
Method Article
RNA Yapısının Dimetil Sülfat Mutasyonel Profillemesi ile İn Vitro ve İn Hücre Dizilimi ile İncelenmesi
Bu Makalede
Özet
Protokol, mutasyonel profilleme deneyleri için RNA'nın dimetil sülfat ile modifiye edilmesi için talimat sağlar. İki alternatif kütüphane hazırlama yöntemi ile in vitro ve in vivo problama içerir.
Özet
RNA yapısının hemen hemen her biyolojik süreçteki rolü, özellikle son on yılda, giderek daha belirgin hale gelmiştir. Bununla birlikte, RNA kristalografisi veya kriyo-EM gibi RNA yapısını çözmeye yönelik klasik yaklaşımlar, hızla gelişen alana ve yüksek verimli çözümlere duyulan ihtiyaca ayak uyduramamıştır. Dimetil sülfat (DMS) MaPseq kullanılarak dizileme ile mutasyonel profilleme, RNA yapısını bir bazın DMS ile reaktivitesinden çıkarmak için dizilemeye dayalı bir yaklaşımdır. DMS, baz eşleşmediğinde adenosinlerdeki N1 azotunu ve sitozinlerdeki N3'ü Watson-Crick yüzlerinde metilleştirir. Modifiye RNA'nın termostabil grup II intron ters transkriptaz (TGIRT-III) ile ters transkripsiyonu, metillenmiş bazların cDNA'ya mutasyon olarak dahil edilmesine yol açar. Elde edilen cDNA'yı sıralarken ve bir referans transkriptine geri eşleştirirken, her baz için göreceli mutasyon oranları, bazın eşleştirilmiş veya eşleşmemiş olarak "durumunun" göstergesidir. DMS reaktiviteleri hem in vitro hem de hücrelerde yüksek sinyal-gürültü oranına sahip olsa da, bu yöntem işleme prosedürlerinde önyargıya duyarlıdır. Bu önyargıyı azaltmak için, bu makale hücrelerde DMS ve in vitro transkribe RNA ile RNA tedavisi için bir protokol sunmaktadır.
Giriş
RNA'nın hem yapısal1,2 hem de katalitik3 özelliklere sahip olduğunun keşfedilmesinden bu yana, RNA'nın önemi ve düzenleyici işlevinin çok sayıda biyolojik süreçte yavaş yavaş ortaya çıkarılmıştır. Nitekim RNA yapısının gen regülasyonu üzerindeki etkisi giderek artan bir ilgi görmüştür4. Proteinler gibi, RNA da nükleotitlerin dizisine, baz eşleştirme etkileşimlerinin 2B haritalanmasına ve bu baz eşleştirilmiş yapıların 3B katlanmasına atıfta bulunan birincil, ikincil ve üçüncül yapılara sahiptir. Üçüncül yapının belirlenmesi, RNA'ya bağımlı süreçlerin arkasındaki kesin mekanizmaları anlamanın anahtarı olsa da, ikincil yapı da RNA fonksiyonu hakkında oldukça bilgilendiricidir ve daha fazla 3D katlama5'in temelidir.
Bununla birlikte, RNA yapısının belirlenmesi, geleneksel yaklaşımlarla özünde zor olmuştur. Proteinler için, kristalografi, nükleer manyetik rezonans (NMR) ve kriyojenik elektron mikroskobu (kriyo-EM), yapısal motiflerin çeşitliliğini belirlemeyi mümkün kılarken, yalnızca diziden yapı tahminine izin verir6, bu yaklaşımlar RNA'lara yaygın olarak uygulanamaz. Gerçekten de, RNA'lar, amino asit muadillerine kıyasla çok daha fazla konformasyonel ve rotasyonel özgürlüğe sahip yapı taşlarına (nükleotitler) sahip esnek moleküllerdir. Ayrıca, baz eşleştirme yoluyla etkileşimler, amino asit kalıntılarınınkinden daha dinamik ve çok yönlüdür. Sonuç olarak, klasik yaklaşımlar sadece iyi tanımlanmış, oldukça kompakt yapılara sahip nispeten küçük RNA'lar için başarılı olmuştur7.
RNA yapısını belirlemek için bir başka yaklaşım, yeni nesil dizileme (NGS) ile birleştirilmiş kimyasal problamadır. Bu strateji, bir RNA dizisindeki her bir bazın bağlanma durumu (yani ikincil yapısı) hakkında bilgi üretir. Kısacası, bir RNA molekülündeki baz eşleştirmesine katılmayan bazlar, küçük kimyasal bileşikler tarafından farklı şekilde modifiye edilir. Bu RNA'ların özel ters transkriptazlarla (RT'ler) ters transkripsiyonu, modifikasyonları mutasyonlar olarak tamamlayıcı deoksiribonükleik aside (cDNA) dahil eder. Bu cDNA molekülleri daha sonra polimeraz zincir reaksiyonu (PCR) ile çoğaltılır ve sıralanır. Bağlı veya bağlanmamış olarak "durumları" hakkında bilgi edinmek için, ilgilenilen bir RNA'daki her bir bazdaki mutasyon frekansları hesaplanır ve kısıtlamalar8 olarak yapı tahmin yazılımına girilir. En yakın komşu kuralları9 ve minimum serbest enerji hesaplamaları10'a dayanarak, bu yazılım elde edilen deneysel verilere en uygun yapı modelleri üretir11,12.
DMS-MaPseq, Watson-Crick yüzlerinde adenosinlerdeki N1 azotunu ve sitozinlerdeki N3 azotunu oldukça spesifik bir şekilde metilleştiren DMS'yi kullanır13. Ters transkripsiyonda termostabil grup II intron ters transkriptaz (TGIRT-III) kullanılması, benzeri görülmemiş sinyal-gürültü oranlarına sahip mutasyonel profiller oluşturur, hatta iki veya daha fazla alternatif konformasyon tarafından üretilen örtüşen profillerin dekonvolüsyonuna izin verir14,15. Ayrıca, DMS hücre zarlarına ve tüm dokulara nüfuz edebilir ve fizyolojik bağlamlarda sondalamayı mümkün kılar. Bununla birlikte, işleme prosedüründeki değişiklikler sonuçları etkileyebileceğinden, kaliteli verilerin oluşturulması zordur. Bu nedenle, önyargıyı azaltmak ve yeni gelenleri karşılaşabilecekleri zorluklarla yönteme yönlendirmek için hem in vitro hem de hücre içi DMS-MaPseq için ayrıntılı bir protokol sağlıyoruz. Özellikle son SARS-CoV2 pandemisi ışığında, RNA virüsleri hakkındaki yüksek kaliteli veriler, gen ekspresyonunu incelemek ve olası terapötikleri bulmak için önemli bir araçtır.
Protokol
NOT: Bu protokolde kullanılan tüm malzemeler, yazılımlar, reaktifler, aletler ve hücrelerle ilgili ayrıntılar için Malzeme Tablosu'na bakın.
1. Gen-spesifik in vitro DMS-MaP
- RNA in vitro transkripsiyonu
- İlgilenilen RNA'nın dizisini çift sarmallı (ds) DNA olarak elde edin (örneğin, önceden var olan / genomik DNA'dan DNA fragmanları, plazmidler veya PCR olarak). DNA dizisi bir polimeraz promotoru içeriyorsa, adım 3'e atlayın.
- İstenilen DNA fragmanının yukarı yönünde bir RNA polimeraz promotor bağlamak için örtüşen PCR gerçekleştirin (T7 polimeraz için ileri astar: 5' TAATACGACTCACTATAGG + hedef dizi 3'ün ilk bazları).
- In vitro DNA fragmanını RNA'ya aktarın. RNA'yı daima buzun üzerinde tutun.
- DNA'yı bir DNaz kullanarak sindirin.
- RNA'yı sütun tabanlı bir yaklaşım (adım 2.4) veya etanol çökeltmesi (adım 2.5) kullanarak izole edin. Uygun bir hacimde sülte, ~ 50 μg'lik bir verim bekleyin.
- RNA bütünlüğünü bir agaroz jeli üzerinde çalıştırarak sağlayın; RNA'yı çalıştırmadan önce 70 ° C'de 2-3 dakika boyunca denatüre edin.
NOT: Tampon ve agaroz, RNA'yı bozan ve RNA örneğini kirletebilecek RNazlar içerebilir. Prekast agaroz jelleri daha önce bu laboratuvarda kullanılmıştır; Sonuçlar (özellikle RNA ile) zaman zaman belirsiz olmuştur. En iyi sonuçlar agaroz veya PAGE jelleri ile elde edildi. - RNA'nın doğrudan kullanımı, çözülmeden sonra bozulma görülmedikçe RNA'yı birkaç ay boyunca -80 ° C'de saklayın.
- İn vitro DMS modifikasyonu (105 mM DMS'de)
- Yeterli miktarda yeniden katlanır tampon hazırlayın (0.4 M sodyum kakodilat, pH 7.2, 6 mM MgCl2 içerir).
NOT: Her reaksiyon için (100 μL'lik son hacim), 89 μL yeniden katlanır tampon ekleyin. - Her reaksiyon için, 89 μL yeniden katlanır tamponu belirlenmiş bir 1,5 mL tüpe aktarın ve kimyasal bir davlumbazın altına yerleştirilmiş bir termoshaker'da 37 ° C'de ön ısıtın.
NOT: DMS oldukça toksiktir ve bir indirgeyici madde tarafından söndürülene kadar daima kimyasal bir davlumbazın altında tutulmalıdır. - 10 μL nükleaz içermeyen suda (NF H2O) 1-10 pmol RNA salınımlı; bir PCR tüpüne transfer.
- RNA'yı denatüre etmek için 95 ° C'de bir termosikler içinde 1 dakika boyunca inkübe edin.
- Yanlış katlanmayı önlemek için hemen bir buz bloğuna yerleştirin.
- RNA örneğini 37 ° C'de yeniden katlanır tampon ile belirlenen tüpe ekleyin, iyice karıştırın ve RNA'yı yeniden şekillendirmek için 10-20 dakika boyunca inkübe edin.
NOT: Çoğu RNA, milisaniyeler ila saniyeler arasında katlanacaktır, ancak istisnalar16 vardır. - RNA örneğine 1 μL% 100 (10.5 M) DMS ekleyin ve dakikada 800-1.400 dönüşte (rpm) sallanırken 5 dakika boyunca inkübe edin.
NOT: DMS hidrofobik olduğundan ve yeniden katlanan tamponda tamamen çözünmeyebileceğinden, bu adımda sallama (veya diğer karıştırma yöntemleri) çok önemlidir. Reaksiyon sürelerindeki sapmalar, DMS reaktivitelerinin tekrarlanabilirliğini etkileyebilir. Pipetleme hatasını en aza indirmek için DMS, numuneye eklenmeden önce %100 etanol içinde çözülebilir, eğer son konsantrasyon %1 (105 mM) DMS korunursa. Arıtılmamış bir kontrol için DMS, dimetil sülfoksit (DMSO) veya su ile değiştirilebilir. - 5 dakikalık reaksiyon süresinden sonra, 60 μL% 100 β-merkaptoetanol (BME) ile söndürün, iyice karıştırın ve RNA'yı hemen buzun üzerine yerleştirin.
NOT: RNA, temizlemek için reaksiyonu BME ile söndürdükten sonra davlumbazdan güvenli bir şekilde çıkarılabilir. Bununla birlikte, BME'nin çevreye doğrudan maruz kalması, güçlü kokusu ve tahriş edici özellikleri nedeniyle hala önlenmelidir. - RNA'yı sodyum asetat-etanol çökeltmesi (bkz. adım 2.5) veya sütun bazlı bir yaklaşımla (bkz. adım 2.6) temizleyin ve 10 μL suda salın.
- RNA'yı bir spektrofotometre kullanarak sayısallaştırın.
- Modifiye RNA'yı -80 ° C'de depolamanın doğrudan kullanımı.
NOT: DMS tedavisinden sonra RNA daha az kararlı olduğu için uzun süreli depolamadan kaçınılmalıdır.
- Yeterli miktarda yeniden katlanır tampon hazırlayın (0.4 M sodyum kakodilat, pH 7.2, 6 mM MgCl2 içerir).
- Modifiye RNA'nın gen-spesifik RT-PCR'si
NOT: DMS ile işlenmiş parçaların RT-PCR kurulumu için Şekil 1'e bakın.- 10 μL nükleaz içermeyen (NF)H2O'da 100 ng modifiye RNA'nın salınımı Bir PCR tüpüne transfer.
- Tüpe 4 μL 5x birinci iplik tamponu (FSB), 1 μL dNTP karışımı (her biri 10 mM), 1 μL 0.1 M ditiyotreitol (DTT) (donma-çözülme döngülerinden kaçının), 1 μL RNaz inhibitörü, 1 μL 10 μM ters primer (tek astar veya bir astar havuzu) ve 1 μL TGIRT III ekleyin.
NOT: Bir astar havuzu için, her bir astarın 10 μM'lik 1 μL'sini doğrudan RT'ye eklemeyin; Bunun yerine, önce astarları karıştırın ve karışımdan 1 μL ekleyin (toplam 10 μM astar konsantrasyonunda). - Bir termosiklette 30 dakika ila 1,5 saat arasında 57 ° C'de (tipik olarak, 500 nt'lik bir ürün yapmak için 30 dakika yeterlidir) inkübe edin.
- 1 μL 4 M NaOH ekleyin, pipetle karıştırın ve RNA'yı bozmak için 3 dakika boyunca 95 ° C'de inkübe edin.
NOT: Bu adım, RNA'yı bozarak TGIRT'yi cDNA'dan serbest bıraktığı için çok önemlidir. Atlanırsa, aşağı akış PCR etkilenebilir. - Astarları yeterince uzaklaştıran ve 10 μL NF H 2 O'da salınım yapan sütun tabanlı bir yaklaşım (bkz. adım2.6) kullanarak temizleyin.
- PCR-reaksiyonun 25 μL'si başına ters transkripsiyon ürününün 1 μL'sini kullanarak cDNA'yı, verim ve sadakati dengelemek için tasarlanmış bir PCR kiti ile yükseltin.
NOT: Astarlar ~ 60 °C erime sıcaklığına sahip olmalıdır. - PCR başarısını doğrulamak için PCR ürününün 2 μL'sini bir agaroz jeli veya prekast agaroz jeli üzerinde çalıştırın.
- İdeal olarak, PCR'den sonra sadece bir bant gösterilmelidir. Öyleyse, sütun tabanlı bir yaklaşım kullanarak reaksiyonu temizleyin. Alternatif bantlar varsa, jelden doğru bandı çıkarmak için kalan PCR reaksiyonunu kullanın. Yeterince küçük bir hacimde (örneğin, 10 μL) sülte.
- Bir spektrofotometre kullanarak çıkarılan parçaları ölçün.
- İstenilen dizileme platformuna uygun bir yaklaşım kullanarak dizileme için dsDNA parçalarını dizine ekleyin.

Resim 1: DMS ile işlenmiş büyük parçaların RT-PCR'si için deney düzeneği. Modifiye edilmiş bir RNA üzerinde ters transkripsiyon yapılırken, astarın tavlandığı dizi üzerindeki modifikasyonlar kaydedilmeyecektir. Bu nedenle, parçaların uzunluğu 400-500 bp'yi aştığında, burada örneklendiği gibi, astar bölgelerinde üst üste binen parçaların tasarlanması gerekir. Parçaların uzunluğu, sıralama ihtiyaçlarına bağlıdır. Eşleştirilmiş uçlu 150 döngü dizilimi kullanıldığında, parçalar 300 bp'yi geçmemelidir. Kısaltmalar: RT-PCR = ters transkripsiyon polimeraz zincir reaksiyonu; DMS = dimetil sülfat. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
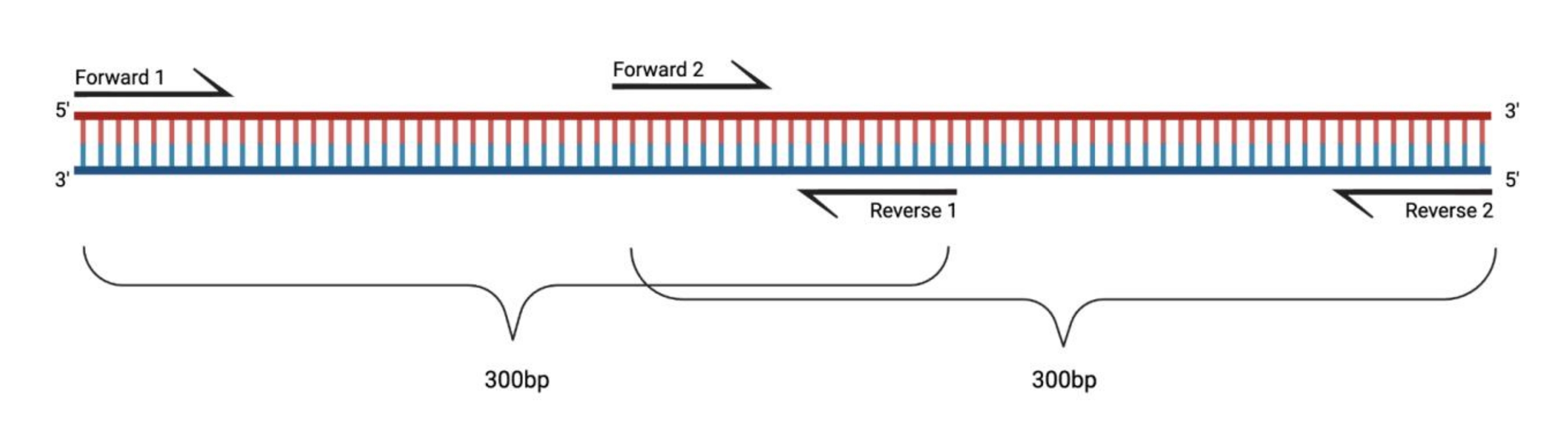{kind=link}
2. Virüs bulaşmış hücreleri kullanan tüm genom DMS-MaP
NOT: Hücrelerde, DMS tedavisi yukarıda açıklanan gen-spesifik amplifikasyon yaklaşımı ile de birleştirilebilir. Tüm genom kütüphanesi, tek bir gen üzerinde tam kapsama alanı elde etmek için muazzam bir dizileme derinliği gerektirir. Bununla birlikte, viral RNA'lar ekstraksiyondan sonra ribodeplemiş RNA'nın önemli bir bölümünü oluşturuyorsa, tüm genom dizilimi uygun olacaktır. Ayrıca, diğer zenginleştirme yöntemleri tüm genom kütüphanesi oluşturma yöntemiyle birleştirilebilir.
- DMS tedavisi
- İstenilen enfeksiyon aşamasına kadar virüsle enfekte olmuş hücreleri büyütün.
- Hücre kabını, hem virüsleri gerekli biyogüvenlik seviyesinde hem de DMS gibi ajanlar tarafından üretilen kimyasal dumanları işlemek için uygun olan özel bir duman davlumbazına aktarın.
- Kültür ortamına% 2,5'lik bir DMS hacmi ekleyin ve kabı (tipik olarak 10 cm'lik bir plaka) parafilm ile kapatın.
NOT: DMS ile az değişiklik yapmak ve aşırı değiştirmek kolaydır. DMS'yi doğrudan hücrelere eklerken, iyice karıştırmak çok önemlidir. Alternatif olarak, yeni ortamı 37 ° C'de 50 mL'lik bir konik tüp içinde önceden ısıtın ve DMS'yi doğrudan kuvvetlice sallayarak ekleyin. Harcanan ortamı hücreler üzerinde boşaltın ve DMS içeren ortamda yavaşça pipet çekin. - 5 dakika boyunca 37 °C'lik bir inkübatöre aktarın.
NOT: DMS'nin inkübatörün dışında tutulması için gereken süreye bağlı olarak, 5 dakikanın aşırı modifikasyona yol açması mümkündür. DMS'yi eklemekten inkübasyona kadar geçen süreyi ≤1 dakikaya kadar tutun. Deney ilk kez gerçekleştiriliyorsa, optimum modifikasyon oranını bulmak ve sonuçların bir konsantrasyon penceresinde sağlam olmasını sağlamak için bir DMS titrasyonu yapmanız ve inkübasyon süresini (3 dakika ile 10 dakika arasında) değiştirmeniz önerilir. - DMS içeren ortamı dikkatlice pipetle çıkarın (uygun kimyasal atıklara) ve nazikçe 10 mL durdurma tamponu ekleyin (% 30 BME içeren PBS [örneğin, 3 mL BME ve 7 mL PBS]).
NOT: DMS ve BME ilavesi, hücreler kuvvetli bir şekilde yapışmamışsa, hücreleri plakadan kaldırabilir. Hücreler kalkıyorsa, DMS içeren ortamı çıkarmak yerine süspansiyon hücreleri olarak muamele edilebilir, durdurma tamponunu doğrudan ekleyin ve DMS ve BME içeren hücreleri 50 mL'lik bir konik tüpe kazıyın. 3.000 × g'da 3 dakika santrifüj yaparak hücreleri peletleyin; Büyük damlacıklarda hücrelerin altına pelet edebilen herhangi bir kalıntı DMS'den kurtulduğunuzdan emin olun. DMS ortamı başlangıçta çıkarılamıyorsa,% 30 BME'de ekstra bir yıkama adımı önerilir. - Hücreleri kazıyın ve 15 mL'lik bir konik tüpe aktarın.
- 3.000 × g'da 3 dakika boyunca santrifüjleme ile pelet.
- Süpernatantı çıkarın ve 10 mL PBS ile 2 kat yıkayın.
- Mümkün olduğunca fazla PBS kalıntısını dikkatlice çıkarın.
- Peleti uygun miktarda RNA izolasyon reaktifi içinde çözün (örneğin; bir T75 kültür şişesi için 3 mL, 10 cm'lik bir plaka için 1 mL).
NOT: Reaktifin yetersiz miktarları RNA verimini etkileyebilir.
- RNA ekstraksiyonu ve ribozomal RNA (rRNA) tükenmesi
- RNA izolasyon reaktifindeki 1 mL homojenize hücreye, 200 μL kloroform, parlak pembeye kadar 15-20 s vorteks ekleyin ve daha sonra faz ayrımı görünene kadar 3 dakikaya kadar inkübe edin.
NOT: Pembe lipit fazı altta yerleşmelidir. Eğer durum böyle değilse, vorteks zamanı muhtemelen yetersizdi. - 4 ° C'de 15 dakika boyunca maksimum hızda (~ 20.000 × g) döndürün.
- Üst sulu fazı yeni bir tüpe aktarın.
- RNA'yı sodyum asetat-etanol çökeltmesi (bkz. adım 2.5) veya sütun bazlı bir yaklaşımla (bkz. adım 2.6) temizleyin ve yeterli miktarda NFH2O hacminde salın.
- Bir agaroz jeli üzerindeki RNA bütünlüğünü kontrol edin. İki ribozomal alt birime karşılık gelen iki bant arayın.
- Tercih edilen yaklaşımı kullanarak rRNA'ları tüketin veNF H 2O'nun yeterli hacminde (tipik olarak 20-50 μL) salınımlı.
NOT: Aşağı akış uygulamaları için, 8 μL'lik bir hacimde ~ 500 ng toplam RNA önerilmektedir. ribozomal olmayan RNA'lar tipik olarak toplam RNA'nın sadece% 5-10'unu oluşturur. - Bir spektrofotometre kullanarak sayısallaştırın.
- RNA izolasyon reaktifindeki 1 mL homojenize hücreye, 200 μL kloroform, parlak pembeye kadar 15-20 s vorteks ekleyin ve daha sonra faz ayrımı görünene kadar 3 dakikaya kadar inkübe edin.
- Kütüphane oluşturma
- Kütüphaneler oluşturmak için genlere özgü RT-PCR veya diğer yaklaşımları kullanın15. Astarlama için rastgele hexamers kullanıyorsanız, hexamer tavlamasına izin vermek için düşük bir Tm'de (37-42 °C) bir inkübasyon adımı ekleyin.
NOT: Standart kütüphane üretim kitleri, RT enzimini TGIRT ile değiştirerek ve RT sıcaklığını 57 °C'ye değiştirerek de kullanılabilir.
- Kütüphaneler oluşturmak için genlere özgü RT-PCR veya diğer yaklaşımları kullanın15. Astarlama için rastgele hexamers kullanıyorsanız, hexamer tavlamasına izin vermek için düşük bir Tm'de (37-42 °C) bir inkübasyon adımı ekleyin.
- RNA Temizleme ve Konsantratör sütunlarını kullanarak sütun tabanlı RNA temizleme
NOT: Tüm adımlar oda sıcaklığında yapılmalıdır.- 50 μL'lik bir hacme getirmek için numune tüpüne NF H2O ekleyin.
- Numuneye 100 μL bağlayıcı tampon ve 150 μL %100 etanol ekleyin.
- Karıştırın ve bir döndürme sütununa aktarın.
- 30 s için 10.000-16.000 × g'de döndürün; akış geçişini atın.
- 400 μL RNA hazırlık tamponu ekleyin.
- 30 s için 10.000-16.000 × g'de döndürün; akış geçişini atın.
- 700 μL RNA yıkama tamponu ekleyin.
- 30 s için 10.000-16.000 × g'da döndürün; akış geçişini atın.
- 400 μL RNA yıkama tamponu ekleyin.
- 30 s için 10.000-16.000 × g'de döndürün; akış geçişini atın.
- (İsteğe bağlı) Kolonu yeni bir toplama tüpüne aktarın ve 2 dakika boyunca 10.000-16.000 × g'de döndürün.
- Kolonu temiz bir RNAse içermeyen tüpe aktarın ve uygun miktarda NF H2O ekleyin.
- 1 dakika boyunca 10.000-16.000 × g'de döndürün.
- Asit fenol-kloroform RNA ekstraksiyonu.
- Eşit miktarda asit fenol ekleyin:kloroform:izoamil alkol.
- Vorteksi iyice alın ve 5 dakika boyunca 14.000 × g'de santrifüj yapın.
- Faz ayrımı yoksa, 20 μL 2 M NaCl ekleyin ve santrifüjlemeyi tekrarlayın.
- Sulu fazı yeni bir tüpe aktarın.
- 500 μL izopropanol ve 2 μL ko-çökeltici ekleyin.
- RT'de 3 dakika boyunca karıştırın ve kuluçkaya yatırın; daha sonra, gece boyunca -80 ° C'de inkübe edin.
- RNA'yı 4 ° C'de 30 dakika boyunca maksimum hızda (~ 20.000 × g) santrifüjleme ile peletleyin.
- Peleti 200 μL buz soğuk% 70 etanol ile yıkayın.
- 5 dakika boyunca maksimum hızda (~ 20.000 × g) döndürün; akış geçişini atın.
- Peleti uygun miktarda NFH2O ile yeniden askıya alın.
- Oligo Clean ve Concentrator sütunlarını kullanarak sütun tabanlı cDNA temizliği
NOT: Tüm adımlar oda sıcaklığında yapılmalıdır.- 50 μL'lik bir hacme getirmek için numune tüpüne NF H2O ekleyin.
- 100 μL bağlayıcı tampon ve 400 μL% 100 etanol ekleyin.
- Karıştırın ve bir döndürme sütununa aktarın.
- 30 s için 10.000-16.000 × g'de döndürün; akış geçişini atın.
- 750 μL DNA yıkama tamponu ekleyin.
- 30 s için 10.000-16.000 × g'de döndürün; akış geçişini atın.
- (İsteğe bağlı) Kolonu yeni bir toplama tüpüne aktarın ve 2 dakika boyunca 10.000-16.000 × g'de döndürün.
- Kolonu temiz bir RNAse içermeyen tüpe aktarın ve uygun miktarda NF H2O ekleyin.
- 1 dakika boyunca 10.000-16.000 × g'da döndürün.
3. Sıralama verilerinin analizi
NOT: DMS-MaP dizileme verilerinden RNA ikincil yapı modelleri oluşturmak için, elde edilen .fastq dosyalarının birkaç farklı adımda işlenmesi gerekir. Bu adımlar,
- Adaptör dizilerini TrimGalore veya Cutadapt ile kırpın.
- Bowtie2 kullanarak okumaları referans dizileriyle (.fasta formatı) eşleyin.
- Özel RNA yapı yazılımı (örneğin, DREEM14, RNA-Framework17 veya benzeri) ile okumaları sayın ve reaktivite profilleri oluşturun.
- (İsteğe bağlı) DREEM14, DRACO17, DANCE-MaP18 veya benzerlerini kullanarak alternatif RNA konformasyonlarını bulmak için okumaları kümeleyin.
- RNAStructure12, ViennaRNA veya benzerlerini kullanarak reaktivite profillerine dayanarak minimum serbest enerji yapısını tahmin edin.
- RNA11 yapısını VARNA (https://varna.lri.fr/) veya benzerini kullanarak görselleştirin.
NOT: Pratiklik için, DREEM (www.rnadreem.org) ve RNA-Framework19 gibi yazılımlar, analiz sürecini kolaylaştıran 1--5 adımlarını boru hatlarına büyük ölçüde dahil eder. Bununla birlikte, herhangi bir yapı tahmini dikkatle ele alınmalıdır (örneğin, yapının verilerle olan anlaşmasını doğrulayarak20.
Sonuçlar
Gen-spesifik in vitro DMS-MaP
SARS2'nin 5'UTR'sini incelemek için, virüsün ilk 300 bp'si, üç primerin yanı sıra bir gBlock dizisi olarak sipariş edildi. Bunlar, parçayı ("FW" ve "RV") PCR aracılığıyla yaymak için iki astar ve T7 promotorunu ("FW-T7") bağlamak için bir tane içeriyordu. Bu diziler Tablo 1'de görülebilir.
| Ad | Sıralama (5'->3') |
| FW | ATTAAAGGTTTATACCTTCCCAGGTAAC |
| RV | GCAAACTGAGTTGGACGTGT |
| FW-T7 Serisi | TAATACGACTCACTATAGG ATTAAAGGTTTATACCTTCCCAGGTAAC |
Tablo 1: SARS-CoV2 5'UTR'nin DMS-MaP RT-PCR için primer dizisi. Burada, in vitro transkripsiyon için bir DNA şablonu oluşturmak için FW-T7 ve RV'ye ihtiyaç vardır, RV ters transkripsiyonda kullanılır ve FW-RV primer-çifti, cDNA'nın sonraki PCR amplifikasyonunda kullanılır. Primerler, SARS-CoV2 genomunun (FW) en başlangıcına ve ilgili bölgenin hemen aşağısındaki diziye tavlanır. Kısaltmalar: DMS-MaP = Dimetil sülfat kullanarak dizileme ile mutasyonel profilleme; RT-PCR = ters transkripsiyon polimeraz zincir reaksiyonu; SARS-CoV2 = şiddetli akut solunum sendromu-koronavirüs 2; UTR = çevrilmemiş bölge; RV = ters astar; FW = ileri astar.
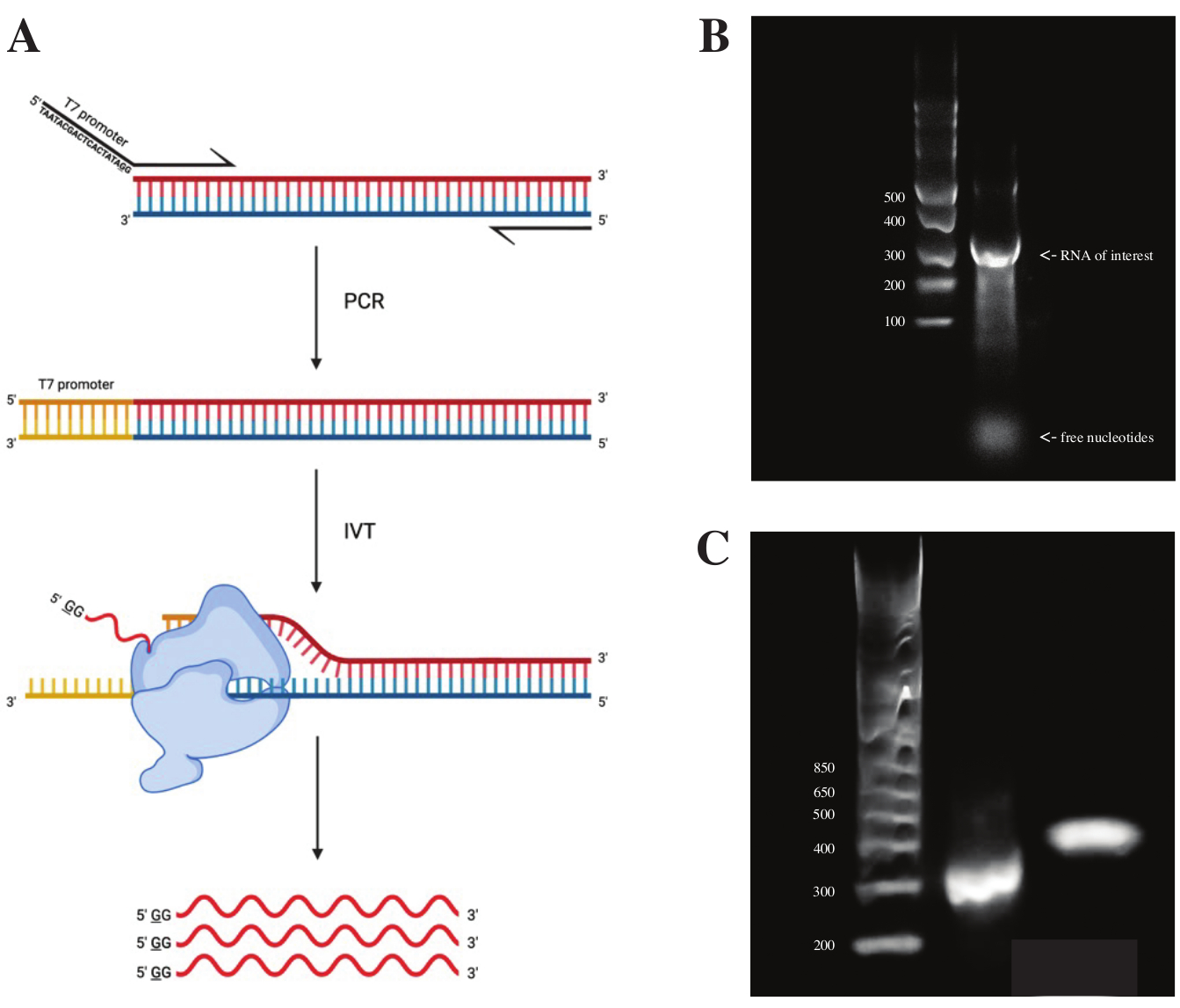
gBlock fragmanından RNA üretmek için, T7 polimeraz promotorunun dizisi, Şekil 2A'da görülen şemaya göre PCR ön karışımı kullanılarak üst üste binen PCR kullanılarak bağlanmıştır. Uzatılmış fragmandan, RNA T7 Transkripsiyon Kiti kullanılarak üretildi. DNA şablonu daha sonra RNA Clean & Concentrator sütunları kullanılarak izole edilen DNaz ve RNA kullanılarak sindirildi.
İn vitro transkripsiyonun kalite kontrolü, RNA ürününün bir ssRNA merdiveni yanında% 1'lik bir agaroz jeli üzerinde çalıştırılmasıyla yapıldı. Sadece bir bant görülebildiği için in vitro DMS problaması ve RT-PCR yapıldı (bkz. Şekil 2B).
PCR reaksiyonunun başarısını doğrulamak için, numune bir dsDNA merdiveni kullanılarak% 2'lik bir agaroz jeli üzerinde çalıştırıldı. İndekslemeden sonra, bant aynı jel üzerinde ~ 150 bp daha yüksek çalışmalı ve indeksleme primerlerinin boyutunu hesaba katmalıdır.

Şekil 2: DNA şablonunun in vitro transkripsiyonu. (A) Henüz içsel bir RNA polimeraz promotoruna sahip olmayan bir DNA şablonunu in vitro olarak transkripte etmek için, şablon önce örtüşen PCR ile eklenmelidir. Bu, istenen parçayla örtüşen ilk bazların yukarı akışı olan TAATACGACTCACTATAGG (T7 RNA polimeraz durumunda) dizisini içeren bir ileri astar kullanılarak yapılır. Buradaki altı çizili taban, polimerazın transkripsiyon başlangıç bölgesini sembolize eder. Promotör dsDNA fragmanına bağlandıktan sonra, T7 polimeraz tarafından transkripte edilebilir. Önemli olarak, polimeraz, söz konusu promotör dizisine karşı olan ipliği şablon (mavi) olarak kullanır ve etkili bir şekilde, belirtilen promotör dizisinin (kırmızı) hemen aşağısındaki diziyle aynı RNA'yı oluşturur. (B) Bir ssRNA Merdiveni (şerit 1) ve in vitro transkribe RNA ürünü 300 nt'de (şerit 2) bulunan% 1'lik bir agaroz jeli. (C) GeneRuler 1 kb artı Merdiven (şerit 1) ile %2 agaroz jeli, RT-PCR'den sonra 300 bp'de (şerit 2) çalışan PCR ürünü ve 470 bp'de (şerit 3) çalışan kütüphane hazırlığından sonra indekslenmiş parça. Kısaltmalar: RT-PCR = ters transkripsiyon polimeraz zincir reaksiyonu; DMS = dimetil sülfat; nt = nükleotidler; dsDNA = çift sarmallı DNA; ssRNA = tek sarmallı RNA. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
Virüs bulaşmış hücreleri kullanarak in vivo DMS-MaP tüm genom
DMS tedavisinden önce, HCT-8 hücreleri OC43 ile enfekte edildi. Enfeksiyondan 4 gün sonra (dpi) sitopatik bir etki (CPE) gözlendiğinde ( Şekil 3A'da görüldüğü gibi), bu hücreler tedavi edildi ve RNA ekstrakte edildi ve ribodepledi. Toplam RNA'yı bir agaroz jeli üzerinde çalıştırırken, toplam RNA kütlesinin yaklaşık% 95'ini oluşturan ribozomun 40S ve 60S alt birimlerini oluşturan iki parlak bant görülebiliyordu (bkz. Şekil 3B). RNA ekstraksiyonu başarısız olduğunda veya bozulduğunda (örneğin, çoklu donma-çözülme döngüleri ile), RNA bozunma ürünleri jelin dibinde görülebiliyordu (bkz. Şekil 3C, ikinci şerit). Ayrıca, rRNA tükenmesinden sonra, iki parlak bant ortadan kayboldu ve şeritte bir yayma bıraktı (bkz. Şekil 3C, üçüncü şerit). Son olarak, kütüphane hazırlığından sonra, örnekler farklı boyut dağılımlarına sahipti ve son PAGE jeli üzerinde bir yayma olarak gösterildi. Bant, bu kütüphaneleri analiz etmek için planlanan 150 x 150 çift uçlu dizileme çalışması ile uyumlu olarak 200 nükleotid (nt) ile 500 nt arasında eksize edildi. En önemlisi, ~ 150 nt'de çalışan adaptör dimerleri ayrıldı (bkz. Şekil 3D).

Şekil 3: Virüs bulaşmış hücrelerle in vivo DMS-MaP'nin kontrol noktaları . (A) Virüs bulaşmış HCT-8 hücrelerinin ışık mikroskobu görüntüsü, 4 gün dpi. Hücre ölümüne bağlı olumsuz etkileri en aza indirirken toplam RNA'dan mümkün olan en yüksek viral RNA verimini elde etmek için, resimde görüldüğü gibi, CPE başladığında veya hatta bundan önce DMS eklenmelidir. (B) 1 μg toplam RNA'nın altı örneğine sahip% 1'lik bir agaroz jeli. Her şeritte, 40S ve 60S alt birimlerini oluşturan iki parlak bant görülebilir, çünkü ribozomal RNA toplam RNA'nın ~% 95'ini oluşturur. Not: Hücre içi DMS tedavisi bazı RNA parçalanmasına ve bulaşmasına neden olur, ancak iki rRNA bandı hala görünür olmalıdır. Modifikasyon sonrası hafif parçalanma tolere edilir, çünkü metilasyon işaretini içeren bilgi üretilir ve hücreler hala hayattayken DMS inkübasyonu sırasında RNA yapısı hakkında rapor verir. (C) GeneRuler 1 kb artı merdiven DNA belirtecinin (şerit 1) %1'lik bir Agarose jeli, daha önce 6 ay boyunca -80 ° C'de depolanan toplam RNA (şerit 2) ve ribode tükenmiş RNA (şerit 3). RNA'yı birkaç donma-çözülme döngüsü ile uzun süre saklarken, RNA bozulmaya başlar ve muhtemelen sondalama deneyleri için kullanılmamalıdır. Ayrıca, toplam RNA'yı tükürdükten sonra, ribozomun 40S ve 60S alt birimlerini oluşturan iki parlak bant kaybolur ve artık RNA'ların bir yayması ortaya çıkmaya başlar. (D) GeneRuler 1 kb artı merdiven DNA belirtecinin (şerit 1) bir PAGE jeli ve tüm genom hazırlanmış RNA'nın bir kütüphane örneği. Jel, sıralama ihtiyaçlarına göre eksize edilmelidir. Her iki taraftan 150 döngüyü kapsayan çift uçlu bir sıralama çalışması için, jel 300 bp ile 500 bp arasında eksize edilmelidir. Adaptör dimerleri (170 bp'de çalışan) ayrılmalıdır. Kısaltmalar: DMS-MaP = Dimetil sülfat kullanarak dizileme ile mutasyonel profilleme; dpi = enfeksiyondan sonraki günler; CPE = sitopatik etki. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
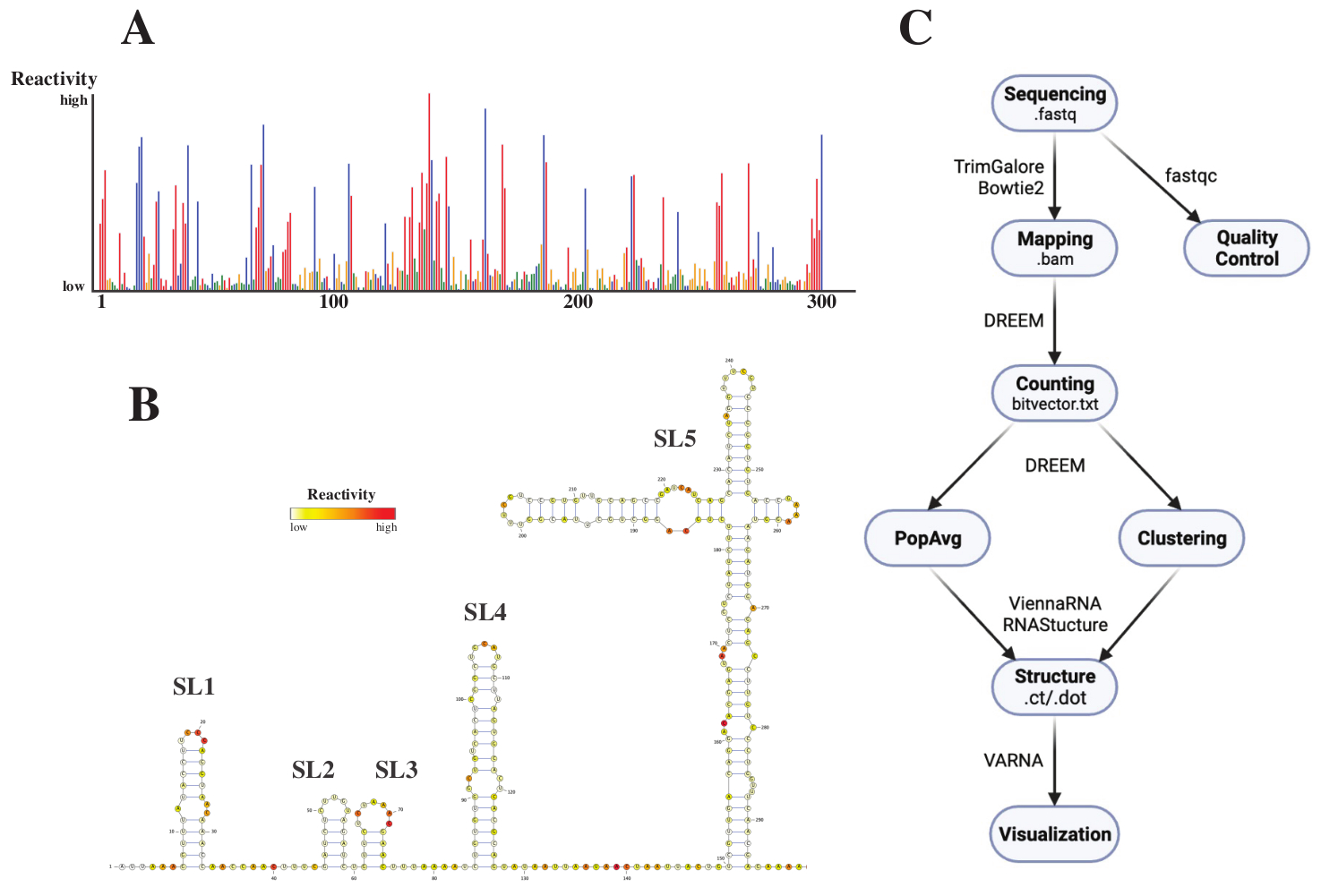
Sıralamadan sonra, .fastq dosyaları bir .fasta başvuru dosyasıyla birlikte DREEM web sunucusuna (http://rnadreem.org/) bir iş gönderilerek analiz edildi. Sunucu tarafından oluşturulan çıktı, fastqc (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) ve TrimGalore (https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/) tarafından oluşturulan kalite kontrol dosyalarının yanı sıra popülasyon ortalama mutasyon frekanslarını içeren diğer çıktı dosyalarını içerir. Mutasyon frekanslarını etkileşimli bir .html (bakınız Şekil 4A) formatıyla gösteren diyagram ve baz başına ham reaktivitleri içeren bir .csv dosyası ve birkaç RNA yapı tahmin yazılımı tarafından okunabilen bir struct_constraint.txt dosyası dışında, bu aynı zamanda okunan mutasyonlar hakkında rapor veren bir bitvector.txt dosyası da içerir. Bunlardan, .fasta ve struct_constraint.txt dosyaları RNAfold web sunucusuna (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi) gönderilerek popülasyon ortalama yapıları hesaplandı. Bu, çevrimiçi olarak görüntülenebilen veya ct veya Viyana formatında indirilebilen minimum serbest enerjiye dayalı yapı tahminleri üretmek için ViennaRNA yazılımını kullanır. RNA yapı modelleri oluşturmak için, bu indirilebilir dosyalar VARNA'ya gönderildi (https://varna.lri.fr/, bkz. Şekil 4B). Son olarak, bitvector.txt dosyaları, alternatif RNA konformasyonlarını aramak için DREEM'in kararlı sürümü (https://codeocean.com/capsule/6175523/tree/v1) tarafından kullanılabilir. DREEM kullanarak iyi yapı modelleri elde etmek için, taban başına 10.000 okuma kapsamı elde edilmelidir; kümeleme için, taban başına en fazla 100.000 okuma gerekebilir. Tüm iş akışına genel bir bakış Şekil 4C'de bulunabilir.

Şekil 4: SARS-CoV2 5'UTR. (A) SARS-CoV2 genomunun ilk 300 bazının baza göre renklendirilmiş reaktivite profilinin kimyasal sondalama deneylerinden elde edilen örnek veriler (A: kırmızı, C: mavi, U: yeşil, G: sarı). Ham reaktiviteler, mutlak mutasyon frekansının kapsama alanına bölünmesiyle hesaplanır. Açık konformasyonlu bazlar yüksek reaktivite değerlerine sahiptir; baz eşleştirme yapan bazlar düşük reaktivite değerlerine sahiptir. siz ve G, DMS tarafından değiştirilmez ve polimeraz sadakatsizliğinden kaynaklanan düşük reaktivite değerlerine sahiptir. Tahminler DREEM web sunucusu ile yapıldı. (B) SARS-CoV2 5'UTR'nin VARNA ile yapılan reaktivite değerlerinden tahmin edilen yapı modeli. Reaktivite değerleri yüksek bazlar kırmızı renktedir; reaktivite değerleri düşük bazlar beyaz renktedir. (C) DMS-MaP analizinin sıralamadan elde edilen .fastq dosyalarıyla başlayan iş akışı. Bunlar fastqc kullanılarak kalite kontrollü olabilir; adaptör dizileri TrimGalore kullanılarak kesilir ve ardından Bowtie2 kullanılarak bir referans dizisine geri eşlenir. Elde edilen .bam dosyalarından, DREEM her okumadaki mutasyonları sayar, bir mutasyon haritası veya .bitvector.txt dosyası oluşturur. Bunlar, her okunanın mutasyonlarını, popülasyon ortalama reaktivite profillerinin oluşturulabileceği konuma bağlı bir şekilde rapor eder. Alternatif olarak, bitvektörler alternatif RNA konformasyonlarını aramak için DREEM kullanılarak kümelenebilir. Son olarak, elde edilen yapı modelleri yazılım (örneğin, VARNA) kullanılarak görselleştirilir. Kısaltmalar: DMS-MaP = Dimetil sülfat kullanarak dizileme ile mutasyonel profilleme; SARS-CoV2 = şiddetli akut solunum sendromu-koronavirüs 2. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
Buradaki protokol, DMS mutasyonel profilleme deneylerini kullanarak RNA'nın in vitro ve hücrelerde nasıl araştırılacağını açıklamaktadır. Ayrıca, genlere özgü veriler oluşturmak ve elde edilen .fastq dosyalarını analiz etmek için Illumina dizilimi için kütüphanelerin nasıl hazırlanacağına dair talimatlar verir. Ek olarak, genom çapında kütüphane yaklaşımları kullanılabilir. Bununla birlikte, gene özgü RT-PCR en yüksek kalitede ve en sağlam verileri üretir. Bu nedenle, örnekler arasında karşılaştırma yapılırsa, kütüphane oluşturma bazı önyargılara neden olduğundan, aynı sıralama stratejileriyle hazırlandıklarından emin olmak önemlidir. Tekrarlanabilirlik her zaman replikalar kullanılarak ölçülmelidir.
Çeşitli önlemler
RNA, hem yüksek sıcaklıklarda hem de RNazlar tarafından bozulmaya duyarlı kararsız bir moleküldür. Bu nedenle, özel önlemler - kişisel koruyucu ekipman (KKD), RNAse içermeyen malzeme ve RNAse inhibitörlerinin kullanılması - tavsiye edilir. En önemlisi, RNA mümkün olduğunca buz üzerinde tutulmalıdır. Bu özellikle yüksek sıcaklıklara karşı daha hassas olan metillenmiş RNA için geçerlidir.
İlgili RNA yapısının DMS konsantrasyonuna ve tampon koşullarına duyarlı olmadığını doğrulamak önemlidir. pH 7-7.5'te 100 mM Tris, 100 mM MOPS ve 100 mM HEPES gibi tamponlar yüksek sinyal verir, ancak reaksiyon sırasında pH'ı korumak için yeterli olmayabilir21. DMS, pH'ı azaltan suda hidrolize olduğundan, modifikasyon reaksiyonu sırasında nötr bir pH'ı korumak için güçlü bir tampon kritik öneme sahiptir. Bisin ilavesinin pH'ın biraz bazik21 olarak korunmasına yardımcı olduğu gösterilmiştir, ancak Gs ve Us üzerinde düşük DMS modifikasyonu ile sonuçlanır, bu da bilgilendirici olabilir, ancak As ve C'den çok daha düşük bir sinyalin üretilmesi nedeniyle ayrı ayrı analiz edilmelidir ve bu protokolde daha fazla tartışılmamıştır.
Gen-spesifik RT-PCR'de, modifiye RNA DNA'ya ters transkribe edilir ve PCR ile fragmanlar halinde çoğaltılır. RNA'nın boyutu teorik olarak sınırsız olabilirken, bu PCR fragmanları, ters transkripsiyon reaksiyonu sırasında önyargıyı önlemek için 400-500 baz çifti (bp) uzunluğunu geçmemelidir. İdeal olarak, parçalar sıralama çalışması kapsamında olmalıdır (yani, sıralama 150 x 150 döngülü çift uçlu bir sıralama programı kullanılarak gerçekleştirilirse, tek bir parça 300 bp'yi geçmemelidir). Daha az döngüye sahip sıralama programları kullanıldığında, PCR ürünleri bir dsDNase kullanılarak parçalanabilir. Ayrıca, primer dizilerdeki diziler herhangi bir yapısal bilgi içermediğinden, sondalanan RNA >1 fragmanından oluştuğunda fragmanlar üst üste binmelidir. RT reaksiyonları, farklı parçalar için birden fazla RT primeri içerebilir (10 farklı RT primerine kadar). Dizilere bağlı olarak, RT astarlarının bir araya getirilmesi, ters transkripsiyonu daha az verimli hale getirebilir, ancak tipik olarak iyi çalışır. Her PCR reaksiyonu ayrı ayrı yapılmalıdır.
RNA'yı DMS ile araştırırken, deneysel koşullar ek bir rol oynar, çünkü birçok RNA termodinamik olarak kararsızdır ve sıcaklık gibi çevresel faktörlere bağlı olarak konformasyonlarını değiştirir. Düzensizlikleri önlemek için, deney koşulları, reaksiyon sürelerine göre de mümkün olduğunca sabit tutulmalıdır. RNA24'ün düzgün bir şekilde katlanmasını sağlamak için tampon koşulları, temel koşullar korunduğunda - tamponlama kapasitesi ve monovalent (Na) ve iki değerli iyonların (Mg) varlığı - belirlibir dereceye kadar değiştirilebilir görünmektedir.
Modifiye RNA'ların kütüphane hazırlığı ile ilgili olarak, çeşitli yönler dikkate alınmalıdır. İlk olarak, daha önce de belirtildiği gibi, modifiye RNA'lar değiştirilmemiş muadillerinden daha az kararlıdır, yani optimal parça boyutu dağılımı için parçalanma sürelerinin optimizasyonunu gerektirebilirler. Ayrıca, bazı RNA kütüphanesi hazırlama kitleri ve diğer birçok RNAseq yaklaşımı, ters transkripsiyon kitinde rastgele primerler kullanır. Bu, özellikle bir genin 3'ünde referansın daha düşük kapsama alanına ve nihayetinde yetersiz kapsama derinliğine yol açabilir. Belirli bir bölgenin kapsama alanı çok düşükse, bu üsleri yapı tahmininden çıkarmak gerekebilir. RT-PCR ve tüm genom RNAseq kitleri dışında, diğer kütüphane hazırlama yaklaşımları da kullanılabilir. RNA'ya 3' ve / veya 5' adaptörlerin bağlanmasını içeren protokoller, RNA'nın küçük parçaları kullanıldığında veya primer bölgelerdeki problama bilgisinin kaybından kaçınılması gerektiğinde avantajlıdır.
Son olarak, kimyasal sondalama deneylerinin analizi her zaman dikkatli bir şekilde yorumlanmalıdır. Şu anda, herhangi bir RNA'nın RNA yapısını tek başına diziden yüksek doğrulukla tahmin eden bir yazılım yoktur. Kimyasal problama kısıtlamaları doğruluğu büyük ölçüde artırsa da, uzun RNA'lar (>500 nt) için iyi modeller üretmek hala zordur. Bu modeller diğer yaklaşımlar ve / veya mutagenez ile daha fazla test edilmelidir. RNA tahmin yazılımı, maksimum baz çifti sayısını optimize eder, böylece RNA katlanmasını doğru bir şekilde temsil etmeyebilecek açık konformasyonları önemli ölçüde cezalandırır5. Bu nedenle, elde edilen yapı modeli, Lan ve ark.20 tarafından örneklendiği gibi, altta yatan kimyasal problama verileriyle (örneğin, AUROC tarafından) ve replikasyonlar arasında (örneğin, mFMI tarafından) tahmin anlaşmasını ölçerek test edilmelidir.
İdeal olarak, elde edilen yapı modeline meydan okumak için farklı sistemlerde birkaç deney, kişinin hipotezini güçlendirmek için kullanılmalıdır. Bunlar, in vitro ve hücre içi yaklaşımların, telafi edici mutasyonların ve farklı hücre hatlarının ve türlerinin kullanımını içerebilir. Dahası, ham reaktiviteler genellikle RNA katlanır topluluğunun "zemin gerçeği" anlık görüntüsünü kaydettikleri için yapı tahminleri kadar veya daha bilgilendiricidir. Bu nedenle, ham reaktifler, farklı koşullar arasındaki yapı değişikliklerini karşılaştırmak için çok uygun ve bilgilendiricidir. Daha da önemlisi, hesaplamalı tahmin ile kimyasal problama kısıtlamaları kullanılarak hesaplanan en düşük serbest enerji yapıları, yalnızca tam bir yapı modeline doğru bir başlangıç hipotezi olarak kullanılmalıdır.
Açıklamalar
Yazarların beyan edecekleri herhangi bir çıkar çatışması yoktur.
Teşekkürler
Hiç kimse
Malzemeler
| Name | Company | Catalog Number | Comments |
| 1 Kb Plus DNA Ladder | 10787018 | Thermo | |
| 2-mercaptoethanol | M6250-250ML | Sigma | |
| Acid-Phenol:Chloroform, pH 4.5 | AM9720 | Thermo | |
| Advantage PCR | 639206 | Takara | |
| CloneAmp HiFi PCR Premix | 639298 | Takara | |
| DMS | D186309 | Sigma | |
| dNTPs 10 mM each | U151B | Promega | |
| E-Gel EX Agarose Gels, 2% | G402022 | Thermo | precast agarose gels |
| Ethanol (200 proof) | E7023-4X4L | Sigma | |
| Falcon tubes, 15 mL, 50 mL | |||
| GlycoBlue | co-precipitant | ||
| HCT-8 cells | ATCC #CCL-244 | ||
| Invitrogen MgCl2 (1 M) | AM9530G | fisherscientific | |
| Isopropanol | 278475 | Sigma | |
| Megascript T7 transcription | AM1334 | Thermo | |
| NanoDrop spectrophotometer | |||
| Novex TBE Gels, 8%, 10 well | EC6215BOX | Thermo | |
| OC43 | ATCC #VR-1558 | ||
| RiboRuler Low Range RNA Ladder | SM1831 | Thermo | |
| RNAse H | M0297L | NEB | |
| Sodium Cacodylate, 0.4 M, pH 7.2 | 102090-964 | VWR | |
| Sodium hydroxide solution | S8263-150ML | Sigma | |
| SuperScript II Reverse Transcriptase for FSB and DTT | 18064014 | Thermo | |
| TGIRT-III Enzyme | TGIRT50 | Ingex | |
| The Oligo Clean & Concentrator | D4060 | Genesee | |
| The RNA Clean & Concentrator kits are RNA clean up kits | R1016 | Genesee | |
| TRIzol Reagents | 15596018 | Thermo | RNA isolation reagent |
| Water, (For RNA Work) (DEPC-Treated, DNASE, RNASE free/Mol. Biol.) | BP561-1 | fisherscientific | |
| xGen Broad-range RNA Library Prep 16rxn | 10009865 | IDT | |
| Zymo RNA clean and concentrator columns |
Referanslar
- Kim, S. H., et al. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA. Science. 185 (4149), 435-440 (1974).
- Robertus, J. D., et al. Structure of yeast phenylalanine tRNA at 3 Å resolution. Nature. 250 (467), 546-551 (1974).
- Zaug, A. J., Cech, T. R. In vitro splicing of the ribosomal RNA precursor in nuclei of Tetrahymena. Cell. 19 (2), 331-338 (1980).
- Zhao, Y., et al. NONCODE 2016: An informative and valuable data source of long non-coding RNAs. Nucleic Acids Research. 44, D203-D208 (2016).
- Vandivier, L. E., Anderson, S. J., Foley, S. W., Gregory, B. D. The conservation and function of RNA secondary structure in plants. Annual Review of Plant Biology. 67, 463 (2016).
- Jumper, J., et al. Highly accurate protein structure prediction with AlphaFold. Nature. 596 (7873), 583-589 (2021).
- Das, R. RNA structure: A renaissance begins. Nature Methods. 18 (5), 439-439 (2021).
- Smola, M. J., Rice, G. M., Busan, S., Siegfried, N. A., Weeks, K. M. Selective 2′-hydroxyl acylation analyzed by primer extension and mutational profiling (SHAPE-MaP) for direct, versatile and accurate RNA structure analysis. Nature Protocols. 10 (11), 1643-1669 (2015).
- Mathews, D. H., et al. Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure. Proceedings of the National Academy of Sciences of the United States of America. 101 (19), 7287-7292 (2004).
- Zuker, M., Stiegler, P. Optimal computer folding of large RNA sequences using thermodynamics and auxiliary information. Nucleic Acids Research. 9 (1), 133-148 (1981).
- Lorenz, R., et al. ViennaRNA Package 2.0. Algorithms for Molecular Biology. 6, (2011).
- Reuter, J. S., Mathews, D. H. RNAstructure: Software for RNA secondary structure prediction and analysis. BMC Bioinformatics. 11, (2010).
- Wells, S. E., Hughes, J. M. X., Igel, A. H., Ares, M. Use of dimethyl sulfate to probe RNA structure in vivo. Methods in Enzymology. , 479-493 (2000).
- Tomezsko, P. J., et al. Determination of RNA structural diversity and its role in HIV-1 RNA splicing. Nature. 582 (7812), (2020).
- Zubradt, M., et al. DMS-MaPseq for genome-wide or targeted RNA structure probing in vivo. Nature Methods. 14 (1), (2017).
- Woodson, S. A. Compact intermediates in RNA folding. Annual Reviews in Biophysics. 39, (2010).
- Morandi, E., et al. Genome-scale deconvolution of RNA structure ensembles. Nature Methods. 18 (3), 249-252 (2021).
- Olson, S. W., et al. Discovery of a large-scale, cell-state-responsive allosteric switch in the 7SK RNA using DANCE-MaP. Molecular Cell. 82 (9), 1708-1723 (2022).
- Incarnato, D., Morandi, E., Simon, L. M., Oliviero, S. RNA Framework: An all-in-one toolkit for the analysis of RNA structures and post-transcriptional modifications. Nucleic Acids Research. 46 (16), (2018).
- Lan, T. C. T., et al. Secondary structural ensembles of the SARS-CoV-2 RNA genome in infected cells. Nature Communications. 13 (1), 1128 (2022).
- Homan, P. J., et al. Single-molecule correlated chemical probing of RNA. Proceedings of the National Academy of Sciences of the United States of America. 111 (38), 13858-13863 (2014).
- Yang, S. L., et al. Comprehensive mapping of SARS-CoV-2 interactions in vivo reveals functional virus-host interactions. Nature Communications. 12 (1), 5113 (2021).
- Manfredonia, I., et al. Genome-wide mapping of SARS-CoV-2 RNA structures identifies therapeutically-relevant elements. Nucleic Acids Research. 48 (22), 12436-12452 (2020).
- Fischer, N. M., Polěto, M. D., Steuer, J., vander Spoel, D. Influence of Na+ and Mg2+ ions on RNA structures studied with molecular dynamics simulations. Nucleic Acids Research. 46 (10), 4872-4882 (2018).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır