163 (2007).
- Ortiz-Alvarez, G., et al. Adult neural stem cells and multiciliated ependymal cells share a common lineage regulated by the geminin family members. Neuron. 102 (1), 159-172 (2019).
- Kaplan, E. S., Ramos-Laguna, K. A., Mihalas, A. B., Daza, R. A. M., Hevner, R. F. Neocortical Sox9+ radial glia generate glutamatergic neurons for all layers, but lack discernible evidence of early laminar fate restriction. Neural Development. 12 (1), 14 (2017).
- Lv, X., et al. TBR2 coordinates neurogenesis expansion and precise microcircuit organization via Protocadherin 19 in the mammalian cortex. Nature Communications. 10 (1), 3946 (2019).
- Mihalas, A. B., Hevner, R. F. Clonal analysis reveals laminar fate multipotency and daughter cell apoptosis of mouse cortical intermediate progenitors. Development. 145 (17), dev164335 (2018).
- Picco, N., et al. A mathematical insight into cell labelling experiments for clonal analysis. Journal of Anatomy. 235 (3), 687-696 (2019).
- Johnson, C. A., Ghashghaei, H. T. Sp2 regulates late neurogenic but not early expansive divisions of neural stem cells underlying population growth in the mouse cortex. Development. , (2020).
- Espinosa, J. S., Luo, L. Timing Neurogenesis and Differentiation: Insights from Quantitative Clonal Analyses of Cerebellar Granule Cells. The Journal of Neuroscience. 28 (10), 2301 (2008).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Liu, Y., et al. Tamoxifen-independent recombination in the RIP-CreER mouse. PLoS One. 5 (10), e13533 (2010).
- Klingler, E., Jabaudon, D. Do progenitors play dice?. eLife. 9, e54042 (2020).
- Hippenmeyer, S. Dissection of gene function at clonal level using mosaic analysis with double markers. Frontiers in Biology. 8 (6), 557-568 (2013).
- Rulands, S., Simons, B. D. Tracing cellular dynamics in tissue development, maintenance and disease. Current Opinion in Cell Biology. 43, 38-45 (2016).
- Laukoter, S., et al. Imprinted Cdkn1c genomic locus cell-autonomously promotes cell survival in cerebral cortex development. Nature Communications. 11 (1), 195 (2020).
- Daigle, T. L., et al. A Suite of Transgenic Driver and Reporter Mouse Lines with Enhanced Brain-Cell-Type Targeting and Functionality. Cell. 174 (2), 465-480 (2018).
- He, M., et al. Strategies and Tools for Combinatorial Targeting of GABAergic Neurons in Mouse Cerebral Cortex. Neuron. 91 (6), 1228-1243 (2016).
- Yamamoto, M., et al. A multifunctional reporter mouse line for Cre- and FLP-dependent lineage analysis. Genesis. 47 (2), 107-114 (2009).
- Plummer, N. W., et al. Expanding the power of recombinase-based labeling to uncover cellular diversity. Development. 142 (24), 4385 (2015).
- Imayoshi, I., Ohtsuka, T., Metzger, D., Chambon, P., Kageyama, R. Temporal regulation of Cre recombinase activity in neural stem cells. Genesis. 44 (5), 233-238 (2006).
- Sasaki, S., et al. Complete loss of Ndel1 results in neuronal migration defects and early embryonic lethality. Molecular and Cellular Biology. 25 (17), 7812-7827 (2005).
- Acampora, D., et al. Epilepsy and brain abnormalities in mice lacking the Otx1 gene. Nature Genetics. 14 (2), 218-222 (1996).
- Soeda, T., et al. Sox9-expressing precursors are the cellular origin of the cruciate ligament of the knee joint and the limb tendons. Genesis. 48 (11), 635-644 (2010).
- Klezovitch, O., Fernandez, T. E., Tapscott, S. J., Vasioukhin, V. Loss of cell polarity causes severe brain dysplasia in Lgl1 knockout mice. Genes & Development. 18 (5), 559-571 (2004).
- Pimeisl, I. M., et al. Generation and characterization of a tamoxifen-inducible EomesCreER mouse line. Genesis. 51 (10), 725-733 (2013).
- Nakagawa, N., et al. Memo1-Mediated Tiling of Radial Glial Cells Facilitates Cerebral Cortical Development. Neuron. 103 (5), 836-852 (2019).
- Nowotschin, S., et al. The T-box transcription factor Eomesodermin is essential for AVE induction in the mouse embryo. Genes & Development. 27 (9), 997-1002 (2013).
- Balordi, F., Fishell, G. Mosaic removal of hedgehog signaling in the adult SVZ reveals that the residual wild-type stem cells have a limited capacity for self-renewal. Journal of Neuroscience. 27 (52), 14248-14259 (2007).
- Liang, H., et al. Neural development is dependent on the function of specificity protein 2 in cell cycle progression. Development. 140 (3), 552-561 (2013).
- Guo, C., Yang, W., Lobe, C. G. A Cre recombinase transgene with mosaic, widespread tamoxifen-inducible action. Genesis. 32 (1), 8-18 (2002).
- Ahn, S., Joyner, A. L. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature. 437 (7060), 894-897 (2005).
- Storm, R., et al. The bHLH transcription factor Olig3 marks the dorsal neuroepithelium of the hindbrain and is essential for the development of brainstem nuclei. Development. 136 (2), 295-305 (2009).
- Koundakjian, E. J., Appler, J. L., Goodrich, L. V. Auditory neurons make stereotyped wiring decisions before maturation of their targets. Journal of Neuroscience. 27 (51), 14078-14088 (2007).
- Dor, Y., Brown, J., Martinez, O. I., Melton, D. A. Adult pancreatic β-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 429 (6987), 41-46 (2004).
- Sohal, D. S., et al. Temporally regulated and tissue-specific gene manipulations in the adult and embryonic heart using a tamoxifen-inducible Cre protein. Circulation Research. 89 (1), 20-25 (2001).
- Ventura, A., et al. Restoration of p53 function leads to tumour regression in vivo. Nature. 445 (7128), 661-665 (2007).
- Johnson, L., et al. Somatic activation of the K-ras oncogene causes early onset lung cancer in mice. Nature. 410 (6832), 1111-1116 (2001).
- Tasic, B., et al. Extensions of MADM (mosaic analysis with double markers) in mice. PLoS One. 7 (3), e33332 (2012).
- Yu, J., Carroll, T. J., McMahon, A. P. Sonic hedgehog regulates proliferation and differentiation of mesenchymal cells in the mouse metanephric kidney. Development. 129 (22), 5301-5312 (2002).
- Zhao, H., et al. Role of fibroblast growth factor receptors 1 and 2 in the ureteric bud. Developmental Biology. 276 (2), 403-415 (2004).
- Schuchardt, A., D'Agati, V., Larsson-Blomberg, L., Costantini, F., Pachnis, V. Defects in the kidney and enteric nervous system of mice lacking the tyrosine kinase receptor Ret. Nature. 367 (6461), 380-383 (1994).
- Livet, J., et al. ETS gene Pea3 controls the central position and terminal arborization of specific motor neuron pools. Neuron. 35 (5), 877-892 (2002).
- Volz, K. S., et al. Pericytes are progenitors for coronary artery smooth muscle. eLife. 4, e10036 (2015).
- Cai, C. L., et al. A myocardial lineage derives from Tbx18 epicardial cells. Nature. 454 (7200), 104-108 (2008).
- Lasrado, R., et al. Lineage-dependent spatial and functional organization of the mammalian enteric nervous system. Science. 356 (6339), 722-726 (2017).
- Matsuoka, T., et al. Neural crest origins of the neck and shoulder. Nature. 436 (7049), 347-355 (2005).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены