Aby wyświetlić tę treść, wymagana jest subskrypcja JoVE. Zaloguj się lub rozpocznij bezpłatny okres próbny.
Method Article
Inoculation Strategies to Infect Plant Roots with Soil-Borne Microorganisms
W tym Artykule
Podsumowanie
This protocol presents a detailed summary of strategies to inoculate plant roots with soil-borne microbes. Exemplified for the fungi Verticillium longisporum and Verticillium dahliae, three different root infection systems are described. Potential applications and possible downstream analyses are highlighted, and advantages or disadvantages are discussed for each system.
Streszczenie
The rhizosphere harbors a highly complex microbial community in which plant roots are constantly challenged. Roots are in close contact with a wide variety of microorganisms, but studies on soil-borne interactions are still behind those performed on aboveground organs. Although some inoculation strategies for infecting model plants with model root pathogens are described in the literature, it remains difficult to get a comprehensive methodological overview. To address this problem, three different root inoculation systems are precisely described that can be applied to gain insights into the biology of root-microbe interactions. For illustration, Verticillium species (namely, V. longisporum and V. dahliae) were employed as root invading model pathogens. However, the methods can be easily adapted to other root colonizing microbes - both pathogenic and beneficial. By colonizing the plant xylem, vascular soil-borne fungi such as Verticillium spp. exhibit a unique lifestyle. After root invasion, they spread via the xylem vessels acropetally, reach the shoot, and elicit disease symptoms. Three representative plant species were chosen as model hosts: Arabidopsis thaliana, economically important oilseed rape (Brassica napus), and tomato (Solanum lycopersicum). Step-by-step protocols are given. Representative results of pathogenicity assays, transcriptional analyses of marker genes, and independent confirmations by reporter constructs are shown. Furthermore, the advantages and disadvantages of each inoculation system are thoroughly discussed. These proven protocols can assist in providing approaches for research questions on root-microbe interactions. Knowing how plants cope with microbes in the soil is crucial for developing new strategies to improve agriculture.
Wprowadzenie
Natural soils are inhabited by an astonishing number of microbes that can be neutral, harmful, or beneficial to plants1. Many plant pathogens are soil-borne, surround the roots, and attack the subterranean organ. These microorganisms belong to a wide variety of clades: fungi, oomycetes, bacteria, nematodes, insects, and some viruses1,2. Once environmental conditions favor infection, susceptible plants will become diseased and crop yields decline. The effects of climate change, such as global warming and weather extremes, will increase the proportion of soil-borne plant pathogens3. Therefore, it will become more and more important to study these destructive microbes and their impact on food and feed production, but also on natural ecosystems. Additionally, there are microbial mutualists in the soil that tightly interact with roots and promote plant growth, development, and immunity. When confronted with pathogens, plants can actively recruit specific opponents in the rhizosphere that can support host survival by suppressing pathogens4,5,6,7. However, mechanistic details and pathways involved in beneficial root-microbe interactions are often still unknown6.
It is, therefore, essential to expand the general understanding of root-microbe interactions. Reliable methods for inoculating roots with soil-borne microorganisms are necessary to perform model studies and transfer the findings to agricultural applications. Beneficial interactions in the soil are studied, for example, with Serendipita indica (formerly known as Piriformospora indica), nitrogen-fixing Rhizobium spp., or mycorrhizal fungi, while known soil-borne plant pathogens include Ralstonia solanacearum, Phytophthora spp., Fusarium spp., and Verticillium spp.1. The latter two are fungal genera that are globally distributed and cause vascular diseases2. Verticillium spp. (Ascomycota) can infect hundreds of plant species - largely dicotyledons, including herbaceous annuals, woody perennials, and many crop plants2,8. Hyphae of Verticillium enter the root and grow both intercellularly and intracellularly toward the central cylinder to colonize the xylem vessels2,9. In these vessels, the fungus remains for most of its life cycle. As the xylem sap is nutrient-poor and carries plant defense compounds, the fungus must adapt to this unique environment. This is accomplished by the secretion of colonization-related proteins that enable the pathogen to survive in its host10,11. After reaching the root vasculature, the fungus can spread within the xylem vessels acropetally to the foliage, which leads to systemic colonization of the host9,12. At this point, the plant is negatively affected in growth9,10,13. For instance, stunting and yellow leaves occur as well as premature senescence13,14,15,16.
One member of this genus is Verticillium longisporum, which is highly adapted to brassicaceous hosts, such as the agronomically important oilseed rape, cauliflower, and the model plant Arabidopsis thaliana12. Several studies combined V. longisporum and A. thaliana to gain extensive insights into soil-borne vascular diseases and the resulting root defense responses13,15,16,17. Straightforward susceptibility testing can be realized by using the V. longisporum / A. thaliana model system and well-established genetic resources are available for both organisms. Closely related to V. longisporum is the pathogen Verticillium dahliae. Although both fungal species perform a similar vascular life-style and invasion process, their propagation efficiency from roots to leaves and the elicited disease symptoms in A. thaliana are different: while V. longisporum usually induces early senescence, V. dahliae infection results in wilting18. Recently, a methodological summary presented different root inoculation strategies for infecting A. thaliana with V. longisporum or V. dahliae, assisting in planning experimental setups19. In the field, V. longisporum occasionally causes significant damage in oilseed rape production12, whereas V. dahliae has a very broad host range comprising several cultivated species, such as grapevine, potato, and tomato8. This makes both pathogens economically interesting models to study.
Thus, the following protocols use both V. longisporum and V. dahliae as model root pathogens to exemplify possible approaches for root inoculations. Arabidopsis (Arabidopsis thaliana), oilseed rape (Brassica napus), and tomato (Solanum lycopersicum) were chosen as model hosts. Detailed descriptions of the methodologies can be found in the text below and the accompanying video. Advantages and disadvantages for each inoculation system are discussed. Taken together, this protocol collection can help to identify a suitable method for specific research questions in the context of root-microbe interactions.
Protokół
1. Media for fungal cultures and plant inoculation systems
- Liquid Potato Dextrose Broth (PDB): Prepare 21 g/L PDB in ultrapure water in a heat-stable flask.
- Liquid Czapek Dextrose Broth (CDB): Prepare 42 g/L CDB in ultrapure water in a heat-stable flask.
- Medium for the Petri dish inoculation system: Prepare a heat-stable flask with 1.5 g/L Murashige and Skoog medium (MS) and 8 g/L agar in ultrapure water.
NOTE: Avoid sugar in this medium as it will lead to excessive fungal growth after inoculation. - Medium for the plastic-cup-based inoculation system: Prepare a heat-stable flask with 4.4 g/L MS, 0.2 g/L MgSO4, 1 g/L KNO3, 0.5 g/L 2-(N-morpholino)ethanesulfonic acid (MES), and 6.0 g/L agar in ultrapure water and adjust pH to 5.7 with 5 M KOH.
NOTE: Avoid sugar in this medium as it will lead to excessive fungal growth after inoculation. - ¼ MS medium: Prepare 1.2 g/L MS in ultrapure water.
- Use the autoclave to sterilize all the above solutions. Put the glass flasks into the basket, close the lid and sterilize for 15 min at 121 °C and 98.9 kPa.
2. Sterilizing the surface of plant seeds
NOTE: Use the below protocol always to sterilize the surface of seeds from Arabidopsis, oilseed rape, and tomato prior to sowing.
- Transfer the seeds into a 2 mL reaction tube. Place the tube in an exsiccator with an internal capacity of 5.8 L.
- Generate chlorine gas in the exsiccator by adding 6 mL of 33% hydrochloric acid (HCl) into 100 mL of 12% aqueous sodium hypochlorite (NaClO).
- Immediately close the lid of the exsiccator and incubate the seeds for 3 h in the gas.
3. Preparing the inoculum with Verticillium spores (asexual derived conidia)
NOTE: Cultivate V. dahliae (strain JR2) in the same way as V. longisporum (strain Vl43)17,18,19. Ensure that all the equipment and media are germ-free and that all steps are performed in a laminar flow hood to keep the inoculum axenic.
- Fill 150 mL of liquid PDB (step 1.1) in a 500 mL chicanery flask and supplement the medium with 500 mg/L cefotaxime.
- Add Verticillium conida from glycerol-stock storage to the PDB medium. Close the flask with a sterile foam stopper.
- Incubate the culture for 7-10 days in a dark box at room temperature (RT) under continuous, horizontal shaking (rotary shaker; 60 rpm). This results in small, white mycelia spheres.
- Remove and discard the PDB supernatant carefully. Most of the mycelia should remain in the flask.
- Add 100 mL of liquid CDB (step 1.2) on the mycelia in the chicanery flask and supplement the medium with 500 mg/L cefotaxime.
- Incubate another 4-5 days in a dark box at RT under continuous, horizontal shaking (rotary shaker, 60 rpm) to induce sporulation. The supernatant will turn yellowish-greyish as conidia are released.
- Filter a portion (5-10 mL) of the conidia-containing liquid through a filter paper (particle retention level of 8-12 µm) into a sterile 50 mL collection tube. This separates spores from mycelia.
- Determine the spore concentration by using a cell counting chamber and a microscope. Dilute with germ-free ¼ MS medium in ultrapure water until the spore concentrations given below are obtained.
NOTE: Under the microscope, the conidia from V. longisporum are mostly long-drawn and 7.1-8.8 µm in size, while V. dahliae conidia are shorter (3.5-5.5 µm) and rather spherical20. - Use these freshly harvested conidia as inoculum. Ensure to conduct the experiments always with freshly harvested conidia and not with frozen stocks, as freezing significantly reduces the number of viable spores19.
- For long-term storage, freeze the spores as high concentrated spore solution (approximately 1 x 108 spores/mL) in 25% glycerol at -80 °C (storable up to 1 year). For the next experiments, use these glycerol stocks to inoculate the PDB medium in step 3.2.
4. A sterile in vitro inoculation system based on Petri dishes
NOTE: For the Petri dish system17, ensure that all equipment and media are germ-free and that all steps are performed in a laminar flow hood.
- After autoclaving, pour the medium (see step 1.3) into Petri dishes.
- After the hardening of the medium, repack the Petri dishes in a sterile plastic bag and store them upside-down overnight in the refrigerator (4-10 °C). A chilled medium helps to prevent sliding of the medium in the next steps.
- Cut and remove an infection channel and the upper third of the solidified medium with a scalpel (Figure 1A). Avoid getting liquid or air under the agar medium while cutting; otherwise, the medium will slip and close the infection channel.
- Distribute 50-100 surface-sterilized Arabidopsis seeds with a sterile pipette tip on the cut upper surface. Put the seeds in the angle where the cut agar surface contacts the wall of the Petri dish so that roots can grow between the medium and the Petri dish wall. This will facilitate the inoculation later.
- Close the Petri dishes and seal them with air-permeable adhesive tape to allow gas exchange.
- After stratification for 2 days in the darkness at 4 °C, place the plates vertically in a suitable rack and grow the plants at 22 °C ± 1 °C under long-day conditions (16 h light / 8 h darkness) in a growth chamber.
- When the majority of the roots reach the infection channel (about 9-11-day old seedlings), lay the plates horizontally, open them and add 500 µL of freshly harvested Verticillium conidia with a concentration of 4 x 105 spores/mL directly into the infection channel, making sure that the liquid is evenly distributed in the channel.
- Similarly, prepare control plates by adding 500 µL of a mock solution instead of spores (germ-free ¼ MS medium).
- Incubate the plates horizontally for a couple of minutes until the liquid has soaked in and cannot leak out when the plates are set up vertically again. Then, close the lid and seal the plates with air-permeable adhesive tape.
- Incubate the plates vertically in the growth chamber. Optionally, cover the root parts with black paper-boxes to darken roots and soil-borne fungus (see19).
- Perform the analyses at the preferred time points after inoculation depending on the research question (refer to the figure legends for the exact time points used here). Following are some suggestions.
- Cut the leaves from the roots and harvest both separately. Take the agar strips out of the Petri dishes to easily access the roots and carefully pull them out of the agar using forceps. Freeze all plant material immediately in liquid nitrogen.
- Grind the samples in liquid nitrogen. Extract total DNA from 100 mg of leaf material to determine via a quantitative PCR (qPCR) the amount of fungal DNA relative to plant DNA (see19).
- Grind the samples in liquid nitrogen. Take 100 mg of plant material and extract total RNA. Conduct quantitative reverse transcription PCR (qRT-PCR) to determine the expression of plant genes (or fungal genes) during infestation (see19).
- Carefully remove the roots from the agar avoiding injury and examine them under the fluorescent microscope.
- Determine induction of marker genes in plant reporter lines (e.g., luciferase, β-glucuronidase, or fluorescent reporters17,19,21).
- Visualize fungal propagation at the root by using fungal reporter lines (e.g., V. longisporum constitutively expressing enhanced Green Fluorescent Protein, Vl-sGFP9) or by staining techniques (e.g., through 5-bromo-4-chloro-3-indoxyl-N-acetyl-beta-d-glucosaminide (X-beta-D-Glc-Nac)18).
- Cut the leaves from the roots and harvest both separately. Take the agar strips out of the Petri dishes to easily access the roots and carefully pull them out of the agar using forceps. Freeze all plant material immediately in liquid nitrogen.
5. A sterile in vitro inoculation system organized with plastic cups
NOTE: As noted in the first description of this technique19, ensure that all equipment and media are germ-free and that all steps are performed in a laminar flow hood.
- Utilize transparent plastic cups with a total volume of 500 mL and sterilize them in a 70%-75% ethanol bath for at least 20 min. Dry the cups in the laminar flow hood.
- Pour the autoclaved medium (see step 1.4) into the plastic cups. Optionally, add cefotaxime (final concentration of 50 mg/L) to the autoclaved medium to prevent bacterial contaminations. Use 150 mL of medium per cup for experiments with Arabidopsis or more medium (250-300 mL per cup) for experiments with larger plant species (oilseed rape, tomato).
- Place a plastic layer (sterilized before by incubating in 70%-75% ethanol for 20 min) on the medium before it solidifies (Figure 1B).
NOTE: This plastic layer contains four prefabricated holes at the corners for placing surface-sterilized seeds. This allows the seeds to access the medium. Later, this separating layer prevents the leaves from touching the fungus-containing medium, so that the microbes cannot directly attack the leaves and must take the root pathway. Another hole is in the center, enabling cutting the infection channel. - When the medium has solidified, cut the agar with a scalpel through the prefabricated center hole to a depth of about 1.5 cm. Remove the cut agar to create an infection channel into which the fungal spores can be added later.
- Slightly scratch the agar medium with a pipette tip in the four smaller holes to interrupt the solidified skin (this allows the seeds to soak up water from the watery agar medium). Place the seeds using a pipette tip into the smaller holes.
- Close the plastic cup with a second, inverted plastic cup and seal with air-permeable adhesive tape. The tape must allow gas exchange.
- After stratification for 3 days in darkness at 4 °C, incubate the cup systems under 12 h light / 12 h darkness (Arabidopsis, oilseed rape) or 16 h light / 8 h darkness conditions (tomato) in growth chambers at a constant temperature of 22 °C and 60% humidity.
- Follow the recommended age of plants for inoculation: 21 days for Arabidopsis; 5-7 days for oilseed rape; 12 days for tomato.
- Inoculate plantlets with Verticillium by adding 1 mL of conidia solution (recommended concentration: 4 x 105 spores/mL) into the infection channel. To prepare control samples, add 1 mL of mock solution without spores (germ-free ¼ MS medium) into the channel.
- Perform the analyses at the preferred time points after inoculation depending on the research question (refer to the figure legends for the exact time points used here). Following are some suggestions.
- Take photographs of the plants with a digital camera from above keeping the distance the same for each photo. Quantify the leaf area (e.g., with ImageJ22 or BlattFlaeche17,19; use the length of the cups to set the scale) and compare infected and control groups. Categorize the development of disease symptoms (e.g., smaller, more yellowish, or necrotic leaves).
NOTE: If there are any stems on Arabidopsis, remove them to get better photos of the rosettes. - Remove the roots and define the biomass (fresh weight) of shoots from infected and control samples by weighing. Determine the relative fresh weight19.
- Collect the samples for molecular analyses as follows.
- Arabidopsis: Remove stems if there are any. Cut the rosettes at the base from the roots. Make sure to exclude all root material from the sample and harvest whole rosettes. Combine 4-5 rosettes from different plants into one sample and freeze the leaf material in liquid nitrogen.
- Pull the roots carefully out of the medium with forceps, press and dab them with a paper towel to remove agar remains, and combine 4-5 roots from different plants into one sample. Freeze immediately in liquid nitrogen.
- Oilseed rape/tomato: Cut stem segments from the hypocotyl (e.g., 1 cm in length; always take the same stem region). Combine material from 4-5 plants into each sample and freeze in liquid nitrogen.
- Grind the samples in liquid nitrogen. Extract total DNA from 100 mg of the leaf or stem material to determine via qPCR the amount of fungal DNA relative to plant DNA (see19).
- Grind the samples in liquid nitrogen, take 100 mg of plant material and extract total RNA. Conduct qRT-PCR to determine the expression of plant genes (or fungal genes) upon infestation (see19).
- Take photographs of the plants with a digital camera from above keeping the distance the same for each photo. Quantify the leaf area (e.g., with ImageJ22 or BlattFlaeche17,19; use the length of the cups to set the scale) and compare infected and control groups. Categorize the development of disease symptoms (e.g., smaller, more yellowish, or necrotic leaves).
6. A soil-based inoculation system in pots
- Thoroughly mix soil and sand in a 3:1 (soil:sand) volumetric ratio to facilitate washing the substrate off the roots. Pour the mixture into an autoclave bag. If the mixture is too dry, add an appropriate amount of water and mix it into the substrate. Steam at 80 °C for 20 min in an autoclave to minimize microbial contaminations.
NOTE: Avoid heating to over 80 °C, as this may affect organic soil nutrients. - Fill pots with the soil-sand mixture and transfer them into trays. Add water into the trays about 1/3 the height of a pot, so that the soil-sand mixture gets thoroughly soaked with water. Additionally, water-spray the substrate with a spray bottle to ensure wet starting conditions.
- Sow 3-4 seeds in each pot (Figure 1C) ensuring that the seeds have enough distance from each other. Keep them for 3 days in darkness at 4 °C for stratification to synchronize germination.
NOTE: Pre-cultivate an excess of plants, which enables a selection of plants of similar size for the inoculation experiments and reduces deviations due to individual differences. - Let the seedlings grow under long-day conditions (16 h light / 8 h darkness; constant temperature of 22 °C; 60% humidity) with regular watering.
- Follow the recommended age of plants for inoculation: 21 days for Arabidopsis, 7 days for oilseed rape, and 10 days for tomato. Pick plants of similar size to perform the "root dip inoculation"15,17,23,24. Take the soil out of the pots and carefully excavate the roots.
- Gently wash only the roots in a water container and keep the rosettes out of the water. Incubate the washed roots for 60 min in a Petri dish containing the Verticillium spore solution (recommended concentration: 2 x 106 spores/mL). For the non-infected control group, incubate the roots for 60 min in the mock solution without spores (germ-free ¼ MS medium).
- Prepare new pots with moist, steam-sterilized soil (80 °C for 20 min) without sand. Use a pipette tip to make one hole in the center of the soil in each pot.
- Directly place the roots into the hole (transfer only one plant per pot). After inserting the roots, make sure to refill the holes carefully with soil. Avoid pressing the soil, otherwise repotting can cause stress symptoms such as purple leaves.
- Cultivate infected and control groups under long-day conditions (16 h light / 8 h darkness; a constant temperature of 22 °C; 60% humidity) with regular watering.
- Perform the analyses at the preferred time points after inoculation depending on the research question (refer to the figure legends for the exact time points used here). Following are some suggestions.
- Take photographs of the plants with a digital camera from above, keeping the distance the same for each photo. Quantify the leaf area (e.g., with ImageJ22 or BlattFlaeche17,19; use the diameter of the pots to set the scale) and compare infected and control groups. Categorize the development of disease symptoms (e.g., smaller, more yellowish, or necrotic leaves)13.
NOTE: Removing stems of Arabidopsis facilitates taking photos of the rosettes. - Remove the roots and define the biomass (fresh weight) of shoots from infected and control samples by weighing. Determine the relative fresh weight19.
- Alternatively, measure the plant height or categorize fungal outgrowth from stem segments to evaluate disease severity13.
- Collect samples for molecular analyses as follows.
- Arabidopsis: Remove the stems. Cut the rosettes at the root crown. Combine 4-5 rosettes from different plants into one sample. Freeze the leaf material in liquid nitrogen.
NOTE: In the case of roots, it is difficult to clean them sufficiently from the soil without reprogramming gene expression through washing. - Oilseed rape/tomato: Cut stem segments from the hypocotyl (e.g., 1 cm in length; always take the same stem region). Combine material from 4-5 plants into one sample and freeze it in liquid nitrogen.
- Grind the samples in liquid nitrogen. Extract total DNA from 100 mg leaf or stem material to determine via qPCR the amount of fungal DNA relative to plant DNA (see19).
- Grind the samples in liquid nitrogen, take 100 mg of plant material and extract total RNA. Conduct qRT-PCR to determine the expression of plant genes (or fungal genes) upon infestation (see19).
- Take photographs of the plants with a digital camera from above, keeping the distance the same for each photo. Quantify the leaf area (e.g., with ImageJ22 or BlattFlaeche17,19; use the diameter of the pots to set the scale) and compare infected and control groups. Categorize the development of disease symptoms (e.g., smaller, more yellowish, or necrotic leaves)13.
7. Analyzing the data
- Calculate the average and standard deviation (± SD) based on the biological replicates.
- Calculate the relative values by dividing all the results from the infected group by the result of the control. Display the average as, for example, "relative to mock" or "relative to wild-type".
- Determine statistical significance between groups.

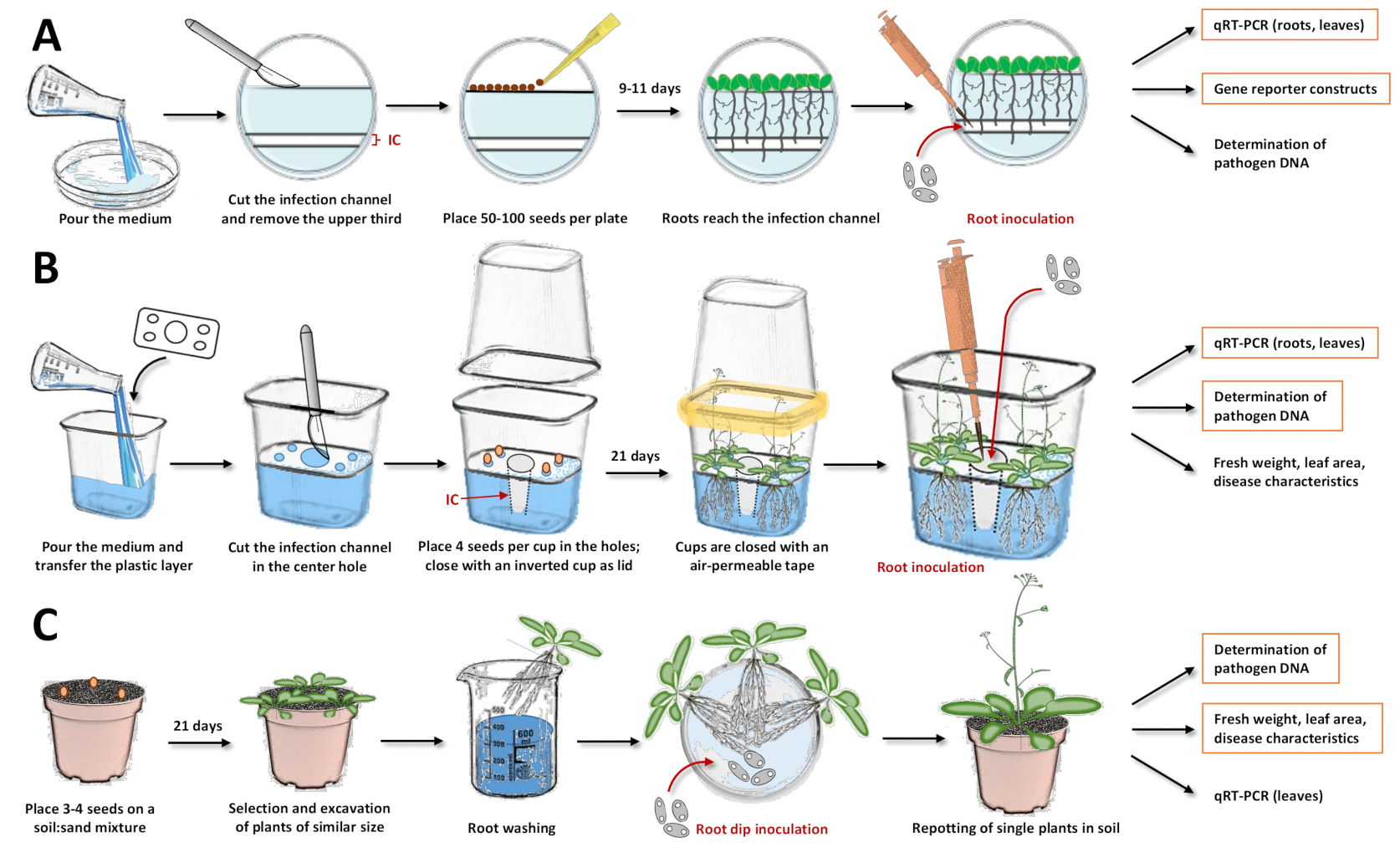
Figure 1: Compilation of the three inoculation systems and individual steps in the protocols. These figures illustrate the systems with the model plant Arabidopsis thaliana. For other plant species, the timing must be adjusted. Orange boxes highlight, for which subsequent analyses are most recommended with the respective system. (A) For the inoculation system in Petri dishes17, pour the medium and let it solidify. Keep the plates in the fridge overnight. Then, cut and remove the upper third as well as the infection channel (IC) with a scalpel (white areas in the illustration were removed from the agar, while bluish areas represent the agar). Place the seeds on the cut surface and close the Petri dishes. After stratification, place the plates vertically and let the plants grow. Once most of the roots have reached the infection channel, add the spore solution with a pipette directly into the channel. Make sure that the solution is evenly distributed. Close the Petri dishes and incubate them vertically in a growth chamber. Approaches that may follow are expressional analysis with quantitative reverse transcription PCR (qRT-PCR), microscopy with reporter lines, and quantification of microbial DNA. (B) For the inoculation system in plastic cups19, pour the medium and transfer the separating plastic layer with the prefabricated holes (four small holes in the corners for placing the seeds and one large hole in the center for the infection channel). Let the medium solidify. Cut and remove the agar medium in the center hole with a scalpel to obtain the infection channel (IC). Scratch the medium in the smaller holes and transfer the seeds. Close the cup with an inverted cup and seal with air-permeable tape (symbolized in yellow). Let the plants grow. For inoculation, add the spore solution with a pipette directly into the infection channel. Close the system and continue cultivation in the growth chamber. Approaches that may follow are expressional analysis with qRT-PCR, quantification of microbial DNA, and determination of fresh weight, leaf area, or other disease characteristics. (C) "Root dip inoculation"15,17,23,24: for the soil-based inoculation system, fill pots with a soil:sand mixture. Transfer the seeds and let the seedlings grow. Excavate plants of similar size and wash the roots in water. Place the washed roots in a Petri dish holding the solution with the spores. After incubation, insert single plants in pots with soil. Approaches that may follow are expressional analysis in leaves with qRT-PCR, quantification of microbial DNA, and determination of fresh weight, leaf area, or other disease characteristics. Please click here to view a larger version of this figure.
{kind=link}
Wyniki
Following the protocol, the plants were cultivated and inoculated with V. longisporum (strain Vl4325) or V. dahliae (isolate JR218). Various scenarios were designed to prove the effectiveness and to highlight some capabilities of the given protocols. Representative outcomes are shown.
Expressional induction of genes involved in the antimicrobial indol-glucosinolate (IG) biosynthesis is a reliable indicator for the evalu...
Dyskusje
Due to the tremendous yield losses caused by soil-borne phytopathogens1, an improvement of farming strategies or crop varieties is required. The limited insight into the pathogenesis of soil-borne diseases hinders the development of more resistant plants. Underlying pathomechanisms need to be explored, for which a robust methodological platform is required. Reported inoculation procedures have shown that multifactorial events in root-microbe interactions can be well dissected by combining differen...
Ujawnienia
The authors have nothing to disclose.
Podziękowania
The authors acknowledge Tim Iven and Jaqueline Komorek for previous work on these methods, the group of Wolfgang Dröge-Laser (Department of Pharmaceutical Biology, University of Würzburg, Germany) for providing the equipment and the resources needed for this work, and Wolfgang Dröge-Laser as well as Philipp Kreisz (both University of Würzburg) for critical proofreading of the manuscript. This study was supported by the "Deutsche Forschungsgemeinschaft" (DFG, DR273/15-1,2).
Materiały
| Name | Company | Catalog Number | Comments |
| Agar (Gelrite) | Carl Roth | Nr. 0039 | all systems described require Gelrite |
| Arabidopsis thaliana wild-type | NASC stock | Col-0 (N1092) | |
| Autoclave | Systec | VE-100 | |
| BlattFlaeche | Datinf GmbH | BlattFlaeche | software to determine leaf areas |
| Brassica napus wild-type | see Floerl et al., 2008 | rapid-cycling rape | genome ACaacc |
| Cefotaxime sodium | Duchefa | C0111 | |
| Chicanery flask 500 mL | Duran Group / neoLab | E-1090 | Erlenmeyer flask with four baffles |
| Collection tubes 50 mL | Sarstedt | 62.547.254 | 114 x 28 mm |
| Czapek Dextrose Broth medium | Duchefa | C1714 | |
| Digital camera | Nikon | D3100 18-55 VR | |
| Exsiccator (Desiccator ) | Duran Group | 200 DN, 5.8 L | Seal with lid to hold chlorine gas |
| Fluorescence Microscope | Leica | Leica TCS SP5 II | |
| HCl | Carl Roth | P074.3 | |
| KNO3 | Carl Roth | P021.1 | ≥ 99 % |
| KOH | Carl Roth | 6751 | |
| Leukopor | BSN medical GmbH | 2454-00 AP | non-woven tape 2.5 cm x 9.2 m |
| MES (2-(N-morpholino)ethanesulfonic acid) | Carl Roth | 4256.2 | Pufferan ≥ 99 % |
| MgSO4 | Carl Roth | T888.1 | Magnesiumsulfate-Heptahydrate |
| Murashige & Skoog medium (MS) | Duchefa | M0222 | MS including vitamins |
| NaClO | Carl Roth | 9062.1 | |
| Percival growth chambers | CLF Plant Climatics GmbH | AR-66L2 | |
| Petri-dishes | Sarstedt | 82.1473.001 | size ØxH: 92 × 16 mm |
| Plastic cups (500 mL, transparent) | Pro-pac, salad boxx | 5070 | size: 108 × 81 × 102 mm |
| Pleated cellulose filter | Hartenstein | FF12 | particle retention level 8–12 μm |
| poly klima growth chamber | poly klima GmbH | PK 520 WLED | |
| Potato Dextrose Broth medium | SIGMA Aldrich | P6685 | for microbiology |
| Pots | Pöppelmann GmbH | TO 7 D or TO 9,5 D | Ø 7 cm resp. Ø 9.5 cm |
| PromMYB51::YFP | see Poncini et al., 2017 | MYB51 reporter line | YFP (i.e. 3xmVenus with NLS) |
| Reaction tubes 2 mL | Sarstedt | 72.695.400 | PCR Performance tested |
| Rotary (orbital) shaker | Edmund Bühler | SM 30 C control | |
| Sand (bird sand) | Pet Bistro, Müller Holding | 786157 | |
| Soil | Einheitserde spezial | SP Pikier (SP ED 63 P) | |
| Solanum lycopersicum wild-type | see Chavarro-Carrero et al., 2021 | Type: Moneymaker | |
| Thoma cell counting chamber | Marienfeld | 642710 | depth 0.020 mm; 0.0025 mm2 |
| Ultrapure water (Milli-Q purified water) | MERK | IQ 7003/7005 | water obtained after purification |
| Verticillium dahliae | see Reusche et al., 2014 | isolate JR2 | |
| Verticillium longisporum | Zeise and von Tiedemann, 2002 | strain Vl43 |
Odniesienia
- Mendes, R., Garbeva, P., Raaijmakers, J. M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiology Review. 37 (5), 634-663 (2013).
- Yadeta, K. A., Thomma, B. P. H. J. The xylem as battleground for plant hosts and vascular wilt pathogens. Frontiers in Plant Science. 4, 97 (2013).
- Delgado-Baquerizo, M., et al. The proportion of soil-borne pathogens increases with warming at the global scale. Nature Climate Change. 10 (6), 550-554 (2020).
- Berendsen, R. L., et al. Disease-induced assemblage of a plant-beneficial bacterial consortium. The ISME Journal. 12 (6), 1496-1507 (2018).
- Yuan, J., et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome. 6 (1), 156 (2018).
- Liu, H., et al. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytologist. 229 (5), 2873-2885 (2021).
- Wang, H., Liu, R., You, M. P., Barbetti, M. J., Chen, Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): role of bacterial diversity. Microorganisms. 9 (9), 1988 (2021).
- Inderbitzin, P., Subbarao, K. V. Verticillium systematics and evolution: how confusion impedes Verticillium wilt management and how to resolve it. Phytopathology. 104 (6), 564-574 (2014).
- Eynck, C., Koopmann, B., Grunewaldt-Stoecker, G., Karlowsky, P., von Tiedemann, A. Differential interactions of Verticillium longisporum und V. dahliae with Brassica napus with molecular and histological techniques. European Journal of Plant Pathology. 118 (3), 259-274 (2007).
- Floerl, S., et al. Defence reactions in the apoplastic proteome of oilseed rape (Brassica napus var. napus) attenuate Verticillium longisporum growth but not disease symptoms. BMC Plant Biology. 8, 129 (2008).
- Leonard, M., et al. Verticillium longisporum elicits media-dependent secretome responses with capacity to distinguish between plant-related environments. Frontiers in Microbiology. 11, 1876 (2020).
- Depotter, J. R. L., et al. Verticillium longisporum, the invisible threat to oilseed rape and other brassicaceous plant hosts. Molecular Plant Pathology. 17 (7), 1004-1016 (2016).
- Fröschel, C., et al. A gain-of-function screen reveals redundant ERF transcription factors providing opportunities for resistance breeding toward the vascular fungal pathogen Verticillium longisporum. Molecular Plant-Microbe Interactions. 32 (9), 1095-1109 (2019).
- Zhou, L., Hu, Q., Johansson, A., Dixelius, C. Verticillium longisporum and V. dahliae: infection and disease in Brassica napus. Plant Pathology. 55 (1), 137-144 (2006).
- Ralhan, A., et al. The vascular pathogen Verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots. Plant Physiology. 159 (3), 1192-1203 (2012).
- Reusche, M., et al. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium longisporum. Molecular Plant-Microbe Interactions. 26 (8), 850-860 (2013).
- Iven, T., et al. Transcriptional activation and production of tryptophan-derived secondary metabolites in Arabidopsis roots contributes to the defense against the fungal vascular pathogen Verticillium longisporum. Molecular Plant. 5 (6), 1389-1402 (2012).
- Reusche, M., et al. Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana. Environmental and Experimental Botany. 108, 23-37 (2014).
- Fröschel, C. In-depth evaluation of root infection systems using the vascular fungus Verticillium longisporum as soil-borne model pathogen. Plant Methods. 17 (1), 57 (2021).
- Karapapa, V. K., Bainbridge, B. W., Heale, J. B. Morphological and molecular characterization of Verticillium longisporum comb, nov., pathogenic to oilseed rape. Mycological Research. 101 (11), 1281-1294 (1997).
- Poncini, L., et al. In roots of Arabidopsis thaliana, the damage-associated molecular pattern AtPep1 is a stronger elicitor of immune signalling than flg22 or the chitin heptamer. PLoS One. 12 (10), 1-21 (2017).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Fradin, E. F., et al. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiology. 150 (1), 320-332 (2009).
- Singh, S., et al. The plant host Brassica napus induces in the pathogen Verticillium longisporum the expression of functional catalase peroxidase which is required for the late phase of disease. Molecular Plant-Microbe Interactions. 25 (4), 569-581 (2012).
- Zeise, K., von Tiedemann, A. Application of RAPD-PCR for virulence type analysis within Verticillium dahliae and Verticillium longisporum. Journal of Phytopathology. 150 (10), 557-563 (2002).
- Fröschel, C., et al. Plant roots employ cell-layer-specific programs to respond to pathogenic and beneficial microbes. Cell Host & Microbe. 29 (2), 299-310 (2021).
- Gigolashvili, T., et al. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. The Plant Journal. 50 (5), 886-901 (2007).
- Back, M. A., Haydock, P. P. J., Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathology. 51 (6), 683-697 (2002).
- Behrens, F. H., et al. Suppression of abscisic acid biosynthesis at the early infection stage of Verticillium longisporum in oilseed rape (Brassica napus). Molecular Plant Pathology. 20 (12), 1645-1661 (2019).
- Vorholt, J. A., Vogel, C., Carlström, C. I., Müller, D. B. Establishing causality: opportunities of synthetic communities for plant microbiome research. Cell Host & Microbe. 22 (2), 142-155 (2017).
Przedruki i uprawnienia
Zapytaj o uprawnienia na użycie tekstu lub obrazów z tego artykułu JoVE
Zapytaj o uprawnieniaThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Wszelkie prawa zastrzeżone