Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Bitki Köklerini Toprak Kaynaklı Mikroorganizmalarla Enfekte Etmek için Aşılama Stratejileri
Bu Makalede
Özet
Bu protokol, bitki köklerini toprak kaynaklı mikroplarla aşılamak için stratejilerin ayrıntılı bir özetini sunar. Verticillium longisporum ve Verticillium dahliae mantarları için örneklendirilen üç farklı kök enfeksiyon sistemi tanımlanmıştır. Potansiyel uygulamalar ve olası aşağı akış analizleri vurgulanır ve her sistem için avantajlar veya dezavantajlar tartışılır.
Özet
Rizosfer, bitki köklerinin sürekli olarak zorlandığı oldukça karmaşık bir mikrobiyal topluluğu barındırır. Kökler çok çeşitli mikroorganizmalarla yakın temas halindedir, ancak toprak kaynaklı etkileşimler üzerine yapılan çalışmalar hala yer üstü organlarında yapılanların gerisindedir. Model bitkileri model kök patojenlerle enfekte etmek için bazı aşılama stratejileri literatürde tanımlanmış olsa da, kapsamlı bir metodolojik genel bakış elde etmek zor olmaya devam etmektedir. Bu sorunu çözmek için, kök-mikrop etkileşimlerinin biyolojisi hakkında fikir edinmek için uygulanabilecek üç farklı kök aşılama sistemi tam olarak tanımlanmıştır. Örnek olarak, Verticillium türleri (yani, V. longisporum ve V. dahliae) kök istilacı model patojenler olarak kullanılmıştır. Bununla birlikte, yöntemler hem patojenik hem de faydalı olan diğer kök kolonize edici mikroplara kolayca uyarlanabilir. Bitki ksilemini kolonize ederek, Verticillium spp. gibi vasküler toprak kaynaklı mantarlar benzersiz bir yaşam tarzı sergiler. Kök istilasından sonra, akropetal olarak ksilem damarları yoluyla yayılırlar, sürgüne ulaşırlar ve hastalık semptomları ortaya çıkarırlar. Model konakçı olarak üç temsili bitki türü seçildi: Arabidopsis thaliana, ekonomik açıdan önemli yağlı tohumlu kolza (Brassica napus) ve domates (Solanum lycopersicum). Adım adım protokoller verilmiştir. Patojenite tahlillerinin temsili sonuçları, belirteç genlerinin transkripsiyonel analizleri ve muhabir yapıları tarafından bağımsız doğrulamalar gösterilmiştir. Ayrıca, her bir aşılama sisteminin avantajları ve dezavantajları kapsamlı bir şekilde tartışılmaktadır. Bu kanıtlanmış protokoller, kök-mikrop etkileşimleri hakkındaki araştırma soruları için yaklaşımlar sağlamada yardımcı olabilir. Bitkilerin topraktaki mikroplarla nasıl başa çıktığını bilmek, tarımı iyileştirmek için yeni stratejiler geliştirmek için çok önemlidir.
Giriş
Doğal topraklarda, nötr, zararlı veya bitkilere faydalı olabilecek şaşırtıcı sayıda mikropyaşar 1. Birçok bitki patojeni toprak kaynaklıdır, kökleri çevreler ve yeraltı organına saldırır. Bu mikroorganizmalar çok çeşitli kladlara aittir: mantarlar, oomisetler, bakteriler, nematodlar, böcekler ve bazı virüsler 1,2. Çevresel koşullar enfeksiyonu desteklediğinde, duyarlı bitkiler hastalıklı hale gelecek ve mahsul verimi düşecektir. Küresel ısınma ve aşırı hava koşulları gibi iklim değişikliğinin etkileri, toprak kaynaklı bitki patojenlerinin oranını artıracaktır3. Bu nedenle, bu yıkıcı mikropları ve bunların gıda ve yem üretimi üzerindeki etkilerini, aynı zamanda doğal ekosistemler üzerindeki etkilerini incelemek giderek daha önemli hale gelecektir. Ek olarak, toprakta köklerle sıkı bir şekilde etkileşime giren ve bitki büyümesini, gelişimini ve bağışıklığını teşvik eden mikrobiyal karşılıkçılar vardır. Patojenlerle karşı karşıya kaldıklarında, bitkiler rizosferde patojenleri baskılayarak konakçı sağkalımını destekleyebilecek spesifik rakipleri aktif olarak işe alabilir 4,5,6,7. Bununla birlikte, yararlı kök-mikrop etkileşimlerinde yer alan mekanik detaylar ve yollar genellikle hala bilinmemektedir6.
Bu nedenle, kök-mikrop etkileşimlerinin genel anlayışını genişletmek esastır. Model çalışmaları yapmak ve bulguları tarımsal uygulamalara aktarmak için köklerin toprak kaynaklı mikroorganizmalarla aşılanması için güvenilir yöntemler gereklidir. Topraktaki faydalı etkileşimler, örneğin, Serendipita indica (eski adıyla Piriformospora indica), azot sabitleyici Rhizobium spp. veya mikorizal mantarlarla incelenirken, bilinen toprak kaynaklı bitki patojenleri arasında Ralstonia solanacearum, Phytophthora spp., Fusarium spp. ve Verticillium spp.1 bulunur. Son ikisi, küresel olarak dağılmış ve vasküler hastalıklara neden olan mantar cinsleridir2. Verticillium spp. (Ascomycota) yüzlerce bitki türünü enfekte edebilir - otsu yıllıklar, odunsu çok yıllık bitkiler ve birçok mahsul bitkisi de dahil olmak üzere büyük ölçüde dikotiledonlar 2,8. Verticillium'un hifleri köke girer ve ksilem damarlarınıkolonize etmek için merkezi silindire doğru hem hücreler arası hem de hücre içi olarak büyür 2,9. Bu damarlarda, mantar yaşam döngüsünün çoğunda kalır. Ksilem özü besin açısından fakir olduğundan ve bitki savunma bileşikleri taşıdığından, mantar bu eşsiz ortama uyum sağlamalıdır. Bu, patojenin konakçısı10,11'de hayatta kalmasını sağlayan kolonizasyonla ilgili proteinlerin salgılanmasıyla gerçekleştirilir. Kök vaskülatürüne ulaştıktan sonra, mantar ksilem damarları içinde akroleptal olarak yeşilliklere yayılabilir ve bu da konakçının sistemik kolonizasyonuna yol açar 9,12. Bu noktada bitki büyümesinde olumsuz etkilenmektedir 9,10,13. Örneğin, bodurluk ve sarı yaprakların yanı sıra erken yaşlanma13,14,15,16 meydana gelir.
Bu cinsin bir üyesi, tarımsal olarak önemli yağlı tohumlu kolza, karnabahar ve model bitki Arabidopsis thaliana12 gibi sütyen konakçılara oldukça adapte olmuş Verticillium longisporum'dur. Birkaç çalışma, toprak kaynaklı vasküler hastalıklar ve ortaya çıkan kök savunma yanıtları13,15,16,17 hakkında kapsamlı bilgiler edinmek için V. longisporum ve A. thaliana'yı birleştirdi. Basit duyarlılık testi, V. longisporum / A. thaliana model sistemi kullanılarak gerçekleştirilebilir ve her iki organizma için de köklü genetik kaynaklar mevcuttur. V. longisporum ile yakından ilişkili olan patojen Verticillium dahliae'dir. Her iki mantar türü de benzer bir vasküler yaşam tarzı ve istila süreci gerçekleştirse de, köklerden yapraklara yayılma etkinlikleri ve A. thaliana'da ortaya çıkan hastalık semptomları farklıdır: V. longisporum genellikle erken yaşlanmayı indüklerken, V. dahliae enfeksiyonu solgunluğa neden olur18. Son zamanlarda, metodolojik bir özet, A. thaliana'yı V. longisporum veya V. dahliae ile enfekte etmek için farklı kök aşılama stratejileri sunmuş ve deneysel kurulumların planlanmasına yardımcı olmuştur19. Tarlada, V. longisporum zaman zaman yağlı tohumlu kolzaüretiminde önemli hasara neden olurken, V. dahliae asma, patates ve domates8 gibi çeşitli ekili türlerden oluşan çok geniş bir konakçı yelpazesine sahiptir. Bu, her iki patojeni de incelemek için ekonomik olarak ilginç modeller haline getirir.
Bu nedenle, aşağıdaki protokoller, kök aşılamaları için olası yaklaşımları örneklemek için hem V. longisporum hem de V. dahliae'yi model kök patojenleri olarak kullanmaktadır. Arabidopsis (Arabidopsis thaliana), yağlı tohumlu kolza (Brassica napus) ve domates (Solanum lycopersicum) model konakçı olarak seçildi. Metodolojilerin ayrıntılı açıklamaları aşağıdaki metinde ve beraberindeki videoda bulunabilir. Her aşılama sistemi için avantaj ve dezavantajlar tartışılmıştır. Birlikte ele alındığında, bu protokol koleksiyonu, kök-mikrop etkileşimleri bağlamında belirli araştırma soruları için uygun bir yöntemin belirlenmesine yardımcı olabilir.
Protokol
1. Mantar kültürleri ve bitki aşılama sistemleri için ortam
- Sıvı Patates Dekstroz Suyu (PDB): Isıya dayanıklı bir şişede ultra saf suda 21 g / L PDB hazırlayın.
- Sıvı Czapek Dekstroz Suyu (CDB): Isıya dayanıklı bir şişede ultra saf suda 42 g / L CDB hazırlayın.
- Petri kabı aşılama sistemi için ortam: Ultra saf suda 1,5 g / L Murashige ve Skoog ortamı (MS) ve 8 g / L agar ile ısıya dayanıklı bir şişe hazırlayın.
NOT: Bu ortamda şekerden kaçının, çünkü aşılamadan sonra aşırı mantar büyümesine yol açacaktır. - Plastik fincan bazlı aşılama sistemi için ortam: Ultra saf suda 4.4 g / L MS, 0.2 g / L MgSO4, 1 g / L KNO3, 0.5 g / L 2-(N-morfolino) etansülfonik asit (MES) ve 6.0 g / L agar ile ısıya dayanıklı bir şişe hazırlayın ve pH'ı 5 M KOH ile 5.7'ye ayarlayın.
NOT: Bu ortamda şekerden kaçının, çünkü aşılamadan sonra aşırı mantar büyümesine yol açacaktır. - 1/4 MS ortamı: Ultra saf suda 1,2 g/L MS hazırlayın.
- Yukarıdaki tüm çözeltileri sterilize etmek için otoklav kullanın. Cam şişeleri sepete koyun, kapağı kapatın ve 121 ° C ve 98.9 kPa'da 15 dakika sterilize edin.
2. Bitki tohumlarının yüzeyinin sterilize edilmesi
NOT: Ekimden önce Arabidopsis, yağlı tohum kolza ve domates tohumlarının yüzeyini sterilize etmek için her zaman aşağıdaki protokolü kullanın.
- Tohumları 2 mL'lik bir reaksiyon tüpüne aktarın. Tüpü 5,8 L'lik bir iç kapasiteye sahip bir eksizatöre yerleştirin.
- 100 mL% 12 sulu sodyum hipoklorite (NaClO) 6 mL% 33 hidroklorik asit (HCl) ekleyerek eksikatörde klor gazı üretin.
- Hemen eksikatörün kapağını kapatın ve tohumları gazda 3 saat boyunca inkübe edin.
3. İnokulumun Verticillium sporları (aseksüel türevli konidia) ile hazırlanması
NOT: V. dahliae (JR2 suşu) ile V. longisporum (suş Vl43)17,18,19 ile aynı şekilde yetiştirilir. Tüm ekipmanın ve ortamın mikropsuz olduğundan ve inokulumu aksenik tutmak için tüm adımların laminer bir akış başlığında gerçekleştirildiğinden emin olun.
- 500 mL'lik bir chicanery şişesine 150 mL sıvı PDB (adım 1.1) doldurun ve ortamı 500 mg / L sefotaksim ile tamamlayın.
- Verticillium conida'yı gliserol stok deposundan PDB ortamına ekleyin. Şişeyi steril bir köpük tıpa ile kapatın.
- Kültürü, sürekli, yatay sallama (döner çalkalayıcı; 60 rpm) altında oda sıcaklığında (RT) karanlık bir kutuda 7-10 gün boyunca inkübe edin. Bu, küçük, beyaz misel küreleri ile sonuçlanır.
- PDB supernatant'ı dikkatlice çıkarın ve atın. Miselinin çoğu şişede kalmalıdır.
- Chicanery şişesindeki miselya üzerine 100 mL sıvı CDB (adım 1.2) ekleyin ve ortamı 500 mg / L sefotaksim ile tamamlayın.
- Sporülasyonu indüklemek için RT'deki karanlık bir kutuda sürekli, yatay sallama (döner çalkalayıcı, 60 rpm) altında 4-5 gün daha inkübe edin. Süpernatant, konidia serbest bırakıldıkça sarımsı-grimsi hale gelecektir.
- Konidya içeren sıvının bir kısmını (5-10 mL) bir filtre kağıdından (partikül tutma seviyesi 8-12 μm) steril bir 50 mL toplama tüpüne filtreleyin. Bu, sporları miselden ayırır.
- Bir hücre sayma odası ve mikroskop kullanarak spor konsantrasyonunu belirleyin. Aşağıda verilen spor konsantrasyonları elde edilene kadar ultra saf suda mikropsuz 1/4 MS ortamı ile seyreltin.
NOT: Mikroskop altında, V. longisporum'dan gelen konidia çoğunlukla uzun çekilmiş ve 7.1-8.8 μm boyutundayken, V. dahliae conidia daha kısadır (3.5-5.5 μm) ve oldukça küresel20'dir. - Bu taze hasat edilmiş konidiayı inokül olarak kullanın. Deneyleri her zaman dondurulmuş stoklarla değil, taze hasat edilmiş konidia ile yaptığınızdan emin olun, çünkü donma canlı sporların sayısını önemli ölçüde azaltır19.
- Uzun süreli depolama için, sporları -80 ° C'de% 25 gliserol içinde (1 yıla kadar saklanabilir) yüksek konsantrasyonlu spor çözeltisi (yaklaşık 1 x 10 8 spor / mL) olarak dondurun. Sonraki deneyler için, adım 3.2'de PDB ortamını aşılamak için bu gliserol stoklarını kullanın.
4. Petri kaplarına dayalı steril bir in vitro aşılama sistemi
NOT: Petri çanak sistemi17 için, tüm ekipman ve ortamların mikropsuz olduğundan ve tüm adımların laminer bir akış başlığında gerçekleştirildiğinden emin olun.
- Otoklavlamadan sonra, ortamı (bkz. adım 1.3) Petri kaplarına dökün.
- Ortamın sertleşmesinden sonra, Petri bulaşıklarını steril bir plastik torbada yeniden paketleyin ve gece boyunca buzdolabında (4-10 ° C) baş aşağı saklayın. Soğutulmuş bir ortam, sonraki adımlarda ortamın kaymasını önlemeye yardımcı olur.
- Bir enfeksiyon kanalını ve katılaşmış ortamın üst üçte birini bir neşterle kesin ve çıkarın (Şekil 1A). Kesim sırasında agar ortamının altına sıvı veya hava almaktan kaçının; Aksi takdirde, ortam kayacak ve enfeksiyon kanalını kapatacaktır.
- 50-100 yüzey sterilize Arabidopsis tohumunu, kesilmiş üst yüzeye steril bir pipet ucu ile dağıtın. Tohumları, kesilmiş agar yüzeyinin Petri kabının duvarına temas ettiği açıya koyun, böylece kökler ortam ile Petri çanağı duvarı arasında büyüyebilir. Bu daha sonra aşılamayı kolaylaştıracaktır.
- Petri çanaklarını kapatın ve gaz değişimine izin vermek için hava geçirgen yapışkan bantla kapatın.
- Karanlıkta 4 °C'de 2 gün boyunca tabakalaşmadan sonra, plakaları dikey olarak uygun bir rafa yerleştirin ve bitkileri bir büyüme odasında uzun gün koşullarında (16 saat ışık / 8 saat karanlık) 22 ° C ± 1 ° C'de yetiştirin.
- Köklerin çoğunluğu enfeksiyon kanalına ulaştığında (yaklaşık 9-11 günlük fideler), plakaları yatay olarak yerleştirin, açın ve sıvının kanalda eşit olarak dağıldığından emin olmak için doğrudan enfeksiyon kanalına 4 x 10 5 spor / mL konsantrasyonda500 μL taze hasat edilmiş Verticillium conidia ekleyin.
- Benzer şekilde, sporlar yerine 500 μL sahte bir çözelti ekleyerek kontrol plakaları hazırlayın (mikropsuz 1/4 MS ortamı).
- Sıvı ıslanana kadar plakaları yatay olarak birkaç dakika inkübe edin ve plakalar tekrar dikey olarak ayarlandığında sızıntı yapamazsınız. Ardından, kapağı kapatın ve plakaları hava geçirgen yapışkan bantla kapatın.
- Plakaları büyüme odasında dikey olarak inkübe edin. İsteğe bağlı olarak, kökleri ve toprak kaynaklı mantarları koyulaştırmak için kök kısımlarını siyah kağıt kutularla örtün (bkz.19).
- Araştırma sorusuna bağlı olarak aşılamadan sonra analizleri tercih edilen zaman noktalarında gerçekleştirin (burada kullanılan kesin zaman noktaları için şekil açıklamalarına bakın). Aşağıda bazı öneriler verilmiştir.
- Yaprakları köklerden kesin ve her ikisini de ayrı ayrı hasat edin. Köklere kolayca erişmek için agar şeritlerini Petri bulaşıklarından çıkarın ve forseps kullanarak dikkatlice agardan çıkarın. Tüm bitki materyallerini hemen sıvı azotta dondurun.
- Numuneleri sıvı azotta öğütün. Kantitatif bir PCR (qPCR) yoluyla bitki DNA'sına göre mantar DNA'sı miktarını belirlemek için 100 mg yaprak materyalinden toplam DNA'yı çıkarın (bkz.19).
- Numuneleri sıvı azotta öğütün. 100 mg bitki materyali alın ve toplam RNA'yı çıkarın. İstila sırasında bitki genlerinin (veya mantar genlerinin) ekspresyonunu belirlemek için kantitatif ters transkripsiyon PCR (qRT-PCR) uygulayın (bkz.19).
- Kökleri yaralanmayı önlemek için agardan dikkatlice çıkarın ve floresan mikroskop altında inceleyin.
- Bitki raportör hatlarında belirteç genlerinin indüksiyonunu belirleyin (örneğin, lusiferaz, β-glukuronidaz veya floresan muhabirler17,19,21).
- Mantar raporlayıcı çizgileri kullanarak (örneğin, gelişmiş Yeşil Floresan Proteini, Vl-sGFP9'u yapısal olarak ifade eden V. longisporum) veya boyama teknikleriyle (örneğin, 5-bromo-4-kloro-3-indoksil-N-asetil-beta-d-glukozaminid (X-beta-D-Glc-Nac)18) kullanarak kökteki mantar yayılımını görselleştirin.
- Yaprakları köklerden kesin ve her ikisini de ayrı ayrı hasat edin. Köklere kolayca erişmek için agar şeritlerini Petri bulaşıklarından çıkarın ve forseps kullanarak dikkatlice agardan çıkarın. Tüm bitki materyallerini hemen sıvı azotta dondurun.
5. Plastik bardaklarla organize edilmiş steril in vitro aşılama sistemi
NOT: Bu tekniğin ilk açıklamasında belirtildiği gibi19, tüm ekipman ve ortamların mikropsuz olduğundan ve tüm adımların laminer bir akış başlığında gerçekleştirildiğinden emin olun.
- Toplam hacmi 500 mL olan şeffaf plastik kaplar kullanın ve bunları en az 20 dakika boyunca% 70-75 etanol banyosunda sterilize edin. Bardakları laminer akış davlumbazında kurulayın.
- Otoklavlanmış ortamı (bkz. adım 1.4) plastik bardaklara dökün. İsteğe bağlı olarak, bakteriyel kontaminasyonları önlemek için otoklavlanmış ortama sefotaksim (50 mg / L'lik son konsantrasyon) ekleyin. Arabidopsis ile yapılan deneyler için fincan başına 150 mL veya daha büyük bitki türleri (yağlı tohumlu kolza, domates) ile deneyler için daha fazla ortam (fincan başına 250-300 mL) kullanın.
- Katılaşmadan önce ortama plastik bir tabaka (20 dakika boyunca% 70-75% etanol içinde inkübe edilerek sterilize edilmiş) yerleştirin (Şekil 1B).
NOT: Bu plastik tabaka, yüzey sterilize edilmiş tohumların yerleştirilmesi için köşelerde dört prefabrik delik içerir. Bu, tohumların ortama erişmesini sağlar. Daha sonra, bu ayırıcı tabaka yaprakların mantar içeren ortama dokunmasını önler, böylece mikroplar yapraklara doğrudan saldıramaz ve kök yolunu almalıdır. Merkezde başka bir delik daha bulunur ve enfeksiyon kanalının kesilmesini sağlar. - Ortam katılaştığında, agar'ı prefabrik merkez delikten yaklaşık 1,5 cm derinliğe kadar bir neşterle kesin. Mantar sporlarının daha sonra eklenebileceği bir enfeksiyon kanalı oluşturmak için kesilmiş agar'ı çıkarın.
- Katılaşmış cildi kesmek için agar ortamını dört küçük delikte bir pipet ucu ile hafifçe çizin (bu, tohumların sulu agar ortamından su emmesini sağlar). Tohumları bir pipet ucu kullanarak daha küçük deliklere yerleştirin.
- Plastik bardağı ikinci, ters çevrilmiş bir plastik kapla kapatın ve hava geçirgen yapışkan bantla kapatın. Bant gaz değişimine izin vermelidir.
- 4 °C'de karanlıkta 3 gün tabakalaşmadan sonra, fincan sistemlerini 12 saat ışık / 12 saat karanlık (Arabidopsis, yağlı tohum kolzası) veya 16 saat ışık / 8 saat karanlık koşullar (domates) altında büyüme odalarında 22 °C sabit sıcaklıkta ve% 60 nemde inkübe edin.
- Aşılama için önerilen bitki yaşını takip edin: Arabidopsis için 21 gün; Yağlı tohumlu kolza için 5-7 gün; Domates için 12 gün.
- Enfeksiyon kanalına 1 mL konidia çözeltisi (önerilen konsantrasyon: 4 x 105 spor / mL) ekleyerek plantletleri Verticillium ile aşılayın. Kontrol numuneleri hazırlamak için, kanala sporsuz (mikropsuz 1/4 MS ortam) 1 mL sahte çözelti ekleyin.
- Araştırma sorusuna bağlı olarak aşılamadan sonra analizleri tercih edilen zaman noktalarında gerçekleştirin (burada kullanılan kesin zaman noktaları için şekil açıklamalarına bakın). Aşağıda bazı öneriler verilmiştir.
- Her fotoğraf için mesafeyi aynı tutarak yukarıdan bir dijital kamera ile bitkilerin fotoğraflarını çekin. Yaprak alanını ölçün (örneğin, ImageJ22 veya BlattFlaeche17,19 ile; ölçeği ayarlamak için bardakların uzunluğunu kullanın) ve enfekte olmuş ve kontrol gruplarını karşılaştırın. Hastalık semptomlarının gelişimini kategorilere ayırın (örneğin, daha küçük, daha sarımsı veya nekrotik yapraklar).
NOT: Arabidopsis'te herhangi bir sap varsa, rozetlerin daha iyi fotoğraflarını çekmek için bunları çıkarın. - Kökleri çıkarın ve enfekte olmuş örneklerden sürgünlerin biyokütlesini (taze ağırlık) tanımlayın ve tartarak kontrol edin. Göreceli taze ağırlığı belirleyin19.
- Moleküler analizler için örnekleri aşağıdaki gibi toplayın.
- Arabidopsis: Varsa sapları çıkarın. Tabandaki rozetleri köklerden kesin. Tüm kök materyallerini numuneden çıkardığınızdan ve tüm rozetleri topladığınızdan emin olun. Farklı bitkilerden 4-5 rozeti tek bir numunede birleştirin ve yaprak malzemesini sıvı azotta dondurun.
- Kökleri forsepslerle ortamdan dikkatlice çekin, agar kalıntılarını çıkarmak için bir kağıt havluyla bastırın ve darpın ve farklı bitkilerden 4-5 kökü tek bir numunede birleştirin. Hemen sıvı azotta dondurun.
- Yağlı tohumlu kolza / domates: Hipokotilden sap segmentlerini kesin (örneğin, 1 cm uzunluğunda; her zaman aynı kök bölgesini alın). 4-5 bitkiden gelen malzemeyi her numunede birleştirin ve sıvı azotta dondurun.
- Numuneleri sıvı azotta öğütün. Bitki DNA'sına göre mantar DNA'sı miktarını qPCR yoluyla belirlemek için 100 mg yaprak veya kök materyalinden toplam DNA'yı çıkarın (bkz.19).
- Numuneleri sıvı azotta öğütün, 100 mg bitki materyali alın ve toplam RNA'yı çıkarın. İstila üzerine bitki genlerinin (veya mantar genlerinin) ekspresyonunu belirlemek için qRT-PCR uygulayın (bkz.19).
- Her fotoğraf için mesafeyi aynı tutarak yukarıdan bir dijital kamera ile bitkilerin fotoğraflarını çekin. Yaprak alanını ölçün (örneğin, ImageJ22 veya BlattFlaeche17,19 ile; ölçeği ayarlamak için bardakların uzunluğunu kullanın) ve enfekte olmuş ve kontrol gruplarını karşılaştırın. Hastalık semptomlarının gelişimini kategorilere ayırın (örneğin, daha küçük, daha sarımsı veya nekrotik yapraklar).
6. Saksılarda toprak bazlı aşılama sistemi
- Substratın köklerden yıkanmasını kolaylaştırmak için toprak ve kumu 3: 1 (toprak: kum) hacimsel oranında iyice karıştırın. Karışımı bir otoklav torbasına dökün. Karışım çok kuruysa, uygun miktarda su ekleyin ve substrata karıştırın. Mikrobiyal kontaminasyonları en aza indirmek için otoklavda 20 dakika boyunca 80 °C'de buharlayın.
NOT: 80 ° C'nin üzerinde ısıtmaktan kaçının, çünkü bu organik toprak besinlerini etkileyebilir. - Saksıları toprak-kum karışımıyla doldurun ve tepsilere aktarın. Tepsilere bir tencerenin yüksekliğinin yaklaşık 1 / 3'ü kadar su ekleyin, böylece toprak-kum karışımı suyla iyice ıslatılır. Ek olarak, ıslak başlangıç koşullarını sağlamak için alt tabakayı bir sprey şişesi ile su püskürtün.
- Her tencereye 3-4 tohum ekin (Şekil 1C), tohumların birbirlerinden yeterli mesafeye sahip olmasını sağlayın. Çimlenmeyi senkronize etmek için tabakalaşma için 4 ° C'de karanlıkta 3 gün saklayın.
NOT: Aşılama deneyleri için benzer büyüklükteki bitkilerin seçilmesini sağlayan ve bireysel farklılıklardan kaynaklanan sapmaları azaltan fazla bitki yetiştirin. - Fidelerin düzenli sulama ile uzun gün koşullarında (16 saat ışık / 8 saat karanlık; 22 ° C sabit sıcaklık; % 60 nem) büyümesine izin verin.
- Aşılama için önerilen bitki yaşını takip edin: Arabidopsis için 21 gün, yağlı tohumlu kolza için 7 gün ve domates için 10 gün. "Kök daldırma aşılaması" nı gerçekleştirmek için benzer büyüklükteki bitkileri seçin15,17,23,24. Toprağı saksılardan çıkarın ve kökleri dikkatlice kazın.
- Sadece kökleri bir su kabında nazikçe yıkayın ve rozetleri sudan uzak tutun. Yıkanmış kökleri Verticillium spor çözeltisi içeren bir Petri kabında 60 dakika boyunca inkübe edin (önerilen konsantrasyon: 2 x 106 spor / mL). Enfekte olmayan kontrol grubu için, kökleri sporsuz sahte çözeltide 60 dakika boyunca inkübe edin (mikropsuz 1/4 MS ortamı).
- Kumsuz, nemli, buharla sterilize edilmiş toprakla (20 dakika boyunca 80 °C) yeni saksılar hazırlayın. Her tencerede toprağın ortasında bir delik açmak için bir pipet ucu kullanın.
- Kökleri doğrudan deliğe yerleştirin (saksı başına sadece bir bitki aktarın). Kökleri yerleştirdikten sonra, delikleri dikkatlice toprakla doldurduğunuzdan emin olun. Toprağa basmaktan kaçının, aksi takdirde saksılama mor yapraklar gibi stres semptomlarına neden olabilir.
- Enfekte ve kontrol gruplarını uzun gün koşullarında (16 saat ışık / 8 saat karanlık; 22 ° C'lik sabit bir sıcaklık; % 60 nem) düzenli sulama ile yetiştirin.
- Araştırma sorusuna bağlı olarak aşılamadan sonra analizleri tercih edilen zaman noktalarında gerçekleştirin (burada kullanılan kesin zaman noktaları için şekil açıklamalarına bakın). Aşağıda bazı öneriler verilmiştir.
- Yukarıdan bir dijital kamera ile bitkilerin fotoğraflarını çekin, mesafeyi her fotoğraf için aynı tutun. Yaprak alanını ölçün (örneğin, ImageJ22 veya BlattFlaeche17,19 ile; ölçeği ayarlamak için saksıların çapını kullanın) ve enfekte olmuş ve kontrol gruplarını karşılaştırın. Hastalık semptomlarının gelişimini kategorize edin (örneğin, daha küçük, daha sarımsı veya nekrotik yapraklar)13.
NOT: Arabidopsis'in saplarını çıkarmak, rozetlerin fotoğrafını çekmeyi kolaylaştırır. - Kökleri çıkarın ve enfekte olmuş örneklerden sürgünlerin biyokütlesini (taze ağırlık) tanımlayın ve tartarak kontrol edin. Göreceli taze ağırlığı belirleyin19.
- Alternatif olarak, bitki boyunu ölçün veya hastalık şiddetini değerlendirmek için kök segmentlerinden mantar büyümesini kategorize edin13.
- Aşağıdaki gibi moleküler analizler için örnekler toplayın.
- Arabidopsis: Sapları çıkarın. Rozetleri kök tacında kesin. Farklı bitkilerden 4-5 rozeti tek bir numunede birleştirin. Yaprak malzemesini sıvı azotta dondurun.
NOT: Kökler söz konusu olduğunda, yıkama yoluyla gen ekspresyonunu yeniden programlamadan onları topraktan yeterince temizlemek zordur. - Yağlı tohumlu kolza / domates: Hipokotilden sap segmentlerini kesin (örneğin, 1 cm uzunluğunda; her zaman aynı kök bölgesini alın). 4-5 bitkiden gelen malzemeyi tek bir numunede birleştirin ve sıvı azotta dondurun.
- Numuneleri sıvı azotta öğütün. Bitki DNA'sına göre mantar DNA'sı miktarını qPCR yoluyla belirlemek için 100 mg yaprak veya kök materyalden toplam DNA'yı çıkarın (bkz.19).
- Numuneleri sıvı azotta öğütün, 100 mg bitki materyali alın ve toplam RNA'yı çıkarın. İstila üzerine bitki genlerinin (veya mantar genlerinin) ekspresyonunu belirlemek için qRT-PCR uygulayın (bkz.19).
- Yukarıdan bir dijital kamera ile bitkilerin fotoğraflarını çekin, mesafeyi her fotoğraf için aynı tutun. Yaprak alanını ölçün (örneğin, ImageJ22 veya BlattFlaeche17,19 ile; ölçeği ayarlamak için saksıların çapını kullanın) ve enfekte olmuş ve kontrol gruplarını karşılaştırın. Hastalık semptomlarının gelişimini kategorize edin (örneğin, daha küçük, daha sarımsı veya nekrotik yapraklar)13.
7. Verilerin analiz edilmesi
- Biyolojik kopyalara dayanarak ortalama ve standart sapmayı (± SD) hesaplayın.
- Enfekte gruptan gelen tüm sonuçları kontrolün sonucuna bölerek göreceli değerleri hesaplayın. Ortalamayı örneğin "alaya göre" veya "vahşi türe göre" olarak görüntüleyin.
- Gruplar arasındaki istatistiksel anlamlılığı belirler.

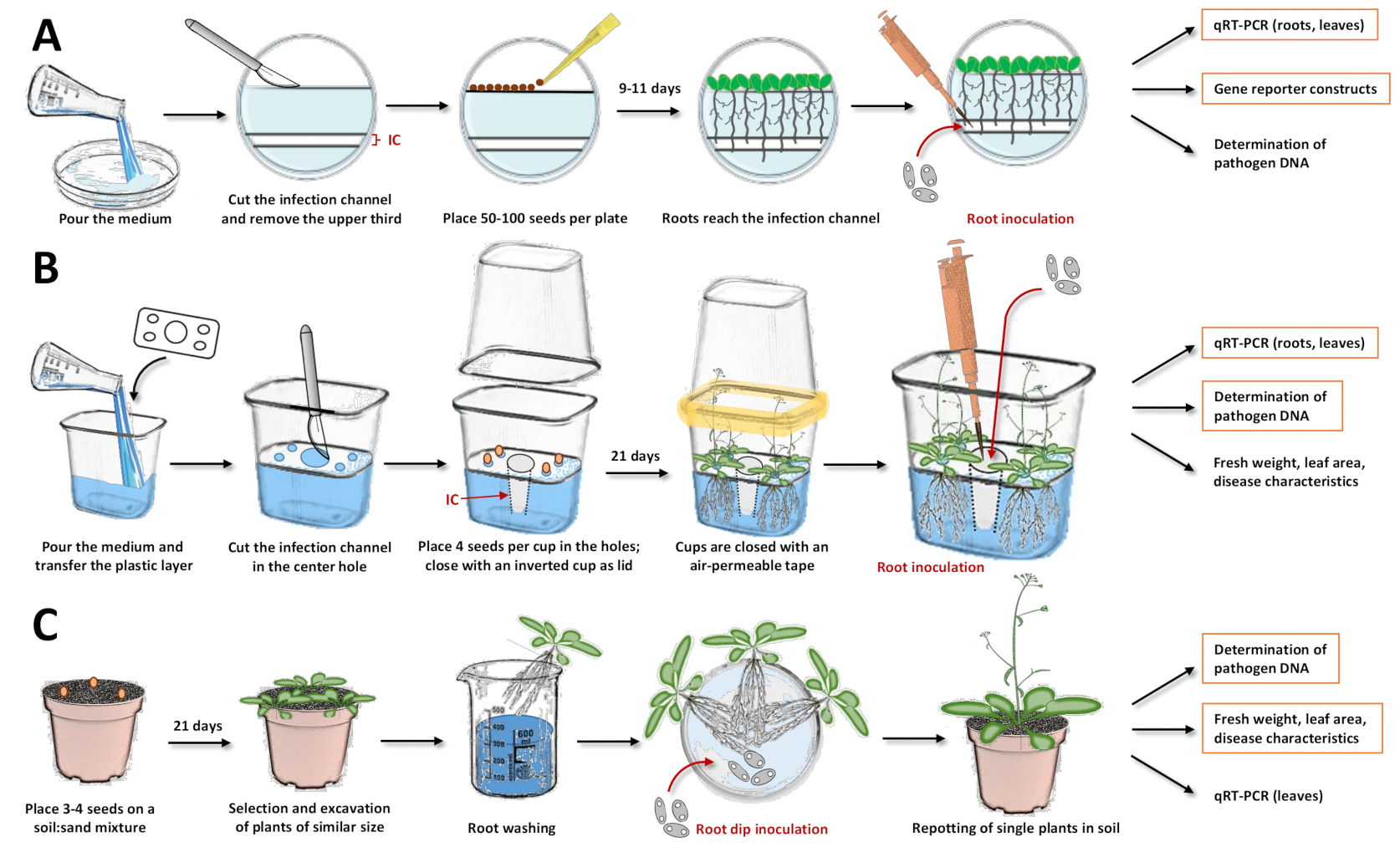
Şekil 1: Üç aşılama sisteminin ve protokollerdeki bireysel adımların derlenmesi. Bu rakamlar, Arabidopsis thaliana model tesisi ile sistemleri göstermektedir. Diğer bitki türleri için zamanlama ayarlanmalıdır. Turuncu kutular, ilgili sistemle sonraki analizlerin en çok önerildiği vurgulanır. (A) Petri tabaklarındaki aşılama sistemiiçin 17, ortamı dökün ve katılaşmasına izin verin. Tabakları gece boyunca buzdolabında saklayın. Daha sonra, üst üçte birini ve enfeksiyon kanalını (IC) bir neşterle kesin ve çıkarın (resimdeki beyaz alanlar agardan çıkarılırken, mavimsi alanlar agar'ı temsil eder). Tohumları kesilmiş yüzeye yerleştirin ve Petri bulaşıklarını kapatın. Tabakalaşmadan sonra, plakaları dikey olarak yerleştirin ve bitkilerin büyümesine izin verin. Köklerin çoğu enfeksiyon kanalına ulaştığında, spor solüsyonunu bir pipetle doğrudan kanala ekleyin. Çözümün eşit olarak dağıtıldığından emin olun. Petri tabaklarını kapatın ve bir büyüme odasında dikey olarak inkübe edin. İzlenebilecek yaklaşımlar kantitatif ters transkripsiyon PCR (qRT-PCR) ile ekspresyonel analiz, muhabir çizgileri ile mikroskopi ve mikrobiyal DNA'nın nicelleştirilmesidir. (B) Plastik kaplar19'daki aşılama sistemi için, ortamı dökün ve ayırıcı plastik tabakayı prefabrik deliklerle (tohumları yerleştirmek için köşelerde dört küçük delik ve enfeksiyon kanalı için merkezde bir büyük delik) aktarın. Ortamın katılaşmasına izin verin. Enfeksiyon kanalını (IC) elde etmek için orta delikteki agar ortamını bir neşterle kesin ve çıkarın. Ortamı daha küçük deliklerde çizin ve tohumları aktarın. Bardağı ters çevrilmiş bir bardakla kapatın ve hava geçirgen bantla kapatın (sarı renkle sembolize edilir). Bitkilerin büyümesine izin verin. Aşılama için, spor çözeltisini bir pipetle doğrudan enfeksiyon kanalına ekleyin. Sistemi kapatın ve büyüme odasında ekime devam edin. İzlenebilecek yaklaşımlar, qRT-PCR ile ekspresyonel analiz, mikrobiyal DNA'nın nicelleştirilmesi ve taze ağırlık, yaprak alanı veya diğer hastalık özelliklerinin belirlenmesidir. (C) "Kök daldırma aşılaması"15,17,23,24: toprak bazlı aşılama sistemi için, saksıları bir toprak: kum karışımı ile doldurun. Tohumları aktarın ve fidelerin büyümesine izin verin. Benzer büyüklükteki bitkileri kazın ve kökleri suda yıkayın. Yıkanmış kökleri, çözeltiyi sporlarla birlikte tutan bir Petri kabına yerleştirin. Kuluçkadan sonra, topraklı saksılara tek bitkiler yerleştirin. İzlenebilecek yaklaşımlar, qRT-PCR'li yapraklarda ekspresyonel analiz, mikrobiyal DNA'nın nicelleştirilmesi ve taze ağırlık, yaprak alanı veya diğer hastalık özelliklerinin belirlenmesidir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
Sonuçlar
Protokolü takiben, bitkiler V. longisporum ( suş Vl4325) veya V. dahliae (izole JR218) ile yetiştirildi ve aşılandı. Etkinliği kanıtlamak ve verilen protokollerin bazı yeteneklerini vurgulamak için çeşitli senaryolar tasarlanmıştır. Temsili sonuçlar gösterilir.
Antimikrobiyal indol-glukozinolat (IG) biyosentezinde rol oynayan genlerin ekspresyonel indüksiyonu, Verticillium enfeksiyonunun değe...
Tartışmalar
Toprak kaynaklı fitopatojenlerin neden olduğu muazzam verim kayıpları nedeniyle1, tarım stratejilerinin veya mahsul çeşitlerinin iyileştirilmesi gerekmektedir. Toprak kaynaklı hastalıkların patogenezine ilişkin sınırlı içgörü, daha dirençli bitkilerin gelişimini engellemektedir. Altta yatan patomekanizmaların araştırılması gerekir, bunun için sağlam bir metodolojik platform gereklidir. Bildirilen aşılama prosedürleri, kök-mikrop etkileşimlerindeki çok faktörlü ol...
Açıklamalar
Yazarların açıklayacak hiçbir şeyleri yoktur.
Teşekkürler
Yazarlar, Tim Iven ve Jaqueline Komorek'e bu yöntemler üzerindeki önceki çalışmaları için, Wolfgang Dröge-Laser grubuna (Farmasötik Biyoloji Bölümü, Würzburg Üniversitesi, Almanya) bu çalışma için gerekli ekipman ve kaynakları sağladıkları için ve Wolfgang Dröge-Laser ve Philipp Kreisz'e (her ikisi de Würzburg Üniversitesi) makalenin eleştirel redaksiyonu için teşekkür etmektedir. Bu çalışma "Deutsche Forschungsgemeinschaft" (DFG, DR273/15-1,2) tarafından desteklenmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Agar (Gelrite) | Carl Roth | Nr. 0039 | all systems described require Gelrite |
| Arabidopsis thaliana wild-type | NASC stock | Col-0 (N1092) | |
| Autoclave | Systec | VE-100 | |
| BlattFlaeche | Datinf GmbH | BlattFlaeche | software to determine leaf areas |
| Brassica napus wild-type | see Floerl et al., 2008 | rapid-cycling rape | genome ACaacc |
| Cefotaxime sodium | Duchefa | C0111 | |
| Chicanery flask 500 mL | Duran Group / neoLab | E-1090 | Erlenmeyer flask with four baffles |
| Collection tubes 50 mL | Sarstedt | 62.547.254 | 114 x 28 mm |
| Czapek Dextrose Broth medium | Duchefa | C1714 | |
| Digital camera | Nikon | D3100 18-55 VR | |
| Exsiccator (Desiccator ) | Duran Group | 200 DN, 5.8 L | Seal with lid to hold chlorine gas |
| Fluorescence Microscope | Leica | Leica TCS SP5 II | |
| HCl | Carl Roth | P074.3 | |
| KNO3 | Carl Roth | P021.1 | ≥ 99 % |
| KOH | Carl Roth | 6751 | |
| Leukopor | BSN medical GmbH | 2454-00 AP | non-woven tape 2.5 cm x 9.2 m |
| MES (2-(N-morpholino)ethanesulfonic acid) | Carl Roth | 4256.2 | Pufferan ≥ 99 % |
| MgSO4 | Carl Roth | T888.1 | Magnesiumsulfate-Heptahydrate |
| Murashige & Skoog medium (MS) | Duchefa | M0222 | MS including vitamins |
| NaClO | Carl Roth | 9062.1 | |
| Percival growth chambers | CLF Plant Climatics GmbH | AR-66L2 | |
| Petri-dishes | Sarstedt | 82.1473.001 | size ØxH: 92 × 16 mm |
| Plastic cups (500 mL, transparent) | Pro-pac, salad boxx | 5070 | size: 108 × 81 × 102 mm |
| Pleated cellulose filter | Hartenstein | FF12 | particle retention level 8–12 μm |
| poly klima growth chamber | poly klima GmbH | PK 520 WLED | |
| Potato Dextrose Broth medium | SIGMA Aldrich | P6685 | for microbiology |
| Pots | Pöppelmann GmbH | TO 7 D or TO 9,5 D | Ø 7 cm resp. Ø 9.5 cm |
| PromMYB51::YFP | see Poncini et al., 2017 | MYB51 reporter line | YFP (i.e. 3xmVenus with NLS) |
| Reaction tubes 2 mL | Sarstedt | 72.695.400 | PCR Performance tested |
| Rotary (orbital) shaker | Edmund Bühler | SM 30 C control | |
| Sand (bird sand) | Pet Bistro, Müller Holding | 786157 | |
| Soil | Einheitserde spezial | SP Pikier (SP ED 63 P) | |
| Solanum lycopersicum wild-type | see Chavarro-Carrero et al., 2021 | Type: Moneymaker | |
| Thoma cell counting chamber | Marienfeld | 642710 | depth 0.020 mm; 0.0025 mm2 |
| Ultrapure water (Milli-Q purified water) | MERK | IQ 7003/7005 | water obtained after purification |
| Verticillium dahliae | see Reusche et al., 2014 | isolate JR2 | |
| Verticillium longisporum | Zeise and von Tiedemann, 2002 | strain Vl43 |
Referanslar
- Mendes, R., Garbeva, P., Raaijmakers, J. M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiology Review. 37 (5), 634-663 (2013).
- Yadeta, K. A., Thomma, B. P. H. J. The xylem as battleground for plant hosts and vascular wilt pathogens. Frontiers in Plant Science. 4, 97 (2013).
- Delgado-Baquerizo, M., et al. The proportion of soil-borne pathogens increases with warming at the global scale. Nature Climate Change. 10 (6), 550-554 (2020).
- Berendsen, R. L., et al. Disease-induced assemblage of a plant-beneficial bacterial consortium. The ISME Journal. 12 (6), 1496-1507 (2018).
- Yuan, J., et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome. 6 (1), 156 (2018).
- Liu, H., et al. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytologist. 229 (5), 2873-2885 (2021).
- Wang, H., Liu, R., You, M. P., Barbetti, M. J., Chen, Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): role of bacterial diversity. Microorganisms. 9 (9), 1988 (2021).
- Inderbitzin, P., Subbarao, K. V. Verticillium systematics and evolution: how confusion impedes Verticillium wilt management and how to resolve it. Phytopathology. 104 (6), 564-574 (2014).
- Eynck, C., Koopmann, B., Grunewaldt-Stoecker, G., Karlowsky, P., von Tiedemann, A. Differential interactions of Verticillium longisporum und V. dahliae with Brassica napus with molecular and histological techniques. European Journal of Plant Pathology. 118 (3), 259-274 (2007).
- Floerl, S., et al. Defence reactions in the apoplastic proteome of oilseed rape (Brassica napus var. napus) attenuate Verticillium longisporum growth but not disease symptoms. BMC Plant Biology. 8, 129 (2008).
- Leonard, M., et al. Verticillium longisporum elicits media-dependent secretome responses with capacity to distinguish between plant-related environments. Frontiers in Microbiology. 11, 1876 (2020).
- Depotter, J. R. L., et al. Verticillium longisporum, the invisible threat to oilseed rape and other brassicaceous plant hosts. Molecular Plant Pathology. 17 (7), 1004-1016 (2016).
- Fröschel, C., et al. A gain-of-function screen reveals redundant ERF transcription factors providing opportunities for resistance breeding toward the vascular fungal pathogen Verticillium longisporum. Molecular Plant-Microbe Interactions. 32 (9), 1095-1109 (2019).
- Zhou, L., Hu, Q., Johansson, A., Dixelius, C. Verticillium longisporum and V. dahliae: infection and disease in Brassica napus. Plant Pathology. 55 (1), 137-144 (2006).
- Ralhan, A., et al. The vascular pathogen Verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots. Plant Physiology. 159 (3), 1192-1203 (2012).
- Reusche, M., et al. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium longisporum. Molecular Plant-Microbe Interactions. 26 (8), 850-860 (2013).
- Iven, T., et al. Transcriptional activation and production of tryptophan-derived secondary metabolites in Arabidopsis roots contributes to the defense against the fungal vascular pathogen Verticillium longisporum. Molecular Plant. 5 (6), 1389-1402 (2012).
- Reusche, M., et al. Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana. Environmental and Experimental Botany. 108, 23-37 (2014).
- Fröschel, C. In-depth evaluation of root infection systems using the vascular fungus Verticillium longisporum as soil-borne model pathogen. Plant Methods. 17 (1), 57 (2021).
- Karapapa, V. K., Bainbridge, B. W., Heale, J. B. Morphological and molecular characterization of Verticillium longisporum comb, nov., pathogenic to oilseed rape. Mycological Research. 101 (11), 1281-1294 (1997).
- Poncini, L., et al. In roots of Arabidopsis thaliana, the damage-associated molecular pattern AtPep1 is a stronger elicitor of immune signalling than flg22 or the chitin heptamer. PLoS One. 12 (10), 1-21 (2017).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Fradin, E. F., et al. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiology. 150 (1), 320-332 (2009).
- Singh, S., et al. The plant host Brassica napus induces in the pathogen Verticillium longisporum the expression of functional catalase peroxidase which is required for the late phase of disease. Molecular Plant-Microbe Interactions. 25 (4), 569-581 (2012).
- Zeise, K., von Tiedemann, A. Application of RAPD-PCR for virulence type analysis within Verticillium dahliae and Verticillium longisporum. Journal of Phytopathology. 150 (10), 557-563 (2002).
- Fröschel, C., et al. Plant roots employ cell-layer-specific programs to respond to pathogenic and beneficial microbes. Cell Host & Microbe. 29 (2), 299-310 (2021).
- Gigolashvili, T., et al. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. The Plant Journal. 50 (5), 886-901 (2007).
- Back, M. A., Haydock, P. P. J., Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathology. 51 (6), 683-697 (2002).
- Behrens, F. H., et al. Suppression of abscisic acid biosynthesis at the early infection stage of Verticillium longisporum in oilseed rape (Brassica napus). Molecular Plant Pathology. 20 (12), 1645-1661 (2019).
- Vorholt, J. A., Vogel, C., Carlström, C. I., Müller, D. B. Establishing causality: opportunities of synthetic communities for plant microbiome research. Cell Host & Microbe. 22 (2), 142-155 (2017).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır