
JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
식물 뿌리를 토양 매개 미생물로 감염시키는 접종 전략
요약
이 프로토콜은 식물 뿌리에 토양 매개 미생물을 접종하는 전략에 대한 자세한 요약을 제공합니다. 진균 Verticillium longisporum 및 Verticillium dahliae에 대해 예시된 세 가지 상이한 뿌리 감염 시스템이 기술된다. 잠재적 응용 분야와 가능한 다운스트림 분석이 강조되고 각 시스템에 대한 장단점이 논의됩니다.
초록
근권은 식물 뿌리가 끊임없이 도전받는 매우 복잡한 미생물 공동체를 가지고 있습니다. 뿌리는 다양한 미생물과 밀접한 접촉을하지만, 토양 매개 상호 작용에 대한 연구는 여전히 지상 기관에서 수행 된 연구 뒤에 있습니다. 모델 뿌리 병원균으로 모델 식물을 감염시키기 위한 일부 접종 전략이 문헌에 기술되어 있지만, 포괄적인 방법론적 개요를 얻는 것은 여전히 어렵다. 이 문제를 해결하기 위해, 뿌리-미생물 상호작용의 생물학에 대한 통찰을 얻기 위해 적용될 수 있는 세 가지 상이한 뿌리 접종 시스템이 정확하게 기술된다. 예를 들어, Verticillium 종 (즉, V. longisporum 및 V. dahliae)은 뿌리 침입 모델 병원균으로 사용되었습니다. 그러나, 상기 방법은 병원성 및 유익한 미생물 둘 다의 다른 뿌리 식민지화에 쉽게 적응될 수 있다. 식물 자일렘을 식민지화함으로써, Verticillium spp.와 같은 혈관 토양 매개 진균은 독특한 생활 방식을 나타낸다. 뿌리 침범 후, 그들은 자일렘 혈관을 통해 작물 적으로 퍼지고, 싹에 도달하고, 질병 증상을 유발합니다. 세 가지 대표적인 식물 종들이 모델 숙주로 선택되었다: 애기장대 탈리아나, 경제적으로 중요한 오일시드 유채(브라시카 나푸스), 토마토(솔라눔 리코페르시쿰). 단계별 프로토콜이 제공됩니다. 병원성 분석, 마커 유전자의 전사 분석 및 리포터 구축물에 의한 독립적 확인의 대표적인 결과가 제시된다. 또한, 각 접종 시스템의 장점과 단점이 철저히 논의됩니다. 이러한 입증 된 프로토콜은 뿌리 - 미생물 상호 작용에 대한 연구 질문에 대한 접근 방식을 제공하는 데 도움이 될 수 있습니다. 식물이 토양의 미생물에 어떻게 대처하는지 아는 것은 농업을 개선하기위한 새로운 전략을 개발하는 데 중요합니다.
서문
자연 토양에는 식물에 중성, 유해 또는 유익 할 수있는 놀라운 수의 미생물이 서식합니다1. 많은 식물 병원균은 토양에서 태어나 뿌리를 둘러싸고 지하 기관을 공격합니다. 이 미생물은 곰팡이, oomycetes, 박테리아, 선충류, 곤충 및 일부 바이러스 1,2와 같은 다양한 클래드에 속합니다. 환경 조건이 감염을 선호하면 감염되기 쉬운 식물이 병에 걸리고 작물 수확량이 감소합니다. 지구 온난화와 극심한 날씨와 같은 기후 변화의 영향은 토양 매개 식물 병원균의 비율을 증가시킬 것입니다3. 따라서 이러한 파괴적인 미생물과 그 미생물이 식량 및 사료 생산뿐만 아니라 자연 생태계에 미치는 영향을 연구하는 것이 점점 더 중요해질 것입니다. 또한 토양에는 뿌리와 밀접하게 상호 작용하고 식물의 성장, 발달 및 면역을 촉진하는 미생물 상호 주의자가 있습니다. 병원균에 직면했을 때, 식물은 병원 균 4,5,6,7을 억제함으로써 숙주 생존을 지원할 수있는 근권의 특정 상대를 적극적으로 모집 할 수 있습니다. 그러나 유익한 뿌리 - 미생물 상호 작용과 관련된 기계론적 세부 사항 및 경로는 종종 여전히 알려지지 않았습니다6.
따라서 뿌리 - 미생물 상호 작용에 대한 일반적인 이해를 넓히는 것이 필수적입니다. 토양 매개 미생물에 뿌리를 접종하기위한 신뢰할 수있는 방법은 모델 연구를 수행하고 그 결과를 농업 응용 분야로 옮기는 데 필요합니다. 토양에서의 유익한 상호작용은 예를 들어, 세렌디피타 인디카 (이전의 피리포르모스포라 인디카로 알려짐), 질소-고정 리조븀 spp., 또는 균류 진균과 함께 연구되는 반면, 공지된 토양 매개 식물 병원체는 랄스토니아 솔라나세아럼, 피토프토호라 spp., 푸사리움 spp., 및 Verticillium spp.1을 포함한다. 후자의 두 가지는 전 세계적으로 분포되어 혈관 질환을 일으키는 곰팡이 속입니다2. Verticillium spp. (Ascomycota)는 초본 연간, 우디 다년생 식물 및 많은 작물 식물 2,8을 포함하여 주로 dicotyledons의 수백 종의 식물을 감염시킬 수 있습니다. Verticillium의 Hyphae는 뿌리에 들어가서 자일렘 혈관을 식민지화하기 위해 중앙 실린더를 향해 세포 간 및 세포 내에서 모두 성장합니다 2,9. 이 혈관에서 곰팡이는 수명주기의 대부분을 유지합니다. 자일렘 수액은 영양이 부족하고 식물 방어 화합물을 운반하기 때문에 곰팡이는이 독특한 환경에 적응해야합니다. 이는 병원체가 그의 숙주10,11에서 생존할 수 있게 하는 콜로니화 관련 단백질의 분비에 의해 달성된다. 뿌리 혈관 조직에 도달 한 후, 곰팡이는 자일렘 혈관 내에서 단풍으로 작물 적으로 퍼질 수 있으며, 이는 숙주 9,12의 전신 식민지화로 이어진다. 이 시점에서 식물은 성장 9,10,13에 부정적인 영향을받습니다. 예를 들어, 기절과 노란 잎은 조기 노화13,14,15,16뿐만 아니라 발생합니다.
이 속의 한 구성원은 Verticillium longisporum으로, 농경학적으로 중요한 유채 유채, 콜리 플라워 및 모델 식물 애기장대 탈리아나12와 같은 황동 숙주에 매우 적합합니다. 몇몇 연구는 V. longisporum과 A. thaliana를 결합하여 토양 매개 혈관 질환과 그 결과 뿌리 방어 반응13,15,16,17에 대한 광범위한 통찰력을 얻었습니다. 간단한 감수성 검사는 V. longisporum / A. thaliana 모델 시스템을 사용하여 실현 될 수 있으며 잘 확립 된 유전 자원은 두 유기체 모두에 사용할 수 있습니다. V. longisporum과 밀접한 관련이있는 병원체는 Verticillium dahliae입니다. 두 곰팡이 종 모두 비슷한 혈관 생활 양식과 침윤 과정을 수행하지만, 뿌리에서 잎으로의 전파 효율과 A. thaliana의 유발 된 질병 증상은 다릅니다 : V. longisporum은 일반적으로 조기 노화를 유도하는 반면, V. dahliae 감염은 시들음18을 초래합니다. 최근에, 방법론적 요약은 A. thaliana를 V. longisporum 또는 V. dahliae로 감염시키기 위한 상이한 뿌리 접종 전략을 제시하여, 실험 셋업19를 계획하는 데 도움을 주었다. 현장에서 V. longisporum은 때때로 유채 유채 생산12에서 심각한 손상을 일으키는 반면, V. dahliae는 포도 나무, 감자 및 토마토8과 같은 여러 재배 종으로 구성된 매우 광범위한 숙주 범위를 가지고 있습니다. 이것은 두 병원균을 경제적으로 흥미로운 모델로 연구하게합니다.
따라서, 다음의 프로토콜은 뿌리 접종을 위한 가능한 접근법을 예시하기 위해 모델 뿌리 병원체로서 V. longisporum 및 V. dahliae 둘 다를 사용한다. 애기장대 (애기장대 탈리아나), 유채 유채 (브라시카 나푸스) 및 토마토 (솔라눔 리코 페르시쿰)가 모델 호스트로 선택되었습니다. 방법론에 대한 자세한 설명은 아래 텍스트와 함께 제공되는 비디오에서 찾을 수 있습니다. 각 접종 시스템에 대한 장점과 단점이 논의됩니다. 종합하면,이 프로토콜 수집은 뿌리 - 미생물 상호 작용의 맥락에서 특정 연구 질문에 적합한 방법을 식별하는 데 도움이 될 수 있습니다.
Access restricted. Please log in or start a trial to view this content.
프로토콜
1. 곰팡이 배양 및 식물 접종 시스템을위한 미디어
- 액체 감자 덱스트로스 국물 (PDB) : 열 안정 플라스크에서 초순수로 21 g / L PDB를 준비하십시오.
- Liquid Czapek Dextrose Broth (CDB): 열 안정한 플라스크에서 초순수에 42 g/L CDB를 준비합니다.
- 페트리 접시 접종 시스템용 배지: 초순수에 1.5g/L Murashige 및 Skoog 배지(MS)와 8g/L 한천으로 열에 안정한 플라스크를 준비합니다.
참고 :이 배지의 설탕은 접종 후 과도한 곰팡이 성장을 유발할 수 있으므로이 배지에서 설탕을 피하십시오. - 플라스틱 컵 기반 접종 시스템용 배지: 초순수에서 4.4 g/L MS, 0.2 g/L MgSO4, 1 g/L KNO3, 0.5 g/L 2-(N-모르폴리노)에탄술폰산(MES) 및 6.0 g/L 한천으로 열 안정 플라스크를 준비하고 5 M KOH로 pH를 5.7로 조정하십시오.
참고 :이 배지의 설탕은 접종 후 과도한 곰팡이 성장을 유발할 수 있으므로이 배지에서 설탕을 피하십시오. - 1/4 MS 배지: 초순수에 1.2 g/L MS를 준비합니다.
- 오토클레이브를 사용하여 위의 모든 솔루션을 멸균하십시오. 유리 플라스크를 바구니에 넣고 뚜껑을 닫고 121 ° C 및 98.9 kPa에서 15 분 동안 살균하십시오.
2. 식물 씨앗의 표면 살균
참고 : 파종 전에 Arabidopsis, 유채 및 토마토에서 씨앗의 표면을 살균하기 위해 항상 아래 프로토콜을 사용하십시오.
- 종자를 2 mL 반응 튜브로 옮긴다. 튜브를 내부 용량이 5.8L인 엑시케이터에 넣으십시오.
- 12% 차아염소산나트륨(NaClO) 100mL에 33% 염산(HCl) 6mL를 첨가하여 엑시케이터에서 염소가스를 생성합니다.
- exsiccator의 뚜껑을 즉시 닫고 가스에서 3 시간 동안 씨앗을 배양하십시오.
3. Verticillium 포자(무성애 유래 코니디아)로 접종물 준비
참고: V. longisporum (균주 Vl43)17,18,19와 같은 방법으로 V. dahliae (균주 JR2)를 재배하십시오. 모든 장비와 매체에 세균이 없으며 모든 단계가 층류 후드에서 수행되어 접종 축을 유지하는지 확인하십시오.
- 500 mL 치카네리 플라스크에 액체 PDB 150 mL (단계 1.1)를 채우고 배지에 500 mg / L cefotaxime을 보충하십시오.
- 글리세롤 스톡 저장소로부터 코니다 버티실륨 을 PDB 배지에 첨가한다. 멸균 폼 마개로 플라스크를 닫으십시오.
- 배양물을 연속적이고 수평적인 진탕 (회전 진탕기; 60 rpm) 하에 실온 (RT)의 다크 박스에서 7-10 일 동안 인큐베이션한다. 이것은 작고 하얀 균사체 구체를 초래합니다.
- PDB 상청액을 조심스럽게 제거하고 버립니다. 균사체의 대부분은 플라스크에 남아 있어야합니다.
- chicanery 플라스크의 균사체에 100 mL의 액체 CDB (1.2 단계)를 넣고 배지에 500 mg / L cefotaxime을 보충하십시오.
- 또 다른 4-5일 동안 RT의 다크 박스에서 연속적이고 수평적인 진탕(rotary shaker, 60 rpm) 하에 인큐베이션하여 다공성을 유도하였다. 상청액은 코니디아가 방출되면 황갈색으로 변할 것입니다.
- 코니디아 함유 액체의 일부(5-10 mL)를 여과지(입자 보유 수준 8-12 μm)를 통해 멸균 50 mL 수집 튜브에 여과한다. 이것은 포자와 균사체를 분리합니다.
- 세포 계수 챔버 및 현미경을 이용하여 포자 농도를 결정한다. 아래에 주어진 포자 농도가 얻어질 때까지 무균 1/4 MS 배지를 초순수로 희석한다.
참고 : 현미경 하에서, V. longisporum 의 conidia는 대부분 긴 그려짐과 7.1-8.8 μm 크기이며, V. dahliae conidia는 더 짧고 (3.5-5.5 μm) 구형20입니다. - 이 갓 수확 한 conidia를 접종물로 사용하십시오. 동결은 생존 가능한 포자의 수를 크게 감소시키기 때문에 냉동 재고가 아닌 갓 수확 된 코니디아로 항상 실험을 수행해야합니다19.
- 장기간 보관을 위해, 포자를 -80 °C에서 25% 글리세롤 중의 고농축 포자 용액(약 1 x 10 8포자/mL)으로 동결시킨다(최대 1년까지 저장 가능). 다음 실험을 위해, 이들 글리세롤 스톡을 사용하여 단계 3.2에서 PDB 배지를 접종한다.
4. 페트리 접시에 기초한 멸균 체외 접종 시스템
참고: 페트리 접시 시스템(17)의 경우, 모든 장비 및 매체가 세균이 없고 모든 단계가 층류 후드에서 수행되는지 확인하십시오.
- 오토클레이빙 후 배지(1.3단계 참조)를 페트리 접시에 붓는다.
- 배지를 경화시킨 후 페트리 접시를 멸균 비닐 봉지에 다시 포장하고 냉장고 (4-10 ° C)에 밤새 거꾸로 보관하십시오. 차가운 매체는 다음 단계에서 매체의 슬라이딩을 방지하는 데 도움이됩니다.
- 감염 채널을 절단하고 메스로 응고된 배지의 상부 삼분의 일을 제거한다(도 1A). 절단하는 동안 한천 매체 아래에 액체 또는 공기가 생기지 않도록하십시오. 그렇지 않으면 배지가 미끄러져 감염 채널을 닫습니다.
- 50-100 개의 표면 멸균 애기장대 씨앗을 절단 된 윗면에 멸균 피펫 팁으로 배포하십시오. 자른 한천 표면이 페트리 접시의 벽과 접촉하는 각도에 씨앗을 넣어 뿌리가 배지와 페트리 접시 벽 사이에서 자랄 수 있도록하십시오. 이것은 나중에 접종을 촉진 할 것입니다.
- 페트리 접시를 닫고 공기 투과성 접착 테이프로 밀봉하여 가스 교환을 허용합니다.
- 4°C의 어둠 속에서 2일 동안 층층화한 후, 플레이트를 적합한 랙에 수직으로 놓고 성장 챔버 내의 장일 조건(16 h 빛/8시간 어둠) 하에서 22°C± 1°C에서 식물을 성장시킨다.
- 뿌리의 대부분이 감염 채널 (약 9-11 일 된 묘목)에 도달하면 플레이트를 수평으로 놓고 열어 갓 수확 한 Verticillium conidia의 500 μL를 4 x 105 포자 / mL의 농도로 감염 채널에 직접 첨가하여 액체가 채널에 고르게 분포되어 있는지 확인하십시오.
- 유사하게, 포자 대신에 모의 용액 500 μL를 첨가하여 대조군 플레이트를 준비한다(무균 1/4 MS 배지).
- 액체가 흠뻑 젖을 때까지 플레이트를 수평으로 몇 분 동안 인큐베이션하고 플레이트가 다시 수직으로 설정되면 누출 될 수 없습니다. 그런 다음 뚜껑을 닫고 공기 투과성 접착 테이프로 플레이트를 밀봉하십시오.
- 플레이트를 성장 챔버 내에서 수직으로 인큐베이션한다. 선택적으로 뿌리 부분을 검은 종이 상자로 덮어 뿌리와 토양 매개 곰팡이를 어둡게하십시오 (19 참조).
- 연구 질문에 따라 접종 후 바람직한 시점에서 분석을 수행하십시오 (여기에 사용 된 정확한 시점은 그림 범례를 참조하십시오). 다음은 몇 가지 제안입니다.
- 뿌리에서 잎을 자르고 둘 다 따로 수확하십시오. 페트리 접시에서 한천 스트립을 꺼내 뿌리에 쉽게 접근하고 포셉을 사용하여 조심스럽게 한천에서 꺼내십시오. 모든 식물 재료를 액체 질소에서 즉시 동결하십시오.
- 샘플을 액체 질소로 분쇄하십시오. 잎 물질 100 mg으로부터 총 DNA를 추출하여 식물 DNA에 대한 진균 DNA의 양을 정량적 PCR(qPCR)을 통해 확인하였다(19 참조).
- 샘플을 액체 질소로 분쇄하십시오. 식물 재료 100mg을 취하여 총 RNA를 추출하십시오. 정량적 역전사 PCR(qRT-PCR)을 실시하여 감염 동안 식물 유전자(또는 곰팡이 유전자)의 발현을 확인한다(19 참조).
- 부상을 피하면서 한천에서 뿌리를 조심스럽게 제거하고 형광 현미경으로 검사하십시오.
- 식물 리포터 라인에서 마커 유전자의 유도를 결정한다 (예를 들어, 루시페라제, β-글루쿠로니다제, 또는 형광 리포터17,19,21).
- 진균 리포터 라인(예를 들어, V. longisporum 구성적으로 향상된 녹색 형광 단백질, Vl-sGFP9)을 사용하거나 염색 기술(예를 들어, 5-브로모-4-클로로-3-인독실-N-아세틸-베타-d-글루코사미니드(X-beta-D-Glc-Nac)18)을 통해 뿌리에서의 진균 전파를 시각화한다.
- 뿌리에서 잎을 자르고 둘 다 따로 수확하십시오. 페트리 접시에서 한천 스트립을 꺼내 뿌리에 쉽게 접근하고 포셉을 사용하여 조심스럽게 한천에서 꺼내십시오. 모든 식물 재료를 액체 질소에서 즉시 동결하십시오.
5. 플라스틱 컵으로 조직 된 멸균 체외 접종 시스템
참고: 이 기술(19)의 첫 번째 설명에서 언급한 바와 같이, 모든 장비 및 매체가 세균이 없고 모든 단계가 층류 후드에서 수행되는지 확인하십시오.
- 총 부피가 500mL인 투명 플라스틱 컵을 사용하여 70%-75% 에탄올 욕조에서 최소 20분 동안 살균하십시오. 층류 후드에서 컵을 말리십시오.
- 오토클레이브 매체(1.4단계 참조)를 플라스틱 컵에 붓습니다. 선택적으로 박테리아 오염을 방지하기 위해 오토클레이브 배지에 cefotaxime (최종 농도 50 mg / L)을 첨가하십시오. 애기장대 실험을 위해 컵당 150mL의 배지를 사용하거나 더 큰 식물 종(유채 유채, 토마토)을 실험하기 위해 그 이상의 배지(컵당 250-300mL)를 사용합니다.
- 플라스틱 층(20분 동안 70%-75% 에탄올에서 인큐베이션하여 멸균 전)을 응고되기 전에 배지 상에 놓는다(도 1B).
참고 :이 플라스틱 층에는 표면 살균 된 씨앗을 배치하기 위해 모서리에 네 개의 조립식 구멍이 있습니다. 이것은 씨앗이 배지에 접근 할 수있게합니다. 나중에이 분리 층은 잎이 곰팡이 함유 배지에 닿지 않도록하여 미생물이 잎을 직접 공격 할 수 없으며 뿌리 경로를 취해야합니다. 또 다른 구멍이 중앙에 있어 감염 채널을 절단할 수 있습니다. - 배지가 응고되면 조립식 중앙 구멍을 통해 메스로 한천을 약 1.5cm 깊이로 자릅니다. 절단 된 한천을 제거하여 곰팡이 포자를 나중에 추가 할 수있는 감염 채널을 만듭니다.
- 응고 된 피부를 방해하기 위해 네 개의 작은 구멍에 피펫 팁이있는 한천 배지를 약간 긁으십시오 (이것은 씨앗이 물 한천 배지에서 물을 흡수 할 수있게합니다). 피펫 팁을 사용하여 씨앗을 작은 구멍에 넣으십시오.
- 두 번째 거꾸로 된 플라스틱 컵으로 플라스틱 컵을 닫고 공기 투과성 접착 테이프로 밀봉하십시오. 테이프는 가스 교환을 허용해야 합니다.
- 4°C에서 어둠 속에서 3일 동안 층화한 후, 컵 시스템을 22°C 및 60% 습도의 항온에서 성장 챔버 내에서 12 h 빛/12 h 어둠 (아라비돕시스, 오일시드 유채) 또는 16 h 빛/8 h 어둠 조건 (토마토) 하에 인큐베이션한다.
- 예방 접종을위한 식물의 권장 연령을 따르십시오 : Arabidopsis의 경우 21 일; 오일 시드 유채의 경우 5-7 일; 토마토 12 일.
- 감염 채널에 1mL의 코니디아 용액(권장 농도: 4 x 105포자/mL)을 첨가하여 버티실륨으로 플랜틀릿을 접종합니다. 대조군 샘플을 준비하려면 포자가 없는 모의 용액 1mL(무균 1/4 MS 배지)를 채널에 첨가한다.
- 연구 질문에 따라 접종 후 바람직한 시점에서 분석을 수행하십시오 (여기에 사용 된 정확한 시점은 그림 범례를 참조하십시오). 다음은 몇 가지 제안입니다.
- 위에서 디지털 카메라로 식물의 사진을 찍어 각 사진에 대해 거리를 동일하게 유지하십시오. 잎 면적을 정량화하고(예를 들어, ImageJ22 또는 BlattFlaeche17,19; 컵의 길이를 사용하여 스케일을 설정함) 감염된 그룹과 대조군을 비교한다. 질병 증상의 발달을 분류하십시오 (예 : 더 작은, 더 황색을 띠거나, 괴사 성 잎).
참고 : Arabidopsis에 줄기가있는 경우 줄기를 제거하여 로제트의 더 나은 사진을 얻으십시오. - 뿌리를 제거하고 감염된 싹으로부터 싹의 바이오매스(신선한 무게)를 정의하고 칭량에 의해 샘플을 조절한다. 상대적인 신선한 무게를 결정하십시오19.
- 분자 분석을 위한 샘플을 다음과 같이 수집한다.
- Arabidopsis : 줄기가있는 경우 줄기를 제거하십시오. 뿌리에서 바닥에있는 로제트를 자릅니다. 샘플에서 모든 뿌리 물질을 배제하고 전체 로제트를 수확하십시오. 다른 식물의 4-5 개의 로제트를 하나의 샘플로 결합하고 잎 물질을 액체 질소로 동결시킵니다.
- 포셉으로 배지에서 뿌리를 조심스럽게 꺼내고, 종이 타월로 눌러서 한천 잔해를 제거하고, 다른 식물의 4-5 뿌리를 하나의 샘플로 결합하십시오. 액체 질소에서 즉시 동결하십시오.
- 유채 / 토마토 : hypocotyl에서 줄기 세그먼트를 자릅니다 (예 : 길이 1cm, 항상 동일한 줄기 부위를 취하십시오). 4-5 식물의 재료를 각 샘플에 결합하고 액체 질소에서 동결하십시오.
- 샘플을 액체 질소로 분쇄하십시오. 잎 또는 줄기 물질 100 mg으로부터 총 DNA를 추출하여 qPCR을 통해 식물 DNA에 대한 진균 DNA의 양을 확인하였다(19 참조).
- 샘플을 액체 질소로 갈아서 식물 재료 100mg을 취하여 총 RNA를 추출하십시오. 감염시 식물 유전자 (또는 곰팡이 유전자)의 발현을 확인하기 위해 qRT-PCR을 수행하십시오 (19 참조).
- 위에서 디지털 카메라로 식물의 사진을 찍어 각 사진에 대해 거리를 동일하게 유지하십시오. 잎 면적을 정량화하고(예를 들어, ImageJ22 또는 BlattFlaeche17,19; 컵의 길이를 사용하여 스케일을 설정함) 감염된 그룹과 대조군을 비교한다. 질병 증상의 발달을 분류하십시오 (예 : 더 작은, 더 황색을 띠거나, 괴사 성 잎).
6. 화분에 있는 토양 기반 접종 체계
- 토양과 모래를 3:1 (토양 : 모래) 부피 비율로 철저히 혼합하여 뿌리에서 기질을 쉽게 씻을 수 있습니다. 혼합물을 오토클레이브 백에 붓습니다. 혼합물이 너무 건조하면 적절한 양의 물을 넣고 기질에 섞으십시오. 오토클레이브에서 20분 동안 80°C에서 증기를 공급하여 미생물 오염을 최소화한다.
참고 : 유기 토양 영양소에 영향을 줄 수 있으므로 80 ° C 이상으로 가열하지 마십시오. - 냄비에 흙 - 모래 혼합물을 채우고 쟁반으로 옮깁니다. 냄비 높이의 약 1/3 정도의 트레이에 물을 넣어 토양 - 모래 혼합물이 물에 완전히 담그도록하십시오. 또한, 젖은 시작 조건을 보장하기 위해 스프레이 병으로 기판에 물을 분사하십시오.
- 각 냄비에 3-4 개의 씨앗을 뿌리십시오 (그림 1C) 씨앗이 서로 충분한 거리를 갖도록 하십시오. 발아를 동기화하기 위해 층화를 위해 4 °C의 어둠 속에서 3 일 동안 보관하십시오.
참고 : 접종 실험을 위해 비슷한 크기의 식물을 선택할 수있게하고 개인차로 인한 편차를 줄이는 과량의 식물을 미리 재배하십시오. - 묘목이 규칙적인 급수로 긴 하루 조건 (16 시간 빛 / 8 시간 어둠, 22 ° C의 일정한 온도, 60 % 습도)에서 자라게하십시오.
- 예방 접종을위한 식물의 권장 연령을 따르십시오 : Arabidopsis의 경우 21 일, 유채 유채의 경우 7 일, 토마토의 경우 10 일. "뿌리 딥 접종"15,17,23,24을 수행하기 위해 비슷한 크기의 식물을 선택하십시오. 냄비에서 토양을 꺼내 조심스럽게 뿌리를 굴착하십시오.
- 물 용기에 뿌리 만 부드럽게 씻고 로제트를 물에서 꺼내십시오. 세척된 뿌리를 Verticillium 포자 용액이 들어있는 페트리 접시에서 60분 동안 배양한다(권장 농도: 2 x 106 포자/mL). 감염되지 않은 대조군의 경우, 포자가 없는 모의 용액(무균 1/4 MS 배지)에서 뿌리를 60분 동안 인큐베이션한다.
- 모래없이 촉촉하고 증기 살균 된 토양 (20 분 동안 80 ° C)으로 새로운 냄비를 준비하십시오. 피펫 팁을 사용하여 각 냄비의 토양 중앙에 하나의 구멍을 만듭니다.
- 뿌리를 구멍에 직접 놓습니다 (냄비 당 하나의 식물 만 옮기십시오). 뿌리를 삽입 한 후 조심스럽게 구멍을 흙으로 다시 채우십시오. 토양을 누르지 말고, 그렇지 않으면 리포팅은 보라색 잎과 같은 스트레스 증상을 유발할 수 있습니다.
- 정기적 인 급수로 장일 조건 (16 시간 빛 / 8 시간 어둠, 22 °C의 일정한 온도, 60 % 습도)에서 감염된 그룹과 대조군을 배양하십시오.
- 연구 질문에 따라 접종 후 바람직한 시점에서 분석을 수행하십시오 (여기에 사용 된 정확한 시점은 그림 범례를 참조하십시오). 다음은 몇 가지 제안입니다.
- 위에서 디지털 카메라로 식물의 사진을 찍어 각 사진에 대해 거리를 동일하게 유지하십시오. 잎 면적을 정량화하고(예를 들어, ImageJ22 또는 BlattFlaeche17,19; 화분의 직경을 사용하여 스케일을 설정함) 감염된 그룹과 대조군을 비교한다. 질병 증상(예를 들어, 더 작은, 더 황색을 띤 또는 괴사성 잎)의 발달을 분류한다13.
참고 : 애기장대의 줄기를 제거하면 로제트의 사진을 쉽게 찍을 수 있습니다. - 뿌리를 제거하고 감염된 싹으로부터 싹의 바이오매스(신선한 무게)를 정의하고 칭량에 의해 샘플을 조절한다. 상대적인 신선한 무게를 결정하십시오19.
- 대안적으로, 식물 높이를 측정하거나 줄기 세그먼트로부터의 진균 발육을 분류하여 질병 중증도를 평가한다(13).
- 다음과 같이 분자 분석을 위한 샘플을 수집합니다.
- Arabidopsis : 줄기를 제거하십시오. 뿌리 크라운에서 로제트를 자릅니다. 다른 식물의 4-5 개의 로제트를 하나의 샘플로 결합하십시오. 잎 물질을 액체 질소에 동결하십시오.
참고 : 뿌리의 경우, 세척을 통해 유전자 발현을 재 프로그래밍하지 않고 토양에서 충분히 청소하기가 어렵습니다. - 유채 / 토마토 : hypocotyl에서 줄기 세그먼트를 자릅니다 (예 : 길이 1cm, 항상 동일한 줄기 부위를 취하십시오). 4-5 식물의 재료를 하나의 샘플로 결합하고 액체 질소에서 동결시킵니다.
- 샘플을 액체 질소로 분쇄하십시오. 100 mg 잎 또는 줄기 물질로부터 총 DNA를 추출하여 qPCR을 통해 식물 DNA에 대한 진균 DNA의 양을 결정한다(19 참조).
- 샘플을 액체 질소로 갈아서 식물 재료 100mg을 취하여 총 RNA를 추출하십시오. 감염시 식물 유전자 (또는 곰팡이 유전자)의 발현을 확인하기 위해 qRT-PCR을 수행하십시오 (19 참조).
- 위에서 디지털 카메라로 식물의 사진을 찍어 각 사진에 대해 거리를 동일하게 유지하십시오. 잎 면적을 정량화하고(예를 들어, ImageJ22 또는 BlattFlaeche17,19; 화분의 직경을 사용하여 스케일을 설정함) 감염된 그룹과 대조군을 비교한다. 질병 증상(예를 들어, 더 작은, 더 황색을 띤 또는 괴사성 잎)의 발달을 분류한다13.
7. 데이터 분석
- 생물학적 반복실험에 기초하여 평균 및 표준 편차(± SD)를 계산한다.
- 감염된 그룹의 모든 결과를 대조군의 결과로 나누어 상대 값을 계산합니다. 예를 들어 평균을 "모의 상대" 또는 "야생형에 상대적"으로 표시합니다.
- 그룹 간의 통계적 유의성을 결정합니다.

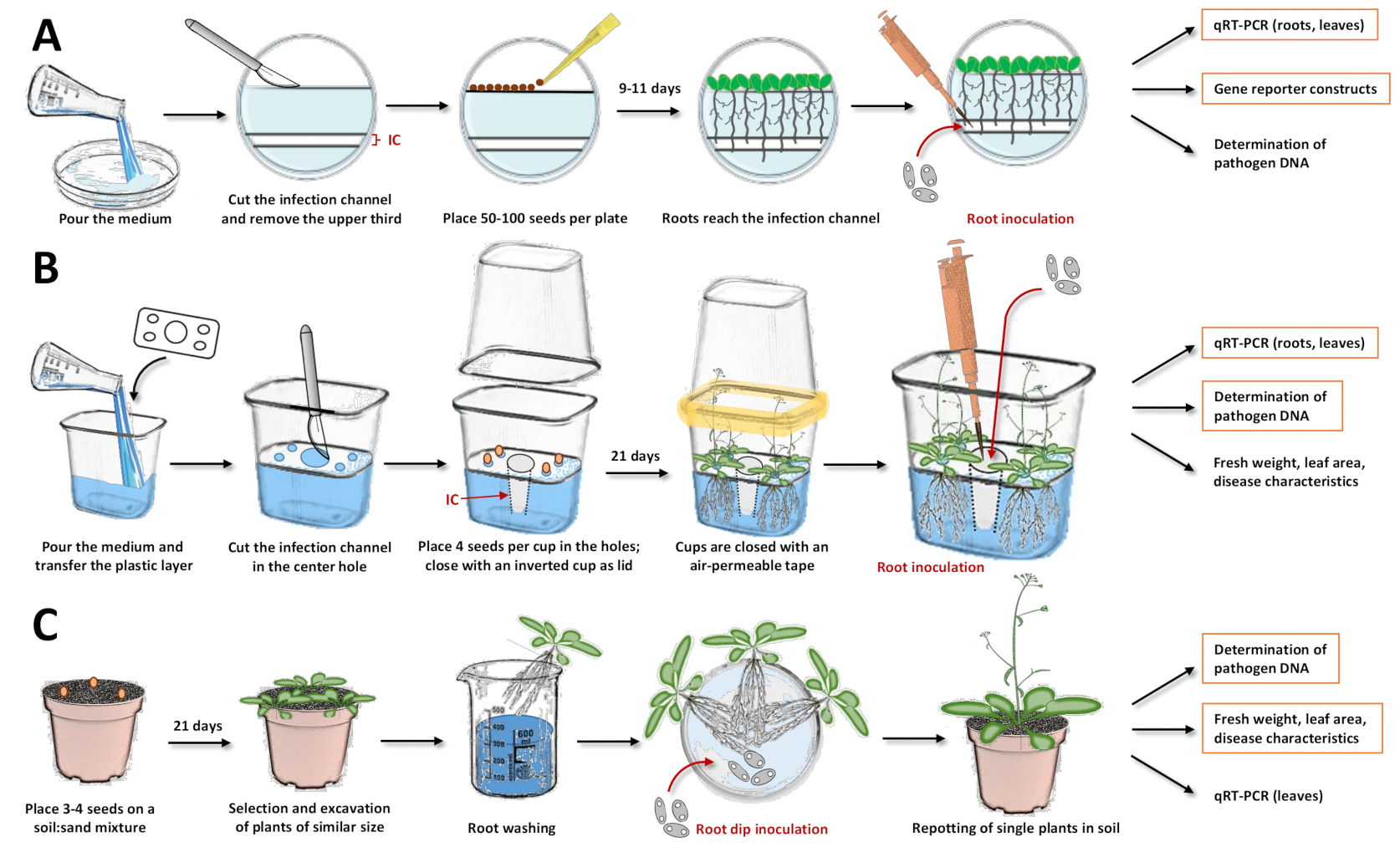
그림 1: 세 가지 접종 시스템 및 프로토콜의 개별 단계 편집. 이 수치는 모델 식물 애기장대 탈리아나를 사용한 시스템을 보여줍니다. 다른 식물 종의 경우시기를 조정해야합니다. 주황색 상자가 강조 표시되며, 각 시스템에서 후속 분석이 가장 권장됩니다. (A) 페트리 접시17의 접종 시스템의 경우, 배지를 부어 고형화시킨다. 접시를 냉장고에 하룻밤 사이에 보관하십시오. 이어서, 메스로 감염 채널(IC)뿐만 아니라 상부 세 번째를 절단하고 제거하였다(그림의 흰색 영역은 한천으로부터 제거되었고, 푸르스름한 영역은 한천을 나타냄). 절단 표면에 씨앗을 놓고 페트리 접시를 닫습니다. 층화 후 플레이트를 수직으로 놓고 식물이 자라게하십시오. 대부분의 뿌리가 감염 채널에 도달하면 피펫이있는 포자 용액을 채널에 직접 추가하십시오. 솔루션이 고르게 분포되어 있는지 확인하십시오. 페트리 접시를 닫고 성장 챔버에서 수직으로 배양하십시오. 뒤따를 수 있는 접근법은 정량적 역전사 PCR(qRT-PCR)을 이용한 발현 분석, 리포터 라인을 이용한 현미경 검사, 미생물 DNA의 정량화이다. (B) 플라스틱 컵(19)에 접종 시스템을 위해, 배지를 붓고 분리된 플라스틱 층을 조립식 구멍(종자를 배치하기 위한 모서리에 네 개의 작은 구멍과 감염 채널의 중앙에 하나의 큰 구멍)으로 옮긴다. 매체가 굳어지게하십시오. 메스로 중앙구멍에 있는 한천 배지를 절단 및 제거하여 감염 채널(IC)을 얻었다. 작은 구멍에서 배지를 긁고 씨앗을 옮깁니다. 컵을 거꾸로 한 컵으로 닫고 공기 투과성 테이프(노란색으로 표시)로 밀봉합니다. 식물이 자라게하십시오. 예방 접종을 위해 피펫이있는 포자 용액을 감염 채널에 직접 추가하십시오. 시스템을 닫고 성장 챔버에서 재배를 계속하십시오. 뒤따를 수 있는 접근법은 qRT-PCR을 이용한 발현 분석, 미생물 DNA의 정량화, 신선한 무게, 잎 면적 또는 다른 질병 특성의 결정이다. (C) "뿌리 딥 접종"15,17,23,24 : 토양 기반 접종 시스템의 경우 냄비에 토양 : 모래 혼합물을 채 웁니다. 씨앗을 옮기고 묘목이 자라게하십시오. 비슷한 크기의 식물을 발굴하고 뿌리를 물로 씻으십시오. 씻은 뿌리를 포자로 용액을 담고있는 페트리 접시에 넣으십시오. 배양 후, 토양이있는 화분에 단일 식물을 넣으십시오. 따를 수 있는 접근법은 qRT-PCR을 갖는 잎에서의 발현 분석, 미생물 DNA의 정량화, 신선한 중량, 잎 면적 또는 다른 질병 특성의 결정이다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
결과
프로토콜에 따라, 식물을 재배하고 V. longisporum (균주 Vl4325) 또는 V. dahliae (JR218 단리물)로 접종하였다. 다양한 시나리오는 효과를 입증하고 주어진 프로토콜의 일부 기능을 강조하기 위해 설계되었습니다. 대표적인 결과가 표시됩니다.
항균성 인돌글루코시놀레이트(IG) 생합성에 관여하는 유전자의 발현 유도는 버티실...
Access restricted. Please log in or start a trial to view this content.
토론
토양 매개 식물 병원균1로 인한 엄청난 수확량 손실로 인해 농업 전략 또는 작물 품종의 개선이 필요합니다. 토양 매개 질병의 발병기전에 대한 제한된 통찰력은 더 저항력이 강한 식물의 발달을 방해합니다. 근본적인 병리학 적 메커니즘을 탐구 할 필요가 있으며, 이를 위해서는 강력한 방법론 적 플랫폼이 필요합니다. 보고된 접종 절차는 뿌리-미생물 상호작용에서의 다인자 ?...
Access restricted. Please log in or start a trial to view this content.
공개
저자는 공개 할 것이 없습니다.
감사의 말
저자들은 Tim Iven과 Jaqueline Komorek이 이러한 방법에 대한 이전 연구, 볼프강 Dröge-Laser (독일 뷔르츠부르크 대학 제약 생물학과)가이 작업에 필요한 장비와 자원을 제공했으며, Wolfgang Dröge-Laser와 Philipp Kreisz (뷔르츠부르크 대학 모두)가 원고의 비판적 교정을 위해 인정합니다. 이 연구는 "Deutsche Forschungsgemeinschaft"(DFG, DR273/15-1,2)에 의해 뒷받침되었다.
Access restricted. Please log in or start a trial to view this content.
자료
| Name | Company | Catalog Number | Comments |
| Agar (Gelrite) | Carl Roth | Nr. 0039 | all systems described require Gelrite |
| Arabidopsis thaliana wild-type | NASC stock | Col-0 (N1092) | |
| Autoclave | Systec | VE-100 | |
| BlattFlaeche | Datinf GmbH | BlattFlaeche | software to determine leaf areas |
| Brassica napus wild-type | see Floerl et al., 2008 | rapid-cycling rape | genome ACaacc |
| Cefotaxime sodium | Duchefa | C0111 | |
| Chicanery flask 500 mL | Duran Group / neoLab | E-1090 | Erlenmeyer flask with four baffles |
| Collection tubes 50 mL | Sarstedt | 62.547.254 | 114 x 28 mm |
| Czapek Dextrose Broth medium | Duchefa | C1714 | |
| Digital camera | Nikon | D3100 18-55 VR | |
| Exsiccator (Desiccator ) | Duran Group | 200 DN, 5.8 L | Seal with lid to hold chlorine gas |
| Fluorescence Microscope | Leica | Leica TCS SP5 II | |
| HCl | Carl Roth | P074.3 | |
| KNO3 | Carl Roth | P021.1 | ≥ 99 % |
| KOH | Carl Roth | 6751 | |
| Leukopor | BSN medical GmbH | 2454-00 AP | non-woven tape 2.5 cm x 9.2 m |
| MES (2-(N-morpholino)ethanesulfonic acid) | Carl Roth | 4256.2 | Pufferan ≥ 99 % |
| MgSO4 | Carl Roth | T888.1 | Magnesiumsulfate-Heptahydrate |
| Murashige & Skoog medium (MS) | Duchefa | M0222 | MS including vitamins |
| NaClO | Carl Roth | 9062.1 | |
| Percival growth chambers | CLF Plant Climatics GmbH | AR-66L2 | |
| Petri-dishes | Sarstedt | 82.1473.001 | size ØxH: 92 × 16 mm |
| Plastic cups (500 mL, transparent) | Pro-pac, salad boxx | 5070 | size: 108 × 81 × 102 mm |
| Pleated cellulose filter | Hartenstein | FF12 | particle retention level 8–12 μm |
| poly klima growth chamber | poly klima GmbH | PK 520 WLED | |
| Potato Dextrose Broth medium | SIGMA Aldrich | P6685 | for microbiology |
| Pots | Pöppelmann GmbH | TO 7 D or TO 9,5 D | Ø 7 cm resp. Ø 9.5 cm |
| PromMYB51::YFP | see Poncini et al., 2017 | MYB51 reporter line | YFP (i.e. 3xmVenus with NLS) |
| Reaction tubes 2 mL | Sarstedt | 72.695.400 | PCR Performance tested |
| Rotary (orbital) shaker | Edmund Bühler | SM 30 C control | |
| Sand (bird sand) | Pet Bistro, Müller Holding | 786157 | |
| Soil | Einheitserde spezial | SP Pikier (SP ED 63 P) | |
| Solanum lycopersicum wild-type | see Chavarro-Carrero et al., 2021 | Type: Moneymaker | |
| Thoma cell counting chamber | Marienfeld | 642710 | depth 0.020 mm; 0.0025 mm2 |
| Ultrapure water (Milli-Q purified water) | MERK | IQ 7003/7005 | water obtained after purification |
| Verticillium dahliae | see Reusche et al., 2014 | isolate JR2 | |
| Verticillium longisporum | Zeise and von Tiedemann, 2002 | strain Vl43 |
참고문헌
- Mendes, R., Garbeva, P., Raaijmakers, J. M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiology Review. 37 (5), 634-663 (2013).
- Yadeta, K. A., Thomma, B. P. H. J. The xylem as battleground for plant hosts and vascular wilt pathogens. Frontiers in Plant Science. 4, 97(2013).
- Delgado-Baquerizo, M., et al. The proportion of soil-borne pathogens increases with warming at the global scale. Nature Climate Change. 10 (6), 550-554 (2020).
- Berendsen, R. L., et al. Disease-induced assemblage of a plant-beneficial bacterial consortium. The ISME Journal. 12 (6), 1496-1507 (2018).
- Yuan, J., et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome. 6 (1), 156(2018).
- Liu, H., et al. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytologist. 229 (5), 2873-2885 (2021).
- Wang, H., Liu, R., You, M. P., Barbetti, M. J., Chen, Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): role of bacterial diversity. Microorganisms. 9 (9), 1988(2021).
- Inderbitzin, P., Subbarao, K. V. Verticillium systematics and evolution: how confusion impedes Verticillium wilt management and how to resolve it. Phytopathology. 104 (6), 564-574 (2014).
- Eynck, C., Koopmann, B., Grunewaldt-Stoecker, G., Karlowsky, P., von Tiedemann, A. Differential interactions of Verticillium longisporum und V. dahliae with Brassica napus with molecular and histological techniques. European Journal of Plant Pathology. 118 (3), 259-274 (2007).
- Floerl, S., et al. Defence reactions in the apoplastic proteome of oilseed rape (Brassica napus var. napus) attenuate Verticillium longisporum growth but not disease symptoms. BMC Plant Biology. 8, 129(2008).
- Leonard, M., et al. Verticillium longisporum elicits media-dependent secretome responses with capacity to distinguish between plant-related environments. Frontiers in Microbiology. 11, 1876(2020).
- Depotter, J. R. L., et al. Verticillium longisporum, the invisible threat to oilseed rape and other brassicaceous plant hosts. Molecular Plant Pathology. 17 (7), 1004-1016 (2016).
- Fröschel, C., et al. A gain-of-function screen reveals redundant ERF transcription factors providing opportunities for resistance breeding toward the vascular fungal pathogen Verticillium longisporum. Molecular Plant-Microbe Interactions. 32 (9), 1095-1109 (2019).
- Zhou, L., Hu, Q., Johansson, A., Dixelius, C. Verticillium longisporum and V. dahliae: infection and disease in Brassica napus. Plant Pathology. 55 (1), 137-144 (2006).
- Ralhan, A., et al. The vascular pathogen Verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots. Plant Physiology. 159 (3), 1192-1203 (2012).
- Reusche, M., et al. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium longisporum. Molecular Plant-Microbe Interactions. 26 (8), 850-860 (2013).
- Iven, T., et al. Transcriptional activation and production of tryptophan-derived secondary metabolites in Arabidopsis roots contributes to the defense against the fungal vascular pathogen Verticillium longisporum. Molecular Plant. 5 (6), 1389-1402 (2012).
- Reusche, M., et al. Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana. Environmental and Experimental Botany. 108, 23-37 (2014).
- Fröschel, C. In-depth evaluation of root infection systems using the vascular fungus Verticillium longisporum as soil-borne model pathogen. Plant Methods. 17 (1), 57(2021).
- Karapapa, V. K., Bainbridge, B. W., Heale, J. B. Morphological and molecular characterization of Verticillium longisporum comb, nov., pathogenic to oilseed rape. Mycological Research. 101 (11), 1281-1294 (1997).
- Poncini, L., et al. In roots of Arabidopsis thaliana, the damage-associated molecular pattern AtPep1 is a stronger elicitor of immune signalling than flg22 or the chitin heptamer. PLoS One. 12 (10), 1-21 (2017).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Fradin, E. F., et al. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiology. 150 (1), 320-332 (2009).
- Singh, S., et al. The plant host Brassica napus induces in the pathogen Verticillium longisporum the expression of functional catalase peroxidase which is required for the late phase of disease. Molecular Plant-Microbe Interactions. 25 (4), 569-581 (2012).
- Zeise, K., von Tiedemann, A. Application of RAPD-PCR for virulence type analysis within Verticillium dahliae and Verticillium longisporum. Journal of Phytopathology. 150 (10), 557-563 (2002).
- Fröschel, C., et al. Plant roots employ cell-layer-specific programs to respond to pathogenic and beneficial microbes. Cell Host & Microbe. 29 (2), 299-310 (2021).
- Gigolashvili, T., et al. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. The Plant Journal. 50 (5), 886-901 (2007).
- Back, M. A., Haydock, P. P. J., Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathology. 51 (6), 683-697 (2002).
- Behrens, F. H., et al. Suppression of abscisic acid biosynthesis at the early infection stage of Verticillium longisporum in oilseed rape (Brassica napus). Molecular Plant Pathology. 20 (12), 1645-1661 (2019).
- Vorholt, J. A., Vogel, C., Carlström, C. I., Müller, D. B. Establishing causality: opportunities of synthetic communities for plant microbiome research. Cell Host & Microbe. 22 (2), 142-155 (2017).
Access restricted. Please log in or start a trial to view this content.
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유