É necessária uma assinatura da JoVE para visualizar este conteúdo. Faça login ou comece sua avaliação gratuita.
Method Article
Estratégias de inoculação para infectar raízes vegetais com microrganismos transportados pelo solo
Neste Artigo
Resumo
Este protocolo apresenta um resumo detalhado das estratégias para inocular raízes vegetais com micróbios transportados pelo solo. Exemplificados para os fungos Verticillium longisporum e Verticillium dahliae, três diferentes sistemas de infecção radicular são descritos. Potenciais aplicações e possíveis análises a jusante são destacadas, e vantagens ou desvantagens são discutidas para cada sistema.
Resumo
A rizosfera abriga uma comunidade microbiana altamente complexa na qual as raízes das plantas são constantemente desafiadas. As raízes estão em contato próximo com uma grande variedade de microrganismos, mas estudos sobre interações transmitidas pelo solo ainda estão por trás daqueles realizados em órgãos acima do solo. Embora algumas estratégias de inoculação para infectar plantas modelo com patógenos radiculares modelos sejam descritas na literatura, ainda é difícil obter uma visão geral metodológica abrangente. Para resolver esse problema, três diferentes sistemas de inoculação radicular são precisamente descritos que podem ser aplicados para obter insights sobre a biologia das interações root-micróbios. Para ilustração, as espécies de Verticillium (ou seja, V. longisporum e V. dahliae) foram empregadas como patógenos de modelo invasores radiculares. No entanto, os métodos podem ser facilmente adaptados a outros micróbios colonizadores radiculares - tanto patogênicos quanto benéficos. Ao colonizar a planta xilema, fungos vasculares transmitidos pelo solo, como Verticillium spp. exibem um estilo de vida único. Após a invasão radicular, eles se espalham pelos vasos xilema acropetalmente, chegam ao tiro e provocam sintomas da doença. Três espécies representativas de plantas foram escolhidas como hospedeiras-modelo: Arabidopsis thaliana, estupro de oleosa economicamente importante (Brassica napus) e tomate (Solanum lycopersicum). Protocolos passo a passo são dados. Resultados representativos de ensaios de patogenicidade, análises transcritivas de genes marcadores e confirmações independentes por construções de repórteres são mostrados. Além disso, as vantagens e desvantagens de cada sistema de inoculação são exaustivamente discutidas. Esses protocolos comprovados podem auxiliar no fornecimento de abordagens para perguntas de pesquisa sobre interações entre micróbios radiculares. Saber como as plantas lidam com micróbios no solo é crucial para o desenvolvimento de novas estratégias para melhorar a agricultura.
Introdução
Solos naturais são habitados por um número surpreendente de micróbios que podem ser neutros, prejudiciais ou benéficos para as plantas1. Muitos patógenos vegetais são transportados pelo solo, cercam as raízes e atacam o órgão subterrâneo. Esses microrganismos pertencem a uma grande variedade de clados: fungos, oomycetes, bactérias, nematoides, insetos e alguns vírus 1,2. Uma vez que as condições ambientais favoreçam a infecção, as plantas suscetíveis se tornarão doentes e a produção de culturas diminuirá. Os efeitos das mudanças climáticas, como o aquecimento global e os extremos climáticos, aumentarão a proporção de patógenos vegetais transportados pelo solo3. Portanto, será cada vez mais importante estudar esses micróbios destrutivos e seu impacto na produção de alimentos e rações, mas também nos ecossistemas naturais. Além disso, há mutualistas microbianos no solo que interagem firmemente com raízes e promovem o crescimento, o desenvolvimento e a imunidade das plantas. Quando confrontadas com patógenos, as plantas podem recrutar ativamente oponentes específicos na rizosfera que podem suportar a sobrevivência do hospedeiro suprimindo patógenos 4,5,6,7. No entanto, detalhes mecanicistas e caminhos envolvidos em interações benéficas entre micróbios radiculares ainda são desconhecidos6.
É, portanto, essencial ampliar a compreensão geral das interações raiz-micróbios. Métodos confiáveis para inocular raízes com microrganismos transportados pelo solo são necessários para realizar estudos de modelo e transferir os achados para aplicações agrícolas. Interações benéficas no solo são estudadas, por exemplo, com serndipita indica (anteriormente conhecida como Piriformospora indica), fixação de nitrogênio Rhizobium spp., ou fungos micorrizais, enquanto os patógenos vegetais suportados pelo solo incluem Ralstonia solanacearum, Phytophthora spp., Fusarium spp., e Verticillium spp.1. Os dois últimos são gêneros fúngicos que são distribuídos globalmente e causam doenças vasculares2. Verticillium spp. (Ascomycota) pode infectar centenas de espécies vegetais - em grande parte dicotyledons, incluindo anuários herbáceos, perenes lenhosos e muitas plantasagrícolas 2,8. A hifa de Verticillium entra na raiz e cresce intercelularmente e intracelularmente em direção ao cilindro central para colonizar os vasos xilema 2,9. Nestes vasos, o fungo permanece durante a maior parte de seu ciclo de vida. Como a seiva de xilema é pobre em nutrientes e carrega compostos de defesa vegetal, o fungo deve se adaptar a este ambiente único. Isso é feito pela secreção de proteínas relacionadas à colonização que permitem que o patógeno sobreviva em seuhospedeiro 10,11. Depois de atingir a vasculatura raiz, o fungo pode se espalhar dentro dos vasos xilem acropetalmente para a folhagem, o que leva à colonização sistêmica do hospedeiro 9,12. Neste ponto, a planta é afetada negativamente no crescimentode 9,10,13. Por exemplo, a desnutrição e as folhas amarelas ocorrem, bem como a senescência prematura 13,14,15,16.
Um membro desse gênero é o Verticillium longisporum, que é altamente adaptado aos hospedeiros brassicaceos, como o estupro da oleosa agronomicamente importante, a couve-flor e a planta modelo Arabidopsis thaliana12. Vários estudos combinaram V. longisporum e A. thaliana para obter extensas percepções sobre doenças vasculares transmitidas pelo solo e as respostas de defesa raiz resultantes 13,15,16,17. Testes de suscetibilidade simples podem ser realizados usando o sistema modelo V. longisporum / A. thaliana e recursos genéticos bem estabelecidos estão disponíveis para ambos os organismos. Intimamente relacionado com V. longisporum é o patógeno Verticillium dahliae. Embora ambas as espécies fúngicas realizem um processo de vida vascular semelhante e de invasão, sua eficiência de propagação de raízes para folhas e os sintomas da doença provocada em A. thaliana são diferentes: enquanto V. longisporum geralmente induz senescência precoce, a infecção por V. dahliae resulta em murchar18. Recentemente, um resumo metodológico apresentou diferentes estratégias de inoculação radicular para infectar A. thaliana com V. longisporum ou V. dahliae, auxiliando no planejamento de configurações experimentais19. No campo, V. longisporum ocasionalmente causa danos significativos na produção de estupro da oleosa12, enquanto V. dahliae tem uma gama hospedeira muito ampla composta por várias espécies cultivadas, como videira, batata e tomate8. Isso torna ambos os modelos economicamente interessantes para estudar.
Assim, os protocolos a seguir usam v . longisporum e V. dahliae como patógenos radiculares modelo para exemplificar possíveis abordagens para inoculações radiculares. Arabidopsis (Arabidopsis thaliana), estupro de oleosa (Brassica napus) e tomate (Solanum lycopersicum) foram escolhidos como hospedeiros modelo. Descrições detalhadas das metodologias podem ser encontradas no texto abaixo e no vídeo que acompanha. São discutidas vantagens e desvantagens para cada sistema de inoculação. Em conjunto, essa coleção de protocolos pode ajudar a identificar um método adequado para questões específicas de pesquisa no contexto de interações root-micróbios.
Protocolo
1. Mídia para culturas fúngicas e sistemas de inoculação de plantas
- Caldo de desxtrose de batata líquida (PDB): Prepare 21 g/L PDB em água ultrapura em um frasco estável a calor.
- Caldo líquido Czapek Dextrose (CDB): Prepare 42 g/L CDB em água ultrauso em um frasco estável a calor.
- Meio para o sistema de inoculação da placa de Petri: Prepare um frasco estável a calor com 1,5 g/L Murashige e Skoog médio (MS) e 8 g/L de ágar em água ultrauso.
NOTA: Evite o açúcar neste meio, pois levará ao crescimento fúngico excessivo após a inoculação. - Meio para o sistema de inoculação à base de copos plásticos: Prepare um frasco estável a calor com 4,4 g/L MS, 0,2 g/L MgSO4, 1 g/L KNO3, 0,5 g/L 2-(N-morpholino)ácido esulfônico (MES) e 6,0 g/L de ágar em água ultrauso e ajustar pH a 5,7 com 5 M KOH.
NOTA: Evite o açúcar neste meio, pois levará ao crescimento fúngico excessivo após a inoculação. - 1/4 MS médio: Prepare 1,2 g/L MS em água ultrapura.
- Use a autoclave para esterilizar todas as soluções acima. Coloque os frascos de vidro na cesta, feche a tampa e esterilize por 15 min a 121 °C e 98,9 kPa.
2. Esterilizando a superfície das sementes vegetais
NOTA: Use sempre o protocolo abaixo para esterilizar a superfície das sementes da Arabidopsis, estupro da oleosa e tomate antes da semeadura.
- Transfira as sementes para um tubo de reação de 2 mL. Coloque o tubo em um exsiccator com uma capacidade interna de 5,8 L.
- Gere gás cloro no exsiccator adicionando 6 mL de ácido clorídrico (HCl) em 100 mL de hipoclorito de sódio aquoso de 12% (NaClO).
- Feche imediatamente a tampa da exsiccator e incuba as sementes por 3h no gás.
3. Preparando o inóculo com esporos de Verticillium (conidia derivada assexual)
NOTA: Cultivar V. dahliae (cepa JR2) da mesma forma que V. longisporum (cepa Vl43)17,18,19. Certifique-se de que todos os equipamentos e mídias estão livres de germes e que todas as etapas sejam executadas em um capô de fluxo laminar para manter o machado inóculo.
- Encha 150 mL de PDB líquido (etapa 1.1) em um frasco de chicanaria de 500 mL e suplemente o médio com 500 mg/L cefotaxime.
- Adicione Verticillium conida do armazenamento de glicerol ao meio PDB. Feche o frasco com uma rolha de espuma estéril.
- Incubar a cultura por 7-10 dias em uma caixa escura à temperatura ambiente (RT) sob agitação contínua e horizontal (agitador rotativo; 60 rpm). Isso resulta em pequenas esferas de micélia branca.
- Remova e descarte cuidadosamente o supernatante PDB. A maior parte da micélio deve permanecer no frasco.
- Adicione 100 mL de CDB líquido (etapa 1.2) na micélia no frasco chicâneo e suplemente o médio com 500 mg/L cefotaxime.
- Incubar mais 4-5 dias em uma caixa escura na RT sob agitação contínua e horizontal (agitador rotativo, 60 rpm) para induzir a esporulação. O supernatante ficará amarelado-acinzento à medida que a conídia for liberada.
- Filtre uma porção (5-10 mL) do líquido contendo conidia através de um papel filtro (nível de retenção de partículas de 8-12 μm) em um tubo de coleta estéril de 50 mL. Isso separa esporos da micélia.
- Determine a concentração de esporos usando uma câmara de contagem de células e um microscópio. Diluir com meio de 1/4 MS sem germes em água ultrapura até que as concentrações de esporos dadas abaixo sejam obtidas.
NOTA: Sob o microscópio, as conidia de V. longisporum são em sua maioria desenhadas longamente e 7,1-8,8 μm de tamanho, enquanto V. dahliae conidia são mais curtas (3,5-5 μm) e bastanteesféricas 20. - Use essas conidia recém-colhidas como inóculo. Certifique-se de realizar os experimentos sempre com conidia recém-colhida e não com estoques congelados, pois o congelamento reduz significativamente o número de esporos viáveis19.
- Para armazenamento a longo prazo, congele os esporos como solução de esporos concentrados (aproximadamente 1 x 108 esporos/mL) em 25% de glicerol a -80 °C (armazenado até 1 ano). Para os próximos experimentos, use esses estoques de glicerol para inocular o meio PDB na etapa 3.2.
4. Um sistema de inoculação in vitro estéril baseado em placas de Petri
NOTA: Para o sistema de placas de Petri17, certifique-se de que todos os equipamentos e mídias são livres de germes e que todas as etapas sejam realizadas em um capô de fluxo laminar.
- Depois de autoclavar, despeje o meio (ver passo 1.3) em pratos de Petri.
- Após o endurecimento do meio, reembale as placas de Petri em um saco plástico estéril e guarde-as de cabeça para baixo durante a noite na geladeira (4-10 °C). Um meio refrigerado ajuda a evitar o deslizamento do meio nos próximos passos.
- Corte e remova um canal de infecção e o terço superior do meio solidificado com um bisturi (Figura 1A). Evite obter líquido ou ar sob o meio ágar durante o corte; caso contrário, o meio escorregará e fechará o canal de infecção.
- Distribua 50-100 sementes arabidopsis esterilizadas pela superfície com uma ponta de pipeta estéril na superfície superior cortada. Coloque as sementes no ângulo onde a superfície do ágar cortada entra em contato com a parede da placa de Petri para que as raízes possam crescer entre o meio e a parede da placa de Petri. Isso facilitará a inoculação mais tarde.
- Feche as placas de Petri e seque-as com fita adesiva permeável a ar para permitir a troca de gás.
- Após a estratificação por 2 dias na escuridão a 4 °C, coloque as placas verticalmente em um rack adequado e cresça as plantas a 22 °C ± 1 °C em condições de longo dia (16h de luz / 8h de escuridão) em uma câmara de crescimento.
- Quando a maioria das raízes chegar ao canal de infecção (cerca de 9-11 dias de idade, colocar as placas horizontalmente, abri-las e adicionar 500 μL de conidia verticillium recém-colhida com uma concentração de 4 x 105 esporos/mL diretamente no canal de infecção, certificando-se de que o líquido seja distribuído uniformemente no canal.
- Da mesma forma, prepare as placas de controle adicionando 500 μL de uma solução simulada em vez de esporos (meio de 1/4 MS sem germes).
- Incubar as placas horizontalmente por alguns minutos até que o líquido esteja encharcado e não possa vazar quando as placas estiverem configuradas verticalmente novamente. Em seguida, feche a tampa e sele as placas com fita adesiva permeável a ar.
- Incubar as placas verticalmente na câmara de crescimento. Opcionalmente, cubra as partes radiculares com caixas de papel pretas para escurecer raízes e fungos transportados pelo solo (ver19).
- Realize as análises nos pontos de tempo preferidos após a inoculação dependendo da questão da pesquisa (consulte as legendas de figura para os pontos de tempo exatos utilizados aqui). A seguir estão algumas sugestões.
- Corte as folhas das raízes e colhe as duas separadamente. Retire as tiras de ágar das placas de Petri para acessar facilmente as raízes e puxá-las cuidadosamente para fora do ágar usando fórceps. Congele todo o material vegetal imediatamente em nitrogênio líquido.
- Triture as amostras em nitrogênio líquido. Extrair DNA total de 100 mg de material de folha para determinar através de um PCR quantitativo (qPCR) a quantidade de DNA fúngico em relação ao DNA vegetal (ver19).
- Triture as amostras em nitrogênio líquido. Pegue 100 mg de material vegetal e extraia RNA total. Realizar transcrição reversa quantitativa PCR (qRT-PCR) para determinar a expressão de genes vegetais (ou genes fúngicos) durante a infestação (ver19).
- Remova cuidadosamente as raízes do ágar evitando lesões e examine-as sob o microscópio fluorescente.
- Determinar a indução de genes marcadores em linhas de repórteres vegetais (por exemplo, luciferase, β-glucuronidase ou repórteres fluorescentes 17,19,21).
- Visualize a propagação fúngica na raiz usando linhas de repórter fúngicos (por exemplo, V. longisporum expressando com alguma forma proteína fluorescente verde aprimorada, VL-sGFP 9) ou por técnicas de coloração (por exemplo, através de 5-bromo-4-cloro-3-indoxyl-N-acetyl-beta-d-glucosaminida (X-beta-D-Glc-Nac)18).
- Corte as folhas das raízes e colhe as duas separadamente. Retire as tiras de ágar das placas de Petri para acessar facilmente as raízes e puxá-las cuidadosamente para fora do ágar usando fórceps. Congele todo o material vegetal imediatamente em nitrogênio líquido.
5. Um sistema de inoculação in vitro estéril organizado com copos plásticos
NOTA: Como observado na primeira descrição desta técnica19, certifique-se de que todos os equipamentos e mídias estão livres de germes e que todas as etapas sejam executadas em uma capa de fluxo laminar.
- Utilize copos plásticos transparentes com um volume total de 500 mL e esterilize-os em um banho de etanol de 70%-75% por pelo menos 20 min. Seque as xícaras no capô de fluxo laminar.
- Despeje o meio autoclavado (ver passo 1.4) nos copos de plástico. Opcionalmente, adicione cefotaxime (concentração final de 50 mg/L) ao meio autoclavado para evitar contaminações bacterianas. Use 150 mL de média por xícara para experimentos com arabidopsis ou mais médios (250-300 mL por xícara) para experimentos com espécies vegetais maiores (estupro de oleosa, tomate).
- Coloque uma camada plástica (esterilizada antes incubando em 70%-75% de etanol por 20 min) no meio antes de solidificar (Figura 1B).
NOTA: Esta camada de plástico contém quatro orifícios pré-fabricados nos cantos para a colocação de sementes esterilizadas pela superfície. Isso permite que as sementes acessem o meio. Posteriormente, esta camada de separação impede que as folhas toquem no fungo que contém o meio, de modo que os micróbios não possam atacar diretamente as folhas e devem tomar o caminho raiz. Outro buraco está no centro, permitindo o corte do canal de infecção. - Quando o meio se solidificar, corte o ágar com um bisturi através do orifício central pré-fabricado até uma profundidade de cerca de 1,5 cm. Remova o ágar cortado para criar um canal de infecção no qual os esporos fúngicos podem ser adicionados mais tarde.
- Arranhe ligeiramente o meio do ágar com uma ponta de pipeta nos quatro orifícios menores para interromper a pele solidificada (isso permite que as sementes absorvam água do meio ágar a aguado). Coloque as sementes usando uma ponta de pipeta nos orifícios menores.
- Feche o copo de plástico com um segundo copo de plástico invertido e vedação com fita adesiva permeável a ar. A fita deve permitir a troca de gás.
- Após estratificação por 3 dias na escuridão a 4 °C, incubar os sistemas de copos sob luz de 12h / 12 h de escuridão (Arabidopsis, estupro da oleosa) ou 16h de luz / 8h de condição de escuridão (tomate) em câmaras de crescimento a uma temperatura constante de 22 °C e 60% de umidade.
- Seguir a idade recomendada das plantas para a inoculação: 21 dias para arabidopsis; 5-7 dias por estupro de oleosa; 12 dias para tomate.
- Plantlets inoculados com Verticillium adicionando 1 mL de solução conidia (concentração recomendada: 4 x 105 esporos/mL) no canal de infecção. Para preparar amostras de controle, adicione 1 mL de solução simulada sem esporos (meio 1/4 MS sem germes) no canal.
- Realize as análises nos pontos de tempo preferidos após a inoculação dependendo da questão da pesquisa (consulte as legendas de figura para os pontos de tempo exatos utilizados aqui). A seguir estão algumas sugestões.
- Tire fotos das plantas com uma câmera digital de cima mantendo a distância igual para cada foto. Quantifique a área da folha (por exemplo, com ImageJ22 ou BlattFlaeche17,19; use o comprimento dos copos para definir a escala) e compare grupos infectados e de controle. Categorizar o desenvolvimento de sintomas da doença (por exemplo, folhas menores, mais amareladas ou necróticas).
NOTA: Se houver alguma haste em Arabidopsis, remova-as para obter melhores fotos das rosetas. - Remova as raízes e defina a biomassa (peso fresco) dos brotos de amostras infectadas e controle por pesagem. Determine o peso fresco relativo19.
- Colete as amostras para análises moleculares da seguinte forma.
- Arabidopsis: Remova hastes se houver alguma. Corte as rosetas na base das raízes. Certifique-se de excluir todo o material raiz da amostra e colher rosas inteiras. Combine 4-5 rosetas de diferentes plantas em uma amostra e congele o material da folha em nitrogênio líquido.
- Retire as raízes cuidadosamente para fora do meio com fórceps, pressione e coloque-as com uma toalha de papel para remover restos de ágar, e combine 4-5 raízes de diferentes plantas em uma amostra. Congele imediatamente em nitrogênio líquido.
- Estupro/tomate obênto: Corte segmentos de caule do hipocotílico (por exemplo, 1 cm de comprimento; sempre tome a mesma região de caule). Misture o material de 4-5 plantas em cada amostra e congele em nitrogênio líquido.
- Triture as amostras em nitrogênio líquido. Extrair DNA total de 100 mg da folha ou material-tronco para determinar via qPCR a quantidade de DNA fúngico em relação ao DNA vegetal (ver19).
- Triture as amostras em nitrogênio líquido, pegue 100 mg de material vegetal e extraia RNA total. Realize qRT-PCR para determinar a expressão de genes vegetais (ou genes fúngicos) após a infestação (ver19).
- Tire fotos das plantas com uma câmera digital de cima mantendo a distância igual para cada foto. Quantifique a área da folha (por exemplo, com ImageJ22 ou BlattFlaeche17,19; use o comprimento dos copos para definir a escala) e compare grupos infectados e de controle. Categorizar o desenvolvimento de sintomas da doença (por exemplo, folhas menores, mais amareladas ou necróticas).
6. Um sistema de inoculação à base de solo em vasos
- Misture completamente o solo e a areia em uma relação volumosa de 3:1 (solo:areia) para facilitar a lavagem do substrato das raízes. Despeje a mistura em um saco de autoclave. Se a mistura estiver muito seca, adicione uma quantidade adequada de água e misture-a no substrato. Vapor a 80 °C por 20 min em uma autoclave para minimizar as contaminações microbianas.
NOTA: Evite o aquecimento a mais de 80 °C, pois isso pode afetar nutrientes orgânicos do solo. - Encha os potes com a mistura de areia do solo e transfira-os para bandejas. Adicione água nas bandejas cerca de 1/3 da altura de uma panela, de modo que a mistura de areia do solo fique completamente encharcada com água. Além disso, pulverizar a água do substrato com uma garrafa de spray para garantir condições de partida molhadas.
- Porca 3-4 sementes em cada pote (Figura 1C) garantindo que as sementes tenham distância suficiente uma da outra. Mantenha-os por 3 dias na escuridão a 4 °C para estratificação para sincronizar a germinação.
NOTA: Pré-cultivar um excesso de plantas, o que permite uma seleção de plantas de tamanho semelhante para os experimentos de inoculação e reduz os desvios devido a diferenças individuais. - Deixe as mudas crescerem em condições de longo dia (16h de luz / 8h de escuridão; temperatura constante de 22 °C; 60% de umidade) com rega regular.
- Siga a idade recomendada das plantas para inoculação: 21 dias para arabidopse, 7 dias para estupro de oleosa e 10 dias para tomate. Escolha plantas de tamanho semelhante para realizar a "inoculação de mergulho raiz"15,17,23,24. Tire o solo das panelas e escava cuidadosamente as raízes.
- Lave suavemente apenas as raízes em um recipiente de água e mantenha as rosetas fora da água. Incubar as raízes lavadas por 60 min em uma placa de Petri contendo a solução de esporos verticillium (concentração recomendada: 2 x 106 esporos/mL). Para o grupo de controle não infectado, incubar as raízes por 60 minutos na solução simulada sem esporos (meio de 1/4 MS sem germes).
- Prepare novas panelas com solo úmido e esterilizado a vapor (80 °C por 20 minutos) sem areia. Use uma ponta de pipeta para fazer um buraco no centro do solo em cada panela.
- Coloque diretamente as raízes no orifício (transfira apenas uma planta por vaso). Depois de inserir as raízes, certifique-se de reabastecer os orifícios cuidadosamente com o solo. Evite pressionar o solo, caso contrário, o repotting pode causar sintomas de estresse, como folhas roxas.
- Cultivar grupos infectados e de controle em condições de longo dia (16h de luz / 8h de escuridão; uma temperatura constante de 22 °C; 60% de umidade) com rega regular.
- Realize as análises nos pontos de tempo preferidos após a inoculação dependendo da questão da pesquisa (consulte as legendas de figura para os pontos de tempo exatos utilizados aqui). A seguir estão algumas sugestões.
- Tire fotos das plantas com uma câmera digital de cima, mantendo a distância igual para cada foto. Quantifique a área da folha (por exemplo, com ImageJ22 ou BlattFlaeche17,19; use o diâmetro dos potes para definir a escala) e compare grupos infectados e de controle. Categorizar o desenvolvimento de sintomas da doença (por exemplo, folhas menores, mais amareladas ou necróticas)13.
NOTA: A remoção de hastes de Arabidopsis facilita tirar fotos das rosetas. - Remova as raízes e defina a biomassa (peso fresco) dos brotos de amostras infectadas e controle por pesagem. Determine o peso fresco relativo19.
- Alternativamente, meça a altura da planta ou categorize o crescimento fúngico dos segmentos de caule para avaliar a gravidade da doença13.
- Coletar amostras para análises moleculares da seguinte forma.
- Arabidopsis: Remova as hastes. Corte as rosetas na coroa raiz. Combine 4-5 rosetas de diferentes plantas em uma amostra. Congele o material da folha em nitrogênio líquido.
NOTA: No caso das raízes, é difícil limpá-las suficientemente do solo sem reprogramar a expressão genética através da lavagem. - Estupro/tomate obênto: Corte segmentos de caule do hipocotílico (por exemplo, 1 cm de comprimento; sempre tome a mesma região de caule). Misture o material de 4-5 plantas em uma amostra e congele-o em nitrogênio líquido.
- Triture as amostras em nitrogênio líquido. Extrair DNA total de 100 mg de folha ou material-tronco para determinar via qPCR a quantidade de DNA fúngico em relação ao DNA vegetal (ver19).
- Triture as amostras em nitrogênio líquido, pegue 100 mg de material vegetal e extraia RNA total. Realize qRT-PCR para determinar a expressão de genes vegetais (ou genes fúngicos) após a infestação (ver19).
- Tire fotos das plantas com uma câmera digital de cima, mantendo a distância igual para cada foto. Quantifique a área da folha (por exemplo, com ImageJ22 ou BlattFlaeche17,19; use o diâmetro dos potes para definir a escala) e compare grupos infectados e de controle. Categorizar o desenvolvimento de sintomas da doença (por exemplo, folhas menores, mais amareladas ou necróticas)13.
7. Analisando os dados
- Calcule o desvio médio e padrão (± SD) com base nas réplicas biológicas.
- Calcule os valores relativos dividindo todos os resultados do grupo infectado pelo resultado do controle. Mostre a média como, por exemplo, "relativa a zombaria" ou "relativa ao tipo selvagem".
- Determinar a significância estatística entre os grupos.

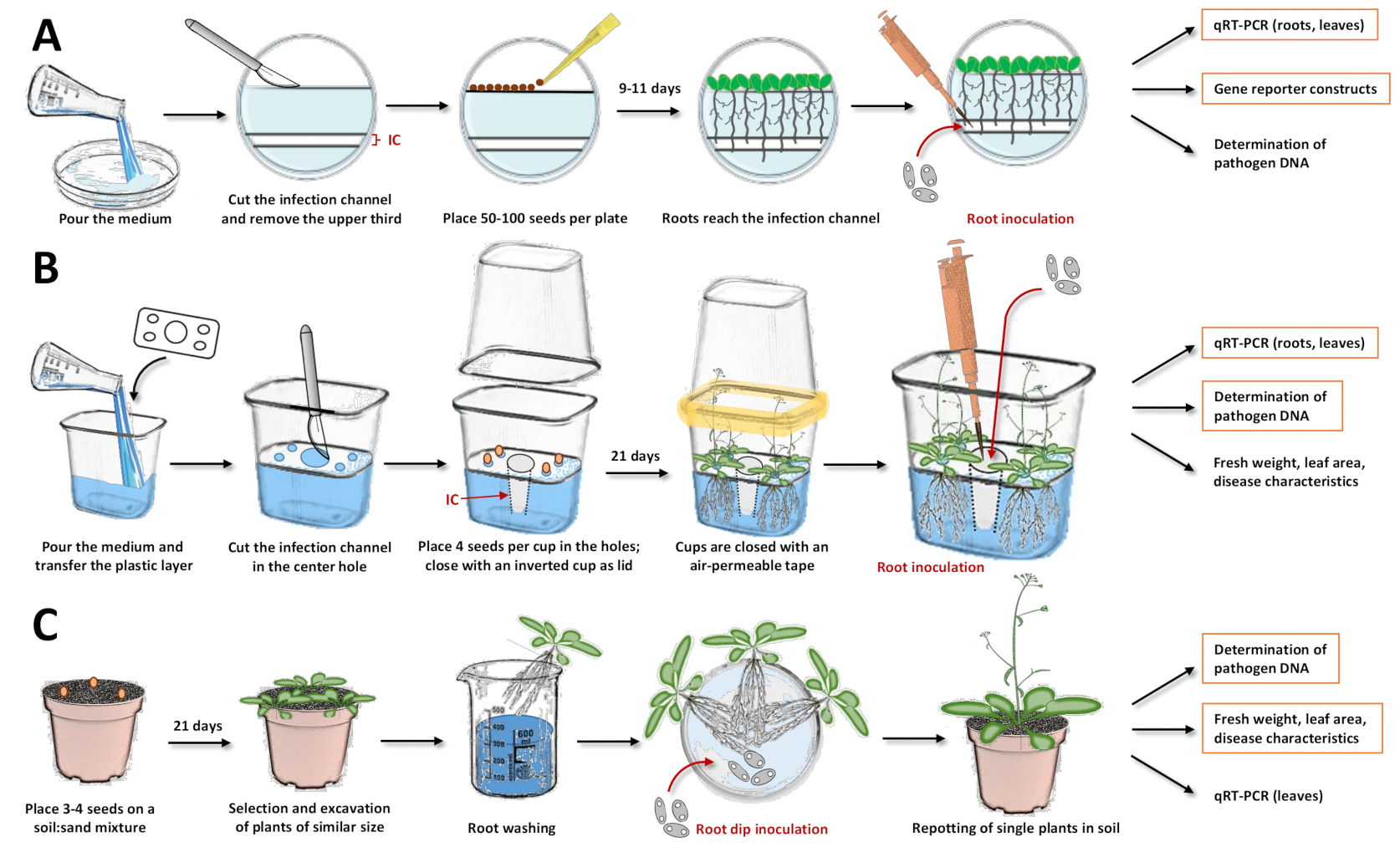
Figura 1: Compilação dos três sistemas de inoculação e etapas individuais nos protocolos. Esses números ilustram os sistemas com a planta modelo Arabidopsis thaliana. Para outras espécies de plantas, o tempo deve ser ajustado. Destaque para caixas laranjas, para as quais as análises subsequentes são mais recomendadas com o respectivo sistema. (A) Para o sistema de inoculação em placas de Petri17, despeje o meio e deixe-o solidificar. Mantenha os pratos na geladeira durante a noite. Em seguida, corte e remova o terço superior, bem como o canal de infecção (IC) com um bisturi (áreas brancas na ilustração foram removidas do ágar, enquanto as áreas azuladas representam o ágar). Coloque as sementes na superfície cortada e feche as placas de Petri. Após a estratificação, coloque as placas verticalmente e deixe as plantas crescerem. Uma vez que a maioria das raízes tenha atingido o canal de infecção, adicione a solução de esporos com uma pipeta diretamente no canal. Certifique-se de que a solução está distribuída uniformemente. Feche as placas de Petri e incuba-as verticalmente em uma câmara de crescimento. As abordagens que podem seguir são análises expressionais com transcrição reversa quantitativa PCR (qRT-PCR), microscopia com linhas de repórteres e quantificação do DNA microbiano. (B) Para o sistema de inoculação em copos plásticos19, despeje o meio e transfira a camada plástica que separa os orifícios pré-fabricados (quatro pequenos orifícios nos cantos para colocação das sementes e um grande buraco no centro para o canal de infecção). Deixe o meio solidificar. Corte e remova o meio ágar no orifício central com um bisturi para obter o canal de infecção (IC). Coce o meio nos orifícios menores e transfira as sementes. Feche o copo com um copo invertido e vedação com fita permeável de ar (simbolizada em amarelo). Deixe as plantas crescerem. Para a inoculação, adicione a solução de esporo com uma pipeta diretamente no canal de infecção. Feche o sistema e continue o cultivo na câmara de crescimento. As abordagens que podem seguir são análise expressional com qRT-PCR, quantificação do DNA microbiano e determinação de peso fresco, área da folha ou outras características da doença. (C) "Inoculação de mergulho raiz"15,17,23,24: para o sistema de inoculação à base de solo, encha os potes com uma mistura de solo:areia. Transfira as sementes e deixe as mudas crescerem. Escavar plantas de tamanho semelhante e lavar as raízes na água. Coloque as raízes lavadas em uma placa de Petri segurando a solução com os esporos. Após a incubação, insira plantas únicas em vasos com solo. As abordagens que podem seguir são análise expressional em folhas com qRT-PCR, quantificação do DNA microbiano e determinação de peso fresco, área da folha ou outras características da doença. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Resultados
Seguindo o protocolo, as plantas foram cultivadas e inoculadas com V. longisporum (cepa Vl4325) ou V. dahliae (isolar JR218). Vários cenários foram projetados para provar a eficácia e destacar algumas capacidades dos protocolos determinados. Resultados representativos são mostrados.
A indução expressional de genes envolvidos na biossíntese indol-glucosinolato antimicrobiana (IG) é um indicador confiável para a ...
Discussão
Devido às enormes perdas de rendimento causadas pelos fitopatógenos transportados pelo solo1, é necessária uma melhoria das estratégias agrícolas ou variedades de culturas. A visão limitada da patogênese das doenças transmitidas pelo solo dificulta o desenvolvimento de plantas mais resistentes. Os mecanismos subjacentes precisam ser explorados, para os quais uma plataforma metodológica robusta é necessária. Os procedimentos de inoculação relatados mostraram que eventos multifatoriais...
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Os autores reconhecem Tim Iven e Jaqueline Komorek por trabalhos anteriores sobre esses métodos, o grupo de Wolfgang Dröge-Laser (Departamento de Biologia Farmacêutica da Universidade de Würzburg, Alemanha) por fornecer os equipamentos e os recursos necessários para este trabalho, e Wolfgang Dröge-Laser, bem como Philipp Kreisz (ambos da Universidade de Würzburg) para a revisão crítica do manuscrito. Este estudo foi apoiado pela "Deutsche Forschungsgemeinschaft" (DFG, DR273/15-1,2).
Materiais
| Name | Company | Catalog Number | Comments |
| Agar (Gelrite) | Carl Roth | Nr. 0039 | all systems described require Gelrite |
| Arabidopsis thaliana wild-type | NASC stock | Col-0 (N1092) | |
| Autoclave | Systec | VE-100 | |
| BlattFlaeche | Datinf GmbH | BlattFlaeche | software to determine leaf areas |
| Brassica napus wild-type | see Floerl et al., 2008 | rapid-cycling rape | genome ACaacc |
| Cefotaxime sodium | Duchefa | C0111 | |
| Chicanery flask 500 mL | Duran Group / neoLab | E-1090 | Erlenmeyer flask with four baffles |
| Collection tubes 50 mL | Sarstedt | 62.547.254 | 114 x 28 mm |
| Czapek Dextrose Broth medium | Duchefa | C1714 | |
| Digital camera | Nikon | D3100 18-55 VR | |
| Exsiccator (Desiccator ) | Duran Group | 200 DN, 5.8 L | Seal with lid to hold chlorine gas |
| Fluorescence Microscope | Leica | Leica TCS SP5 II | |
| HCl | Carl Roth | P074.3 | |
| KNO3 | Carl Roth | P021.1 | ≥ 99 % |
| KOH | Carl Roth | 6751 | |
| Leukopor | BSN medical GmbH | 2454-00 AP | non-woven tape 2.5 cm x 9.2 m |
| MES (2-(N-morpholino)ethanesulfonic acid) | Carl Roth | 4256.2 | Pufferan ≥ 99 % |
| MgSO4 | Carl Roth | T888.1 | Magnesiumsulfate-Heptahydrate |
| Murashige & Skoog medium (MS) | Duchefa | M0222 | MS including vitamins |
| NaClO | Carl Roth | 9062.1 | |
| Percival growth chambers | CLF Plant Climatics GmbH | AR-66L2 | |
| Petri-dishes | Sarstedt | 82.1473.001 | size ØxH: 92 × 16 mm |
| Plastic cups (500 mL, transparent) | Pro-pac, salad boxx | 5070 | size: 108 × 81 × 102 mm |
| Pleated cellulose filter | Hartenstein | FF12 | particle retention level 8–12 μm |
| poly klima growth chamber | poly klima GmbH | PK 520 WLED | |
| Potato Dextrose Broth medium | SIGMA Aldrich | P6685 | for microbiology |
| Pots | Pöppelmann GmbH | TO 7 D or TO 9,5 D | Ø 7 cm resp. Ø 9.5 cm |
| PromMYB51::YFP | see Poncini et al., 2017 | MYB51 reporter line | YFP (i.e. 3xmVenus with NLS) |
| Reaction tubes 2 mL | Sarstedt | 72.695.400 | PCR Performance tested |
| Rotary (orbital) shaker | Edmund Bühler | SM 30 C control | |
| Sand (bird sand) | Pet Bistro, Müller Holding | 786157 | |
| Soil | Einheitserde spezial | SP Pikier (SP ED 63 P) | |
| Solanum lycopersicum wild-type | see Chavarro-Carrero et al., 2021 | Type: Moneymaker | |
| Thoma cell counting chamber | Marienfeld | 642710 | depth 0.020 mm; 0.0025 mm2 |
| Ultrapure water (Milli-Q purified water) | MERK | IQ 7003/7005 | water obtained after purification |
| Verticillium dahliae | see Reusche et al., 2014 | isolate JR2 | |
| Verticillium longisporum | Zeise and von Tiedemann, 2002 | strain Vl43 |
Referências
- Mendes, R., Garbeva, P., Raaijmakers, J. M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiology Review. 37 (5), 634-663 (2013).
- Yadeta, K. A., Thomma, B. P. H. J. The xylem as battleground for plant hosts and vascular wilt pathogens. Frontiers in Plant Science. 4, 97 (2013).
- Delgado-Baquerizo, M., et al. The proportion of soil-borne pathogens increases with warming at the global scale. Nature Climate Change. 10 (6), 550-554 (2020).
- Berendsen, R. L., et al. Disease-induced assemblage of a plant-beneficial bacterial consortium. The ISME Journal. 12 (6), 1496-1507 (2018).
- Yuan, J., et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome. 6 (1), 156 (2018).
- Liu, H., et al. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytologist. 229 (5), 2873-2885 (2021).
- Wang, H., Liu, R., You, M. P., Barbetti, M. J., Chen, Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): role of bacterial diversity. Microorganisms. 9 (9), 1988 (2021).
- Inderbitzin, P., Subbarao, K. V. Verticillium systematics and evolution: how confusion impedes Verticillium wilt management and how to resolve it. Phytopathology. 104 (6), 564-574 (2014).
- Eynck, C., Koopmann, B., Grunewaldt-Stoecker, G., Karlowsky, P., von Tiedemann, A. Differential interactions of Verticillium longisporum und V. dahliae with Brassica napus with molecular and histological techniques. European Journal of Plant Pathology. 118 (3), 259-274 (2007).
- Floerl, S., et al. Defence reactions in the apoplastic proteome of oilseed rape (Brassica napus var. napus) attenuate Verticillium longisporum growth but not disease symptoms. BMC Plant Biology. 8, 129 (2008).
- Leonard, M., et al. Verticillium longisporum elicits media-dependent secretome responses with capacity to distinguish between plant-related environments. Frontiers in Microbiology. 11, 1876 (2020).
- Depotter, J. R. L., et al. Verticillium longisporum, the invisible threat to oilseed rape and other brassicaceous plant hosts. Molecular Plant Pathology. 17 (7), 1004-1016 (2016).
- Fröschel, C., et al. A gain-of-function screen reveals redundant ERF transcription factors providing opportunities for resistance breeding toward the vascular fungal pathogen Verticillium longisporum. Molecular Plant-Microbe Interactions. 32 (9), 1095-1109 (2019).
- Zhou, L., Hu, Q., Johansson, A., Dixelius, C. Verticillium longisporum and V. dahliae: infection and disease in Brassica napus. Plant Pathology. 55 (1), 137-144 (2006).
- Ralhan, A., et al. The vascular pathogen Verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots. Plant Physiology. 159 (3), 1192-1203 (2012).
- Reusche, M., et al. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium longisporum. Molecular Plant-Microbe Interactions. 26 (8), 850-860 (2013).
- Iven, T., et al. Transcriptional activation and production of tryptophan-derived secondary metabolites in Arabidopsis roots contributes to the defense against the fungal vascular pathogen Verticillium longisporum. Molecular Plant. 5 (6), 1389-1402 (2012).
- Reusche, M., et al. Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana. Environmental and Experimental Botany. 108, 23-37 (2014).
- Fröschel, C. In-depth evaluation of root infection systems using the vascular fungus Verticillium longisporum as soil-borne model pathogen. Plant Methods. 17 (1), 57 (2021).
- Karapapa, V. K., Bainbridge, B. W., Heale, J. B. Morphological and molecular characterization of Verticillium longisporum comb, nov., pathogenic to oilseed rape. Mycological Research. 101 (11), 1281-1294 (1997).
- Poncini, L., et al. In roots of Arabidopsis thaliana, the damage-associated molecular pattern AtPep1 is a stronger elicitor of immune signalling than flg22 or the chitin heptamer. PLoS One. 12 (10), 1-21 (2017).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Fradin, E. F., et al. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiology. 150 (1), 320-332 (2009).
- Singh, S., et al. The plant host Brassica napus induces in the pathogen Verticillium longisporum the expression of functional catalase peroxidase which is required for the late phase of disease. Molecular Plant-Microbe Interactions. 25 (4), 569-581 (2012).
- Zeise, K., von Tiedemann, A. Application of RAPD-PCR for virulence type analysis within Verticillium dahliae and Verticillium longisporum. Journal of Phytopathology. 150 (10), 557-563 (2002).
- Fröschel, C., et al. Plant roots employ cell-layer-specific programs to respond to pathogenic and beneficial microbes. Cell Host & Microbe. 29 (2), 299-310 (2021).
- Gigolashvili, T., et al. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. The Plant Journal. 50 (5), 886-901 (2007).
- Back, M. A., Haydock, P. P. J., Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathology. 51 (6), 683-697 (2002).
- Behrens, F. H., et al. Suppression of abscisic acid biosynthesis at the early infection stage of Verticillium longisporum in oilseed rape (Brassica napus). Molecular Plant Pathology. 20 (12), 1645-1661 (2019).
- Vorholt, J. A., Vogel, C., Carlström, C. I., Müller, D. B. Establishing causality: opportunities of synthetic communities for plant microbiome research. Cell Host & Microbe. 22 (2), 142-155 (2017).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados