È necessario avere un abbonamento a JoVE per visualizzare questo. Accedi o inizia la tua prova gratuita.
Method Article
Strategie di inoculazione per infettare le radici delle piante con microrganismi trasmessi dal suolo
In questo articolo
Riepilogo
Questo protocollo presenta un riepilogo dettagliato delle strategie per inoculare le radici delle piante con microbi trasmessi dal suolo. Esemplificato per i funghi Verticillium longisporum e Verticillium dahliae, vengono descritti tre diversi sistemi di infezione delle radici. Vengono evidenziate le potenziali applicazioni e le possibili analisi a valle e vengono discussi vantaggi o svantaggi per ciascun sistema.
Abstract
La rizosfera ospita una comunità microbica altamente complessa in cui le radici delle piante sono costantemente sfidate. Le radici sono a stretto contatto con un'ampia varietà di microrganismi, ma gli studi sulle interazioni trasmesse dal suolo sono ancora indietro rispetto a quelli eseguiti su organi fuori terra. Sebbene alcune strategie di inoculazione per infettare piante modello con patogeni radicali modello siano descritte in letteratura, rimane difficile ottenere una panoramica metodologica completa. Per affrontare questo problema, vengono descritti con precisione tre diversi sistemi di inoculazione delle radici che possono essere applicati per ottenere informazioni sulla biologia delle interazioni radice-microbo. Per esempio, le specie di Verticillium (vale a dire, V. longisporum e V. dahliae) sono state impiegate come patogeni modello di invasione delle radici. Tuttavia, i metodi possono essere facilmente adattati ad altri microbi colonizzanti le radici, sia patogeni che benefici. Colonizzando lo xilema vegetale, i funghi vascolari trasmessi dal suolo come il Verticillium spp. esibiscono uno stile di vita unico. Dopo l'invasione delle radici, si diffondono attraverso i vasi xilemici acropetalmente, raggiungono il germoglio e suscitano sintomi di malattia. Tre specie vegetali rappresentative sono state scelte come ospiti modello: Arabidopsis thaliana, colza economicamente importante (Brassica napus) e pomodoro (Solanum lycopersicum). Vengono forniti protocolli passo-passo. Vengono mostrati risultati rappresentativi di saggi di patogenicità, analisi trascrizionali di geni marcatori e conferme indipendenti da parte di costrutti reporter. Inoltre, i vantaggi e gli svantaggi di ciascun sistema di inoculazione sono discussi in modo approfondito. Questi protocolli collaudati possono aiutare a fornire approcci per domande di ricerca sulle interazioni radice-microbo. Sapere come le piante affrontano i microbi nel terreno è fondamentale per sviluppare nuove strategie per migliorare l'agricoltura.
Introduzione
I terreni naturali sono abitati da un numero sorprendente di microbi che possono essere neutri, dannosi o benefici per le piante1. Molti agenti patogeni delle piante sono trasmessi dal suolo, circondano le radici e attaccano l'organo sotterraneo. Questi microrganismi appartengono a un'ampia varietà di cladi: funghi, oomiceti, batteri, nematodi, insetti e alcuni virus 1,2. Una volta che le condizioni ambientali favoriscono l'infezione, le piante sensibili si ammaleranno e le rese delle colture diminuiranno. Gli effetti dei cambiamenti climatici, come il riscaldamento globale e gli estremi meteorologici, aumenteranno la percentuale di patogeni vegetali trasmessi dal suolo3. Pertanto, diventerà sempre più importante studiare questi microbi distruttivi e il loro impatto sulla produzione di alimenti e mangimi, ma anche sugli ecosistemi naturali. Inoltre, ci sono mutualisti microbici nel terreno che interagiscono strettamente con le radici e promuovono la crescita, lo sviluppo e l'immunità delle piante. Di fronte agli agenti patogeni, le piante possono reclutare attivamente avversari specifici nella rizosfera che possono supportare la sopravvivenza dell'ospite sopprimendo i patogeni 4,5,6,7. Tuttavia, i dettagli meccanicistici e i percorsi coinvolti nelle interazioni benefiche radice-microbo sono spesso ancora sconosciuti6.
È quindi essenziale espandere la comprensione generale delle interazioni radice-microbo. Sono necessari metodi affidabili per inoculare radici con microrganismi trasmessi dal suolo per eseguire studi modello e trasferire i risultati alle applicazioni agricole. Le interazioni benefiche nel suolo sono studiate, ad esempio, con Serendipita indica (precedentemente nota come Piriformospora indica), Rhizobium spp. che fissa l'azoto o funghi micorrizici, mentre noti patogeni vegetali trasmessi dal suolo includono Ralstonia solanacearum, Phytophthora spp., Fusarium spp. e Verticillium spp.1. Gli ultimi due sono generi fungini che sono distribuiti a livello globale e causano malattie vascolari2. Verticillium spp. (Ascomycota) può infettare centinaia di specie vegetali - in gran parte dicotiledoni, tra cui annuali erbacee, piante perenni legnose e molte piante coltivate 2,8. Le Ife di Verticillium entrano nella radice e crescono sia intercellulari che intracellulari verso il cilindro centrale per colonizzare i vasi xilema 2,9. In questi vasi, il fungo rimane per la maggior parte del suo ciclo vitale. Poiché la linfa dello xilema è povera di nutrienti e trasporta composti di difesa delle piante, il fungo deve adattarsi a questo ambiente unico. Ciò si ottiene con la secrezione di proteine correlate alla colonizzazione che consentono all'agente patogeno di sopravvivere nel suo ospite10,11. Dopo aver raggiunto la vascolarizzazione radicale, il fungo può diffondersi all'interno dei vasi xilema acropetalmente al fogliame, il che porta alla colonizzazione sistemica dell'ospite 9,12. A questo punto, la pianta è influenzata negativamente nella crescita 9,10,13. Ad esempio, si verificano arresto della crescita e foglie gialle e senescenza prematura 13,14,15,16.
Un membro di questo genere è verticillium longisporum, che è altamente adattato agli ospiti brassicacei, come la colza agronomicamente importante, il cavolfiore e la pianta modello Arabidopsis thaliana12. Diversi studi hanno combinato V. longisporum e A. thaliana per ottenere ampie informazioni sulle malattie vascolari trasmesse dal suolo e sulle conseguenti risposte di difesa delle radici 13,15,16,17. Semplici test di suscettibilità possono essere realizzati utilizzando il sistema modello V. longisporum / A. thaliana e risorse genetiche consolidate sono disponibili per entrambi gli organismi. Strettamente correlato a V. longisporum è l'agente patogeno Verticillium dahliae. Sebbene entrambe le specie fungine svolgano uno stile di vita vascolare e un processo di invasione simili, la loro efficienza di propagazione dalle radici alle foglie e i sintomi della malattia suscitati in A. thaliana sono diversi: mentre V. longisporum di solito induce la senescenza precoce, l'infezione da V. dahliae provoca l'appassimento18. Recentemente, un riassunto metodologico ha presentato diverse strategie di inoculazione radicale per infettare A. thaliana con V. longisporum o V. dahliae, assistendo nella pianificazione di configurazioni sperimentali19. Nel campo, V. longisporum causa occasionalmente danni significativi nella produzione di colza12, mentre V. dahliae ha una gamma di ospiti molto ampia che comprende diverse specie coltivate, come vite, patate e pomodoro8. Ciò rende entrambi i patogeni modelli economicamente interessanti da studiare.
Pertanto, i seguenti protocolli utilizzano sia V. longisporum che V. dahliae come patogeni radicali modello per esemplificare possibili approcci per le inoculazioni radicali. Arabidopsis (Arabidopsis thaliana), colza (Brassica napus) e pomodoro (Solanum lycopersicum) sono stati scelti come ospiti modello. Descrizioni dettagliate delle metodologie sono disponibili nel testo sottostante e nel video di accompagnamento. Vengono discussi vantaggi e svantaggi per ciascun sistema di inoculazione. Nel complesso, questa raccolta di protocolli può aiutare a identificare un metodo adatto per domande di ricerca specifiche nel contesto delle interazioni radice-microbo.
Protocollo
1. Mezzi per colture fungine e sistemi di inoculazione vegetale
- Brodo di destrosio di patate liquide (PDB): preparare 21 g/L di PDB in acqua ultrapura in un matraccio termostabile.
- Brodo liquido di destrosio Czapek (CDB): Preparare 42 g/L CDB in acqua ultrapura in un matraccio termostabile.
- Mezzo per il sistema di inoculazione della capsula di Petri: preparare un pallone termostabile con 1,5 g/L di Murashige e Skoog medium (MS) e 8 g/L di agar in acqua ultrapura.
NOTA: Evitare lo zucchero in questo mezzo in quanto porterà a un'eccessiva crescita fungina dopo l'inoculazione. - Mezzo per il sistema di inoculazione a base di bicchieri di plastica: preparare un matraccio termostabile con 4,4 g/L MS, 0,2 g/L MgSO4, 1 g/L KNO3, 0,5 g/L 2-(N-morfolino)etanosolfonico (MES) e 6,0 g/L di agar in acqua ultrapura e regolare il pH a 5,7 con 5 M KOH.
NOTA: Evitare lo zucchero in questo mezzo in quanto porterà a un'eccessiva crescita fungina dopo l'inoculazione. - 1/4 MS medium: Preparare 1,2 g/L MS in acqua ultrapura.
- Utilizzare l'autoclave per sterilizzare tutte le soluzioni di cui sopra. Mettere i palloni di vetro nel cestello, chiudere il coperchio e sterilizzare per 15 minuti a 121 °C e 98,9 kPa.
2. Sterilizzare la superficie dei semi delle piante
NOTA: Utilizzare sempre il protocollo sottostante per sterilizzare la superficie dei semi di Arabidopsis, colza e pomodoro prima della semina.
- Trasferire i semi in un tubo di reazione da 2 ml. Posizionare il tubo in un essiccatore con una capacità interna di 5,8 L.
- Generare gas di cloro nell'essiccatore aggiungendo 6 mL di acido cloridrico al 33% (HCl) in 100 ml di ipoclorito di sodio acquoso al 12% (NaClO).
- Chiudere immediatamente il coperchio dell'essiccatore e incubare i semi per 3 ore nel gas.
3. Preparazione dell'inoculo con spore di Verticillium (conidi derivati asessuati)
NOTA: Coltivare V. dahliae (ceppo JR2) allo stesso modo di V. longisporum (ceppo Vl43)17,18,19. Assicurarsi che tutte le apparecchiature e i supporti siano privi di germi e che tutti i passaggi vengano eseguiti in una cappa a flusso laminare per mantenere l'inoculo axenico.
- Riempire 150 mL di PDB liquido (fase 1.1) in un matraccio da 500 mL e integrare il mezzo con 500 mg/L di cefotaxime.
- Aggiungere Verticillium conida dalla conservazione del glicerolo al supporto PDB. Chiudere il pallone con un tappo di schiuma sterile.
- Incubare la coltura per 7-10 giorni in una scatola buia a temperatura ambiente (RT) sotto agitazione orizzontale continua (agitatore rotante; 60 giri / min). Ciò si traduce in piccole sfere micelie bianche.
- Rimuovere ed eliminare il surnatante PDB con attenzione. La maggior parte dei miceli dovrebbe rimanere nel pallone.
- Aggiungere 100 mL di CDB liquido (fase 1.2) sul micelio nel matraccio e integrare il mezzo con 500 mg/L di cefotaxima.
- Incubare altri 4-5 giorni in una scatola scura a RT sotto scuotimento continuo e orizzontale (agitatore rotativo, 60 giri / min) per indurre la sporulazione. Il surnatante diventerà giallastro-grigiastro quando i conidi vengono rilasciati.
- Filtrare una porzione (5-10 mL) del liquido contenente conidi attraverso una carta da filtro (livello di ritenzione delle particelle di 8-12 μm) in un tubo di raccolta sterile da 50 mL. Questo separa le spore dai miceli.
- Determinare la concentrazione di spore utilizzando una camera di conteggio delle cellule e un microscopio. Diluire con mezzo 1/4 MS privo di germi in acqua ultrapura fino ad ottenere le concentrazioni di spore indicate di seguito.
NOTA: Al microscopio, i conidi di V. longisporum sono per lo più disegnati a lungo e di dimensioni 7,1-8,8 μm, mentre i conidi V. dahliae sono più corti (3,5-5,5 μm) e piuttosto sferici20. - Usa questi conidi appena raccolti come inoculo. Assicurarsi di condurre gli esperimenti sempre con conidi appena raccolti e non con scorte congelate, poiché il congelamento riduce significativamente il numero di spore vitali19.
- Per la conservazione a lungo termine, congelare le spore come soluzione di spore ad alta concentrazione (circa 1 x 108 spore/ml) in glicerolo al 25% a -80 °C (conservabile fino a 1 anno). Per i prossimi esperimenti, utilizzare questi stock di glicerolo per inoculare il mezzo PDB nel passaggio 3.2.
4. Un sistema di inoculazione sterile in vitro a base di piastre di Petri
NOTA: Per il sistema a capsula di Petri17, assicurarsi che tutte le apparecchiature e i supporti siano privi di germi e che tutti i passaggi vengano eseguiti in una cappa a flusso laminare.
- Dopo l'autoclave, versare il mezzo (vedere il punto 1.3) nelle piastre di Petri.
- Dopo l'indurimento del mezzo, reimballare le piastre di Petri in un sacchetto di plastica sterile e conservarle capovolta per una notte in frigorifero (4-10 °C). Un mezzo refrigerato aiuta a prevenire lo scorrimento del mezzo nei passaggi successivi.
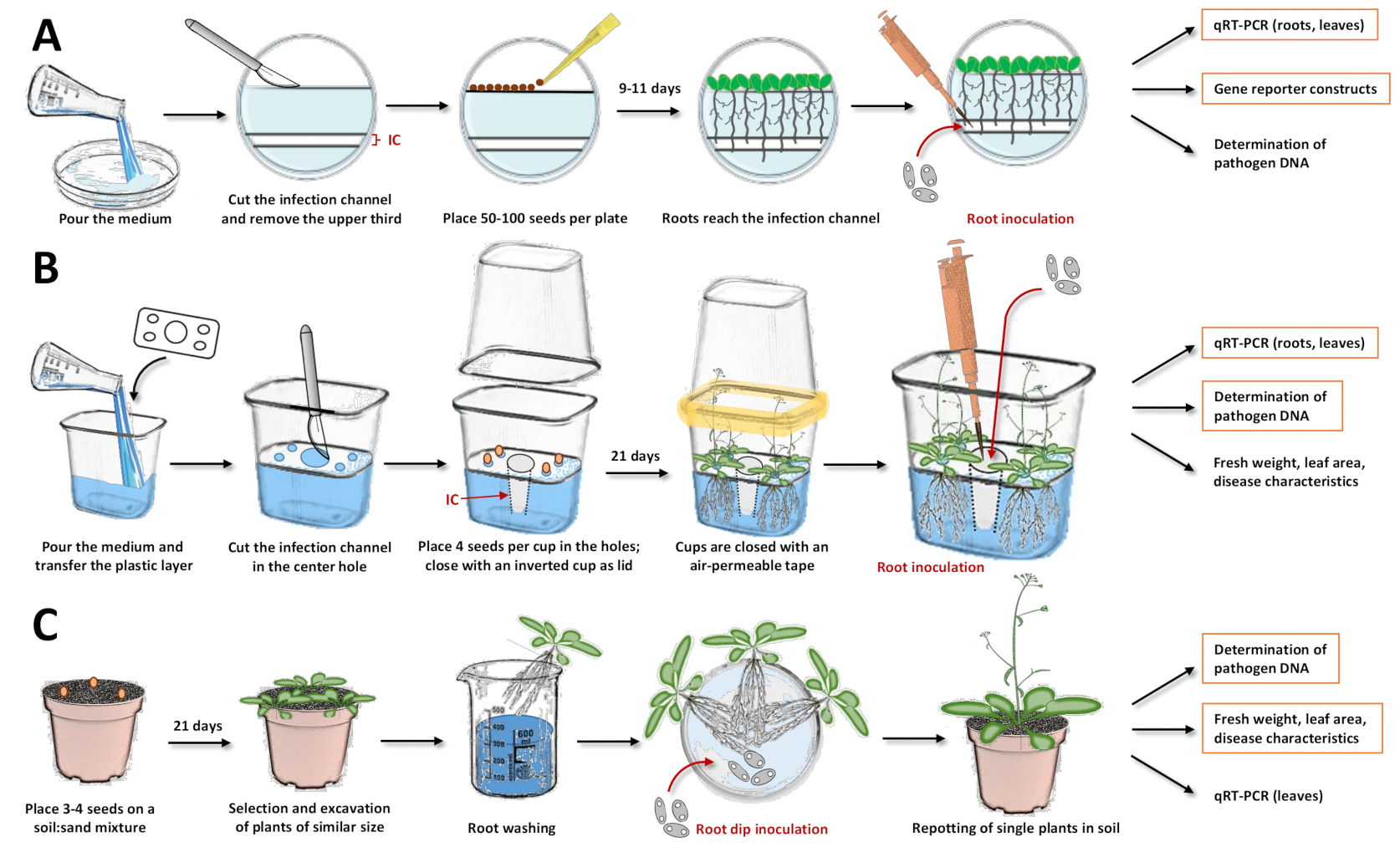
- Tagliare e rimuovere un canale di infezione e il terzo superiore del mezzo solidificato con un bisturi (Figura 1A). Evitare di ottenere liquido o aria sotto il mezzo agar durante il taglio; altrimenti, il mezzo scivolerà e chiuderà il canale di infezione.
- Distribuire 50-100 semi di Arabidopsis sterilizzati in superficie con una punta di pipetta sterile sulla superficie superiore tagliata. Metti i semi nell'angolo in cui la superficie dell'agar tagliata entra in contatto con la parete della capsula di Petri in modo che le radici possano crescere tra il mezzo e la parete della capsula di Petri. Ciò faciliterà l'inoculazione in seguito.
- Chiudere le piastre di Petri e sigillarle con nastro adesivo permeabile all'aria per consentire lo scambio di gas.
- Dopo la stratificazione per 2 giorni nell'oscurità a 4 °C, posizionare le piastre verticalmente in un rack adatto e far crescere le piante a 22 °C ± 1 °C in condizioni di lunga giornata (16 h luce / 8 h buio) in una camera di crescita.
- Quando la maggior parte delle radici raggiunge il canale di infezione (piantine di circa 9-11 giorni), adagiare le piastre orizzontalmente, aprirle e aggiungere 500 μL di Verticillium conidia appena raccolto con una concentrazione di 4 x 105 spore/mL direttamente nel canale di infezione, assicurandosi che il liquido sia distribuito uniformemente nel canale.
- Allo stesso modo, preparare piastre di controllo aggiungendo 500 μL di una soluzione simulata invece di spore (mezzo 1/4 MS privo di germi).
- Incubare le piastre orizzontalmente per un paio di minuti fino a quando il liquido non si è immerso e non può fuoriuscire quando le piastre vengono nuovamente posizionate verticalmente. Quindi, chiudere il coperchio e sigillare le piastre con nastro adesivo permeabile all'aria.
- Incubare le piastre verticalmente nella camera di crescita. Facoltativamente, coprire le parti della radice con scatole di carta nera per scurire le radici e i funghi trasmessi dal suolo (vedi19).
- Eseguire le analisi nei punti temporali preferiti dopo l'inoculazione a seconda della domanda di ricerca (fare riferimento alle legende delle figure per i punti temporali esatti utilizzati qui). Di seguito sono riportati alcuni suggerimenti.
- Tagliare le foglie dalle radici e raccoglierle entrambe separatamente. Togliere le strisce di agar dalle piastre di Petri per accedere facilmente alle radici e tirarle fuori con cura dall'agar usando una pinza. Congelare immediatamente tutto il materiale vegetale in azoto liquido.
- Macinare i campioni in azoto liquido. Estrarre il DNA totale da 100 mg di materiale fogliare per determinare tramite una PCR quantitativa (qPCR) la quantità di DNA fungino rispetto al DNA vegetale (vedere19).
- Macinare i campioni in azoto liquido. Prendi 100 mg di materiale vegetale ed estrai l'RNA totale. Condurre la PCR quantitativa a trascrizione inversa (qRT-PCR) per determinare l'espressione dei geni delle piante (o dei geni fungini) durante l'infestazione (vedere19).
- Rimuovere con cura le radici dall'agar evitando lesioni ed esaminarle al microscopio fluorescente.
- Determinare l'induzione di geni marcatori in linee reporter vegetali (ad esempio, luciferasi, β-glucuronidasi o reporter fluorescenti 17,19,21).
- Visualizza la propagazione fungina alla radice utilizzando linee reporter fungine (ad esempio, V. longisporum che esprime costitutivamente la proteina fluorescente verde potenziata, Vl-sGFP9) o mediante tecniche di colorazione (ad esempio, attraverso 5-bromo-4-cloro-3-indossil-N-acetil-beta-d-glucosaminide (X-beta-D-Glc-Nac)18).
- Tagliare le foglie dalle radici e raccoglierle entrambe separatamente. Togliere le strisce di agar dalle piastre di Petri per accedere facilmente alle radici e tirarle fuori con cura dall'agar usando una pinza. Congelare immediatamente tutto il materiale vegetale in azoto liquido.
5. Un sistema di inoculazione sterile in vitro organizzato con bicchieri di plastica
NOTA: Come indicato nella prima descrizione di questa tecnica19, assicurarsi che tutte le apparecchiature e i fluidi siano privi di germi e che tutti i passaggi vengano eseguiti in una cappa a flusso laminare.
- Utilizzare bicchieri di plastica trasparente con un volume totale di 500 ml e sterilizzarli in un bagno di etanolo al 70% -75% per almeno 20 minuti. Asciugare le tazze nella cappa a flusso laminare.
- Versare il mezzo autoclavato (vedere punto 1.4) nei bicchieri di plastica. Facoltativamente, aggiungere cefotaxime (concentrazione finale di 50 mg/L) al mezzo autoclavato per evitare contaminazioni batteriche. Utilizzare 150 ml di mezzo per tazza per esperimenti con Arabidopsis o più medio (250-300 ml per tazza) per esperimenti con specie vegetali più grandi (colza, pomodoro).
- Posizionare uno strato di plastica (sterilizzato prima incubando in etanolo al 70%-75% per 20 minuti) sul mezzo prima che si solidifichi (Figura 1B).
NOTA: questo strato di plastica contiene quattro fori prefabbricati agli angoli per posizionare semi sterilizzati in superficie. Ciò consente ai semi di accedere al mezzo. Successivamente, questo strato separatore impedisce alle foglie di toccare il mezzo contenente funghi, in modo che i microbi non possano attaccare direttamente le foglie e debbano prendere il percorso radicale. Un altro foro è al centro, consentendo di tagliare il canale di infezione. - Quando il mezzo si è solidificato, tagliare l'agar con un bisturi attraverso il foro centrale prefabbricato ad una profondità di circa 1,5 cm. Rimuovere l'agar tagliato per creare un canale di infezione in cui le spore fungine possono essere aggiunte in seguito.
- Grattare leggermente il mezzo agar con una punta di pipetta nei quattro fori più piccoli per interrompere la pelle solidificata (questo consente ai semi di assorbire l'acqua dal mezzo acquoso di agar). Posizionare i semi usando una punta di pipetta nei fori più piccoli.
- Chiudere il bicchiere di plastica con un secondo bicchiere di plastica invertito e sigillare con nastro adesivo permeabile all'aria. Il nastro deve consentire lo scambio di gas.
- Dopo la stratificazione per 3 giorni al buio a 4 °C, incubare i sistemi di tazze sotto 12 ore di luce / 12 ore di oscurità (Arabidopsis, colza) o 16 ore di luce / 8 ore di oscurità (pomodoro) in camere di crescita a una temperatura costante di 22 °C e 60% di umidità.
- Seguire l'età raccomandata delle piante per l'inoculazione: 21 giorni per Arabidopsis; 5-7 giorni per la colza; 12 giorni per il pomodoro.
- Inoculare le piantine con Verticillium aggiungendo 1 mL di soluzione di conidi (concentrazione raccomandata: 4 x 105 spore/mL) nel canale di infezione. Per preparare i campioni di controllo, aggiungere 1 mL di soluzione simulata senza spore (mezzo 1/4 MS privo di germi) nel canale.
- Eseguire le analisi nei punti temporali preferiti dopo l'inoculazione a seconda della domanda di ricerca (fare riferimento alle legende delle figure per i punti temporali esatti utilizzati qui). Di seguito sono riportati alcuni suggerimenti.
- Scatta fotografie delle piante con una fotocamera digitale dall'alto mantenendo la stessa distanza per ogni foto. Quantificare l'area fogliare (ad esempio, con ImageJ22 o BlattFlaeche17,19; utilizzare la lunghezza delle tazze per impostare la scala) e confrontare i gruppi infetti e di controllo. Classificare lo sviluppo dei sintomi della malattia (ad esempio, foglie più piccole, più giallastre o necrotiche).
NOTA: Se ci sono steli su Arabidopsis, rimuoverli per ottenere foto migliori delle rosette. - Rimuovere le radici e definire la biomassa (peso fresco) dei germogli da campioni infetti e controllare i campioni mediante pesatura. Determinare il peso fresco relativo19.
- Raccogliere i campioni per le analisi molecolari come segue.
- Arabidopsis: Rimuovere gli steli se ce ne sono. Tagliare le rosette alla base dalle radici. Assicurati di escludere tutto il materiale radicale dal campione e raccogli rosette intere. Combinare 4-5 rosette di piante diverse in un unico campione e congelare il materiale fogliare in azoto liquido.
- Estrarre accuratamente le radici dal mezzo con una pinza, premere e tamponarle con un tovagliolo di carta per rimuovere i resti di agar e combinare 4-5 radici di piante diverse in un unico campione. Congelare immediatamente in azoto liquido.
- Colza / pomodoro: tagliare i segmenti del gambo dall'ipocotile (ad esempio, 1 cm di lunghezza; prendere sempre la stessa regione del gambo). Combinare il materiale di 4-5 piante in ogni campione e congelare in azoto liquido.
- Macinare i campioni in azoto liquido. Estrarre il DNA totale da 100 mg del materiale fogliare o dello stelo per determinare tramite qPCR la quantità di DNA fungino rispetto al DNA vegetale (vedere19).
- Macinare i campioni in azoto liquido, prelevare 100 mg di materiale vegetale ed estrarre l'RNA totale. Condurre la qRT-PCR per determinare l'espressione dei geni delle piante (o dei geni fungini) al momento dell'infestazione (vedere19).
- Scatta fotografie delle piante con una fotocamera digitale dall'alto mantenendo la stessa distanza per ogni foto. Quantificare l'area fogliare (ad esempio, con ImageJ22 o BlattFlaeche17,19; utilizzare la lunghezza delle tazze per impostare la scala) e confrontare i gruppi infetti e di controllo. Classificare lo sviluppo dei sintomi della malattia (ad esempio, foglie più piccole, più giallastre o necrotiche).
6. Un sistema di inoculazione a base di suolo in vaso
- Mescolare accuratamente il terreno e la sabbia in un rapporto volumetrico 3: 1 (terra: sabbia) per facilitare il lavaggio del substrato dalle radici. Versare il composto in un sacchetto di autoclave. Se la miscela è troppo secca, aggiungere una quantità appropriata di acqua e mescolarla nel substrato. Vapore a 80 °C per 20 minuti in autoclave per ridurre al minimo le contaminazioni microbiche.
NOTA: Evitare il riscaldamento a oltre 80 °C, in quanto ciò potrebbe influire sui nutrienti organici del suolo. - Riempire i vasi con la miscela di terra e sabbia e trasferirli in vassoi. Aggiungere acqua nei vassoi a circa 1/3 dell'altezza di una pentola, in modo che la miscela di terra e sabbia venga completamente imbevuta d'acqua. Inoltre, spruzzare il substrato con un flacone spray per garantire condizioni di partenza bagnate.
- Semina 3-4 semi in ogni vaso (Figura 1C) assicurandoti che i semi abbiano una distanza sufficiente l'uno dall'altro. Tenerli per 3 giorni al buio a 4 °C per la stratificazione per sincronizzare la germinazione.
NOTA: Pre-coltivare un eccesso di piante, che consente una selezione di piante di dimensioni simili per gli esperimenti di inoculazione e riduce le deviazioni dovute alle differenze individuali. - Lasciare che le piantine crescano in condizioni di lunga giornata (16 h di luce / 8 h di buio; temperatura costante di 22 °C; 60% di umidità) con annaffiature regolari.
- Seguire l'età raccomandata delle piante per l'inoculazione: 21 giorni per Arabidopsis, 7 giorni per la colza e 10 giorni per il pomodoro. Scegli piante di dimensioni simili per eseguire l'"inoculazione a immersione radicale"15,17,23,24. Prendi il terreno dai vasi e scava con cura le radici.
- Lavare delicatamente solo le radici in un contenitore d'acqua e tenere le rosette fuori dall'acqua. Incubare le radici lavate per 60 minuti in una capsula di Petri contenente la soluzione di spore verticillium (concentrazione consigliata: 2 x 106 spore/mL). Per il gruppo di controllo non infetto, incubare le radici per 60 minuti nella soluzione simulata senza spore (mezzo 1/4 MS privo di germi).
- Preparare nuovi vasi con terreno umido e sterilizzato a vapore (80 °C per 20 min) senza sabbia. Usa una punta della pipetta per fare un buco al centro del terreno in ogni pentola.
- Posizionare direttamente le radici nel foro (trasferire solo una pianta per vaso). Dopo aver inserito le radici, assicurarsi di riempire accuratamente i fori con il terreno. Evitare di premere il terreno, altrimenti il rinvaso può causare sintomi di stress come foglie viola.
- Coltivare gruppi infetti e di controllo in condizioni di lunga giornata (16 ore di luce / 8 ore di buio; una temperatura costante di 22 °C; 60% di umidità) con irrigazione regolare.
- Eseguire le analisi nei punti temporali preferiti dopo l'inoculazione a seconda della domanda di ricerca (fare riferimento alle legende delle figure per i punti temporali esatti utilizzati qui). Di seguito sono riportati alcuni suggerimenti.
- Scatta fotografie delle piante con una fotocamera digitale dall'alto, mantenendo la stessa distanza per ogni foto. Quantificare l'area fogliare (ad esempio, con ImageJ22 o BlattFlaeche17,19; utilizzare il diametro dei vasi per impostare la scala) e confrontare i gruppi infetti e di controllo. Classificare lo sviluppo dei sintomi della malattia (ad esempio, foglie più piccole, più giallastre o necrotiche)13.
NOTA: La rimozione degli steli di Arabidopsis facilita lo scatto di foto delle rosette. - Rimuovere le radici e definire la biomassa (peso fresco) dei germogli da campioni infetti e controllare i campioni mediante pesatura. Determinare il peso fresco relativo19.
- In alternativa, misurare l'altezza della pianta o classificare la crescita fungina dai segmenti staminali per valutare la gravità della malattia13.
- Raccogliere campioni per analisi molecolari come segue.
- Arabidopsis: rimuovere gli steli. Tagliare le rosette alla corona della radice. Combina 4-5 rosette di piante diverse in un unico campione. Congelare il materiale fogliare in azoto liquido.
NOTA: Nel caso delle radici, è difficile pulirle sufficientemente dal terreno senza riprogrammare l'espressione genica attraverso il lavaggio. - Colza / pomodoro: tagliare i segmenti del gambo dall'ipocotile (ad esempio, 1 cm di lunghezza; prendere sempre la stessa regione del gambo). Combina il materiale di 4-5 piante in un campione e congelalo in azoto liquido.
- Macinare i campioni in azoto liquido. Estrarre il DNA totale da 100 mg di materiale fogliare o stelo per determinare tramite qPCR la quantità di DNA fungino rispetto al DNA vegetale (vedere19).
- Macinare i campioni in azoto liquido, prelevare 100 mg di materiale vegetale ed estrarre l'RNA totale. Condurre la qRT-PCR per determinare l'espressione dei geni delle piante (o dei geni fungini) al momento dell'infestazione (vedere19).
- Scatta fotografie delle piante con una fotocamera digitale dall'alto, mantenendo la stessa distanza per ogni foto. Quantificare l'area fogliare (ad esempio, con ImageJ22 o BlattFlaeche17,19; utilizzare il diametro dei vasi per impostare la scala) e confrontare i gruppi infetti e di controllo. Classificare lo sviluppo dei sintomi della malattia (ad esempio, foglie più piccole, più giallastre o necrotiche)13.
7. Analisi dei dati
- Calcola la deviazione media e standard (± SD) in base alle repliche biologiche.
- Calcolare i valori relativi dividendo tutti i risultati del gruppo infetto per il risultato del controllo. Visualizzare la media come, ad esempio, "relativa al finto" o "relativa al wild-type".
- Determinare la significatività statistica tra i gruppi.

Figura 1: Compilazione dei tre sistemi di inoculazione e dei singoli passaggi nei protocolli. Queste figure illustrano i sistemi con l'impianto modello Arabidopsis thaliana. Per altre specie vegetali, i tempi devono essere regolati. Evidenziano le caselle arancioni, per le quali le analisi successive sono più raccomandate con il rispettivo sistema. (A) Per il sistema di inoculazione nelle piastre di Petri17, versare il mezzo e lasciarlo solidificare. Conservare i piatti in frigorifero per una notte. Quindi, tagliare e rimuovere il terzo superiore e il canale di infezione (IC) con un bisturi (le aree bianche nell'illustrazione sono state rimosse dall'agar, mentre le aree bluastre rappresentano l'agar). Posizionare i semi sulla superficie tagliata e chiudere le piastre di Petri. Dopo la stratificazione, posizionare le piastre verticalmente e lasciare che le piante crescano. Una volta che la maggior parte delle radici ha raggiunto il canale di infezione, aggiungere la soluzione di spore con una pipetta direttamente nel canale. Assicurarsi che la soluzione sia distribuita uniformemente. Chiudere le piastre di Petri e incubarle verticalmente in una camera di crescita. Gli approcci che possono seguire sono l'analisi espressionale con PCR quantitativa a trascrizione inversa (qRT-PCR), la microscopia con linee reporter e la quantificazione del DNA microbico. (B) Per il sistema di inoculazione in bicchieri di plastica19, versare il mezzo e trasferire lo strato di plastica separatore con i fori prefabbricati (quattro piccoli fori negli angoli per posizionare i semi e un grande foro al centro per il canale di infezione). Lascia che il mezzo si solidifichi. Tagliare e rimuovere il mezzo agar nel foro centrale con un bisturi per ottenere il canale di infezione (IC). Gratta il mezzo nei fori più piccoli e trasferisci i semi. Chiudere la tazza con una tazza rovesciata e sigillare con nastro impermeabile all'aria (simboleggiato in giallo). Lascia che le piante crescano. Per l'inoculazione, aggiungere la soluzione di spore con una pipetta direttamente nel canale di infezione. Chiudere il sistema e continuare la coltivazione nella camera di crescita. Gli approcci che possono seguire sono l'analisi espressionale con qRT-PCR, la quantificazione del DNA microbico e la determinazione del peso fresco, dell'area fogliare o di altre caratteristiche della malattia. (C) "Inoculazione a immersione radicale"15,17,23,24: per il sistema di inoculazione a base di suolo, riempire i vasi con una miscela di terra:sabbia. Trasferisci i semi e lascia crescere le piantine. Scava piante di dimensioni simili e lava le radici in acqua. Posizionare le radici lavate in una capsula di Petri tenendo la soluzione con le spore. Dopo l'incubazione, inserire singole piante in vasi con terra. Gli approcci che possono seguire sono l'analisi espressionale nelle foglie con qRT-PCR, la quantificazione del DNA microbico e la determinazione del peso fresco, dell'area fogliare o di altre caratteristiche della malattia. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Risultati
Seguendo il protocollo, le piante sono state coltivate e inoculate con V. longisporum (ceppo Vl4325) o V. dahliae (isolato JR218). Vari scenari sono stati progettati per dimostrare l'efficacia e per evidenziare alcune capacità dei protocolli dati. Vengono mostrati i risultati rappresentativi.
L'induzione espressionale di geni coinvolti nella biosintesi antimicrobica indol-glucosinolato (IG) è un indicatore affidabile ...
Discussione
A causa delle enormi perdite di resa causate dai fitopatogeni trasmessi dal suolo1, è necessario un miglioramento delle strategie agricole o delle varietà di colture. La limitata comprensione della patogenesi delle malattie trasmesse dal suolo ostacola lo sviluppo di piante più resistenti. È necessario esplorare i meccanismi patologici sottostanti, per i quali è necessaria una solida piattaforma metodologica. Le procedure di inoculazione riportate hanno dimostrato che gli eventi multifattoria...
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Gli autori riconoscono Tim Iven e Jaqueline Komorek per il lavoro precedente su questi metodi, il gruppo di Wolfgang Dröge-Laser (Dipartimento di Biologia Farmaceutica, Università di Würzburg, Germania) per aver fornito le attrezzature e le risorse necessarie per questo lavoro, e Wolfgang Dröge-Laser e Philipp Kreisz (entrambi Università di Würzburg) per la correzione critica del manoscritto. Questo studio è stato sostenuto dalla "Deutsche Forschungsgemeinschaft" (DFG, DR273/15-1,2).
Materiali
| Name | Company | Catalog Number | Comments |
| Agar (Gelrite) | Carl Roth | Nr. 0039 | all systems described require Gelrite |
| Arabidopsis thaliana wild-type | NASC stock | Col-0 (N1092) | |
| Autoclave | Systec | VE-100 | |
| BlattFlaeche | Datinf GmbH | BlattFlaeche | software to determine leaf areas |
| Brassica napus wild-type | see Floerl et al., 2008 | rapid-cycling rape | genome ACaacc |
| Cefotaxime sodium | Duchefa | C0111 | |
| Chicanery flask 500 mL | Duran Group / neoLab | E-1090 | Erlenmeyer flask with four baffles |
| Collection tubes 50 mL | Sarstedt | 62.547.254 | 114 x 28 mm |
| Czapek Dextrose Broth medium | Duchefa | C1714 | |
| Digital camera | Nikon | D3100 18-55 VR | |
| Exsiccator (Desiccator ) | Duran Group | 200 DN, 5.8 L | Seal with lid to hold chlorine gas |
| Fluorescence Microscope | Leica | Leica TCS SP5 II | |
| HCl | Carl Roth | P074.3 | |
| KNO3 | Carl Roth | P021.1 | ≥ 99 % |
| KOH | Carl Roth | 6751 | |
| Leukopor | BSN medical GmbH | 2454-00 AP | non-woven tape 2.5 cm x 9.2 m |
| MES (2-(N-morpholino)ethanesulfonic acid) | Carl Roth | 4256.2 | Pufferan ≥ 99 % |
| MgSO4 | Carl Roth | T888.1 | Magnesiumsulfate-Heptahydrate |
| Murashige & Skoog medium (MS) | Duchefa | M0222 | MS including vitamins |
| NaClO | Carl Roth | 9062.1 | |
| Percival growth chambers | CLF Plant Climatics GmbH | AR-66L2 | |
| Petri-dishes | Sarstedt | 82.1473.001 | size ØxH: 92 × 16 mm |
| Plastic cups (500 mL, transparent) | Pro-pac, salad boxx | 5070 | size: 108 × 81 × 102 mm |
| Pleated cellulose filter | Hartenstein | FF12 | particle retention level 8–12 μm |
| poly klima growth chamber | poly klima GmbH | PK 520 WLED | |
| Potato Dextrose Broth medium | SIGMA Aldrich | P6685 | for microbiology |
| Pots | Pöppelmann GmbH | TO 7 D or TO 9,5 D | Ø 7 cm resp. Ø 9.5 cm |
| PromMYB51::YFP | see Poncini et al., 2017 | MYB51 reporter line | YFP (i.e. 3xmVenus with NLS) |
| Reaction tubes 2 mL | Sarstedt | 72.695.400 | PCR Performance tested |
| Rotary (orbital) shaker | Edmund Bühler | SM 30 C control | |
| Sand (bird sand) | Pet Bistro, Müller Holding | 786157 | |
| Soil | Einheitserde spezial | SP Pikier (SP ED 63 P) | |
| Solanum lycopersicum wild-type | see Chavarro-Carrero et al., 2021 | Type: Moneymaker | |
| Thoma cell counting chamber | Marienfeld | 642710 | depth 0.020 mm; 0.0025 mm2 |
| Ultrapure water (Milli-Q purified water) | MERK | IQ 7003/7005 | water obtained after purification |
| Verticillium dahliae | see Reusche et al., 2014 | isolate JR2 | |
| Verticillium longisporum | Zeise and von Tiedemann, 2002 | strain Vl43 |
Riferimenti
- Mendes, R., Garbeva, P., Raaijmakers, J. M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiology Review. 37 (5), 634-663 (2013).
- Yadeta, K. A., Thomma, B. P. H. J. The xylem as battleground for plant hosts and vascular wilt pathogens. Frontiers in Plant Science. 4, 97 (2013).
- Delgado-Baquerizo, M., et al. The proportion of soil-borne pathogens increases with warming at the global scale. Nature Climate Change. 10 (6), 550-554 (2020).
- Berendsen, R. L., et al. Disease-induced assemblage of a plant-beneficial bacterial consortium. The ISME Journal. 12 (6), 1496-1507 (2018).
- Yuan, J., et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome. 6 (1), 156 (2018).
- Liu, H., et al. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytologist. 229 (5), 2873-2885 (2021).
- Wang, H., Liu, R., You, M. P., Barbetti, M. J., Chen, Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): role of bacterial diversity. Microorganisms. 9 (9), 1988 (2021).
- Inderbitzin, P., Subbarao, K. V. Verticillium systematics and evolution: how confusion impedes Verticillium wilt management and how to resolve it. Phytopathology. 104 (6), 564-574 (2014).
- Eynck, C., Koopmann, B., Grunewaldt-Stoecker, G., Karlowsky, P., von Tiedemann, A. Differential interactions of Verticillium longisporum und V. dahliae with Brassica napus with molecular and histological techniques. European Journal of Plant Pathology. 118 (3), 259-274 (2007).
- Floerl, S., et al. Defence reactions in the apoplastic proteome of oilseed rape (Brassica napus var. napus) attenuate Verticillium longisporum growth but not disease symptoms. BMC Plant Biology. 8, 129 (2008).
- Leonard, M., et al. Verticillium longisporum elicits media-dependent secretome responses with capacity to distinguish between plant-related environments. Frontiers in Microbiology. 11, 1876 (2020).
- Depotter, J. R. L., et al. Verticillium longisporum, the invisible threat to oilseed rape and other brassicaceous plant hosts. Molecular Plant Pathology. 17 (7), 1004-1016 (2016).
- Fröschel, C., et al. A gain-of-function screen reveals redundant ERF transcription factors providing opportunities for resistance breeding toward the vascular fungal pathogen Verticillium longisporum. Molecular Plant-Microbe Interactions. 32 (9), 1095-1109 (2019).
- Zhou, L., Hu, Q., Johansson, A., Dixelius, C. Verticillium longisporum and V. dahliae: infection and disease in Brassica napus. Plant Pathology. 55 (1), 137-144 (2006).
- Ralhan, A., et al. The vascular pathogen Verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots. Plant Physiology. 159 (3), 1192-1203 (2012).
- Reusche, M., et al. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium longisporum. Molecular Plant-Microbe Interactions. 26 (8), 850-860 (2013).
- Iven, T., et al. Transcriptional activation and production of tryptophan-derived secondary metabolites in Arabidopsis roots contributes to the defense against the fungal vascular pathogen Verticillium longisporum. Molecular Plant. 5 (6), 1389-1402 (2012).
- Reusche, M., et al. Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana. Environmental and Experimental Botany. 108, 23-37 (2014).
- Fröschel, C. In-depth evaluation of root infection systems using the vascular fungus Verticillium longisporum as soil-borne model pathogen. Plant Methods. 17 (1), 57 (2021).
- Karapapa, V. K., Bainbridge, B. W., Heale, J. B. Morphological and molecular characterization of Verticillium longisporum comb, nov., pathogenic to oilseed rape. Mycological Research. 101 (11), 1281-1294 (1997).
- Poncini, L., et al. In roots of Arabidopsis thaliana, the damage-associated molecular pattern AtPep1 is a stronger elicitor of immune signalling than flg22 or the chitin heptamer. PLoS One. 12 (10), 1-21 (2017).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Fradin, E. F., et al. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiology. 150 (1), 320-332 (2009).
- Singh, S., et al. The plant host Brassica napus induces in the pathogen Verticillium longisporum the expression of functional catalase peroxidase which is required for the late phase of disease. Molecular Plant-Microbe Interactions. 25 (4), 569-581 (2012).
- Zeise, K., von Tiedemann, A. Application of RAPD-PCR for virulence type analysis within Verticillium dahliae and Verticillium longisporum. Journal of Phytopathology. 150 (10), 557-563 (2002).
- Fröschel, C., et al. Plant roots employ cell-layer-specific programs to respond to pathogenic and beneficial microbes. Cell Host & Microbe. 29 (2), 299-310 (2021).
- Gigolashvili, T., et al. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. The Plant Journal. 50 (5), 886-901 (2007).
- Back, M. A., Haydock, P. P. J., Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathology. 51 (6), 683-697 (2002).
- Behrens, F. H., et al. Suppression of abscisic acid biosynthesis at the early infection stage of Verticillium longisporum in oilseed rape (Brassica napus). Molecular Plant Pathology. 20 (12), 1645-1661 (2019).
- Vorholt, J. A., Vogel, C., Carlström, C. I., Müller, D. B. Establishing causality: opportunities of synthetic communities for plant microbiome research. Cell Host & Microbe. 22 (2), 142-155 (2017).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati