
このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
植物の根に土壌媒介微生物を感染させる接種戦略
要約
このプロトコルは、植物の根に土壌媒介微生物を接種するための戦略の詳細な要約を提示する。真菌 Verticillium longisporum および Verticillium dahliaeについて例示された、3つの異なる根感染系が記載されている。潜在的なアプリケーションと可能なダウンストリーム解析が強調表示され、各システムについて長所または短所が議論されます。
要約
根圏には、植物の根が常に挑戦されている非常に複雑な微生物群集があります。根は多種多様な微生物と密接に接触しているが、土壌媒介相互作用に関する研究は地上の臓器で行われている研究にはまだ遅れをとっている。モデル植物をモデル根病原体に感染させるためのいくつかの接種戦略が文献に記載されているが、包括的な方法論的概要を得ることは依然として困難である。この問題に対処するために、根と微生物の相互作用の生物学への洞察を得るために適用することができる3つの異なる根接種システムが正確に記述されている。説明のために、 Verticillium 種(すなわち、 V. longisporum および V. dahliae)を根侵入モデル病原体として使用した。しかし、この方法は、病原性および有益な他の根コロニー形成微生物に容易に適合させることができる。植物の木部に定着することにより、 バーティシリウム 属などの維管束性土壌媒介性真菌は独特の生活様式を示す。根の侵入後、それらは木部血管 を介して 無作為に広がり、芽に達し、そして病気の症状を誘発する。モデル宿主として、 シロイヌナズナ、経済的に重要な油糧種子菜種(アブラナナ)、トマト(Solanum lycopersicum)の3つの代表的な植物種が選ばれた。ステップバイステップのプロトコルが与えられます。病原性アッセイ、マーカー遺伝子の転写解析、およびレポーター構築物による独立した確認の代表的な結果が示されている。さらに、各接種系の長所と短所を徹底的に議論する。これらの実証済みのプロトコルは、根と微生物の相互作用に関する研究上の質問に対するアプローチを提供するのに役立ちます。植物が土壌中の微生物にどのように対処するかを知ることは、農業を改善するための新しい戦略を開発するために不可欠です。
概要
天然土壌には、植物にとって中性、有害、または有益な驚くほど多くの微生物が生息しています1。多くの植物病原菌は土壌に生着し、根を囲み、地下の器官を攻撃する。これらの微生物は、真菌、卵菌類、細菌、線虫、昆虫、およびいくつかのウイルス1,2の多種多様なクレードに属する。環境条件が感染に有利になると、影響を受けやすい植物は病気になり、作物収量は減少します。地球温暖化や極端な気象などの気候変動の影響により、土壌に媒介する植物病原菌の割合が増加します3。したがって、これらの破壊的な微生物と、それらが食料や飼料の生産だけでなく、自然の生態系に与える影響を研究することはますます重要になります。さらに、土壌中には根と密接に相互作用し、植物の成長、発達、免疫を促進する微生物の相互主義者がいます。病原体に直面したとき、植物は病原体を抑制することによって宿主の生存をサポートできる根圏内の特定の敵を積極的に募集することができる4、5、6、7。しかし、有益な根と微生物の相互作用に関与する機構的な詳細と経路は、しばしばまだ不明である6。
したがって、根と微生物の相互作用の一般的な理解を広げることが不可欠です。土壌媒介微生物を根に接種するための信頼できる方法は、モデル研究を行い、その結果を農業用途に移転するために必要である。土壌中の有益な相互作用は、例えば、セレンディピタ・インディカ(以前はピリホルモスポラ・インディカとして知られていた)、窒素固定リゾビウム属、または菌根菌との研究されているが、既知の土壌媒介性植物病原体には、ラルストニア・ソラナセアラム、フィトフトラ属、フザリウム属、およびバーティシリウム属が含まれる。後者の2つは、世界中に分布し、血管疾患を引き起こす真菌属である2。Verticillium spp.(Ascomycota)は、何百もの植物種に感染する可能性があります - 草本一年生植物、木質多年生植物、および多くの作物植物を含む主に双子葉類2,8。Verticilliumの菌糸は根に入り、中央シリンダーに向かって細胞間および細胞内の両方で成長し、木部血管2,9に定着する。これらの血管では、真菌はそのライフサイクルのほとんどに留まる。木部樹液は栄養価が低く、植物防御化合物を運ぶので、真菌はこのユニークな環境に適応しなければなりません。これは、病原体がその宿主で生存することを可能にするコロニー形成関連タンパク質の分泌によって達成される10,11。根の血管系に到達した後、真菌は木部血管内で葉に無作為に広がることができ、これは宿主9,12の全身コロニー形成をもたらす。この時点で、植物は成長9、10、13に悪影響を受けます。例えば、発育不全および黄色い葉は、早期老化と同様に起こる13,14,15,16。
この属のメンバーの1つはVerticillium longisporumであり、これは農業学的に重要な油糧種子菜種、カリフラワー、およびモデル植物シロイヌナズナ12などのアブラナ科宿主に高度に適応している。いくつかの研究は、V. longisporumとA. thalianaを組み合わせて、土壌媒介性血管疾患およびその結果生じる根防御応答に関する広範な洞察を得た13,15,16,17。V. longisporum / A. thalianaモデルシステムを使用することによって、簡単な感受性試験を実現することができ、十分に確立された遺伝資源が両方の生物に利用可能である。V. longisporumと密接に関連しているのは、病原体Verticillium dahliaeである。両方の真菌種は同様の血管ライフスタイルおよび侵入プロセスを実行するが、根から葉へのそれらの増殖効率およびA. thalianaにおける誘発された疾患症状は異なる:V. longisporumは通常早期老化を誘発するが、V. dahliae感染は萎縮をもたらす18。最近、方法論的要約は、A. thalianaにV. longisporumまたはV. dahliaeを感染させるための異なる根接種戦略を提示し、実験セットアップの計画を支援した19。畑では、V. longisporumは時折油糧種子菜種生産に重大な損傷を与える12が、V. dahliaeはブドウ、ジャガイモ、トマトなどのいくつかの栽培種を含む非常に広い宿主範囲を有する8。これにより、両方の病原体を研究するのに経済的に興味深いモデルになります。
したがって、以下のプロトコルは、V. longisporum および V. dahliae の両方をモデル根病原体として使用して、根接種のための可能なアプローチを例示する。シロイヌナズナ(Arabidopsis thaliana)、油糧種子菜種(アブラナナプス)、およびトマト(Solanum lycopersicum)がモデル宿主として選択された。方法論の詳細な説明は、以下のテキストと付属のビデオで見つけることができます。各接種系の長所と短所について考察する。まとめると、このプロトコルコレクションは、根と微生物の相互作用の文脈における特定の研究課題に適した方法を特定するのに役立ちます。
Access restricted. Please log in or start a trial to view this content.
プロトコル
1. 真菌培養用培地および植物接種システム
- 液体ジャガイモデキストロースブロス(PDB):熱安定性フラスコ内の超純水中に21g/L PDBを調製する。
- 液体チャペックデキストロースブロス(CDB):熱安定性フラスコ内の超純水中に42g / L CDBを調製する。
- シャーレ接種システム用培地:超純水中に1.5g/Lのムラシゲ培地とスクーグ培地(MS)と8g/L寒天を入れた熱安定性フラスコを用意する。
注:この培地中の砂糖は、接種後に過剰な真菌の増殖につながるため、避けてください。 - プラスチックカップベースの接種システム用培地:超純水中で4.4 g/L MS、0.2 g/L MgSO4、1 g/L KNO3、0.5 g/L 2-(N-モルホリノ)エタンスルホン酸(MES)、および6.0 g/L寒天を含む熱安定性フラスコを調製し、5 M KOHでpHを5.7に調整する。
注:この培地中の砂糖は、接種後に過剰な真菌の増殖につながるため、避けてください。 - 1/4 MS 培地: 超純水に 1.2 g/L MS を調製します。
- オートクレーブを使用して、上記のすべての溶液を滅菌します。ガラスフラスコをバスケットに入れ、蓋を閉じ、121°C、98.9kPaで15分間殺菌する。
2.植物種子の表面を殺菌する
注:播種前にシロイヌナズナ、油糧種子菜種、およびトマトからの種子の表面を滅菌するには、常に以下のプロトコルを使用してください。
- 種子を2mL反応管に移す。チューブを内部容量5.8Lのエクスシケーターに入れます。
- 100mLの12%次亜塩素酸ナトリウム水溶液(NaClO)に6mLの33%塩酸(HCl)を加えて、エキシケーター内で塩素ガスを生成する。
- すぐにエキサイケーターの蓋を閉じ、種子をガス中で3時間インキュベートする。
3. バーティシリウム 胞子(無性由来分生子)の接種剤の調製
注:V. dahliae(JR2株)をV. longisporum(Vl43株)17,18,19と同じ方法で栽培する。すべての機器と培地に無菌であること、および接種源を軸状に保つためにすべてのステップが層流フード内で実行されることを確認します。
- 150 mL の液体 PDB (ステップ 1.1) を 500 mL のシカンリーフラスコに充填し、培地に 500 mg/L のセフォタキシムを補充します。
- グリセロールストック貯蔵からの バーティシリウム ・コニダをPDB培地に加える。フラスコを滅菌フォームストッパーで閉じます。
- 培養物を室温(RT)の暗箱中で7〜10日間、連続的な水平振とう(ロータリーシェーカー;60rpm)下で培養する。これにより、小さな白い菌糸球体が得られます。
- PDBの上清を慎重に取り除いて捨てます。菌糸体のほとんどはフラスコに残るはずです。
- 100mLの液体CDBをシカンリーフラスコ内の菌糸体に加え(ステップ1.2)、培地に500mg/Lセフォタキシムを補充する。
- RTの暗箱中でさらに4〜5日間インキュベートし、連続的な水平振とう(ロータリーシェーカー、60rpm)下で胞子形成を誘導する。上清は分生子が放出されると黄色がかった灰色に変わります。
- 分生子含有液の一部(5〜10mL)を濾紙(粒子保持レベル8〜12μm)を通して滅菌50mL回収管に濾過する。これは胞子を菌糸体から分離する。
- 細胞計数室および顕微鏡を用いて胞子濃度を決定する。無菌の1/4MS培地を超純水で希釈し、下記の胞子濃度が得られるまで希釈する。
注:顕微鏡下では、 V. longisporum の分生子はほとんどが長く描かれ、サイズは7.1-8.8 μmですが、 V. dahliae conidiaはより短く(3.5-5.5 μm)、かなり球形20です。 - これらの収穫したての分生子を接種剤として使用します。凍結すると生存可能な胞子の数が大幅に減少するため、凍結ストックではなく、収穫したての分生子で常に実験を行うようにしてください19。
- 長期保存の場合は、胞子を高濃度胞子溶液(約1 x 108胞子/mL)として-80 °Cで25%グリセロールで凍結します(最大1年間保存可能)。次の実験のために、これらのグリセロールストックを使用して、ステップ3.2でPDB培地を接種する。
4. ペトリ皿に基づく滅菌 インビトロ 接種システム
注:ペトリ皿システム17の場合、すべての機器および媒体が無菌であり、すべてのステップが層流フード内で実行されることを確認してください。
- オートクレーブ処理後、培地をペトリ皿に注ぎます(ステップ1.3を参照)。
- 培地の硬化後、ペトリ皿を滅菌ビニール袋に詰め直し、冷蔵庫(4〜10°C)に一晩逆さまに保管する。冷却された培地は、次のステップでの培地の滑りを防ぐのに役立ちます。
- 感染チャネルおよび固化培地の上3分の1をメスで切断して除去する(図1A)。切断中に寒天培地の下に液体や空気を得ることを避けます;さもなければ、培地は滑って感染チャネルを閉じるでしょう。
- 50-100個の表面滅菌シロイヌナズナ種子を、切断された上面に滅菌ピペットチップで分配する。切り取った寒天表面がペトリ皿の壁に接触する角度に種子を入れて、根が培地とペトリ皿の壁の間に成長できるようにします。これにより、後で接種が容易になります。
- ペトリ皿を閉じ、通気性粘着テープで密封してガス交換を可能にする。
- 4°Cの暗所で2日間成層化した後、プレートを適当なラックに垂直に置き、生育室内で22°C±1°Cの長日条件下(16時間光/暗8時間)で植物を生育させた。
- 根の大部分が感染チャネル(約9〜11日齢の苗)に到達したら、プレートを水平に置き、それらを開き、4 x 105胞子/ mLの濃度で収穫したばかりのVerticillium conidiaを500μL感染チャネルに直接加え、液体がチャネル内に均等に分布することを確認する。
- 同様に、胞子の代わりに500 μLの偽溶液(無菌1/4 MS培地)を加えてコントロールプレートを調製する。
- 液体が浸かり、プレートが再び垂直にセットアップされたときに漏れ出さなくなるまで、プレートを水平に数分間インキュベートします。その後、蓋を閉じ、通気性粘着テープでプレートを密封します。
- プレートを成長チャンバー内で垂直にインキュベートする。必要に応じて,根と土壌媒介菌を暗くするために,根の部分を黒い紙箱で覆います(19参照)。
- 研究課題に応じて、接種後の優先時点で分析を実行します(ここで使用される正確な時間点については、図の凡例を参照してください)。以下はいくつかの提案です。
- 根から葉を切り取り、両方を別々に収穫する。ペトリ皿から寒天ストリップを取り出して根に簡単にアクセスし、鉗子を使用して寒天から慎重に引き出します。すべての植物材料を液体窒素で直ちに凍結する。
- サンプルを液体窒素で粉砕する。100mgの葉材料から全DNAを抽出し、植物DNAに対する真菌DNAの量を定量PCR(qPCR) を介して 決定する(19参照)。
- サンプルを液体窒素で粉砕する。植物材料100mgを採取し、トータルRNAを抽出する。定量的逆転写PCR(qRT-PCR)を実施して、侵入中の植物遺伝子(または真菌遺伝子)の発現を決定する(19参照)。
- 怪我を避けて寒天から根を慎重に取り除き、蛍光顕微鏡で調べます。
- 植物レポーター系統におけるマーカー遺伝子(例えば、ルシフェラーゼ、β-グルクロニダーゼ、または蛍光レポーター17、19、21)の誘導を決定する。
- 真菌レポーター株(例えば、増強された緑色蛍光タンパク質を恒常的に発現するV.ロンギスポラム、Vl-sGFP9)を使用するか、染色技術(例えば、5-ブロモ-4-クロロ-3-インドキシル-N-アセチル-β-d-グルコサミニド(X-β-D-Glc-Nac)を介して)によって、根元での真菌の繁殖を視覚化する18。
- 根から葉を切り取り、両方を別々に収穫する。ペトリ皿から寒天ストリップを取り出して根に簡単にアクセスし、鉗子を使用して寒天から慎重に引き出します。すべての植物材料を液体窒素で直ちに凍結する。
5. プラスチックカップで組織された滅菌 インビトロ 接種システム
注:この技術19の最初の説明で述べたように、すべての機器および媒体が無菌であり、すべてのステップが層流フード内で実行されることを確認してください。
- 総容量500mLの透明なプラスチックカップを使用し、70%〜75%エタノール浴中で少なくとも20分間滅菌する。層流フードでカップを乾かします。
- オートクレーブ処理した媒体(ステップ1.4参照)をプラスチックカップに注ぎます。必要に応じて、セフォタキシム(最終濃度50mg/L)をオートクレーブ処理培地に添加し、細菌汚染を防止します。シロイヌナズナの実験には1カップあたり150mLの培地を使用し、より大きな植物種(油糧種子菜種、トマト)の実験にはそれ以上の培地(カップあたり250〜300mL)を使用します。
- それが固化する前に、プラスチック層(70%〜75%エタノール中で20分間インキュベートすることによって滅菌前)を培地上に置く(図1B)。
注:このプラスチック層には、表面滅菌種子を配置するためのコーナーに4つのプレハブ穴が含まれています。これにより、シードはメディアにアクセスできるようになります。その後、この分離層は、葉が真菌含有培地に触れるのを防ぎ、微生物が葉を直接攻撃することができず、根の経路をとらなければならない。中央にはもう1つの穴があり、感染経路を切断することができます。 - 培地が固化したら、プレハブの中央の穴を通してメスで寒天を約1.5cmの深さに切ります。カットされた寒天を除去して、真菌胞子を後で追加できる感染チャネルを作成する。
- 寒天培地を4つの小さな穴にピペットチップでわずかに引っ掻き、固化した皮膚を中断します(これにより、種子が水っぽい寒天培地から水を吸収することができます)。ピペットチップを使用して種子を小さな穴に入れます。
- 2番目の反転プラスチックカップでプラスチックカップを閉じ、通気性粘着テープでシールします。テープはガス交換を可能にする必要があります。
- 4°Cの暗闇で3日間層別化した後、カップシステムを22°Cの一定温度および湿度60%の成長チャンバー内で12時間光/ 12時間暗闇(シロイヌナズナ、油糧種子菜種)または16時間光/ 8時間暗さ条件(トマト)下でインキュベートする。
- 接種のための植物の推奨年齢に従ってください:シロイヌナズナのための21日;油糧種子菜種のための5-7日;トマトの12日間。
- 感染チャネルに1mLの分生子溶液(推奨濃度:4 x105胞子/ mL)を加えて、小植物にVerticilliumを接種する。対照サンプルを調製するために、胞子を含まない1mLの偽溶液(無菌1/4MS培地)をチャネルに加える。
- 研究課題に応じて、接種後の優先時点で分析を実行します(ここで使用される正確な時間点については、図の凡例を参照してください)。以下はいくつかの提案です。
- 上からデジタルカメラで植物の写真を撮り、写真ごとに距離を同じに保ちます。葉の面積を定量化し(例えば、ImageJ22またはBlattFlaeche17,19で;カップの長さを使用してスケールを設定する)、感染群と対照群を比較する。病気の症状(例えば、より小さい、より黄色がかった、または壊死性の葉)の発症を分類する。
注:シロイヌナズナに茎がある場合は、ロゼットのより良い写真を得るためにそれらを削除してください。 - 根を取り除き、秤量によって感染および対照サンプルからの芽のバイオマス(新鮮な重量)を定義します。相対新鮮重量19を求める。
- 分子分析用のサンプルを以下のように採取する。
- シロイヌナズナ:茎がある場合は取り除いてください。根元にあるロゼットを根元から切ります。サンプルからすべての根材を除外し、ロゼット全体を収穫してください。異なる植物からの4〜5個のロゼットを1つのサンプルに結合し、葉の材料を液体窒素で凍結する。
- 鉗子で培地から根を慎重に引き出し、ペーパータオルで押して軽く叩いて寒天の残骸を取り除き、異なる植物の4〜5本の根を1つのサンプルに組み合わせます。液体窒素ですぐに凍結する。
- 油糧種子菜種/トマト:胚軸から茎の部分を切り取ります(例えば、長さ1cm、常に同じ茎領域を取ります)。4~5本の植物の材料を各サンプルに結合し、液体窒素で凍結します。
- サンプルを液体窒素で粉砕する。100mgの葉または茎材料から総DNAを抽出し、植物DNAに対する真菌DNAの量をqPCR を介して 決定する(19を参照されたい)。
- 液体窒素でサンプルを粉砕し、100mgの植物材料を採取し、トータルRNAを抽出する。qRT-PCRを実施して、侵入時の植物遺伝子(または真菌遺伝子)の発現を決定する(19参照)。
- 上からデジタルカメラで植物の写真を撮り、写真ごとに距離を同じに保ちます。葉の面積を定量化し(例えば、ImageJ22またはBlattFlaeche17,19で;カップの長さを使用してスケールを設定する)、感染群と対照群を比較する。病気の症状(例えば、より小さい、より黄色がかった、または壊死性の葉)の発症を分類する。
6. 鉢への土壌接種システム
- 土と砂を3:1(土:砂)の体積比で徹底的に混合し、基質を根から洗い流しやすくします。混合物をオートクレーブバッグに注ぐ。混合物が乾燥しすぎる場合は、適量の水を加えて基材に混ぜる。微生物汚染を最小限に抑えるために、オートクレーブ内で80°Cで20分間蒸気を流します。
注:有機土壌の栄養素に影響を与える可能性があるため、80°C以上に加熱しないでください。 - 鍋に土砂混合物を入れ、トレイに移します。鍋の高さの約1/3の高さのトレイに水を加えて、土壌と砂の混合物が水に完全に浸されるようにします。さらに、スプレーボトルで基板に水を噴霧して、濡れた開始条件を確保します。
- 各ポットに3〜4個の種子を蒔き(図1C)、種子が互いに十分な距離を持つようにする。発芽を同期させるために層別化のためにそれらを4°Cの暗闇の中で3日間保ちます。
注:過剰の植物を事前に栽培することで、接種実験で同程度の大きさの植物を選択でき、個体差による偏差を低減できます。 - 定期的な散水で長い日の条件(16時間の光/ 8時間の暗闇、22°Cの一定温度、湿度60%)で苗を成長させましょう。
- 接種のための植物の推奨年齢に従ってください:シロイヌナズナのための21日、油糧種子菜種のための7日、そしてトマトのための10日。同様の大きさの植物を摘み取り、「根のディップ接種」15、17、23、24を行う。鉢から土を取り出し、慎重に根を掘ります。
- 根だけを水容器で優しく洗い、ロゼットを水から遠ざけてください。洗浄した根を 、バーティシリウム 胞子溶液(推奨濃度:2 x 106胞子/mL)を含むペトリ皿中で60 分間インキュベートする。非感染対照群の場合、胞子を含まない偽液(無菌1/4MS培地)中で根を60分間インキュベートする。
- 湿った蒸気滅菌土壌(80°Cで20分間)を砂なしで新しいポットを準備します。ピペットチップを使用して、各ポットの土壌の中央に1つの穴を開けます。
- 根を穴に直接入れます(鉢ごとに1つの植物だけを移します)。根を挿入した後、穴に土を慎重に補充してください。土を押さないようにし、さもなければ鉢替えは紫色の葉のようなストレス症状を引き起こす可能性があります。
- 感染群および対照群を、定期的な散水で長い日の条件(16時間の光/ 8時間の暗闇;22°Cの一定温度、湿度60%)で栽培する。
- 研究課題に応じて、接種後の優先時点で分析を実行します(ここで使用される正確な時間点については、図の凡例を参照してください)。以下はいくつかの提案です。
- 上からデジタルカメラで植物の写真を撮り、写真ごとに距離を同じに保ちます。葉の面積を定量化し(例えば、ImageJ22またはBlattFlaeche17,19;鉢の直径を使用してスケールを設定する)、感染群と対照群を比較する。病気の症状(例えば、より小さい、より黄色がかった、または壊死性の葉)の発症を分類する13。
注:シロイヌナズナの茎を取り除くと、ロゼットの写真を撮ることが容易になります。 - 根を取り除き、秤量によって感染および対照サンプルからの芽のバイオマス(新鮮な重量)を定義します。相対新鮮重量19を求める。
- あるいは、草丈を測定するか、または茎セグメントからの真菌の伸長を分類して、疾患の重症度を評価する13。
- 分子分析用のサンプルを以下のように採取する。
- シロイヌナズナ:茎を取り除きます。根の冠でロゼットを切る。異なる植物からの4〜5個のロゼットを1つのサンプルに組み合わせる。葉の材料を液体窒素で凍結する。
注:根の場合、洗浄による遺伝子発現の初期化を行わずに土壌から十分にきれいにすることは困難です。 - 油糧種子菜種/トマト:胚軸から茎の部分を切り取ります(例えば、長さ1cm、常に同じ茎領域を取ります)。4~5本の植物の材料を1つのサンプルに混ぜ合わせ、液体窒素で凍結します。
- サンプルを液体窒素で粉砕する。100mgの葉または茎材料から総DNAを抽出し、植物DNAに対する真菌DNAの量をqPCR を介して 決定する(19を参照されたい)。
- 液体窒素でサンプルを粉砕し、100mgの植物材料を採取し、トータルRNAを抽出する。qRT-PCRを実施して、侵入時の植物遺伝子(または真菌遺伝子)の発現を決定する(19参照)。
- 上からデジタルカメラで植物の写真を撮り、写真ごとに距離を同じに保ちます。葉の面積を定量化し(例えば、ImageJ22またはBlattFlaeche17,19;鉢の直径を使用してスケールを設定する)、感染群と対照群を比較する。病気の症状(例えば、より小さい、より黄色がかった、または壊死性の葉)の発症を分類する13。
7. データの分析
- 生物学的反復に基づいて平均および標準偏差(±SD)を計算する。
- 感染グループからのすべての結果をコントロールの結果で割って、相対値を計算します。平均を、たとえば「モックに対する相対」や「野生型に対する相対」として表示します。
- グループ間の統計的有意性を判断する。

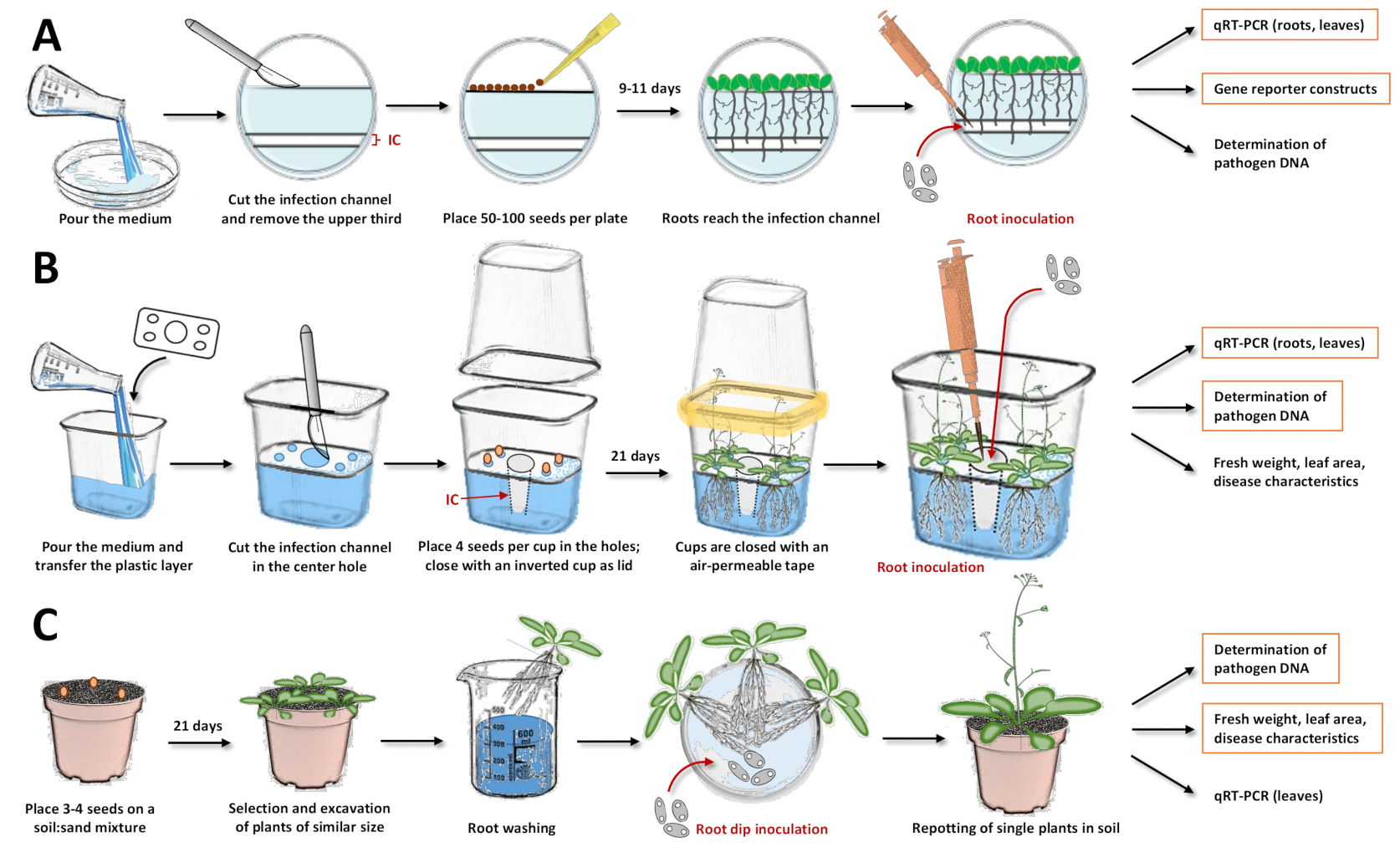
図1:3つの接種システムのコンパイルとプロトコルの個々のステップ。これらの図は、モデル植物シロイヌナズナとのシステムを示しています。他の植物種の場合、タイミングを調整する必要があります。オレンジ色のボックスが強調表示され、以降の解析がそれぞれのシステムで最も推奨されます。(a)シャーレ17への接種系については、培地を注ぎ、固化させる。皿は一晩冷蔵庫に保管してください。次に、上3分の1ならびに感染チャネル(IC)をメスで切断して除去する(図中の白い領域は寒天から除去し、青みがかった領域は寒天を表す)。切断面に種子を置き、ペトリ皿を閉じます。層別化後、プレートを垂直に置き、植物を成長させます。ほとんどの根が感染チャネルに到達したら、胞子溶液をピペットでチャネルに直接加えます。ソリューションが均等に分散されていることを確認します。ペトリ皿を閉じ、成長室で垂直にインキュベートする。従うことができるアプローチは、定量的逆転写PCR(qRT-PCR)による発現分析、レポーターラインによる顕微鏡法、および微生物DNAの定量化である。(b)プラスチックカップ19への接種系の場合、培地を注ぎ、分離プラスチック層を予め作製した穴(種子を配置するための角に4つの小さな穴、感染チャネルの中央に1つの大きな穴)で転写する。培地を固めましょう。中央の穴にある寒天培地をメスで切断除去し、感染チャネル(IC)を得た。小さな穴に媒体を傷つけ、種子を移します。カップを逆さのカップで閉じ、通気性テープ(黄色で表示)で密封します。植物を成長させましょう。接種のために、ピペットで胞子溶液を感染チャネルに直接加える。システムを閉じ、成長室で栽培を続ける。従うことができるアプローチは、qRT-PCRによる発現分析、微生物DNAの定量化、および新鮮な体重、葉面積、または他の疾患特性の決定である。(c)「根浸漬接種」15、17、23、24:土壌ベースの接種システムのために、土壌:砂混合物でポットを充填する。種を移し、苗を成長させます。同様のサイズの植物を発掘し、根を水で洗う。洗浄した根を胞子の入った溶液を保持するペトリ皿に入れる。インキュベーション後、単一の植物を土壌の入った鉢に挿入する。従うことができるアプローチは、qRT-PCRによる葉の発現分析、微生物DNAの定量、および新鮮な体重、葉面積、または他の疾患特性の決定である。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
Access restricted. Please log in or start a trial to view this content.
結果
プロトコールに従って、植物を栽培し、 V. longisporum ( Vl4325株)または V. dahliae (単離JR218)を接種した。有効性を証明し、特定のプロトコルのいくつかの機能を強調するために、さまざまなシナリオが設計されました。代表的な成果が示されています。
抗菌性インドール - グルコシノレート(IG)生合成に関与する遺伝子?...
Access restricted. Please log in or start a trial to view this content.
ディスカッション
土壌媒介性植物病原体1による収量損失が大きいため、農業戦略や作物品種の改善が求められています。土壌媒介性疾患の病因に関する限られた洞察は、より抵抗性の高い植物の発達を妨げる。根底にある病態メカニズムを探求する必要があり、そのためには堅牢な方法論的プラットフォームが必要である。報告された接種手順は、根−微生物相互作用における多因子事象が...
Access restricted. Please log in or start a trial to view this content.
開示事項
著者らは開示するものは何もありません。
謝辞
著者らは、これらの方法に関する以前の研究についてティム・イヴェンとジャクリーン・コモレク、この研究に必要な機器とリソースを提供したヴォルフガング・ドレーゲ・レーザー(ドイツ、ヴュルツブルク大学薬学部)、ヴォルフガング・ドレーゲ・レーザーとフィリップ・クライス(どちらもヴュルツブルク大学)のグループが原稿の批判的な校正を行ったことを認めている。この研究は「Deutsche Forschungsgemeinschaft」(DFG, DR273/15-1,2)の支援を受けた。
Access restricted. Please log in or start a trial to view this content.
資料
| Name | Company | Catalog Number | Comments |
| Agar (Gelrite) | Carl Roth | Nr. 0039 | all systems described require Gelrite |
| Arabidopsis thaliana wild-type | NASC stock | Col-0 (N1092) | |
| Autoclave | Systec | VE-100 | |
| BlattFlaeche | Datinf GmbH | BlattFlaeche | software to determine leaf areas |
| Brassica napus wild-type | see Floerl et al., 2008 | rapid-cycling rape | genome ACaacc |
| Cefotaxime sodium | Duchefa | C0111 | |
| Chicanery flask 500 mL | Duran Group / neoLab | E-1090 | Erlenmeyer flask with four baffles |
| Collection tubes 50 mL | Sarstedt | 62.547.254 | 114 x 28 mm |
| Czapek Dextrose Broth medium | Duchefa | C1714 | |
| Digital camera | Nikon | D3100 18-55 VR | |
| Exsiccator (Desiccator ) | Duran Group | 200 DN, 5.8 L | Seal with lid to hold chlorine gas |
| Fluorescence Microscope | Leica | Leica TCS SP5 II | |
| HCl | Carl Roth | P074.3 | |
| KNO3 | Carl Roth | P021.1 | ≥ 99 % |
| KOH | Carl Roth | 6751 | |
| Leukopor | BSN medical GmbH | 2454-00 AP | non-woven tape 2.5 cm x 9.2 m |
| MES (2-(N-morpholino)ethanesulfonic acid) | Carl Roth | 4256.2 | Pufferan ≥ 99 % |
| MgSO4 | Carl Roth | T888.1 | Magnesiumsulfate-Heptahydrate |
| Murashige & Skoog medium (MS) | Duchefa | M0222 | MS including vitamins |
| NaClO | Carl Roth | 9062.1 | |
| Percival growth chambers | CLF Plant Climatics GmbH | AR-66L2 | |
| Petri-dishes | Sarstedt | 82.1473.001 | size ØxH: 92 × 16 mm |
| Plastic cups (500 mL, transparent) | Pro-pac, salad boxx | 5070 | size: 108 × 81 × 102 mm |
| Pleated cellulose filter | Hartenstein | FF12 | particle retention level 8–12 μm |
| poly klima growth chamber | poly klima GmbH | PK 520 WLED | |
| Potato Dextrose Broth medium | SIGMA Aldrich | P6685 | for microbiology |
| Pots | Pöppelmann GmbH | TO 7 D or TO 9,5 D | Ø 7 cm resp. Ø 9.5 cm |
| PromMYB51::YFP | see Poncini et al., 2017 | MYB51 reporter line | YFP (i.e. 3xmVenus with NLS) |
| Reaction tubes 2 mL | Sarstedt | 72.695.400 | PCR Performance tested |
| Rotary (orbital) shaker | Edmund Bühler | SM 30 C control | |
| Sand (bird sand) | Pet Bistro, Müller Holding | 786157 | |
| Soil | Einheitserde spezial | SP Pikier (SP ED 63 P) | |
| Solanum lycopersicum wild-type | see Chavarro-Carrero et al., 2021 | Type: Moneymaker | |
| Thoma cell counting chamber | Marienfeld | 642710 | depth 0.020 mm; 0.0025 mm2 |
| Ultrapure water (Milli-Q purified water) | MERK | IQ 7003/7005 | water obtained after purification |
| Verticillium dahliae | see Reusche et al., 2014 | isolate JR2 | |
| Verticillium longisporum | Zeise and von Tiedemann, 2002 | strain Vl43 |
参考文献
- Mendes, R., Garbeva, P., Raaijmakers, J. M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiology Review. 37 (5), 634-663 (2013).
- Yadeta, K. A., Thomma, B. P. H. J. The xylem as battleground for plant hosts and vascular wilt pathogens. Frontiers in Plant Science. 4, 97(2013).
- Delgado-Baquerizo, M., et al. The proportion of soil-borne pathogens increases with warming at the global scale. Nature Climate Change. 10 (6), 550-554 (2020).
- Berendsen, R. L., et al. Disease-induced assemblage of a plant-beneficial bacterial consortium. The ISME Journal. 12 (6), 1496-1507 (2018).
- Yuan, J., et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome. 6 (1), 156(2018).
- Liu, H., et al. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytologist. 229 (5), 2873-2885 (2021).
- Wang, H., Liu, R., You, M. P., Barbetti, M. J., Chen, Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): role of bacterial diversity. Microorganisms. 9 (9), 1988(2021).
- Inderbitzin, P., Subbarao, K. V. Verticillium systematics and evolution: how confusion impedes Verticillium wilt management and how to resolve it. Phytopathology. 104 (6), 564-574 (2014).
- Eynck, C., Koopmann, B., Grunewaldt-Stoecker, G., Karlowsky, P., von Tiedemann, A. Differential interactions of Verticillium longisporum und V. dahliae with Brassica napus with molecular and histological techniques. European Journal of Plant Pathology. 118 (3), 259-274 (2007).
- Floerl, S., et al. Defence reactions in the apoplastic proteome of oilseed rape (Brassica napus var. napus) attenuate Verticillium longisporum growth but not disease symptoms. BMC Plant Biology. 8, 129(2008).
- Leonard, M., et al. Verticillium longisporum elicits media-dependent secretome responses with capacity to distinguish between plant-related environments. Frontiers in Microbiology. 11, 1876(2020).
- Depotter, J. R. L., et al. Verticillium longisporum, the invisible threat to oilseed rape and other brassicaceous plant hosts. Molecular Plant Pathology. 17 (7), 1004-1016 (2016).
- Fröschel, C., et al. A gain-of-function screen reveals redundant ERF transcription factors providing opportunities for resistance breeding toward the vascular fungal pathogen Verticillium longisporum. Molecular Plant-Microbe Interactions. 32 (9), 1095-1109 (2019).
- Zhou, L., Hu, Q., Johansson, A., Dixelius, C. Verticillium longisporum and V. dahliae: infection and disease in Brassica napus. Plant Pathology. 55 (1), 137-144 (2006).
- Ralhan, A., et al. The vascular pathogen Verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots. Plant Physiology. 159 (3), 1192-1203 (2012).
- Reusche, M., et al. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium longisporum. Molecular Plant-Microbe Interactions. 26 (8), 850-860 (2013).
- Iven, T., et al. Transcriptional activation and production of tryptophan-derived secondary metabolites in Arabidopsis roots contributes to the defense against the fungal vascular pathogen Verticillium longisporum. Molecular Plant. 5 (6), 1389-1402 (2012).
- Reusche, M., et al. Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana. Environmental and Experimental Botany. 108, 23-37 (2014).
- Fröschel, C. In-depth evaluation of root infection systems using the vascular fungus Verticillium longisporum as soil-borne model pathogen. Plant Methods. 17 (1), 57(2021).
- Karapapa, V. K., Bainbridge, B. W., Heale, J. B. Morphological and molecular characterization of Verticillium longisporum comb, nov., pathogenic to oilseed rape. Mycological Research. 101 (11), 1281-1294 (1997).
- Poncini, L., et al. In roots of Arabidopsis thaliana, the damage-associated molecular pattern AtPep1 is a stronger elicitor of immune signalling than flg22 or the chitin heptamer. PLoS One. 12 (10), 1-21 (2017).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Fradin, E. F., et al. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiology. 150 (1), 320-332 (2009).
- Singh, S., et al. The plant host Brassica napus induces in the pathogen Verticillium longisporum the expression of functional catalase peroxidase which is required for the late phase of disease. Molecular Plant-Microbe Interactions. 25 (4), 569-581 (2012).
- Zeise, K., von Tiedemann, A. Application of RAPD-PCR for virulence type analysis within Verticillium dahliae and Verticillium longisporum. Journal of Phytopathology. 150 (10), 557-563 (2002).
- Fröschel, C., et al. Plant roots employ cell-layer-specific programs to respond to pathogenic and beneficial microbes. Cell Host & Microbe. 29 (2), 299-310 (2021).
- Gigolashvili, T., et al. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. The Plant Journal. 50 (5), 886-901 (2007).
- Back, M. A., Haydock, P. P. J., Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathology. 51 (6), 683-697 (2002).
- Behrens, F. H., et al. Suppression of abscisic acid biosynthesis at the early infection stage of Verticillium longisporum in oilseed rape (Brassica napus). Molecular Plant Pathology. 20 (12), 1645-1661 (2019).
- Vorholt, J. A., Vogel, C., Carlström, C. I., Müller, D. B. Establishing causality: opportunities of synthetic communities for plant microbiome research. Cell Host & Microbe. 22 (2), 142-155 (2017).
Access restricted. Please log in or start a trial to view this content.
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved