
Method Article
Rastreamento de linhagem e análise clonal no desenvolvimento de córtex cerebral usando análise de mosaico com marcadores duplos (MADM)
Neste Artigo
Resumo
Um protocolo para realizar o rastreamento de linhagem e a análise genética funcional dos genes candidatos em um único nível celular usando a análise de mosaico com marcadores duplos (MADM). A análise clonal madm fornece uma estrutura quantitativa para medir o comportamento proliferativo, a saída celular e a relação de linhagem de progenitores individuais e suas células filhas.
Resumo
A partir de uma piscina limitada de progenitores, o córtex cerebral mamífero forma circuitos neurais funcionais altamente organizados. No entanto, os mecanismos celulares e moleculares subjacentes que regulam as transições de linhagem das células-tronco neurais (NSCs) e a eventual produção de neurônios e glia no neuroepithelium em desenvolvimento permanece incerto. Métodos para rastrear padrões de divisão NSC e mapear a linhagem de células clonalmente relacionadas avançaram dramaticamente. No entanto, muitas técnicas contemporâneas de rastreamento de linhagem sofrem com a falta de resolução celular do destino celular progênero, o que é essencial para decifrar padrões de divisão celular progenitora. Apresentado é um protocolo que utiliza análise de mosaico com marcadores duplos (MADM) para realizar análises clonais in vivo. O MADM manipula concomitantemente células progenitoras individuais e visualiza padrões precisos de divisão e progressão de linhagem em resolução celular única sem precedentes. Eventos de recombinação intercromossômica baseados em MADM durante a fase G2-X da mitose, juntamente com o CreERT2temporalmente indutível, fornecem informações exatas sobre as datas de nascimento dos clones e seus padrões de divisão. Assim, o rastreamento de linhagem MADM fornece leituras ópticas qualitativas e quantitativas sem precedentes do modo de proliferação de progenitores de células-tronco no nível único da célula. O MADM também permite o exame dos mecanismos e requisitos funcionais dos genes candidatos na progressão da linhagem NSC. Este método é único na forma de que a análise comparativa de subclonas de controle e mutantes pode ser realizada no mesmo ambiente tecidual in vivo. Aqui, o protocolo é descrito em detalhes, e paradigmas experimentais para empregar MADM para análise clonal e rastreamento de linhagem no córtex cerebral em desenvolvimento são demonstrados. É importante ressaltar que este protocolo pode ser adaptado para realizar análises clonais MADM em qualquer nicho de células-tronco murinas, desde que o driver CreERT2 esteja presente.
Introdução
O córtex cerebral é uma estrutura altamente organizada composta de seis camadas distintas. O córtex contém uma variedade diversificada de tipos celulares, incluindo neurônios e glia, que interagem para formar circuitos neurais funcionais. A maioria, se não todos, os neurônios de projeção excitatória cortical e a glia são derivados de um conjunto comum de células-tronco neurais (NSCs) conhecidos como progenitores gliais radiais (RGPs)1,,2,3. Os próprios RGPs são derivados de células-tronco neuroepiteliais (NESCs) que compõem o neuroepithelium embrionário precoce. No dia 9 (E9) em camundongos, os NESCs começam a fazer a transição para RGPs4. A progressão da linhagem RGP requer regulação temporal e espacial precisa, e quando esse processo é dificultado, doenças neurológicas graves como megalencefalia, microcefalia, lissencefalia ou prejuízos como esquizofrenia e autismo podem resultar5,,6. Na E10, a maioria dos RGPs sofre divisões proliferativas simétricas, resultando em uma expansão da piscina progenitora neural4,7. Os RGPs eventualmente começam a se dividir assimetriamente, produzindo neurônios de projeção cortical de forma temporalmente definida. Através de ondas consecutivas de neurogênese, os neurônios recém-nascidos migram para a placa cortical formando laminae cortical com neurônios nascidos precocemente ocupando camadas profundas e neurônios nascidos tardios residentes nas camadas superficiais8,,9,,10. Como os neurônios piramiológicos relacionados clonalmente migram radialmente para o córtex com muito pouca dispersão tangencial, as células filhas tendem a formar uma estrutura em forma de coluna ou cone referida como uma unidade radial neuronal4,,11,,12,13. Por E17, a expansão neurogênica embrionária está completa em camundongos14. Os RGPs também podem produzir células ependímicas e algumas classes de glia, incluindo astrócitos e oligódendrocitos1,,15,,16,,17,,18,19. O potencial dos RGPs para dar origem a neurônios e astrócitos parece ser consistente em todas as regiões corticais18, com aproximadamente 1/6 de RGPs neurogênicos também produzindo glia11.
Atualmente, os fatores genéticos e epigenéticos que regulam a progressão temporal de uma célula-tronco ao longo de sua linhagem são, em sua maioria, desconhecidos. Padrões temporais de expressão genética podem ter impacto substancial nas decisões de linhagem em RGPs20,21,22,23,24. Não se sabe como essa relação firmemente tricotado entre a padronização temporal e espacial leva à diversidade molecular dos tipos neuronais adultos em áreas corticais. Da mesma forma, como o potencial individual das células-tronco e sua saída celular é modulado no nível celular e molecular é uma importante pergunta sem resposta. Estudos futuros abordarão algumas dessas questões, expandindo nossa compreensão da formação de circuitos corticais funcionais.
A neurobiologia do desenvolvimento busca entender a relação de linhagem que as células do cérebro compartilham entre si. Inicialmente, poucas ferramentas de pesquisa estavam disponíveis para isso, e muitos estudos iniciais se baseavam em observações visuais de padrões de divisão em organismos transparentes, como caenorhabditis elegans25. Nas últimas décadas, houve um aumento dramático no número e sofisticação das técnicas disponíveis13,26,27,28,,29. O surgimento do sistema de edição do genoma CRISPR-Cas9 permite a reconstrução sintética das relações de linhagem celular, introduzindo códigos de barras de DNAem evolução 27,30. Dois exemplos recentes de estratégias de codificação de barras incluem o uso de RNA guia de homing que direciona o CRISPR-Cas9 para loci de código de barras de DNA específico ou uma deaminase de cytidine fundida com nickase Cas9 para atingir regiões de repetição endógenas31,,32. Essas tecnologias fornecem abordagens altamente multiplexadas através da introdução de códigos de barras que progressivamente e stably acumulam mutações únicas ao longo do tempo. As abordagens de edição de genomas são altamente valiosas porque permitem a análise retroativa da relação entre as duas células com base na herança compartilhada desses códigos de barras. No entanto, para ler os códigos de barras em células individuais, o tecido geralmente deve ser interrompido, e, portanto, se perde informações sobre posição, morfologia e números de células absolutas de um progenitor individual.
Paradigmas de rotulagem combinatória preservam informações espaciais e, em princípio, também permitem a distinção entre clones intimamente localizados ou mesmo sobrepostos33,34. Para que um método de rastreamento de linhagem seja informativo, deve rotular os progenitores individuais e seus descendentes de forma esparsa e indelével. Notavelmente, as abordagens Brainbow35 e Confetti36,37 usam repórteres baseados em cre estocásticos baseados em cre recombinase que expressam uma combinação de proteínas fluorescentes a partir de um único lócus. O extenso número de combinações simultâneas de cores que podem ser alcançadas in vivo fazem desta uma ferramenta poderosa ao rastrear clones e astrócitos cortical RGP34. Sistemas baseados em transposon que fornecem integração genômica estável de transgenes codificando repórteres fluorescentes e permitindo o rastreamento de linhagem de progenitores corticais também foram desenvolvidos33,38,,39,,40,,41. Os sistemas baseados em Transposon têm uma vantagem adicional na forma de integrar-se ao genoma e, assim, rotular de forma confiável células filhas relacionadas com linha. Para traçar as linhagens de astrócitos especificamente, foram desenvolvidos vários métodos que envolvem a eletroporação de transposases de piggyBac, incluindo o Star Track,que faz uso de uma combinação de construções codificando diferentes proteínas fluorescentes40,42. Outra abordagem, marcadores MAGIC,introduz os vetores brainbow como transgenes transposáveis. Isso foi usado com sucesso para rastrear progenitores neurais e astrócitos embrionários34,43. Recentemente, a análise do mosaico por dupla troca de mediada por recombinase (MADR) foi encontrada para rotular as células mutantes expressando elementos transgênicos do loci cromossômico44. Essas poderosas técnicas de rotulagem combinatória in vivo forneceram inúmeras percepções sobre a dinâmica de linhagem das células progenitoras. No entanto, essas análises são realizadas em tecido fixo, fornecendo um instantâneo de clones individuais em um estágio de desenvolvimento definido. Para observar mudanças na dinâmica de linhagem de clones únicos ao longo do tempo, métodos crônicos de imagem in vivo semelhantes aos realizados no giro dento adulto precisam ser aplicados45.
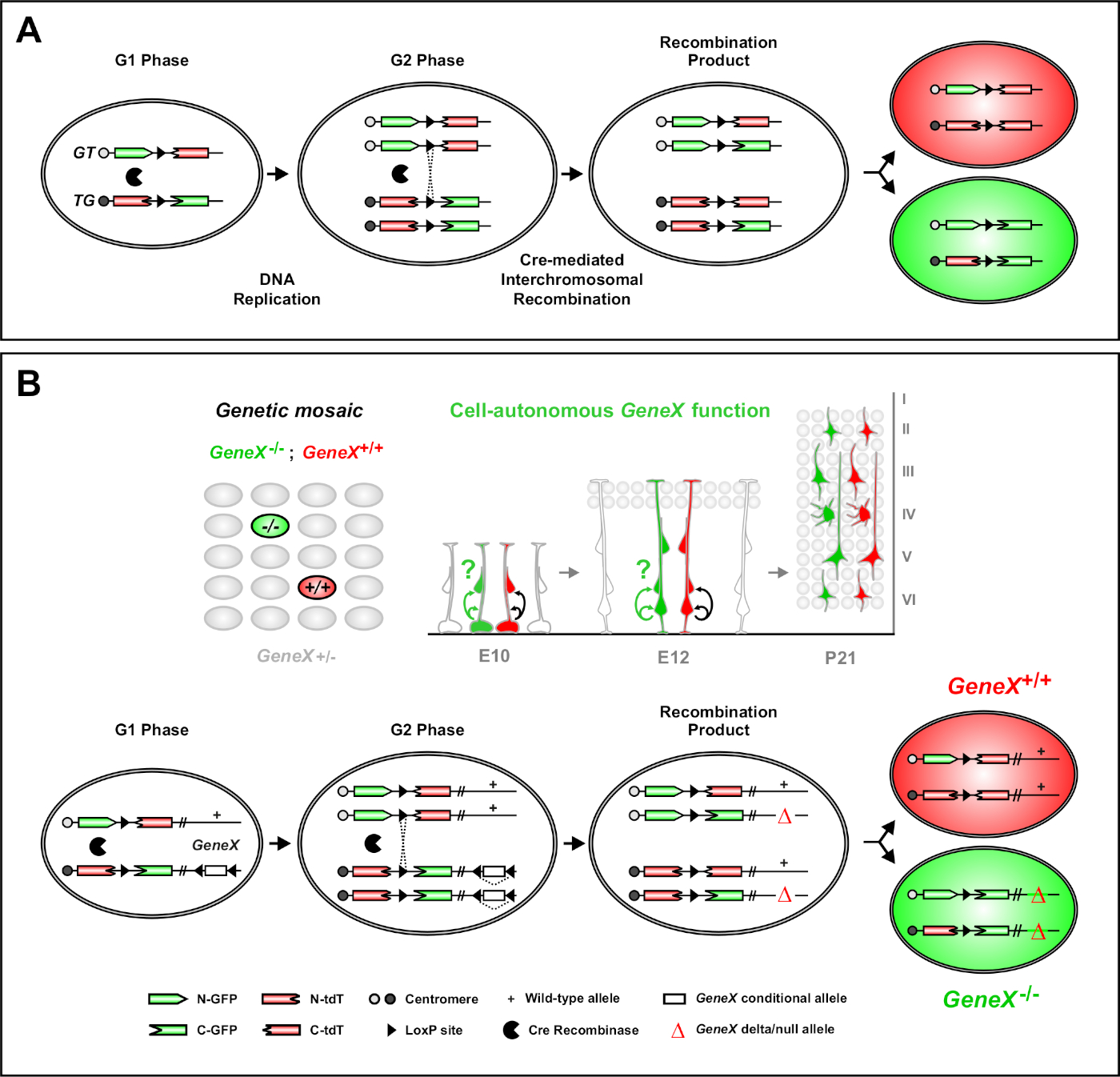
A análise de mosaico com marcadores duplos (MADM) é um poderoso método de rotulagem de cores duplas que permite o rastreamento de linhagem in vivo de células progenitoras individuais em camundongos46,47. Dois componentes são necessários para que os eventos de rotulagem MADM ocorram: Primeiro, as fitas MADM devem ser direcionadas a loci idênticas em cromossomos homólogos. As consistem em dois genes de repórteres fluorescentes quiméricos, eGFP (verde, [G]) e tandem dimer Tomate (vermelho, tdT[T]). O GT contém o N-terminus de eGFP e o C-terminus de tdT, separado por um intron contendo um site loxP. O TG é construído inversamente, com o N-terminus de tdT e o C-terminus do eGFP. Em segundo lugar, a expressão de Cre recombinase na mesma célula contendo as fitas MADM direcionadas é essencial. Na ausência de Cre,as fitas quimricas não expressam eGFP ou tdT funcionais porque suas sequências de codificação são interrompidas. Os sites loxP servem como alvo para a recombinação intercromossômica mediada por Cre, resultando na reconstituição de ambas as fitas de expressão simultaneamente. Se a recombinação ocorrer durante a fase G2 do ciclo celular seguido pela segregação X (G2-X), as duas células filhas expressarão cada uma das duas proteínas fluorescentes. A regulação temporal da atividade CreERT2 utilizando tamoxifen (TM) fornece informações precisas sobre a data de nascimento dos clones MADM e os padrões de divisão de seus descendentes (Figura 1A)29,,46,47.
O MADM pode rotular potencialmente clones individuais com alta resolução de células individuais no cérebro do camundongo semelhante aos métodos tradicionais, mas não específicos e trabalhosos, como a coloração de Golgi48 ou o enchimento decorantes 49. Como apenas o promotor que conduz o CreERT2 determina a especificidade do tipo celular da rotulagem clonal MADM, o MADM pode, em princípio, ser aplicado para rastreamento de linhagem clonal em qualquer órgão e tecido murino47,,50,,51,,52. De fato, estudos já utilizaram o MADM para revelar relações de linhagem em clones derivados de tecidos diversos47,50,51,52,53,54,55,56,57,58,59. Paradigmas experimentais MADM têm sido aplicados para estudar a linhagem em neurônios de projeção cortical, glia e células-tronco pós-natal no neocórtex em desenvolvimento7,11,12,46,60,,61,62,63,64,65. O MADM também tem sido usado para estudar a linhagem celular no giro dento adulto, tálamo, células de grânulo cerebelar e interneurons no nível clonal (ver Tabela 1 para uma lista completa)47,53,,54,,56,,57,66.
Uma característica única do MADM é a capacidade de vincular geneticamente mutações distais a um MADM, criando assim um mosaico genético(Figura 1B e Figura 2). Isso resulta em células filhas do tipo selvagem rotuladas com um marcador fluorescente (tdT na Figura 1B) e irmãos mutantes homozigosos com o outro (eGFP na Figura 1B) em um ambiente heterozigoso sem rótulo. MADM é único na que a análise comparativa de subclonas de controle e mutantes pode ser realizada no mesmo ambiente tecidual in vivo. Originalmente, as fitas MADM eram direcionadas para o lócus Rosa26 47,mas a análise MADM da função genética limitava-se aos genes distais ao lócus. Para superar (pelo menos em parte) essa limitação e ampliar as possibilidades de análises genéticas baseadas em MADM, as fitas MADM foram derrubadas perto dos centrosmers de Chr. 751, Chr. 1146e Chr. 1251. Direcionar todos os 19 autossómos do mouse com MADM está em andamento e permitirá que praticamente qualquer gene seja estudado no futuro, fornecendo uma plataforma incomparável para o estudo de relações de linhagem de desenvolvimento em combinação com a análise genética funcional.
Protocolo
Os protocolos de mouse foram revisados pelo núcleo institucional pré-clínico (PCF) e pelo comitê interno de ética da IST Austria. Toda a reprodução e experimentação foram realizadas sob uma licença aprovada pelo Ministério Federal austríaco de Ciência e Pesquisa de acordo com as leis de animais austríacas e da UE.
1. Reprodução de camundongos experimentais para análise clonal MADM
- Configurar acasalamentos madm experimentais cronometrado (>P56; CD-1) no final da tarde (17h) e verifique se há plugues vaginais na manhã de hoje (8h). Pela manhã, a ficha está presente conta como dia 0,5. Consulte a Figura 2 para obter uma visão geral da configuração experimental de acasalamento do mouse. Certifique-se de que os pontos de tempo para a indução de TM da atividade e análise do CreERT2 sejam apropriados para responder a questões experimentais.
NOTA: Para obter mais informações, consulte a Figura 3 e os resultados representativos abaixo. - Para amostragem pós-natal, configure criações para gerar mães adotivas em paralelo.
NOTA: Estes devem ser iniciados até 1-2 dias antes da criação de criações experimentais.
2. Indução TM em camundongos MADM
- Prepare uma solução de trabalho TM de 20 mg/mL dissolvendo-a em óleo de milho em um tubo de centrífuga cônica de 15 mL ou 50 mL e colocando-a em uma plataforma de balanço por ~4 h à temperatura ambiente (RT), garantindo que o TM seja completamente dissolvido. Armazene a solução de trabalho a 4 °C coberta com papel alumínio e use dentro de 2 semanas.
- Para induzir eventos de recombinação MADM, ofereça uma única injeção de TM intraperitoneally (IP) usando uma seringa tuberculina de 1 mL e uma agulha de 25 G em uma represa grávida cronometrada. Dependendo do estágio da neurogênese cortical, injete TM entre E10-E15 em uma dose de 1-2 mg/represa grávida. Para os primeiros pontos de tempo (ou seja, E10) utilize um máximo de 1 mg/barragem de gestante (25 mg/kg) para evitar complicações durante a gravidez11. Para pontos de tempo entre E11-E15 use 2 mg/represa grávida (50 mg/kg)7.
NOTA: Alternativamente, o TM pode ser administrado com um gavage oral para gestações tardias. - Para análise clonal madm para pontos de tempo pós-natal, recupere embriões vivos em E18-E19 através de cesariana, e depois crie filhotes com uma mãe adotiva.
NOTA: Dependendo do estado de saúde da gestante, pode não ser necessário realizar uma cesariana, mas criar filhotes com uma mãe adotiva ainda é necessário porque a mãe tratada com TM original pode ter problemas para lactar. - Para recuperar embriões vivos por cesariana ou para recuperar pontos de tempo embrionários para análise, sacrifique a represa grávida por deslocamento cervical.
- Coloque o animal em posição supina e desinfete a pele com 70% de etanol. Faça uma pequena incisão na pele no abdômen inferior acima do útero usando fórceps cirúrgicos e tesouras. Faça uma segunda incisão através dos músculos e da parede muscular abdominal para revelar o peritônio.
- Remova o útero separando-se dos tecidos circundantes com uma tesoura. Transfira o útero intacto para uma luva cheia de água morna para aumentar a taxa de sobrevivência do embrião até que cada um seja removido do amnion individualmente.
- Use tesouras e dedos finos para abrir cuidadosamente as paredes uterinas para liberar embriões. Não corte os cordões umbilicais muito perto do corpo para evitar uma grande perda de sangue. Se os embriões forem utilizados para análise, proceda à etapa 3.9. Se os filhotes forem adotados, prossiga para a etapa 2.8.
- Se for necessário adoção, limpe os filhotes antes de transferi-los para a mãe adotiva. Durante a limpeza dos filhotes, pressione suavemente o peito de vez em quando para iniciar a respiração. Coloque de volta em uma segunda luva cheia de água morna para melhorar a taxa de sobrevivência.
NOTA: É importante remover suavemente qualquer amônio e/ou placenta restantes com uma toalha de papel. - Antes de transferir filhotes para a mãe adotiva, remova a mãe adotiva de sua gaiola, remova os filhotes originais e substitua pelos filhotes experimentais. Devolva a mãe adotiva para sua gaiola.
NOTA: Consulte a discussão para obter sugestões adicionais para melhorar as taxas de aceitação de fomento. - Se for necessário genotipagem, colete biópsias entre P6−P8.
NOTA: Realize esta etapa somente se as licenças experimentais animais aprovarem essa prática.
3. Preparação tecidual para clones MADM no cérebro
NOTA: Para experimentos que incluem tecido pós-natal (≥P4), proceda à etapa 3.1. Para pontos de tempo embrionários e pós-natal precoce (P0−P3), continue a etapa 3.9.
- Anestesiar o animal MADM experimental com uma injeção IP de uma solução de cetamina/xilazina/acepromazina (65 mgs, 13 mgs e 2 mg/kg de peso corporal, respectivamente) e confirmar que o rato não responde por beliscar a pata traseira.
NOTA: Tanto os camundongos MADM masculinos quanto femininos (fundo CD-1) são usados para análise. Se for necessário genotipagem, colete biópsias de ouvido neste momento. - Coloque o animal anestesiado na posição supina na bandeja de cirurgia de perfusão e desinfete a pele com 70% de etanol. Para começar a cirurgia, faça cuidadosamente uma incisão com tesouras e fórceps cirúrgicos através da camada externa da pele e, em seguida, uma segunda incisão através da camada muscular. Levante a ponta do esterno e corte o tecido conjuntivo nas laterais, tomando cuidado extra para evitar o corte do fígado. A cavidade torácica será visível.
- Corte o diafragma e levante para revelar o coração. Corte cuidadosamente a caixa torácica e fixe na bandeja cirúrgica para expor o coração. Para filhotes, remova completamente a caixa torácica.
- Insira uma agulha com soro fisiológico tamponado de fosfato (PBS) no ventrículo inferior esquerdo (tecido mais pálido). O uso de uma pequena tesoura de íris faz uma incisão na extremidade posterior do átrio direito (tecido vermelho escuro) para o sangue drenar.
- Realizar perfusão com PBS seguida imediatamente por recém-feito, gelado 4% paraformaldeído (PFA) preparado em PBS. Para filhotes (P4−P10) use seringas para realizar perfusão. Encha uma seringa com 10 mL de PBS e outra com 10 mL de 4% pfa. Certifique-se de que todas as bolhas de ar nas seringas foram removidas. Para animais mais velhos, use uma bomba peristáltica.
- Comece a perfusar com PBS (10 mL a 2-4 mL/min em filhotes; 20 mL a 4-6 mL/min para adultos usando uma bomba peristáltica). O fígado ficará claro e amarelo pálido se a agulha estiver posicionada corretamente.
- Uma vez completa, remova a agulha dos filhotes e insira a agulha contendo PFA no mesmo orifício. Para adultos, pare a bomba peristáltica antes de trocar a solução PBS por PFA gelada, certificando-se de evitar bolhas na tubulação de absorção. Retomar a perfusão com PFA (10 mL a 2-4 mL/min em filhotes; 30 mL a 4-6 mL/min para adultos usando uma bomba peristáltica).
- Quando a perfusão estiver completa, decapite o rato e remova o cérebro através de dissecção cuidadosa. Transferir cérebro para 4% pfa. Use pelo menos 5x os volumes cerebrais (ou seja, ~5-10 mL de PFA em um tubo de centrífuga cônica de 15 mL) e incubar durante a noite a 4 °C para fixação pós-perfusão para garantir a fixação completa do tecido. Continue a passo 3.10.
- Para tecido embrionário e tecido pós-natal precoce (ou seja, P0−P3), após a realização de uma cesariana, decapite os embriões com tesouras. Se for necessário genotipagem, colete a cauda do embrião neste momento. Disseque imediatamente o cérebro e transfira para uma placa de 12 poços contendo 2-3 mL de 4% pfa/bem. Incubar durante a noite a 4 °C para pós-fixação.
- Na próxima manhã troque PFA com 10 mL (adulto) ou 2-3 mL (embrião) de PBS e repita 3x por 15 min na RT. Transfira tecido para solução de 30% de sacarose em tampão de fosfato (PB) e armazene a 4 °C em uma plataforma de balanço até que o tecido afunde na solução.
- Incorpore o cérebro em um composto de temperatura de corte ideal (OCT) em um molde de incorporação, tomando cuidado para orientar o cérebro para secção coronal ou sagital. Congele colocando o molde de incorporação em gelo seco até que o OCT se torne completamente opaco (~10-15 min). Armazene o tecido a -80 °C até que use mais.
4. Preparação do tecido MADM para a imunohistoquímica
- Conecte o bloco de tecido ao disco da amostra no criostat, aplicando um anel de OCT ao disco e colocando o bloco diretamente no OCT quando ele começar a congelar. Certifique-se de que o bloco está corretamente orientado para o plano de corte desejado.
NOTA: Aqui, a seção coronal para investigar clones cortical MADM é descrita em detalhes. - Coloque a temperatura do bloco no criostat para -20 °C e a temperatura da lâmina para -21 °C.
- Deixe que o bloco de tecido se ajuste à temperatura da câmara, montando o disco da amostra no suporte da amostra e deixe em criostat por ~5 minutos antes de iniciar a secção.
- Corte bloco em seções grossas (45-60 μm) até que a região tecidual de interesse seja atingida.
- Uma vez que a borda do córtex esteja claramente visível, pare de seção e bloqueie a lâmina. Certifique-se de que a lâmina está protegida antes de aparar o bloco.
- Apare o excesso de OCT ao redor do tecido com uma lâmina, deixando ~1-2 mm de OCT em todos os lados do cérebro.
- Em seguida oriente o bloco para que uma das bordas laterais do córtex seja orientada para baixo e a outra para cima (ou seja, a borda mais rostral do córtex é apontada para a direita).
- Comece a seção com uma espessura de 45 μm para clones adultos e 30 μm para clones embrionários. Execute cada seção individualmente e use uma pequena escova para manter a área abaixo da faca limpa de quaisquer detritos deixados enquanto apara o bloco.
NOTA: Se isso não for feito e uma seção cair, pode ser difícil determinar a ordem correta das fatias. - Se as seções começarem a enrolar, corte as bordas do bloco e/ou ajuste cuidadosamente a placa antiroll de vidro.
- Para análise de clones embrionários, monte seções diretamente para um slide fosco. Seque em uma placa de aquecimento a 37 °C antes de prosseguir diretamente para a etapa 5.6.
NOTA: Várias seções podem ser adicionadas a um slide, mas certifique-se de que a ordem sequencial seja mantida. - Para coletar clones adultos, prepare 24 placas de poço contendo 1 mL de PBS/well (tipicamente, 5-6 placas por cérebro). A partir do primeiro poço, com fórceps frios coletam seções seriais individuais na PBS na ordem de secção.
NOTA: O método de seção flutuante é adotado para tecido adulto para garantir que nenhuma seção seja perdida e que as seções montadas não contenham rugas. - Pare de se seccionar assim que o fim do neocórtex for atingido.
- Para clones adultos, proceda à montagem de seções flutuantes.
NOTA: As seções podem ser mantidas em PBS a 4 °C por até 24 h.
5. Montagem de tecido adulto para imagem
NOTA: São necessárias as seguintes ferramentas: pincel pequeno, placa de Petri, PBS com 0,5% Tween (PBS-T), lâminas de adesão(Tabela de Materiais),meio de montagem(Tabela de Materiais),tampas e fórceps.
- Encha uma placa de Petri com PBS-T.
NOTA: O detergente é usado para auxiliar no processo de montagem. Se a coloração de antígenos adicionais sensíveis a detergentes (ou seja, glicoproteínas) for necessária, é melhor pular a adição de Interpolação. - Coloque um slide de adesão no PBS-T para que ele esteja quase coberto até o rótulo.
- Transfira a primeira seção para o PBS-T.
- Usando uma pequena escova de tinta, manoira a seção sobre o escorregador e organize-a para preservar a ordem de corte. Prossiga da mesma forma com todas as outras seções.
- Uma vez que todas as seções estejam em posição, coloque o slide (~12−16 seções/slide) em uma câmara de slides escura. Levante ligeiramente a tampa para permitir que as seções sequem completamente (~10-20 min), garantindo que elas permaneçam aderidas nas etapas subsequentes.
- Se realizar a imunohistoquímica para antígenos adicionais, proceda diretamente à seção 6 ou 7.
NOTA: Para pontos de tempo embrionários, é necessário executar etapas de imunossuagem para pelo menos GFP e tdT (seção 6). Para clones adultos, isso só é necessário se a coloração de antígenos adicionais em paralelo (seções 6 e 7). - Reidratar e lavar seções 1x com 1x PBS por 5 minutos para remover PBS-T residuais. Aplique ~1 mL de 4′,6-diamidino-2-fenilôdole (DAPI) diluído em 1x PBS (1 μg/mL) no slide, garantindo que todas as seções estejam cobertas e incubadas por 15 minutos.
- Remova cuidadosamente o DAPI e lave 1x com 1x PBS por 5 min. Remova o excesso de PBS e seque por ~1-2 min antes de incorporar em 110 μL de meio de montagem. Veda com uma tampa de 24 x 60 mm e deixe secar por pelo menos 3 h antes da imagem.
6. Imunostaining apenas para GFP e TDT
NOTA: Esta seção é necessária para clones embrionários.
- Coloque slides horizontalmente em uma câmara de incubação de lâminas umidificadas. Marque limites de deslizamento com um marcador de cera para minimizar a quantidade de tampão necessária.
- Reidratar seções com 1x PBS. Para melhorar a qualidade da coloração, trabalhe com tecido recém-seccionado.
- Adicione 250-400 μL de tampão de bloqueio (0,5% Triton X-100, 2−3% soro normal de burro em 1x PBS) por slide, garantindo que todas as seções estejam cobertas. Incubar por 1h.
NOTA: A concentração de detergente (Triton X-100 ou Tween-20) vai variar dependendo dos anticorpos primários adicionais utilizados porque alguns antígenos são mais sensíveis a detergentes do que outros. - Remova o buffer de bloqueio e adicione anticorpos primários no buffer de bloqueio ao slide (300−400 μL/slide).
NOTA: Um exemplo de uma reação de anticorpos primários padrão para anti-GFP/anti-tdT (MADM) poderia usar frango anti-GFP (1:500) e anti-RFP coelho (1:500). - Incubar com anticorpos primários durante a noite a 4 °C.
NOTA: Os slides devem ser incubados perfeitamente horizontalmente com tampão cobrindo todas as seções. Caso contrário, a coloração desigual ou ruim pode resultar. - Confirme na manhã seguinte que o tampão de bloqueio com anticorpos primários ainda cobre todas as seções no slide. Se não, repita a etapa de incubação por 3-4 h no RT.
- Remova os anticorpos primários e lave 4x com 1x PBS por 10 minutos no RT.
- Adicione anticorpos secundários diluídos no buffer de bloqueio para deslizar (300−400 μL/slide): Alexa Fluor 488 anti-frango IgG (1:500) e Cy3 anti-coelho IgG (1:500).
- Incubar em RT por 2 h. Mantenha slides cobertos de luz para evitar branqueamento de fluoroforos.
- Remova os anticorpos secundários e lave 2x com 1x PBS por 10 minutos.
- Incubar com DAPI diluído em PBS (1:5.000) por 15 min.
- Lave as seções 1x com 1x PBS por 10 minutos.
- Remova o excesso de PBS e seque por ~1-2 min antes de incorporar em 110 μL de meio de montagem.
- Vedar com tampa de 24 x 60 mm e deixe secar por pelo menos 3 h antes da imagem. A imagem desliza dentro de 1-2 semanas após a realização de imunohistoquímica para garantir o sinal ideal.
7. Imunostaining para GFP, tdT e antígenos adicionais
- Realize as etapas 6.1-6.3.
- Remova o buffer de bloqueio e adicione anticorpos primários no buffer de bloqueio ao slide (300−400 μL/slide).
NOTA: Ao colorir para três ou mais antígenos (ou seja, GFP, tdT e uma proteína de interesse) e o anticorpo para a proteína de interesse foi levantado em coelho, recomenda-se usar o anticorpo primário anti-tdT (cabra) a uma diluição de 1:500. Um exemplo de reação primária de anticorpos para três antígenos com coloração alternativa de TDT poderia usar frango anti-GFP (1:500), anti-tdT de cabra (1:500) e anticorpo contra a proteína de interesse (ou seja, coelho). - Realize as etapas 6.5-6.7.
- Adicione uma mistura secundária de anticorpos diluída no tampão de bloqueio para deslizar (300−400 μL/slide): Alexa Fluor 488 anti-frango IgG (1:500), Cy3 anti-cabra IgG (1:500) e Alexa Fluor 647 anti-coelho IgG (1:500).
- Realize as etapas 6.9-6.14.
8. Aquisição de imagem confocal e quantificação de clones MADM
- Identificar e documentar seções cerebrais contendo clones e suas localizações no córtex.
NOTA: O número de seções que um clone abrange varia de acordo com quando o clone foi induzido, o driver CreERT2 e o tempo de análise. Esta etapa pode ser realizada em um microscópio confocal ou em um microscópio de epifluorescência. - Usando um microscópio confocal invertido, comece selecionando e configurando as linhas e filtros laser corretos. Para cérebros MADM, selecione DAPI, GFP e tdT (excitação: 358 nm, 488 nm e 554 nm, respectivamente; emissão máxima: 461 nm, 507 nm e 581 nm, respectivamente). Certifique-se de que o orifício está configurado para 1 unidade arejada para a melhor qualidade de imagem.
- Para configurações específicas de confocal, clones de imagem com um objetivo de 20x e zoom de 1x. Para imagens que serão usadas em quantificações, use um valor de imagem de velocidade de varredura de 1,52-2,06 μs (valores 7-8 no software de aquisição de imagens) sem média. Ajuste a intensidade do laser e obtenha as configurações para cada canal conforme apropriado.
NOTA: Dependendo da qualidade de imagem necessária, as configurações para a velocidade de varredura e a média podem variar. - Uma vez que o clone seja claramente identificado, organize as telhas de imagem para cobrir todas as seções relevantes do clone. Ajuste a pilha z para que todas as células rotuladas MADM no clone sejam capturadas com um intervalo de fatia de pilha de 1,5 μm/z. Ajuste a região de ladrilhos para que toda a largura do córtex seja capturada ao fotografar o clone (ou seja, da superfície pial ao caloso corpus).
- Os clones individuais da imagem abrangem várias seções consecutivamente, garantindo que quaisquer seções sem células dentro de um clone ainda sejam imagens com o propósito de reconstrução 3D e interpretação correta das informações espaciais celulares.
- Analise cada seção contendo células de um clone MADM sequencialmente do rostral até a extremidade caudal do córtex. Distinguir neurônios individuais e glia com base em sua morfologia e/ou coloração de marcadores. Registos posicionais em paralelo com base em respectivas fronteiras de camada definidas pela coloração nuclear (DAPI).
NOTA: Consulte a Figura 4 para resultados representativos para análise embrionária e Figura 5 para resultados representativos para análise adulta.
9. Reconstrução serial 3D de clones
NOTA: A reconstrução 3D de clones individuais imagens sobre seções cerebrais seriais é útil para exibição visual, bem como para a análise de arquiteturas clonais 3D e pode ser realizada de acordo com as seguintes etapas.
- Costura e fusível imagens de ladrilhos confocal com base em parâmetros de aquisição usando software de aquisição de imagens. Abra o arquivo .czi e execute o método Costurar na guia Processamento no software ZEN (Zeiss).
- Exportar pilhas de imagens costuradas como z-planes individuais no formato TIFF. Abra o arquivo .czi costurado e, em seguida, execute o método de exportação de imagens na guia Processamento. Para imagens multicanais, exporte como imagens vermelhas/verdes/azuis para processamento subsequente de imagens.
- Repita as etapas 9.1 e 9.2 para cada seção cerebral serial de um clone.
NOTA: Para uma reconstrução 3D precisa, todas as seções cerebrais dentro de um clone, incluindo as sem células rotuladas, também devem ser processadas. - Concatenar imagens individuais em uma única pilha, a partir do mais rostral ao mais caudal z-plane usando software de processamento de imagem de código aberto, como ImageJ/Fiji67,68.
NOTA: Qualquer imagem em branco nas bordas de cada seção cerebral deve ser removida neste momento. - Se necessário, corrija a pilha de imagens obtida da etapa 9.4 para desalinhamento usando um plugin ImageJ chamado "MultiStackReg" seguindo as etapas 9.5.1−9.5.5. Se o alinhamento da imagem não for necessário, proceda à etapa 9.6.
NOTA: Este plugin executa o alinhamento de imagem do canal com maior contraste (geralmente DAPI) e, em seguida, aplica a transformação gravada para os outros canais, permitindo assim um alinhamento confiável de imagem de pilhas multicanais. Um plugin auxiliar chamado "TurboReg" deve ser pré-instalado.- No ImageJ, instale os plugins "MultiStackReg" e "TurboReg".
- Abra a pilha de imagens de imagens de clone obtidas a partir da etapa 9.4 para ser alinhada. Divida os canais em DAPI (azul), GFP (verde) e tdT (vermelho) na opção Color em aba Imagem.
- Execute o plugin" MultiStackReg" para alinhar o canal DAPI por "Rigid Body" e salvar o arquivo de transformação.
- Aplique o arquivo de transformação salvo nos outros dois canais usando "MultiStackReg".
- Mescle todos os três canais alinhados e salve a pilha alinhada.
- Para orientar o clone no ImageJ gire pilha de imagens clones obtidas a partir da etapa 9.4 (ou passo 9.5.5 após o alinhamento) em uma orientação vertical com a superfície pial na parte superior e o caloso corpus na parte inferior. Crop no plano xim, se necessário.
- Tanto para apresentação qualitativa quanto para análise quantitativa, gere uma imagem máxima de projeção z (etapa 9.8) ou realize renderização 3D (etapa 9.9) do clone.
- No ImageJ, abra a pilha de imagens a partir do passo 9.6 e selecione opção de projeção Z com intensidade tipo de projeção Max. Isso vai gerar uma imagem de todo o clone projetado no mesmo plano.
- No ImageJ, abra a pilha de imagens da etapa 9.6 e selecione a função z do Projeto 3D para gerar uma visualização 3D do clone que pode ser girada.
NOTA: É importante nesta etapa inserir o intervalo de fatia correto equivalente à espessura das pilhas z individuais durante a aquisição de imagens. A ferramenta de interpolação deve ser usada para remover lacunas entre fatias.
Resultados
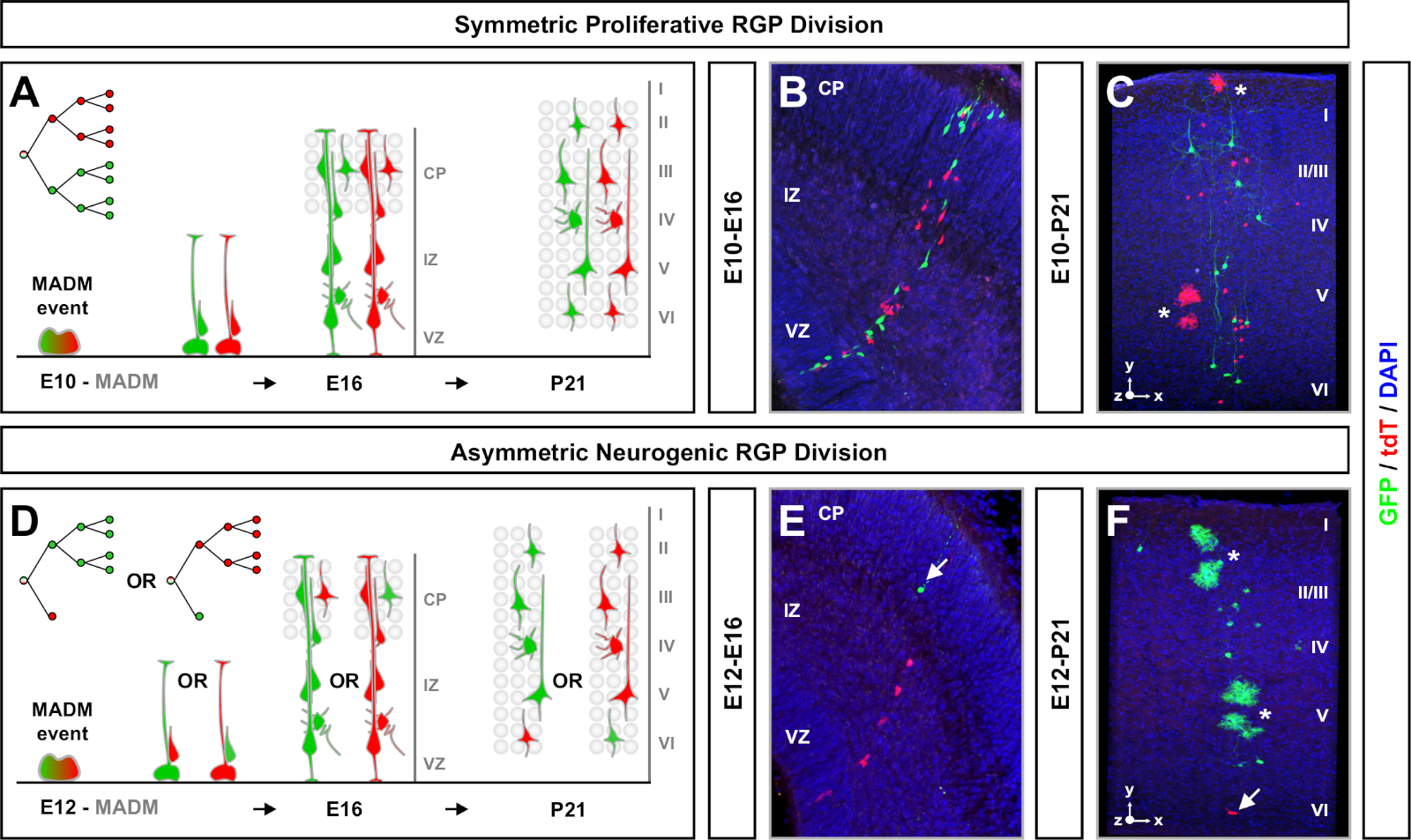
Madm resulta na reconstituição de proteínas fluorescentes verdes e vermelhas funcionais com duas células filhas cada uma expressando uma das duas proteínas fluorescentes em eventos de segregação do cromossomo G2-X(Figura 1A). Como os eventos MADM resultam em rotulagem permanente e distinta das duas linhagens descendentes, pode ser realizada avaliação quantificável de linhagens celulares verdes e vermelhas (subclones). Variáveis incluindo padrão de divisão (por exemplo, simétrica versus assimétrica) e potencial (por exemplo, o número de progenia) do progenitor original podem ser determinadas. Quantificar cada subclone fluorescente é informativo quando determina retroativamente se a célula progenitora original está passando por divisões proliferativas simétricas, ou divisões neurogênicas assimétricas no momento da indução de TM. Estudos anteriores agruparam clones de projeção excitatória derivados em Emx1-CreERT2 ou Nestin-CreERT2 no córtex em duas classes amplas7,,11,46. Os primeiros, denominados "clones proliferativos simétricos", são compostos em média por um número considerável de neurônios, com subclonas verdes e vermelhas contendo quatro ou mais neurônios cada. O segundo grupo, "clones assimétricos" define uma classe de clones onde o subclone "minoritário" contém menos de três neurônios e o subclone "maioria", quatro ou mais11. Essas definições são específicas para RGPs corticais e podem precisar ser revisitadas para outras regiões cerebrais e tecidos. Para ambas as classes de clones corticais, a prole será distribuída por todas as camadas superficiais e profundas.
Ao projetar estudos clonais madm há uma série de aspectos que devem ser levados em consideração. O momento em que os eventos MADM são induzidos pela administração do TM é uma consideração fundamental(Figura 3). Para os clones madm neurônio de projeção excitatória cortical (ou seja, usando Emx1-CreERT2 ou Nestin-CreERT2) no E10, quase todos os RGPs ainda estavam passando por divisões simétricas11. Portanto, a indução no E10 com TM capturou várias rodadas de amplificação de RGP proliferativa e resultou em clones com alto número de neurônios. No entanto, o número de RGPs no E10 era geralmente pequeno e, portanto, a administração de TM gerou muito poucos eventos MADM (às vezes menos de um por cérebro). A maioria dos RGPs mudou de divisões neurogênicas simétricas para assimétricas em torno de E12. Para atingir clones neurogênicos estritamente assimétricos, era melhor induzir em E12 ou posterior(Figura 3). O tempo entre a indução de TM e a observação de eventos de recombinação madm no córtex tendia a ser inferior a 24 h. As injeções de IP eram o método preferido para administrar TM em estágios embrionários para este método, pois levou a uma maior reprodutibilidade na indução clonal. Também é importante manter a dose TM no mínimo por duas razões. Primeiro, se a taxa de recombinação madm aumenta, a probabilidade de induzir múltiplos, talvez sobrepostos, clones é maior. Em segundo lugar, se for entregue muito TM, uma taxa aumentada de aborto, reabsorção de embriões e tamanhos menores de lixo podem ser observados. Abortos em aproximadamente metade de todas as barragens grávidas foram observados quando as injeções de TM foram entregues no E10. Essa frequência diminuiu a partir do E11 em diante e diminuiu para aproximadamente 1/3 das barragens grávidas abortando. Para um resumo das doses de TM, tempos de indução e drivers CreERT2 utilizados em estudos madm anteriores, consulte a Tabela 1. A atividade do repórter na ausência de TM foi observada com alguns drivers CreERT2 indutíveis de TM69. Eventos de recombinação de expressão ectópica ou MADM na ausência de TM não foram observados com o Emx1-CreERT2 dos drivers Nestin-CreERT2. Isso pode ser parcialmente devido ao fato de que as recombinações cromossômicas mediadas por TM ocorrem aproximadamente 1:1.000 a 1:10.000 menor frequência do que as recombinações cis-recombinações, reduzindo a probabilidade de rotulagem MADM ectópica.
Outro fator a considerar ao planejar um experimento de análise clonal madm é a duração do estudo. Variando o tempo entre a indução de TM e quando o experimento foi analisado (A) (janela de tempo) exibe dinâmica de células-tronco ao longo do tempo64. Janelas de tempo embrionárias curtas (i.e., TM/E11−A/E13; TM/E11−A/E16) capturou a dinâmica da neurogênese embrionária(Figura 4). Comparar clones de duas ou mais janelas de tempo fornece uma visão quantitativa sobre o número de células produzidas e como a distribuição de neurônios varia em diferentes estágios de progressão de linhagem64. Para capturar todo o potencial dos clones individuais, é necessário estender a janela de tempo analisada em pontos de tempo pós-natal ou adultos7,,11,,12. Exemplos de clones neocorticais induzidos no embrião e analisados no adulto são mostrados na Figura 5. Note-se que a neurogênese cortical é principalmente completa e a gliogênese aumenta em E17. Aproximadamente 1/6 RGP neurogênico também prossegue para a geração de astrócitos e/ou oligodenrócitos11.
Clones simétricos ocorrem quando os RGPs passam por uma ou mais rodadas de divisãoproliferativa 11. Os clones de RGP induzidos entre E10-E12 foram, em média, maiores em tamanho e forneceram mais características espaciais da distribuição final dos neurônios(Figura 4A-C). Clones com neurônios relativamente distribuídos em camadas profundas e superficiais assumiram uma forma de "cilindro", enquanto clones com neurônios mais dispersos em camadas superficiais do que camadas mais profundas desenvolveram uma forma "cone"11. Para capturar totalmente as informações espaciais e morfológicas de um clone, foi necessário reconstruir computacionalmente cada clone usando imagens sequenciais. Para medir a dispersão clonal, a dispersão lateral máxima (medida em todas as dimensões) em camadas superficiais (LII-VI) de um clone foi comparada à dispersão de neurônios em camadas profundas (LV/LIV). Esta razão (distribuição superior:distribuição inferior) forneceu uma leitura quantificável da forma geral do clone.
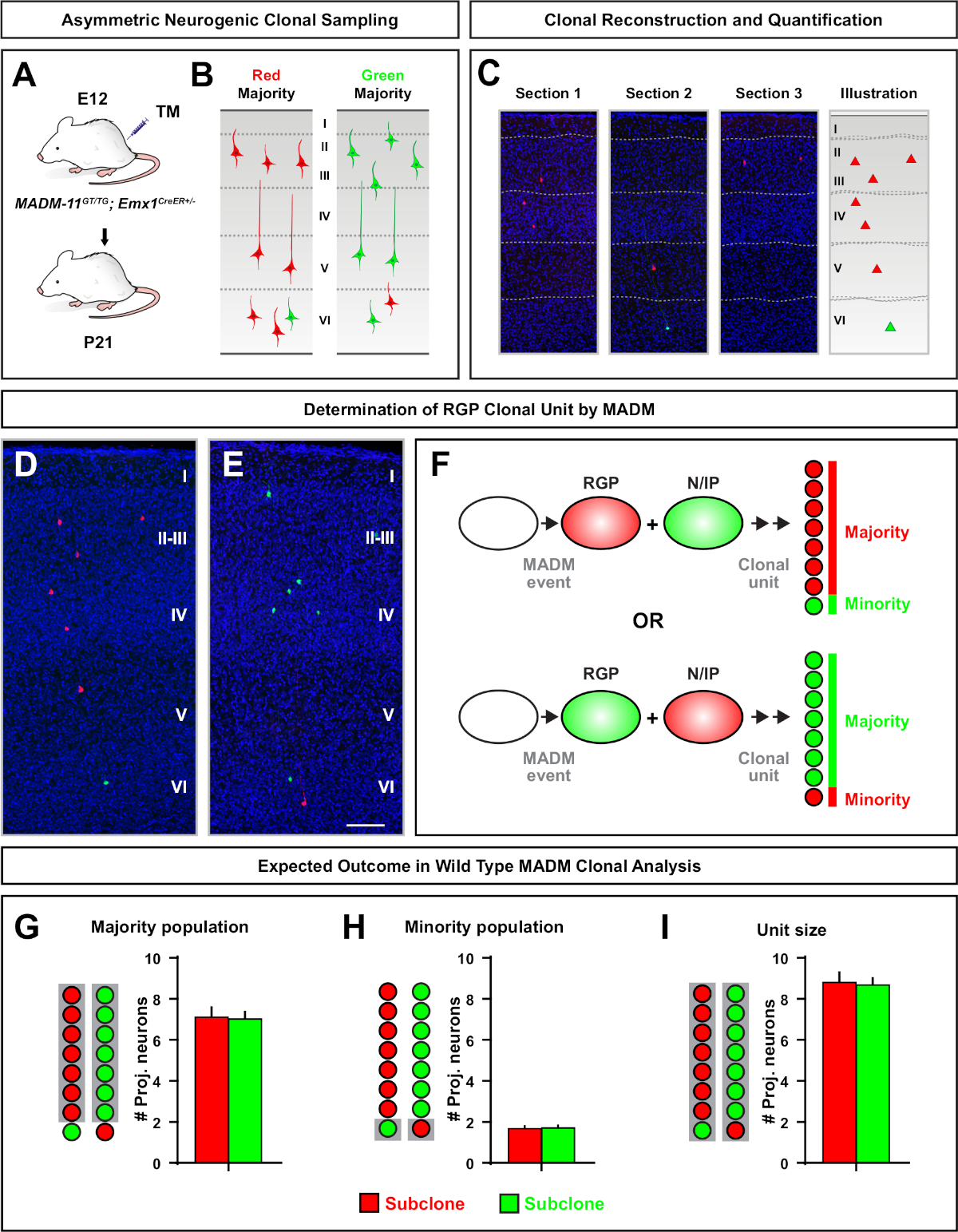
Clones assimétricos, onde o subclone minoritário era de três ou menos, forneceram uma visão da saída neuronal de um único RGP (Figura 4D-F e Figura 5A-F)7,,11,,12. A população majoritária (subclona grande) poderia ser rotulada em vermelho ou verde, com uma média de aproximadamente sete neurônios de projeção excitatória por clone quando induzidas usando um Emx1-CreERT2 ou Nestin-CreERT2(Figura 5G)7,,11,12. O número total de células em um clone MADM poderia ser ainda mais dissecado analisando a distribuição de neurônios no grande subclone através de camadas superficiais e profundas. A população minoritária (pequeno subclone) foi rotulada pela cor recíproca e foi, em média, 1-2 células por clone(Figura 5H). O "tamanho da unidade" total, que era em média 8-9 neurônios, poderia ser calculado adicionando os subclones pequenos e grandes(Figura 5I)7,,11,12. É importante notar que, embora a saída neuronal das RGPs fosse altamente previsível, havia um grau de heterogeneidade clonal12,70.
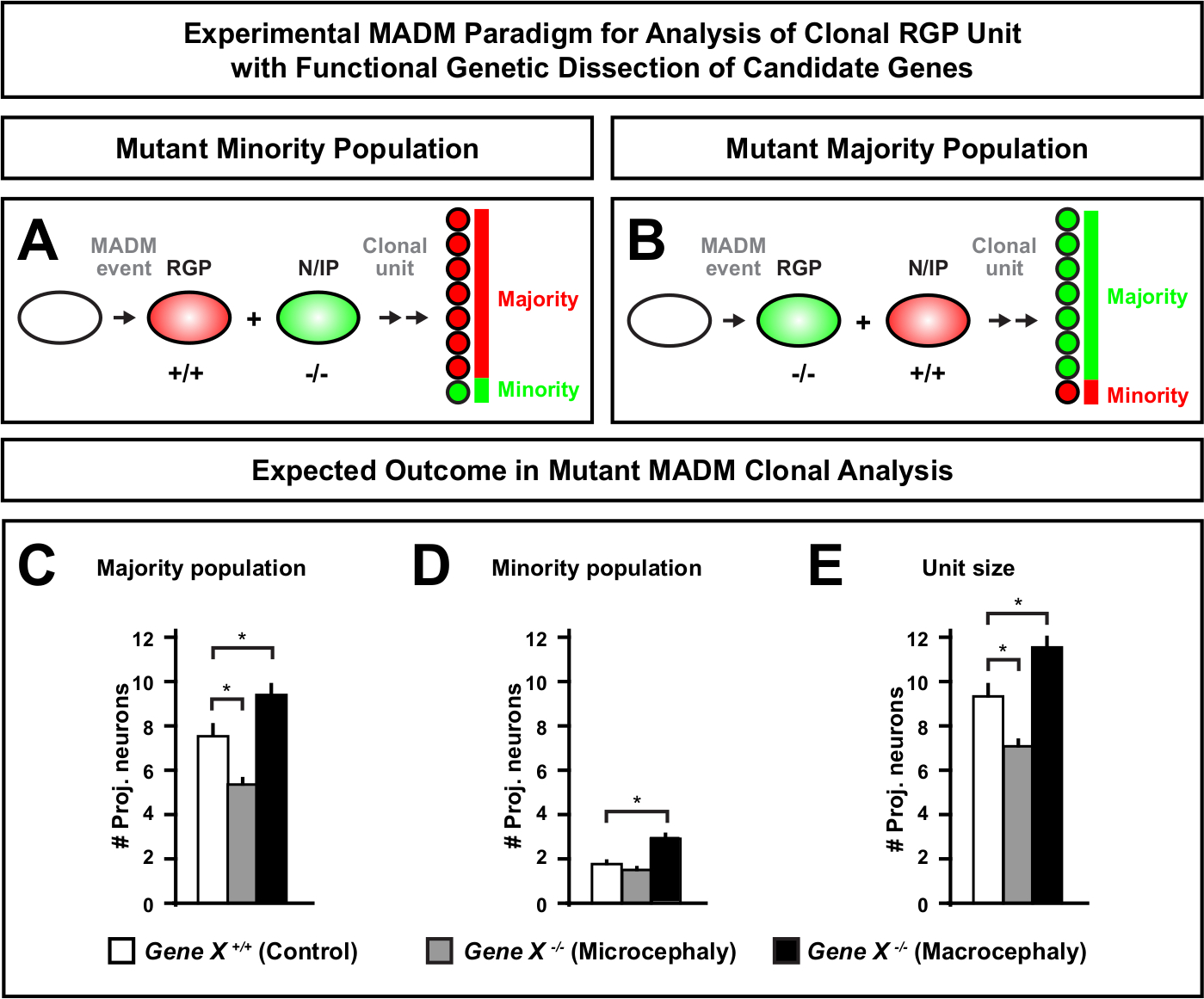
A introdução de uma mutação distal ao MADM permite a geração de mosaicos genéticos, fornecendo um método único para dissecar os reguladores moleculares da progressão da linhagem de células-tronco. Como tal, o MADM fornece uma plataforma experimental incomparável para estudar a função célula-autônoma de um gene (por exemplo, sua associação à microcefalia ou à macrocefalia). Comparando clones induzidos em um mosaico genético MADM a clones induzidos em um controle MADM, uma leitura altamente quantitativa de mudanças no número de neurônios e distribuição pode ser gerada. Estudos anteriores baseados em MADM quantificaram a função célula-autônoma de Otx1 na formação de microcefalia no nível clonal (ver Figura 6A-E para um exemplo representativo)11. Em outro estudo, a análise clonal do MADM demonstrou que o Ndel1 não regula de forma autônoma os números de neurônios de projeção, mas sim a capacidade dos neurônios recém-nascidos de entrar ou migrar dentro da placa cortical, que mais tarde forma o córtex adulto46. Esses estudos demonstraram a natureza altamente quantitativa da análise clonal madm no estudo das funções célula-autônomas dos genes que regulam o desenvolvimento cortical. Atualmente, não há exemplos na literatura usando o MADM para estudar genes implicados na macrocefalia no nível clonal. No entanto, em estudos futuros, a análise de genes relevantes para o controle do tamanho cortical em geral pode fornecer insights altamente desejáveis a nível molecular e celular.

Figura 1: O princípio MADM para rastreamento de linhagem e análise clonal a nível de células-tronco únicas. (A) Para realizar o rastreamento de linhagem e análise clonal com MADM, dois componentes devem estar presentes. Primeiro, as fitas MADM devem ser direcionadas a loci idênticos em cromossomos homólogos. As consistem em dois genes de repórteres fluorescentes quiméricos, eGFP (verde, [G]) e tandem dimer Tomate (vermelho, tdT[T]). O GT contém o N-terminus de eGFP e o C-terminus de tdT, separado por um intron contendo um site loxP. O TG é construído inversamente, com o N-terminus de tdT e o C-terminus do eGFP. Em segundo lugar, a expressão de Cre recombinase deve ocorrer na célula contendo as fitas MADM direcionadas. Os sites loxP servem como um alvo para a recombinação intercromossômica mediada por Cre, resultando na reconstituição de ambas as fitas de expressão simultaneamente. Se a recombinação ocorrer durante a fase G2 do ciclo celular seguido pela segregação X (G2-X), as duas células filhas expressarão uma das duas proteínas fluorescentes. (B) Princípio MADM para análise de mosaico genético em um único nível de clone. Alelos mutantes (mutações de ponto, exclusões, inserções, alelos condicionales flanqueados por loxP como descrito na Figura 1B, etc.) podem ser introduzidos distal à fita TG-MADM via recombinação meiotic (ver Figura 2 e Hippenmeyer et al.46 para obter detalhes sobre como introduzir alelos mutantes no sistema MADM). Se uma recombinação trans intercromossômica mediada por G2-X Cre ocorrer entre MADM resulta em uma célula mutante homozigosa GFP+(GeneX-/-) para o gene de interesse e uma célula tipo selvagem homozigosa tdT+(GeneX+/+) em um ambiente heterozygous não rotulado46,47,71. Os desfechos de rotulagem alternativos não utilizados na análise clonal (ou seja, células amarelas) foram descritos anteriormente no detalhe11,46,47. Clique aqui para ver uma versão maior desta figura.
{kind=link}

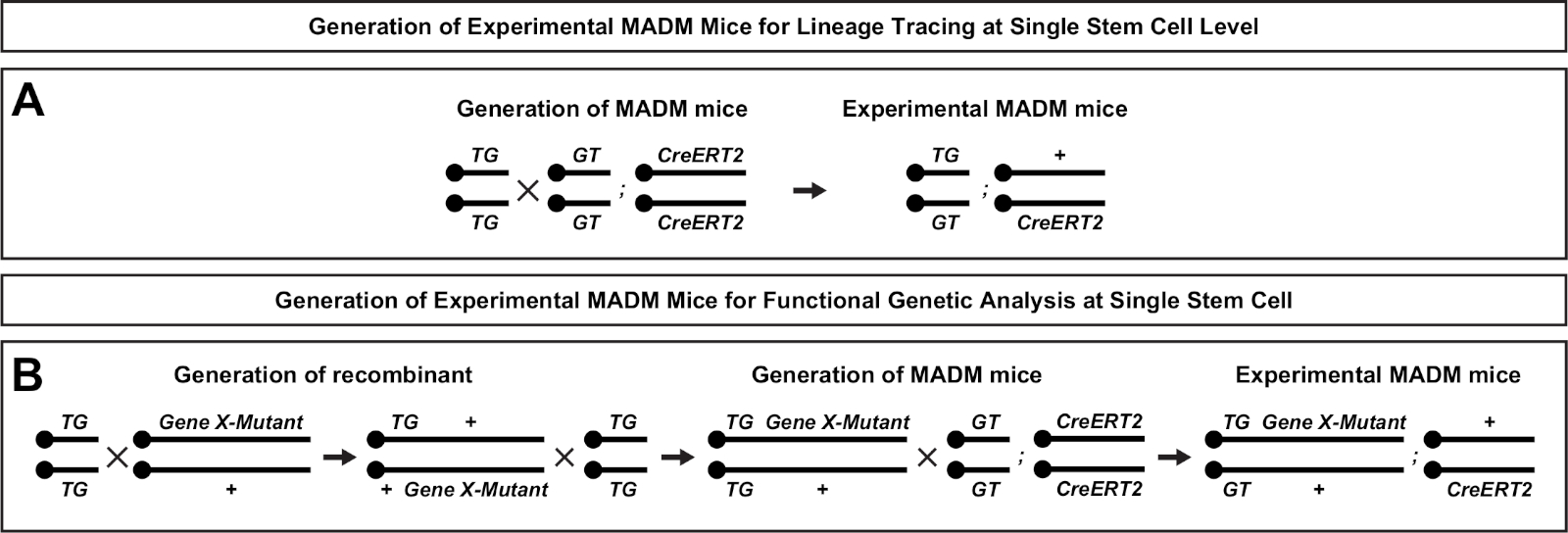
Figura 2: Esquemas de reprodução para geração de camundongos MADM experimentais para rastreamento de linhagem. Esquema de reprodução para a geração de controle MADM (A) e Gene X MADM(B)camundongos madm experimentais para análise clonal. Para obter mais informações sobre paradigmas de reprodução do MADM, consulte Beattie et al.7 e Hippenmeyer et al.7,46. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Paradigmas do curso de tempo para análise de linhagem clonal baseada em MADM. Esquema das janelas de tempo de design experimental. Para paradigmas de amostragem longitudinal, o ponto de tempo da indução de clones permaneceu constante e o tempo antes da análise variou. Na amostragem progressiva do intervalo, o ponto de tempo da análise permaneceu constante, mas o tempo de indução variou. Uma combinação de uma ou ambas as abordagens pode ser usada dependendo das perguntas abordadas. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Análise clonal MADM no neocórtex em desenvolvimento e adulto. Indução de clone MADM mediado em TM em RGPs simetricamente proliferativos (TM at E10)(A-C)e assimétricamente neurogênicos (TM em E12) (D-F)dividindo RGPs. São retratados clones madm individuais in vivo no desenvolvimento (TM/E10−A/E16 e TM/E12−A/E16)(B,E) e adultos (TM/E10−A/P21 e TM/E12−A/P21)(C,F) em MADM-11GT/TG; Nestin-CreERT2+/- (B,E) e MADM-11GT/TG; Emx1-CreERT2+/- (C,F). A saída de neurônios era independente da cor subclone e subclonas de maioria verde/minoria poderiam ser comparadas com subclones de maioria vermelha/minoria sob condições de controle7,,11. Aproximadamente 1/6 dos clones adultos também continham astrócitos e/ou oligodenrócitos, indicados por asteriscos brancos. Os painéis B e F são reproduzidos com permissão de Hippenmeyer et al.46 e Rulands e Simons72, respectivamente. CP = Placa cortical. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: Análise clonal MADM para quantificar a saída de neurônio mediada por RGP. Análise da produção de neurônios excitatórios (unidade) por RGPs neurogênicos individuais no nível clonal utilizando MADM7,11. (A) Paradigma experimental para induzir principalmente clones MADM assimétricos no córtex em desenvolvimento. (B) Possíveis desfechos de clones assimétricos com a subclona majoritária rotulada em seções verdes ou vermelhas(C) Representativos consecutivos abrangendo um único clone assimétrico neurogênico(D,E) imagens de reconstrução 3D de clones assimétricos representativos G2-X MADM com população majoritária em vermelho (D) ou verde(E)em MADM-11GT/TG; Emx1-CreERT2+/- com indução de TM no E12 e análise em P21. Note que as células verde e vermelha são do tipo selvagem. (F) Esquema indicando os dois possíveis resultados experimentais de clones MADM. (G) Quantificação do tamanho da população majoritária decorrente da renovação de RGPs em clones MADM-11. (H) Quantificação do tamanho da população minoritária decorrente da renovação de RGPs em clones MADM-11. (I) Quantificação do tamanho unitário de clones MADM-11 neurogênicos assimétricos. Valores hipotéticos podem representar média ± SEM. Barra de escala = 100 μm(D e E). TM = Tamoxifen. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6: Análise clonal madm para estudar genes que levam à microcefalia e à macrocefalia. A análise clonal hipotética madm resulta ao realizar dissecção genética funcional de genes candidatos que levam à microcefalia ou macrocefalia. Para dissecar as funções célula-autônomas de um gene de interesse (Gene X) na saída de neurônios, madm requer alelos mutantes para serem introduzidos distal às fitas MADM via recombinação meiotic (para detalhes como introduzir alelos mutantes no sistema MADM ver também Figura 2, Hippenmeyer et al.46, e Laukoter et al.46,73). (A,B) Esquema indicando paradigma EXPERIMENTAL MADM para análise funcional de unidades clonais de RGP. O subclone mutante pode formar a população minoritária (A) ou a maioria(B). (C-E) A análise clonal hipotética MADM resulta ao quantificar o controle MADM (barras brancas), a microcefalia Gene-X MADM (barras cinza) e os clones assimétricos da macrocefalia Gene-X MADM. (C) Quantificação do tamanho da população majoritária. (D) Quantificação do tamanho da população minoritária. (E) Quantificação do tamanho unitário de clones neurogênicos assimétricos. Valores hipotéticos podem representar um cenário médio ± SEM. S = Hipotético onde a diferença no número de células subclonas poderia atingir significância, em relação ao controle. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Tabela 1: Estudos clonais madm na literatura. Resumo de estudos na literatura contendo experimentos de linhagem clonal MADM, incluindo driver CreERT2 usado, dose TM e tempo de injeção. Clique aqui para ver esta tabela (Clique com o botão direito do mouse para baixar).
Discussão
Um método para usar o MADM para rastrear a linhagem celular de RGPs individuais in vivo no neocórtex em desenvolvimento é descrito. Quando combinados com o CreERT2indutível TM, os eventos MADM podem ser precisamente cronometrados, fornecendo uma leitura visual altamente qualitativa e quantitativa dos padrões de divisão de células-tronco no nível de célula única. Ao titular a dose de TM entregue, em uma situação ideal pode-se obter uma média de menos de um clone por hemisfério cortical, proporcionando separação espacial adequada para distinguir inequivocamente clones individuais. Ao manter a integridade tecidual, este método também captura informações essenciais sobre posição, morfologia e números de células absolutas. MADM em Chr. 117,11,12,46,56,57, em Chr. 751, e o MADM original em Rosa2647,53,59 foram utilizados em estudos de análise clonal MADM. A alta resolução de células individuais fornece uma visão sem precedentes sobre a morfologia e a relação clonal das células filhas e permite a imagem viva de células-tronco proliferando e clones emergentes46,,52.
A cesariana e o acolhimento de filhotes para análise de clones em momentos pós-natal é um passo necessário e crítico no protocolo. Dependendo do estado de saúde da barragem gestante tratada com TM, pode não ser necessário realizar uma cesariana. No entanto, criar os filhotes com uma mãe adotiva ainda é necessário, porque a mãe tratada com TM pode ter problemas para lactar. Não foram observadas diferenças na necessidade de adoção com diferentes drivers CreERT2. Ambas as linhas MADM e mães adotivas são mantidas em um fundo de CD-1 de raça. Se a cesariana não for necessária, a represa grávida tratada com TM usada para gerar filhotes experimentais pode ser reutilizada para reproduções experimentais adicionais de acordo com os princípios do 3R (note que isso só pode ser feito se as licenças experimentais de animais aprovarem essa prática). Mães adotivas podem ser usadas para adoção de filhotes dentro de 2 dias após o parto, mas maiores taxas de sucesso têm sido observadas quando mães adotivas dão à luz no mesmo dia que os camundongos experimentais que devem ser adotados. Por isso, é importante configurar acasalamentos cronometrados para mães adotivas paralelamente aos acasalamentos experimentais na etapa 1.1. Manter um número de ninhada semelhante ao da ninhada original da mãe adotiva pode melhorar a taxa de sobrevivência dos filhotes adotivos e, portanto, a remoção de alguns para toda a ninhada original pode ser necessária. Etapas adicionais que podem melhorar o acolhimento incluem esfregar as luvas do experimentador com lixo e alimentos (para remover o cheiro das luvas); esfregando os filhotes suavemente após a cesariana com fragmentos da ninhada e ninho sujo da mãe adotiva; e colocação dos filhotes em contato próximo com os filhotes da mãe adotiva antes de sua colocação na gaiola do rato adotivo.
Como em outros métodos de rastreamento de linhagem baseados em repórteres, deve-se tomar uma consideração cuidadosa ao escolher o driver CreERT2 ideal para experimentos clonais MADM. Em primeiro lugar, o promotor utilizado deve expressar a recombina tanto temporal quanto espacialmente na população progenitora de interesse. Encontrar esse promotor pode ser um desafio, pois alguns promotores podem mudar padrões de expressão ou ficar silenciados em diferentes estágios de desenvolvimento. Para melhorar a especificidade do tipo de célula, várias recombinases específicas do local, cada uma impulsionada por promotores separados, foram usadas. Quando uma ou ambas as recombinases são expressas na mesma célula, isso rotula a célula e sua descendência com um repórter fluorescente74,75,76,77. Em resumo, é importante escolher um driver CreERT2 específico para a população de progenitores que está sendo analisado.
O passo mais crítico neste método é a identificação de um clone, pois todas as células devem ser derivadas inequivocamente de um único evento de recombinação (etapa 8.1). A titulação da concentração de TM garante menos de um aglomerado de células vermelhas/verdes por hemisfério cerebral e maximiza a probabilidade de analisar um único clone (passo 2.2)7,11. Os clones devem ser descartados se os aglomerados vizinhos de células ocorrerem dentro de 500 μm do clone de interesse. Portanto, é importante examinar várias seções antes e depois do aparecimento de um clone para garantir que não haja eventos adicionais de recombinação nas proximidades. Devido ao sinal mais fraco dos fluoroforos, é necessário realizar imunohistoquímica para eGFP e tdT em clones embrionários (ver seção 6). Isso só é recomendado em clones adultos se antígenos adicionais forem colabeled. Ao fotografar clones, é importante capturar toda a largura do córtex onde o clone está localizado (ou seja, da superfície pial ao corpo caloso; ver o passo 8.4) para não perder nenhuma célula. Isso também facilita o alinhamento da imagem durante o processamento da imagem (seção 9). A seção 8 do protocolo requer um microscópio confocal invertido, mas pode ser adaptada dependendo da configuração do microscópio disponível. A microscopia de epifluorescência pode ser usada, mas a microscopia confocal é recomendada porque isso leva a uma diminuição na contaminação da luz de fora do plano focal. Também é importante que a intensidade e o ganho do laser sejam ajustados para que as células verdes, vermelhas e amarelas possam ser identificadas inequivocamente. Independentemente da configuração, recomenda-se usar um objetivo de pelo menos 20x para garantir a separação espacial completa de células posicionadas de perto. Além de registrar a profundidade cortical de todas as células (passo 8.6), regiões corticais onde os clones estão localizados devem ser identificadas usando um atlas cerebral, como o atlas do Cérebro de Allen ou outros mapas de coordenadas estereotipadas. Um paradigma de nomeação de arquivos também deve ser adotado para garantir que as imagens de clone sejam facilmente identificáveis. As seguintes informações podem ser incluídas na nomeação do arquivo: identificação de imagem única, imagem de data foi tirada, genótipo de animal, idade de indução, idade de análise, número de imagem em relação ao resto das imagens do mesmo clone.
A introdução de uma mutação distal a uma fita MADM permite distintamente a geração de mosaicos genéticos71 e permite a dissecção de reguladores moleculares de linhagem e diversidade de tipos celulares no nível clonal7,,11,,46,62. Para gerar um mosaico genético com MADM, as fitas MADM devem estar meioticamente ligadas ao mesmo cromossomo que o gene de interesse (ver Figura 2 para o esquema de reprodução). Isso limita a análise clonal atual com MADM aos genes localizados em Chr. 751, Chr. 1146, Chr. 1251, e Chr. 6 distal ao lócus Rosa26 47. Estudos futuros usarão fitas MADM direcionadas a qualquer cromossomo, permitindo a análise do mosaico de praticamente todos os genes do genoma do camundongo no nível clonal.
Finalmente, o MADM não se limita à análise de células progenitoras no neocórtex em desenvolvimento. O estudo de muitos nichos de células-tronco poderia se beneficiar da capacidade de resolver arranjos espáteis de células clonalmente relacionadas. Aplicando MADM a outras regiões do cérebro, condições da doença (por exemplo, câncer), ou em outros tecidos47,50,51,52,53,54,55,56,57,58,59, estudos revelaram relações de linhagem em clones derivados de diversas classes de progenitor e células-tronco (ver Tabela 1 para lista atual de estudos clonais MADM). Outra aplicação futura interessante do MADM é combiná-lo com repórteres funcionais ou subcelulares adicionais, o que aumentaria o grau de informação que pode ser adquirida a partir de clones.
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Agradecemos a todos os membros do laboratório de Hippenmeyer pela discussão, o Centro de Bioimagem, a Instalação de Ciência da Vida e o Centro Pré-Clínico da IST Austria pelo apoio técnico. Este trabalho foi apoiado por fundos institucionais da IST Austria; R.B. recebeu apoio do programa Lise-Meitner (M 2416) do Fundo Austríaco de Ciência (FWF); N.A recebeu apoio do Fundo Austríaco de Ciência (FWF) Firnberg-Programm (T 1031); A GC recebeu apoio do programa de pesquisa e inovação Horizon 2020 da União Europeia no âmbito do acordo de subvenção Marie Skłodowska-Curie nº 754411 como pós-doutorado da ISTplus; A.H. recebeu apoio de um ÖAW DOC (Bolsa de Doutorado da Academia Austríaca de Ciências). Este estudo também foi apoiado pelo European Research Council (ERC) no âmbito do programa de pesquisa e inovação Horizon 2020 da União Europeia (acordo de subvenção nº 725780 LinPro) à S.H.
Materiais
| Name | Company | Catalog Number | Comments |
| 1 mL tuberculin syringe (Omnifix Luer Lock) | Braun | 9204512N | |
| 1,4-diazabicyclooctane (DABCO) | Roth | 0718.2 | |
| 10 mL Syringe (Omnifix Luer Lock) | Braun | 8508429N | |
| 15 mL conical centrifuge | Sarstedt | 65.554.502 | |
| 24 multi-well dishes | Roth/Greiner Bio-one | CE56.1 | |
| 27- gauge x 3/4 needle (Sterican) | Braun | 16010256E | |
| Corn oil | Sigma | C8267-500ML | |
| Coverslips (24 x 60 mm #1) | Thermo Fisher Scientific (Menzel) | 15747592 | |
| Cryostat Cryostar NX70 | Thermo Fisher Scientific | 957000H | |
| Dako Pen (Wax marker) | Agilent | S200230-2 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Invitrogen | D1306 | |
| Disposable microtome blade (MX35 Ultra) | Thermo Fisher Scientific | 705830 | |
| Fine Forceps (Dumont #5) | Fine Science Tools (FST) | 11254-20 | |
| Glass anti-roll plate | Histocom | M 449980 | |
| Glycerol | Sigma | G5516 | |
| LSM 800 Confocal | Zeiss | ||
| Mounting medium | 25 mg/mL DAPCO, 6 g Glycerol, 2.4 g Mowiol 4-88, 6 mL dH2O, 12 mL 0.2 M Tris-HCl (pH 8.5) | ||
| Mowiol 4-88 | Roth | 0713.2 | |
| Normal donkey serum | Innovative Research | IGDNSER100ML | |
| Paraformaldehyde | Sigma | 441244-1KG | |
| Peristaltic pump 323E/D 400RPM | Watson-Marlow | 036.3124.00A | |
| Sucrose | Sigma | S8501-5KG | |
| Superfrost plus glass slides | Thermo Fisher Scientific | J1800AMNT | |
| Tamoxifen | Sigma | T5648 | |
| Tissue Embedding mold T-12 (22mm square) | Polysciences Inc. | 18986-1 | |
| Tissue-Tek O.C.T | Sakura | 4583 | |
| Triton X-100 | Sigma | T8787-250ML | |
| Trizma hydrochloride | Sigma | 93363 | |
| Tween-20 | Sigma | P9416-100ML | |
| Software and Plugins: | |||
| Fiji | 1.52p | Fiji | |
| MultiStackReg | 1.45 | Download link | |
| TurboReg | EPFL Bioimaging | ||
| Zen Blue | 2.6 | Zeiss | |
| Experimental Models: Organisms/Strains: | |||
| Mouse: Emx1-CreER | The Jackson Laboratory | JAX:027784 | |
| Mouse: MADM-11-GT | The Jackson Laboratory | JAX:013749 | |
| Mouse: MADM-11-TG | The Jackson Laboratory | JAX:013751 | |
| Primary antibodies: | |||
| Chicken anti-GFP 1:500 | Aves Labs | GFP-1020 | |
| Goat anti-tdTomato 1:500 | Sicgen Antibodies | AB8181-200 | |
| Rabbit anti-RFP 1:500 | MBL | PM005 | |
| Secondary antibodies: | |||
| Donkey Anti-Chicken Alexa Fluor 488 1:500 | Jackson Immuno Research | 715-475-150 | |
| Donkey Anti-Goat Cy3 1:500 | Jackson Immuno Research | 705-165-147 | |
| Donkey Anti-Rabbit Cy3 1:500 | Jackson Immuno Research | 711-165-152 |
Referências
- Malatesta, P., et al. Neuronal or glial progeny: regional differences in radial glia fate. Neuron. 37 (5), 751-764 (2003).
- Miyata, T., Kawaguchi, A., Okano, H., Ogawa, M. Asymmetric inheritance of radial glial fibers by cortical neurons. Neuron. 31 (5), 727-741 (2001).
- Noctor, S. C., Flint, A. C., Weissman, T. A., Dammerman, R. S., Kriegstein, A. R. Neurons derived from radial glial cells establish radial units in neocortex. Nature. 409 (6821), 714-720 (2001).
- Taverna, E., Götz, M., Huttner, W. B. The Cell Biology of Neurogenesis: Toward an Understanding of the Development and Evolution of the Neocortex. Annual Review of Cell and Developmental Biology. 30 (1), 465-502 (2014).
- Desikan, R. S., Barkovich, A. J. Malformations of cortical development. Annals of Neurology. 80 (6), 797-810 (2016).
- Gao, R., Penzes, P. Common mechanisms of excitatory and inhibitory imbalance in schizophrenia and autism spectrum disorders. Current Molecular Medicine. 15 (2), 146-167 (2015).
- Beattie, R., et al. Mosaic Analysis with Double Markers Reveals Distinct Sequential Functions of Lgl1 in Neural Stem Cells. Neuron. 94 (3), 517-533 (2017).
- Hippenmeyer, S. Molecular pathways controlling the sequential steps of cortical projection neuron migration. Advances in Experimental Medicine and Biology. 800, 1-24 (2014).
- Lodato, S., Arlotta, P. Generating neuronal diversity in the mammalian cerebral cortex. Annual Review of Cell and Developmental Biology. 31, 699-720 (2015).
- Hansen, A. H., Duellberg, C., Mieck, C., Loose, M., Hippenmeyer, S. Cell Polarity in Cerebral Cortex Development-Cellular Architecture Shaped by Biochemical Networks. Frontiers in Cellular Neuroscience. 11, 176 (2017).
- Gao, P., et al. Deterministic progenitor behavior and unitary production of neurons in the neocortex. Cell. 159 (4), 775-788 (2014).
- Llorca, A., et al. A stochastic framework of neurogenesis underlies the assembly of neocortical cytoarchitecture. eLife. 8, e51381 (2019).
- Ma, J., Shen, Z., Yu, Y. C., Shi, S. H. Neural lineage tracing in the mammalian brain. Current Opinion in Neurobiology. 50, 7-16 (2018).
- Caviness, V., Takahashi, T., Nowakowski, R. Numbers, time and neocortical neuronogenesis: a general developmental and evolutionary model. Trends in Neurosciences. 18 (9), 379-383 (1995).
- Schmechel, D. E., Rakic, P. A Golgi study of radial glial cells in developing monkey telencephalon: morphogenesis and transformation into astrocytes. Anatomy and Embryology. 156 (2), 115-152 (1979).
- Kessaris, N., et al. Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nature Neuroscience. 9 (2), 173-179 (2006).
- Magavi, S., Friedmann, D., Banks, G., Stolfi, A., Lois, C. Coincident generation of pyramidal neurons and protoplasmic astrocytes in neocortical columns. The Journal of Neuroscience. 32 (14), 4762-4772 (2012).
- Anthony, T. E., Klein, C., Fishell, G., Heintz, N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 41 (6), 881-890 (2004).
- Voigt, T. Development of glial cells in the cerebral wall of ferrets: direct tracing of their transformation from radial glia into astrocytes. The Journal of Comparative Neurology. 289 (1), 74-88 (1989).
- Amberg, N., Laukoter, S., Hippenmeyer, S. Epigenetic cues modulating the generation of cell-type diversity in the cerebral cortex. Journal of Neurochemistry. 149 (1), 12-26 (2019).
- Beattie, R., Hippenmeyer, S. Mechanisms of Radial Glia Progenitor Cell Lineage Progression. FEBS letters. 591 (24), 3993-4008 (2017).
- Telley, L., et al. Temporal patterning of apical progenitors and their daughter neurons in the developing neocortex. Science. 364 (6440), eaav2522 (2019).
- Oberst, P., et al. Temporal plasticity of apical progenitors in the developing mouse neocortex. Nature. 573 (7774), 370-374 (2019).
- Telley, L., et al. Sequential transcriptional waves direct the differentiation of newborn neurons in the mouse neocortex. Science. 351 (6280), 1443 (2016).
- Deppe, U., et al. Cell lineages of the embryo of the nematode Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America. 75 (1), 376-380 (1978).
- Woodworth, M. B., Girskis, K. M., Walsh, C. A. Building a lineage from single cells: genetic techniques for cell lineage tracking. Nature Reviews Genetics. 18 (4), 230-244 (2017).
- Masuyama, N., Mori, H., Yachie, N. DNA barcodes evolve for high-resolution cell lineage tracing. Current Opinion in Chemical Biology. 52, 63-71 (2019).
- Legue, E., Joyner, A. L. Chapter Ten-Genetic Fate Mapping Using Site-Specific Recombinases. Methods in Enzymology. 477, 153-181 (2010).
- Postiglione, M. P., Hippenmeyer, S. Monitoring neurogenesis in the cerebral cortex: an update. Future Neurology. 9 (3), 323-340 (2014).
- Espinosa-Medina, I., Garcia-Marques, J., Cepko, C., Lee, T. High-throughput dense reconstruction of cell lineages. Open Biology. 9 (12), 190229 (2019).
- Hwang, B., et al. Lineage tracing using a Cas9-deaminase barcoding system targeting endogenous L1 elements. Nature Communications. 10 (1), 1234 (2019).
- Kalhor, R., et al. Developmental barcoding of whole mouse via homing CRISPR. Science. 361 (6405), eaat9804 (2018).
- García-Moreno, F., Vasistha, N. A., Begbie, J., Molnár, Z. CLoNe is a new method to target single progenitors and study their progeny in mouse and chick. Development. 141 (7), 1589-1598 (2014).
- Loulier, K., et al. Multiplex Cell and Lineage Tracking with Combinatorial Labels. Neuron. 81 (3), 505-520 (2014).
- Livet, J., et al. Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature. 450 (7166), 56-62 (2007).
- Snippert, H. J., et al. Intestinal Crypt Homeostasis Results from Neutral Competition between Symmetrically Dividing Lgr5 Stem Cells. Cell. 143 (1), 134-144 (2010).
- Amitai-Lange, A., et al. A method for lineage tracing of corneal cells using multi-color fluorescent reporter mice. Journal of Visualized Experiments. (106), e53370 (2015).
- Vasistha, N. A., et al. Cortical and Clonal Contribution of Tbr2 Expressing Progenitors in the Developing Mouse Brain. Cerebral Cortex. 25 (10), 3290-3302 (2015).
- Chen, F., LoTurco, J. A method for stable transgenesis of radial glia lineage in rat neocortex by piggyBac mediated transposition. Journal of Neuroscience Methods. 207 (2), 172-180 (2012).
- Siddiqi, F., et al. Fate mapping by piggyBac transposase reveals that neocortical GLAST+ progenitors generate more astrocytes than Nestin+ progenitors in rat neocortex. Cerebral Cortex. 24 (2), 508-520 (2014).
- Yoshida, A., et al. Simultaneous expression of different transgenes in neurons and glia by combining in utero electroporation with the Tol2 transposon-mediated gene transfer system. Genes to Cells. 15 (5), 501-512 (2010).
- García-Marqués, J., López-Mascaraque, L. Clonal Identity Determines Astrocyte Cortical Heterogeneity. Cerebral Cortex. 23 (6), 1463-1472 (2013).
- Clavreul, S., et al. Cortical astrocytes develop in a plastic manner at both clonal and cellular levels. Nature Communications. 10 (1), 4884 (2019).
- Kim, G. B., et al. Rapid Generation of Somatic Mouse Mosaics with Locus-Specific, Stably Integrated Transgenic Elements. Cell. 179 (1), 251-267 (2019).
- Pilz, G. A., et al. Live imaging of neurogenesis in the adult mouse hippocampus. Science. 359 (6376), 658 (2018).
- Hippenmeyer, S., et al. Genetic Mosaic Dissection of Lis1 and Ndel1 in Neuronal Migration. Neuron. 68 (4), 695-709 (2010).
- Zong, H., Espinosa, J. S., Su, H. H., Muzumdar, M. D., Luo, L. Mosaic analysis with double markers in mice. Cell. 121 (3), 479-492 (2005).
- Ramón y Cajal, S. . Histologie du système nerveux de l'homme et des vertébrés. , (1911).
- Cowan, W. M. The emergence of modern neuroanatomy and developmental neurobiology. Neuron. 20 (3), 413-426 (1998).
- Ali, S. R., et al. Existing cardiomyocytes generate cardiomyocytes at a low rate after birth in mice. Proceedings of the National Academy of Sciences of the United States of America. 111 (24), 8850-8855 (2014).
- Hippenmeyer, S., Johnson, R. L., Luo, L. Mosaic analysis with double markers reveals cell-type-specific paternal growth dominance. Cell Reports. 3 (3), 960-967 (2013).
- Riccio, P., Cebrian, C., Zong, H., Hippenmeyer, S., Costantini, F. Ret and Etv4 promote directed movements of progenitor cells during renal branching morphogenesis. PLoS Biology. 14 (2), e1002382 (2016).
- Bonaguidi, M. A., et al. In vivo clonal analysis reveals self-renewing and multipotent adult neural stem cell characteristics. Cell. 145 (7), 1142-1155 (2011).
- Mayer, C., et al. Clonally Related Forebrain Interneurons Disperse Broadly across Both Functional Areas and Structural Boundaries. Neuron. 87 (5), 989-998 (2015).
- Muzumdar, M. D., et al. Clonal dynamics following p53 loss of heterozygosity in Kras-driven cancers. Nature Communications. 7, 12685 (2016).
- Shi, W., et al. Ontogenetic establishment of order-specific nuclear organization in the mammalian thalamus. Nature Neuroscience. 20, 516 (2017).
- Wong, S. Z. H., et al. In vivo clonal analysis reveals spatiotemporal regulation of thalamic nucleogenesis. PLoS Biology. 16 (4), e2005211 (2018).
- Xu, H. T., et al. Distinct Lineage-Dependent Structural and Functional Organization of the Hippocampus. Cell. 157 (7), 1552-1564 (2014).
- Brennand, K., Huangfu, D., Melton, D. All β Cells Contribute Equally to Islet Growth and Maintenance. PLoS Biology. 5 (7), e163 (2007).
- Ortiz-Alvarez, G., et al. Adult neural stem cells and multiciliated ependymal cells share a common lineage regulated by the geminin family members. Neuron. 102 (1), 159-172 (2019).
- Kaplan, E. S., Ramos-Laguna, K. A., Mihalas, A. B., Daza, R. A. M., Hevner, R. F. Neocortical Sox9+ radial glia generate glutamatergic neurons for all layers, but lack discernible evidence of early laminar fate restriction. Neural Development. 12 (1), 14 (2017).
- Lv, X., et al. TBR2 coordinates neurogenesis expansion and precise microcircuit organization via Protocadherin 19 in the mammalian cortex. Nature Communications. 10 (1), 3946 (2019).
- Mihalas, A. B., Hevner, R. F. Clonal analysis reveals laminar fate multipotency and daughter cell apoptosis of mouse cortical intermediate progenitors. Development. 145 (17), dev164335 (2018).
- Picco, N., et al. A mathematical insight into cell labelling experiments for clonal analysis. Journal of Anatomy. 235 (3), 687-696 (2019).
- Johnson, C. A., Ghashghaei, H. T. Sp2 regulates late neurogenic but not early expansive divisions of neural stem cells underlying population growth in the mouse cortex. Development. , (2020).
- Espinosa, J. S., Luo, L. Timing Neurogenesis and Differentiation: Insights from Quantitative Clonal Analyses of Cerebellar Granule Cells. The Journal of Neuroscience. 28 (10), 2301 (2008).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Liu, Y., et al. Tamoxifen-independent recombination in the RIP-CreER mouse. PLoS One. 5 (10), e13533 (2010).
- Klingler, E., Jabaudon, D. Do progenitors play dice?. eLife. 9, e54042 (2020).
- Hippenmeyer, S. Dissection of gene function at clonal level using mosaic analysis with double markers. Frontiers in Biology. 8 (6), 557-568 (2013).
- Rulands, S., Simons, B. D. Tracing cellular dynamics in tissue development, maintenance and disease. Current Opinion in Cell Biology. 43, 38-45 (2016).
- Laukoter, S., et al. Imprinted Cdkn1c genomic locus cell-autonomously promotes cell survival in cerebral cortex development. Nature Communications. 11 (1), 195 (2020).
- Daigle, T. L., et al. A Suite of Transgenic Driver and Reporter Mouse Lines with Enhanced Brain-Cell-Type Targeting and Functionality. Cell. 174 (2), 465-480 (2018).
- He, M., et al. Strategies and Tools for Combinatorial Targeting of GABAergic Neurons in Mouse Cerebral Cortex. Neuron. 91 (6), 1228-1243 (2016).
- Yamamoto, M., et al. A multifunctional reporter mouse line for Cre- and FLP-dependent lineage analysis. Genesis. 47 (2), 107-114 (2009).
- Plummer, N. W., et al. Expanding the power of recombinase-based labeling to uncover cellular diversity. Development. 142 (24), 4385 (2015).
- Imayoshi, I., Ohtsuka, T., Metzger, D., Chambon, P., Kageyama, R. Temporal regulation of Cre recombinase activity in neural stem cells. Genesis. 44 (5), 233-238 (2006).
- Sasaki, S., et al. Complete loss of Ndel1 results in neuronal migration defects and early embryonic lethality. Molecular and Cellular Biology. 25 (17), 7812-7827 (2005).
- Acampora, D., et al. Epilepsy and brain abnormalities in mice lacking the Otx1 gene. Nature Genetics. 14 (2), 218-222 (1996).
- Soeda, T., et al. Sox9-expressing precursors are the cellular origin of the cruciate ligament of the knee joint and the limb tendons. Genesis. 48 (11), 635-644 (2010).
- Klezovitch, O., Fernandez, T. E., Tapscott, S. J., Vasioukhin, V. Loss of cell polarity causes severe brain dysplasia in Lgl1 knockout mice. Genes & Development. 18 (5), 559-571 (2004).
- Pimeisl, I. M., et al. Generation and characterization of a tamoxifen-inducible EomesCreER mouse line. Genesis. 51 (10), 725-733 (2013).
- Nakagawa, N., et al. Memo1-Mediated Tiling of Radial Glial Cells Facilitates Cerebral Cortical Development. Neuron. 103 (5), 836-852 (2019).
- Nowotschin, S., et al. The T-box transcription factor Eomesodermin is essential for AVE induction in the mouse embryo. Genes & Development. 27 (9), 997-1002 (2013).
- Balordi, F., Fishell, G. Mosaic removal of hedgehog signaling in the adult SVZ reveals that the residual wild-type stem cells have a limited capacity for self-renewal. Journal of Neuroscience. 27 (52), 14248-14259 (2007).
- Liang, H., et al. Neural development is dependent on the function of specificity protein 2 in cell cycle progression. Development. 140 (3), 552-561 (2013).
- Guo, C., Yang, W., Lobe, C. G. A Cre recombinase transgene with mosaic, widespread tamoxifen-inducible action. Genesis. 32 (1), 8-18 (2002).
- Ahn, S., Joyner, A. L. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature. 437 (7060), 894-897 (2005).
- Storm, R., et al. The bHLH transcription factor Olig3 marks the dorsal neuroepithelium of the hindbrain and is essential for the development of brainstem nuclei. Development. 136 (2), 295-305 (2009).
- Koundakjian, E. J., Appler, J. L., Goodrich, L. V. Auditory neurons make stereotyped wiring decisions before maturation of their targets. Journal of Neuroscience. 27 (51), 14078-14088 (2007).
- Dor, Y., Brown, J., Martinez, O. I., Melton, D. A. Adult pancreatic β-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 429 (6987), 41-46 (2004).
- Sohal, D. S., et al. Temporally regulated and tissue-specific gene manipulations in the adult and embryonic heart using a tamoxifen-inducible Cre protein. Circulation Research. 89 (1), 20-25 (2001).
- Ventura, A., et al. Restoration of p53 function leads to tumour regression in vivo. Nature. 445 (7128), 661-665 (2007).
- Johnson, L., et al. Somatic activation of the K-ras oncogene causes early onset lung cancer in mice. Nature. 410 (6832), 1111-1116 (2001).
- Tasic, B., et al. Extensions of MADM (mosaic analysis with double markers) in mice. PLoS One. 7 (3), e33332 (2012).
- Yu, J., Carroll, T. J., McMahon, A. P. Sonic hedgehog regulates proliferation and differentiation of mesenchymal cells in the mouse metanephric kidney. Development. 129 (22), 5301-5312 (2002).
- Zhao, H., et al. Role of fibroblast growth factor receptors 1 and 2 in the ureteric bud. Developmental Biology. 276 (2), 403-415 (2004).
- Schuchardt, A., D'Agati, V., Larsson-Blomberg, L., Costantini, F., Pachnis, V. Defects in the kidney and enteric nervous system of mice lacking the tyrosine kinase receptor Ret. Nature. 367 (6461), 380-383 (1994).
- Livet, J., et al. ETS gene Pea3 controls the central position and terminal arborization of specific motor neuron pools. Neuron. 35 (5), 877-892 (2002).
- Volz, K. S., et al. Pericytes are progenitors for coronary artery smooth muscle. eLife. 4, e10036 (2015).
- Cai, C. L., et al. A myocardial lineage derives from Tbx18 epicardial cells. Nature. 454 (7200), 104-108 (2008).
- Lasrado, R., et al. Lineage-dependent spatial and functional organization of the mammalian enteric nervous system. Science. 356 (6339), 722-726 (2017).
- Matsuoka, T., et al. Neural crest origins of the neck and shoulder. Nature. 436 (7049), 347-355 (2005).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados