
Method Article
Зондирование структуры РНК с мутационным профилированием диметилсульфата с секвенированием in vitro и в клетках
В этой статье
Резюме
Протокол содержит инструкцию по модификации РНК диметилсульфатом для экспериментов по мутационному профилированию. Он включает в себя зондирование in vitro и in vivo с двумя альтернативными методами подготовки библиотеки.
Аннотация
Роль структуры РНК практически в любом биологическом процессе становится все более очевидной, особенно в последнее десятилетие. Однако классические подходы к решению структуры РНК, такие как кристаллография РНК или крио-ЭМ, не смогли угнаться за быстро развивающейся областью и потребностью в высокопроизводительных решениях. Мутационное профилирование с секвенированием с использованием диметилсульфата (DMS) MaPseq представляет собой основанный на секвенировании подход к выводу структуры РНК из реакционной способности основания с DMS. DMS метилирует азот N1 в аденозинах и N3 в цитозинах на грани Уотсона-Крика, когда основание непарно. Обратная транскрибирование модифицированной РНК термостабильной интронной обратной транскриптазой II группы (TGIRT-III) приводит к включению метилированных оснований в качестве мутаций в кДНК. При секвенировании полученной кДНК и сопоставлении ее с эталонной транскриптом относительные частоты мутаций для каждого основания указывают на «статус» основания как парного или непарного. Несмотря на то, что реактивность DMS имеет высокое отношение сигнал/шум как in vitro , так и в клетках, этот метод чувствителен к смещению в процедурах обработки. Чтобы уменьшить это смещение, в этой статье представлен протокол лечения РНК СДМ в клетках и транскрибированной РНК in vitro .
Введение
С момента открытия того, что РНК обладает как структурными 1,2, так и каталитическимисвойствами 3, важность РНК и ее регуляторной функции во множестве биологических процессов была постепенно раскрыта. Действительно, влияние структуры РНК на регуляцию генов привлекло все большее внимание4. Как и белки, РНК имеет первичные, вторичные и третичные структуры, ссылаясь на последовательность нуклеотидов, 2D-картирование парных взаимодействий оснований и 3D-сворачивание этих парных структур оснований, соответственно. В то время как определение третичной структуры является ключом к пониманию точных механизмов, лежащих в основе РНК-зависимых процессов, вторичная структура также очень информативна в отношении функции РНК и является основой для дальнейшего 3D-сворачивания5.
Тем не менее, определение структуры РНК было по своей сути сложной задачей с традиционными подходами. В то время как для белков кристаллография, ядерный магнитный резонанс (ЯМР) и криогенная электронная микроскопия (крио-ЭМ) позволили определить разнообразие структурных мотивов, что позволяет прогнозировать структуру только по последовательности6, эти подходы не широко применимы к РНК. Действительно, РНК представляют собой гибкие молекулы со строительными блоками (нуклеотидами), которые имеют гораздо большую конформационную и вращательную свободу по сравнению с их аминокислотными аналогами. Кроме того, взаимодействия через спаривание оснований более динамичны и универсальны, чем взаимодействия аминокислотных остатков. В результате классические подходы оказались успешными только для относительно небольших РНК с четко определенными, высококомпактными структурами7.
Другой подход к определению структуры РНК заключается в химическом зондировании в сочетании с секвенированием следующего поколения (NGS). Эта стратегия генерирует информацию о статусе связывания каждого основания в последовательности РНК (т.е. его вторичной структуре). Короче говоря, основания в молекуле РНК, которые не участвуют в спаривании оснований, дифференциально модифицируются небольшими химическими соединениями. Обратная транскрибирование этих РНК со специализированными обратными транскриптазами (RT) включает модификации в комплементарную дезоксирибонуклеиновую кислоту (кДНК) в качестве мутаций. Эти молекулы кДНК затем амплифицируются полимеразной цепной реакцией (ПЦР) и секвенируются. Чтобы получить информацию об их «статусе» как связанном или несвязанном, частоты мутаций на каждом основании в интересующей РНК вычисляются и вводятся в программное обеспечение для прогнозирования структуры в качестве ограничений8. Основываясь на правилахближайшего соседа 9 и расчетах минимальной свободной энергии10, это программное обеспечение генерирует структурные модели, которые наилучшим образом соответствуют полученным экспериментальным данным11,12.
DMS-MaPseq использует DMS, который метилирует азот N1 в аденозинах и азот N3 в цитозинах на их поверхности Watson-Crick оченьспецифическим образом 13. Использование термостабильной интронной обратной транскриптазы II группы (TGIRT-III) в обратной транскрипции создает мутационные профили с беспрецедентным отношением сигнал/шум, даже допуская деконволюцию перекрывающихся профилей, генерируемых двумя или более альтернативными конформациями14,15. Кроме того, DMS может проникать в клеточные мембраны и целые ткани, что делает возможным зондирование в физиологических контекстах. Однако получение данных хорошего качества является сложной задачей, поскольку различия в процедуре обработки могут повлиять на результаты. Поэтому мы предоставляем подробный протокол как для in vitro, так и для внутриклеточного DMS-MaPseq, чтобы уменьшить предвзятость и направить новичков в метод через трудности, с которыми они могут столкнуться. Особенно в свете недавней пандемии SARS-CoV2 высококачественные данные о РНК-вирусах являются важным инструментом для изучения экспрессии генов и поиска возможных терапевтических средств.
протокол
ПРИМЕЧАНИЕ: Смотрите Таблицу материалов для получения подробной информации , связанной со всеми материалами, программным обеспечением, реагентами, инструментами и ячейками, используемыми в этом протоколе.
1. Ген-специфический in vitro DMS-MaP
- Транскрипция РНК in vitro
- Получить последовательность интересующей РНК в виде двухцепочечной (ds)ДНК (например, в виде фрагментов ДНК, плазмид или ПЦР из ранее существовавшей/геномной ДНК). Если последовательность ДНК содержит промотор полимеразы, перейдите к шагу 3.
- Выполняют перекрывающуюся ПЦР для присоединения промотора РНК-полимеразы перед желаемым фрагментом ДНК (форвардный праймер для Т7-полимеразы: 5' TAATACGACTCACTATAGG + первые основания целевой последовательности 3').
- In vitro транскрибируют фрагмент ДНК в РНК. Всегда держите РНК на льду.
- Переваривайте ДНК с помощью ДНКазы.
- Изолируют РНК с помощью столбчатого подхода (стадия 2.4) или путем осаждения этанола (стадия 2.5). Элюют в соответствующем объеме, ожидая выхода ~50 мкг.
- Обеспечить целостность РНК, запустив ее на агарозный гель; денатурировать РНК в течение 2-3 мин при 70 °C перед запуском.
ПРИМЕЧАНИЕ: Буфер и агароза могут содержать РНКазы, которые разлагают РНК и могут загрязнять образец РНК. Сборные агарозные гели ранее использовались в этой лаборатории; результаты (особенно с РНК) временами были неоднозначными. Наилучшие результаты были получены при использовании гелей агарозы или ПЕЙДЖ. - Непосредственное использование хранилища РНК при −80 °C в течение нескольких месяцев, если деградация не видна после оттаивания.
- In vitro Модификация DMS (при 105 мМ DMS)
- Готовят достаточное количество буфера переворачивания (0,4 М какодилата натрия, рН 7,2, содержащего 6 мМMgCl2).
ПРИМЕЧАНИЕ: Для каждой реакции (конечный объем 100 мкл) добавляют 89 мкл буфера повторного складывания. - Для каждой реакции передавайте 89 мкл буфера повторного складывания в специальную трубку объемом 1,5 мл и предварительное нагревание при 37 °C в термоколочебой, помещенном под химический капот.
ПРИМЕЧАНИЕ: DMS очень токсичен и всегда должен храниться под химическим капотом до закалки восстановителем. - Элют 1-10 пмоль РНК в 10 мкл безнуклеазной воды (NFH2O); перевести в ПЦР-трубку.
- Инкубируют в термоциклере при 95 °C в течение 1 мин для денатурации РНК.
- Поместите на ледяную глыбу немедленно , чтобы избежать неправильного складывания.
- Добавьте образец РНК в назначенную пробирку с буфером рефолдирования при 37 °C, хорошо перемешайте и инкубируйте в течение 10-20 мин для повторного складывания РНК.
ПРИМЕЧАНИЕ: Большинство РНК складываются в порядке миллисекунд до секунд, хотя существует16 исключений. - Добавьте 1 мкл 100% (10,5 М) DMS к образцу РНК и инкубируйте в течение 5 мин при встряхивании со скоростью 800-1 400 оборотов в минуту (об/мин).
ПРИМЕЧАНИЕ: Встряхивание (или другие средства смешивания) на этом этапе имеет решающее значение, поскольку DMS является гидрофобным и может не полностью растворяться в буфере рефолдирования. Отклонения во времени реакции могут повлиять на воспроизводимость реактивности DMS. Для сведения к минимуму погрешности дозирования ДМС может быть растворена в 100% этаноле перед добавлением его в образец, если поддерживается конечная концентрация 1% (105 мМ) ДМС. Для необработанного контроля DMS может быть заменена диметилсульфоксидом (DMSO) или водой. - Через 5 мин времени реакции закаляют 60 мкл 100% β-меркаптоэтанола (BME), хорошо перемешивают и сразу же помещают РНК на лед.
ПРИМЕЧАНИЕ: РНК может быть безопасно удалена из капота после гашения реакции с BME, чтобы очистить ее. Тем не менее, прямого воздействия BME на окружающую среду все же следует избегать из-за его сильного запаха и раздражающих свойств. - Очищают РНК путем осаждения ацетата-этанола натрия (см. этап 2.5) или колоночного подхода (см. шаг 2.6) и элюируют в 10 мкл воды.
- Количественно оцените РНК с помощью спектрофотометра.
- Непосредственное использование хранилища модифицированной РНК при −80 °C.
ПРИМЕЧАНИЕ: Следует избегать длительного хранения, так как РНК менее стабильна после лечения ДМС.
- Готовят достаточное количество буфера переворачивания (0,4 М какодилата натрия, рН 7,2, содержащего 6 мМMgCl2).
- Геноспецифическая ОТ-ПЦР модифицированной РНК
ПРИМЕЧАНИЕ: См. рисунок 1 для настройки ОТ-ПЦР фрагментов, обработанных DMS.- Elute 100 нг модифицированной РНК в 10 мкл безнуклеазного (NF)H2O. Перенос в трубку ПЦР.
- В пробирку добавляют 4 мкл 5-кратного буфера первой нити (FSB), 1 мкл смеси dNTP (по 10 мМ), 1 мкл 0,1 М дитиотрейтола (DTT) (избегайте циклов замораживания-оттаивания), 1 мкл ингибитора РНКАЗЫ, 1 мкл обратной грунтовки 10 мкМ (одинарный праймер или пул праймеров) и 1 мкл TGIRT III.
ПРИМЕЧАНИЕ: Для пула праймеров не добавляйте 1 мкл 10 мкМ каждой грунтовки непосредственно к RT; вместо этого сначала смешайте грунтовки и добавьте 1 мкл из смеси (при общей концентрации грунтовки 10 мкМ). - Инкубировать при 57 °C в течение 30-1,5 ч (как правило, 30 мин достаточно для получения продукта 500 нт) в термоциклере.
- Добавьте 1 мкл 4 M NaOH, перемешайте путем пипетки и инкубируйте при 95 °C в течение 3 мин для разложения РНК.
ПРИМЕЧАНИЕ: Этот шаг имеет решающее значение, поскольку он высвобождает TGIRT из кДНК путем деградации РНК. При пропуске может быть затронута последующая ПЦР. - Очистка с использованием колоночного подхода (см. шаг 2.6), который в достаточной степени удаляет грунтовки и элюирует в 10 мкл NFH2O.
- ПЦР-амплифицируют кДНК, используя 1 мкл продукта обратной транскрипции на 25 мкл реакции с помощью набора ПЦР, предназначенного для балансировки выхода и точности.
ПРИМЕЧАНИЕ: Грунтовки должны иметь температуру плавления ~60 °C. - Запустите 2 мкл продукта ПЦР на агарозном геле или сборном агарозном геле, чтобы убедиться в успехе ПЦР.
- В идеале после ПЦР должна показываться только одна полоса. Если это так, очистите реакцию, используя подход на основе столбцов. Если присутствуют альтернативные полосы, используйте оставшуюся реакцию ПЦР, чтобы иссечь правильную полосу из геля. Элюет в достаточно малом объеме (например, 10 мкл).
- Количественно оцените извлеченные фрагменты с помощью спектрофотометра.
- Индексируйте фрагменты dsDNA для секвенирования, используя подход, подходящий для требуемой платформы секвенирования.

Рисунок 1: Экспериментальная установка для ОТ-ПЦР больших фрагментов, обработанных ДМС. При выполнении обратной транскрипции на модифицированной РНК модификации последовательности, к которой отжигается праймер, регистрироваться не будут. Таким образом, когда фрагменты превышают 400-500 bp в длину, фрагменты, перекрывающиеся в областях грунтовки, должны быть спроектированы, как показано здесь. Длина фрагментов зависит от потребностей в последовательности. При использовании парной последовательности 150 циклов фрагменты не должны превышать 300 бит в секунду. Сокращения: ОТ-ПЦР = полимеразная цепная реакция с обратной транскрипцией; DMS = диметилсульфат. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
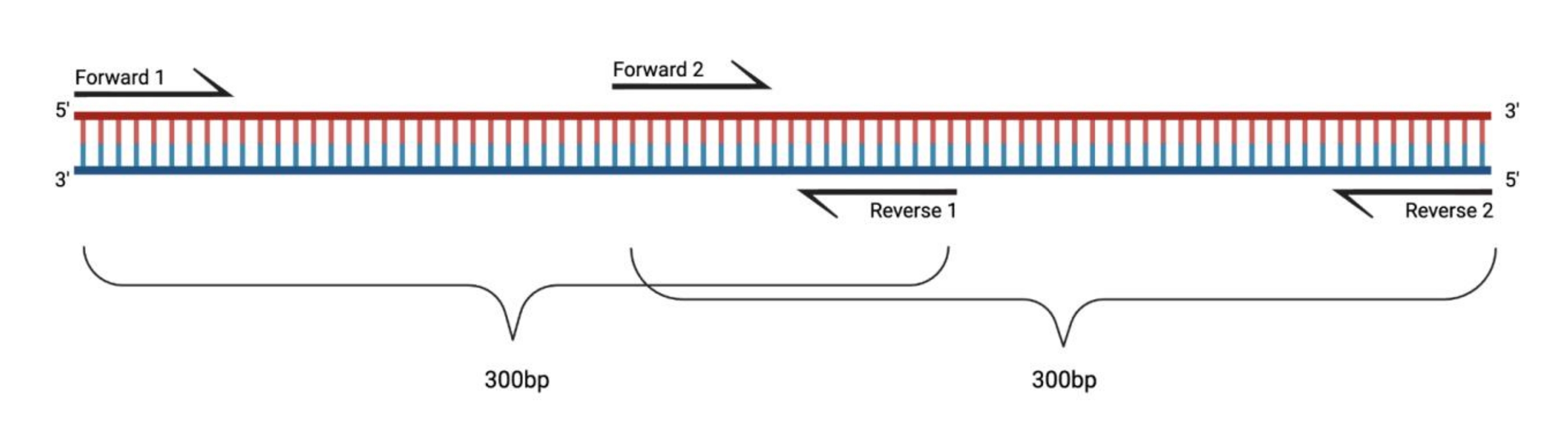{kind=link}
2. Полногеномный DMS-MaP с использованием инфицированных вирусом клеток
ПРИМЕЧАНИЕ: В клетках лечение ДМС также может сочетаться с генно-специфическим подходом амплификации, описанным выше. Библиотека всего генома требует огромной глубины секвенирования для достижения полного охвата одного гена. Однако, если вирусные РНК составляют значительную долю рибоделированной РНК после экстракции, секвенирование всего генома будет целесообразным. Кроме того, другие методы обогащения могут быть объединены с методом генерации библиотеки всего генома.
- Лечение ДМС
- Выращивают клетки, инфицированные вирусом, до нужной стадии заражения.
- Переместите контейнер для клеток в специальный вытяжной шкаф, который подходит для обработки как вирусов на требуемом уровне биобезопасности, так и химических паров, генерируемых такими агентами, как DMS.
- Добавьте 2,5% объема DMS к питательной среде и запечатайте контейнер (обычно пластину 10 см) парапленкой.
ПРИМЕЧАНИЕ: С помощью DMS легко недо-модифицировать и чрезмерно модифицировать. При добавлении DMS непосредственно в клетки очень важно хорошо перемешать. В качестве альтернативы, предварительно разогрейте новую среду в конической трубке объемом 50 мл при 37 °C и добавьте DMS непосредственно энергично встряхивать. Декантируйте отработанную среду на клетках и медленно пипетку в ДМС-содержащую среду. - Переложить в инкубатор при температуре 37 °C в течение 5 мин.
ПРИМЕЧАНИЕ: В зависимости от количества времени, необходимого для обработки DMS вне инкубатора, возможно, что 5 минут приведут к чрезмерной модификации. Не теряйте время от добавления DMS к инкубации до ≤1 мин. Если эксперимент проводится в первый раз, рекомендуется провести титрование DMS и варьировать время инкубации (от 3 до 10 минут), чтобы найти оптимальную скорость модификации и убедиться, что результаты являются надежными в окне концентраций. - Осторожно вылейте ДМС-содержащую среду (в соответствующие химические отходы) и осторожно добавьте 10 мл стоп-буфера (PBS с 30% BME [например, 3 мл BME и 7 мл PBS]).
ПРИМЕЧАНИЕ: Добавление DMS и BME может поднять клетки с пластины, если клетки не сильно адгезивны. Если клетки поднимаются, их можно рассматривать как суспензионные клетки - вместо удаления среды, содержащей DMS, добавить буфер остановки напрямую и соскоблить клетки с DMS и BME в коническую трубку объемом 50 мл. Гранулирование клеток центрифугированием в течение 3 мин при 3000 × г; убедитесь, что вы избавились от любого остаточного DMS, который может гранулироваться под клетками в больших каплях. Дополнительный этап промывки в 30% BME рекомендуется, если среда DMS не может быть удалена изначально. - Соскоблите клетки и перенесите их в коническую трубку объемом 15 мл.
- Гранулы центрифугированием при 3000 × г в течение 3 мин.
- Удалите супернатант и промыть 2 раза 10 мл PBS.
- Осторожно удалите как можно больше остаточного PBS.
- Растворите гранулу в соответствующем количестве реагента, выделяющего РНК (например, 3 мл для колбы для культуры T75, 1 мл для пластины размером 10 см).
ПРИМЕЧАНИЕ: Недостаточное количество реагента может повлиять на выход РНК.
- Экстракция РНК и истощение рибосомальной РНК (рРНК)
- К 1 мл гомогенизированных клеток в реагенте выделения РНК добавляют 200 мкл хлороформа, вихрь в течение 15-20 с до ярко-розового цвета, а затем инкубируют до 3 мин до тех пор, пока не будет видно разделение фаз.
ПРИМЕЧАНИЕ: Розовая липидная фаза должна оседать внизу. Если это не так, то время вихря, вероятно, было недостаточным. - Вращение на максимальной скорости (~ 20 000 × г) в течение 15 мин при 4 °C.
- Перенесите верхнюю водную фазу в новую трубку.
- Очищают РНК путем осаждения ацетата-этанола натрия (см. этап 2.5) или колоночного подхода (см. шаг 2.6) и элюируют в достаточном объеме NFH2O.
- Проверьте целостность РНК на агарозном геле. Ищите две полосы, соответствующие двум рибосомным субъединицам.
- Истощают рРНК, используя предпочтительный подход, и элюируют в достаточном объеме (обычно 20-50 мкл) NFH2O.
ПРИМЕЧАНИЕ: Для последующих применений предлагается ~500 нг общей РНК в объеме 8 мкл. Нерибосомные РНК обычно составляют только 5%-10% от общей РНК. - Количественная оценка с помощью спектрофотометра.
- К 1 мл гомогенизированных клеток в реагенте выделения РНК добавляют 200 мкл хлороформа, вихрь в течение 15-20 с до ярко-розового цвета, а затем инкубируют до 3 мин до тех пор, пока не будет видно разделение фаз.
- Генерация библиотек
- Используйте генно-специфическую ОТ-ПЦР или другие подходы для создания библиотек15. Если для грунтования используются случайные гексамеры, добавьте стадию инкубации принизком T m (37-42 °C), чтобы обеспечить отжиг гексамеров.
ПРИМЕЧАНИЕ: Стандартные наборы генерации библиотек также могут быть использованы путем замены фермента RT на TGIRT и изменения температуры RT до 57 °C.
- Используйте генно-специфическую ОТ-ПЦР или другие подходы для создания библиотек15. Если для грунтования используются случайные гексамеры, добавьте стадию инкубации принизком T m (37-42 °C), чтобы обеспечить отжиг гексамеров.
- Очистка РНК на основе столбцов с использованием колонок RNA Clean & Concentrator
ПРИМЕЧАНИЕ: Все этапы должны проводиться при комнатной температуре.- Добавьте NFH2Oв пробирку, чтобы довести ее до объема 50 мкл.
- Добавьте к образцу 100 мкл связывающего буфера и 150 мкл 100% этанола.
- Перемешайте и переложите в отжимную колонну.
- Отжим при 10 000-16 000 × г в течение 30 с; отбросьте сквозное прохождение.
- Добавьте 400 мкл буфера подготовки РНК.
- Открутка при 10 000-16 000 × г в течение 30 с; отбросьте сквозное прохождение.
- Добавьте 700 мкл буфера промывки РНК.
- Отжим при 10 000-16 000 × г в течение 30 с; отбросьте сквозное прохождение.
- Добавьте 400 мкл РНК промывочного буфера.
- Открутка при 10 000-16 000 × г в течение 30 с; отбросьте сквозное прохождение.
- (Необязательно) Переложите колонну на новую коллекционную трубку и вращайте на 10 000-16 000 × г в течение 2 мин.
- Переложите колонку в чистую трубку, не содержащую РНК, и добавьте соответствующее количество NFH2O.
- Открутите при 10 000-16 000 × г в течение 1 мин.
- Кислотная фенол-хлороформная экстракция РНК.
- Добавьте равный объем кислотного фенола:хлороформа:изоамилового спирта.
- Тщательно вихрь и центрифуга при 14 000 × г в течение 5 мин.
- Если фазовое разделение отсутствует, добавляют 20 мкл 2 М NaCl и повторяют центрифугирование.
- Переведите водную фазу в новую трубку.
- Добавьте 500 мкл изопропанола и 2 мкл соосаждателя.
- Перемешать и инкубировать при RT в течение 3 мин; затем инкубировать при температуре −80 °C в течение ночи.
- Гранулирование РНК центрифугированием на максимальной скорости (~ 20 000 × г) в течение 30 мин при 4 °С.
- Промыть гранулу 200 мкл ледяного 70% этанола.
- Вращение на максимальной скорости (~ 20 000 × g) в течение 5 мин; отбросьте сквозное прохождение.
- Повторно суспендируют гранулу в соответствующем количестве НФH2O.
- Очистка кДНК на основе столбцов с помощью столбцов Oligo Clean и Concentrator
ПРИМЕЧАНИЕ: Все этапы должны проводиться при комнатной температуре.- Добавьте NFH2Oв пробирку, чтобы довести ее до объема 50 мкл.
- Добавьте 100 мкл связывающего буфера и 400 мкл 100% этанола.
- Перемешайте и переложите в отжимную колонну.
- Отжим при 10 000-16 000 × г в течение 30 с; отбросьте сквозное прохождение.
- Добавьте 750 мкл буфера промывки ДНК.
- Открутка при 10 000-16 000 × г в течение 30 с; отбросьте сквозное прохождение.
- (Необязательно) Переведите колонну в новую сборную трубку и открутите при 10 000-16 000 × г в течение 2 мин.
- Переложите колонку в чистую трубку, не содержащую РНК, и добавьте соответствующее количество NFH2O.
- Открутите при 10 000-16 000 × г в течение 1 мин.
3. Анализ данных секвенирования
ПРИМЕЧАНИЕ: Для создания моделей вторичной структуры РНК из данных секвенирования DMS-MaP результирующие файлы .fastq должны быть обработаны несколькими различными шагами. Эти шаги могут быть автоматически выполнены с помощью
- Обрежьте последовательности адаптеров с помощью TrimGalore или Cutadapt.
- Сопоставьте чтение с эталонными последовательностями (формат .fasta) с помощью Bowtie2.
- Подсчитывайте показания с помощью специализированного программного обеспечения структуры РНК (например, DREEM14, RNA-Framework17 или аналогичного) и создавайте профили реактивности.
- (Необязательно) Кластеризуйте показания для поиска альтернативных конформаций РНК с помощью DREEM14, DRACO17, DANCE-MaP18 или аналогичных.
- Прогнозирование минимальной структуры свободной энергии на основе профилей реактивности с использованием RNAStructure12, ViennaRNA или аналогичных.
- Визуализируйте структуру РНК11 с помощью VARNA (https://varna.lri.fr/) или аналогичной.
ПРИМЕЧАНИЕ: Для практичности программное обеспечение, такое как DREEM (www.rnadreem.org) и RNA-Framework19 , широко включает шаги 1-5 в свои конвейеры, что упрощает процесс анализа. Однако с любым прогнозированием структуры следует обращаться с осторожностью (например, путем проверки согласия структуры с данными20).
Результаты
Ген-специфический in vitro DMS-MaP
Для изучения 5'UTR SARS2 первые 300 bp вируса были упорядочены как последовательность gBlock, наряду с тремя праймерами. Они включали в себя два праймера для распространения фрагмента («FW» и «RV») через ПЦР, а также один для прикрепления промотора T7 («FW-T7»). Эти последовательности можно увидеть в таблице 1.
| Имя | Последовательность (5'->3') |
| ФВ | ATTAAAGGTTTATACCTTCCCAGGTAAC |
| RV | GCAAACTGAGTTGGACGTGT |
| ФВ-Т7 | TAATACGACTCACTATAGG ATTAAAGGTTTATACCTTCCCAGGTAAC |
Таблица 1: Последовательность праймеров для DMS-MaP RT-PCR SARS-CoV2 5'UTR. Здесь FW-T7 и RV необходимы для генерации шаблона ДНК для транскрипции in vitro , RV используется в обратной транскрипции, а праймер-пара FW-RV используется в последующей ПЦР-амплификации кДНК. Праймеры отжигают до самого начала генома SARS-CoV2 (FW) и последовательности прямо вниз по течению от интересующей области. Сокращения: DMS-MaP = мутационное профилирование с секвенированием с использованием диметилсульфата; ОТ-ПЦР = полимеразная цепная реакция с обратной транскрипцией; SARS-CoV2 = тяжелый острый респираторный синдром-коронавирус 2; UTR = непереведенный регион; RV = обратная грунтовка; FW = прямая грунтовка.
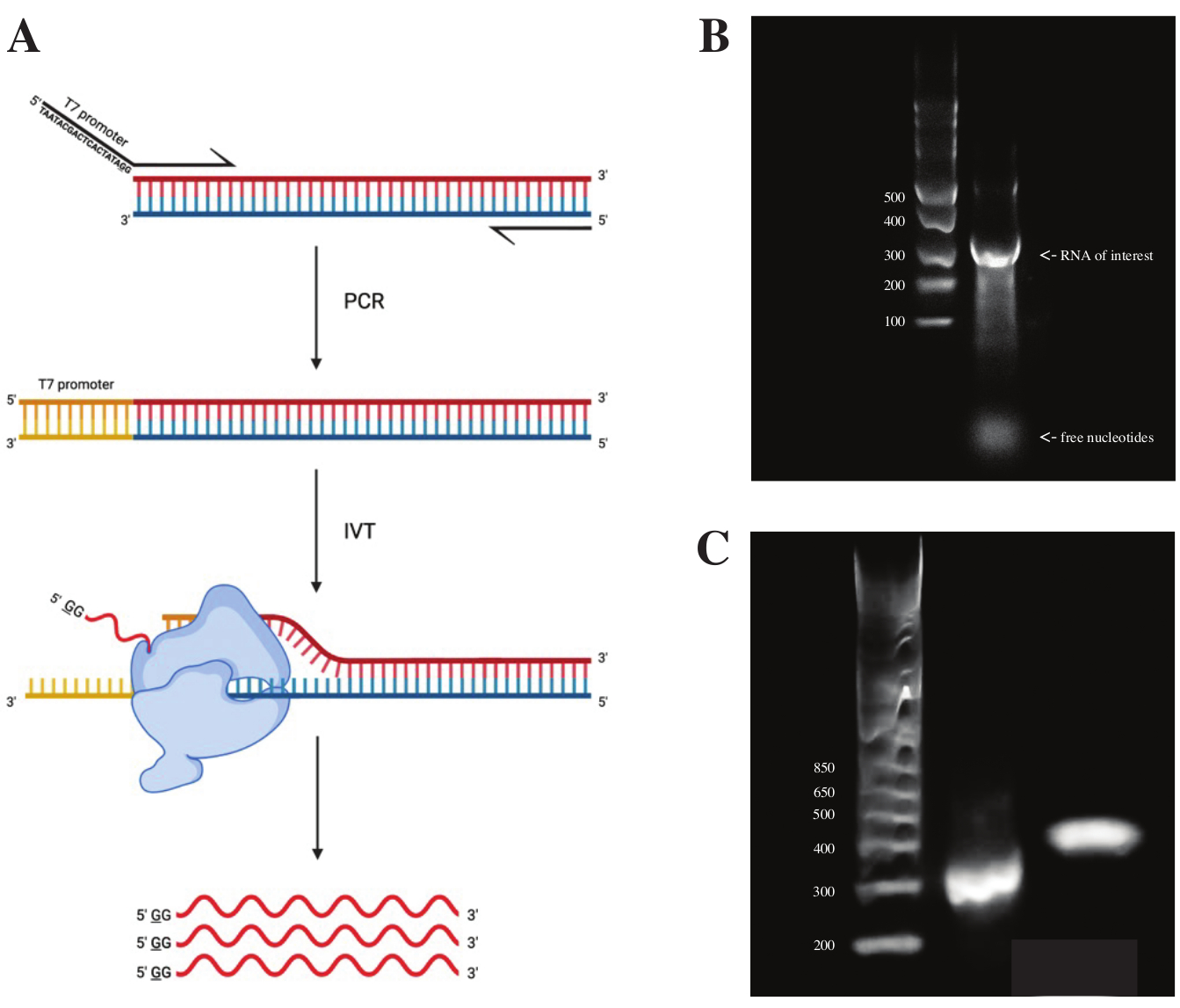
Для генерации РНК из фрагмента gBlock последовательность промотора полимеразы Т7 присоединяли с помощью перекрывающейся ПЦР с использованием премикса ПЦР в соответствии со схемой, показанной на фиг.2А. Из удлиненного фрагмента РНК генерировали с помощью набора транскрипции T7. Шаблон ДНК впоследствии переваривался с использованием ДНКазы и РНК, выделенных с использованием колонок RNA Clean & Concentrator.
Контроль качества транскрипции in vitro осуществлялся путем запуска продукта РНК на 1% агарозном геле вместе с лестницей ssRNA. Поскольку была видна только одна полоса, было проведено зондирование DMS in vitro и ОТ-ПЦР (см. Рисунок 2B).
Чтобы проверить успешность реакции ПЦР, образец запускали на 2% агарозном геле с использованием лестницы dsDNA. После индексации полоса должна работать на ~150 bp выше на том же геле, учитывая размер индексирующих грунтовок.

Рисунок 2: Транскрипция in vitro шаблона ДНК. (А) Чтобы in vitro транскрибировать шаблон ДНК, который еще не имеет внутреннего промотора РНК-полимеразы, шаблон должен быть сначала прикреплен путем перекрывающейся ПЦР. Это делается с помощью прямого праймера, который включает в себя последовательность TAATACGACTCACTATAGG (в случае РНК-полимеразы T7) перед первыми основаниями, перекрывающимися с нужным фрагментом. Подчеркнутое основание здесь символизирует место начала транскрипции полимеразы. После того, как промотор прикрепился к фрагменту дцДНК, он может быть транскрибирован полимеразой Т7. Важно отметить, что полимераза использует нить, противоположную упомянутой промоторной последовательности, в качестве шаблона (синий), эффективно создавая РНК, идентичную последовательности непосредственно после указанной промоторной последовательности (красный). (B) 1% агарозный гель с лестницей ssRNA (полоса 1) и транскрибируемый in vitro продукт РНК при 300 нт (полоса 2). (C) 2% агарозный гель с GeneRuler 1 кб плюс лестница (полоса 1), продукт ПЦР после ОТ-ПЦР, работающий при 300 бп (полоса 2), и индексированный фрагмент после подготовки библиотеки, работающий при 470 бп (полоса 3). Сокращения: ОТ-ПЦР = полимеразная цепная реакция с обратной транскрипцией; DMS = диметилсульфат; nt = нуклеотиды; dsDNA = двухцепочечная ДНК; ssRNA = одноцепочечная РНК. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Полный геном in vivo DMS-MaP с использованием инфицированных вирусом клеток
До лечения DMS клетки HCT-8 были инфицированы OC43. Когда цитопатический эффект (CPE) наблюдался через 4 дня после инфекции (dpi) (как показано на рисунке 3A), эти клетки обрабатывали, а РНК экстрагировали и рибоделировали. При запуске общей РНК на агарозном геле были видны две яркие полосы, составляющие субъединицы 40S и 60S рибосомы, которые составляют примерно 95% от общей массы РНК (см. Рисунок 3B). Когда экстракция РНК была неудачной или деградировала (например, несколькими циклами замораживания-оттаивания), продукты деградации РНК были видны на нижней части геля (см. Рисунок 3C, вторая полоса). Кроме того, после истощения рРНК две яркие полосы исчезли, оставив мазок в полосе (см. Рисунок 3C, третья полоса). Наконец, после подготовки библиотеки образцы имели различное распределение по размеру и были показаны в виде мазка на конечном геле PAGE. Полоса была вырезана между 200 нуклеотидами (nt) и 500 nt, в соответствии с 150 x 150 парным секвенированием, запланированным для анализа этих библиотек. Самое главное, что димеры адаптера, работающие на ~150 нт, были отделены (см. Рисунок 3D).

Рисунок 3: Контрольные точки in vivo DMS-MaP с инфицированными вирусом клетками. (A) Изображение световой микроскопии инфицированных вирусом клеток HCT-8, 4 дня dpi. Чтобы получить максимально возможный выход вирусной РНК из общей РНК при минимизации неблагоприятных эффектов из-за гибели клеток, DMS следует добавлять при начале CPE или даже до этого, как показано на изображении. (B) 1% агарозный гель с шестью образцами по 1 мкг общей РНК. В каждой полосе видны две яркие полосы, составляющие субъединицы 40S и 60S, поскольку рибосомная РНК составляет ~ 95% от общей РНК. Примечание: Внутриклеточное лечение DMS вызывает некоторую фрагментацию и размазывание РНК, но две полосы рРНК все еще должны быть видны. Легкая фрагментация после модификации допускается, потому что информация, содержащая мет метилирование, генерируется и сообщает о структуре РНК во время инкубации DMS, пока клетки еще живы. (C) 1% агарозный гель GeneRuler 1 кб плюс маркер лестничной ДНК (полоса 1), общая РНК, ранее хранившаяся при −80 °C в течение 6 месяцев (полоса 2) и рибоделепированная РНК (полоса 3). При длительном хранении РНК с несколькими циклами замораживания-оттаивания РНК начинает деградировать и, возможно, не должна использоваться для зондирующих экспериментов. Кроме того, после рибодеплирования общей РНК две яркие полосы, составляющие субъединицы 40S и 60S рибосомы, исчезают, и начинает проявляться мазок остаточных РНК. (D) Гель PAGE GeneRuler 1 кб плюс маркер лестничной ДНК (полоса 1) и библиотечный образец полногеномной подготовленной РНК. Гель должен быть иссечен в зависимости от потребностей в секвенировании. Для парного последовательного выполнения, охватывающего 150 циклов с обеих сторон, гель должен быть иссечен между 300 bp и 500 bp. Димеры адаптера (работающие при 170 bp) должны быть отделены. Сокращения: DMS-MaP = мутационное профилирование с секвенированием с использованием диметилсульфата; dpi = дни после заражения; CPE = цитопатический эффект. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
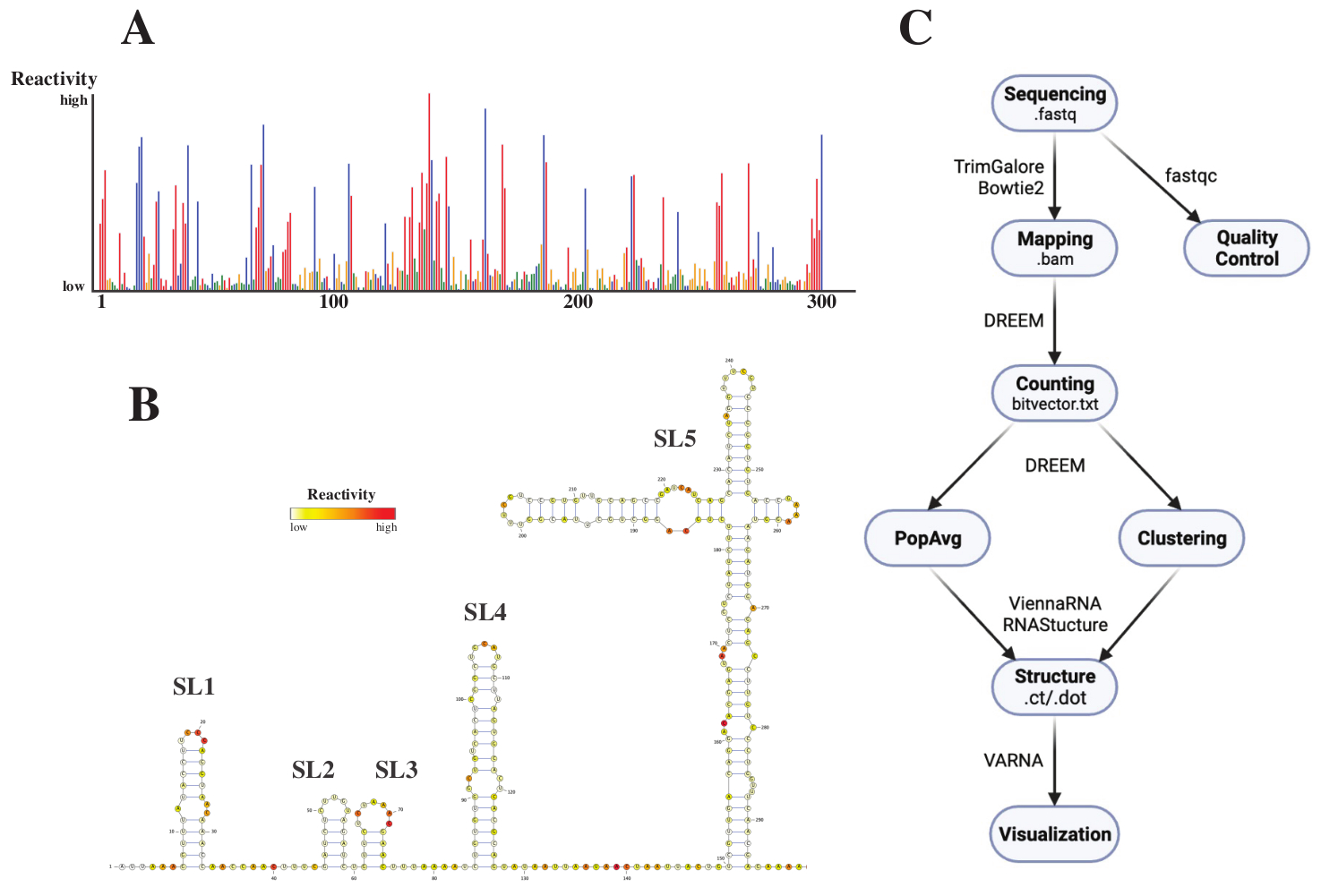
После виртуализации файлы .fastq анализировались путем отправки задания на веб-сервер DREEM (http://rnadreem.org/) вместе с файлом ссылок .fasta. Выходные данные, генерируемые сервером, включают файлы контроля качества, генерируемые fastqc (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) и TrimGalore (https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/), а также другие выходные файлы, содержащие средние частоты мутаций популяции. Помимо диаграммы, показывающей частоты мутаций в формате интерактивного .html (см. Рисунок 4A) и файла .csv с необработанными реактивациями на основание и файла struct_constraint.txt, читаемого несколькими программами прогнозирования структуры РНК, это также включает в себя файл bitvector.txt сообщающий о мутациях при считывании. Из них средние структуры населения были рассчитаны путем отправки файлов .fasta и struct_constraint.txt на веб-сервер RNAfold (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi). Это использует программное обеспечение ViennaRNA для генерации прогнозов структуры на основе минимальной свободной энергии, которые можно просмотреть онлайн или загрузить в формате ct или Vienna. Для генерации моделей структуры РНК эти загружаемые файлы были отправлены в VARNA (https://varna.lri.fr/, см. Рисунок 4B). Наконец, файлы bitvector.txt могут использоваться стабильной версией DREEM (https://codeocean.com/capsule/6175523/tree/v1) для поиска альтернативных конформаций РНК. Для получения моделей хорошей структуры с использованием DREEM следует обеспечить охват в размере 10 000 считываний на базу; для кластеризации может потребоваться до 100 000 операций чтения на базу. Обзор всего рабочего процесса можно найти на рисунке 4C.

Рисунок 4: Примерные данные, полученные в результате экспериментов по химическому зондированию SARS-CoV2 5'UTR. (A) Профиль реакционной способности первых 300 оснований генома SARS-CoV2, окрашенных основанием (A: красный, C: синий, U: зеленый, G: желтый). Необработанные реактивности рассчитываются как абсолютная частота мутаций, деленная на покрытие. Основания с открытой конформацией имеют высокие значения реакционной способности; базы, участвующие в сопряжении оснований, имеют низкие значения реактивности. вы и G не модифицируются DMS и имеют низкие значения реакционной способности, возникающие из-за неверности полимеразы. Прогнозы были сделаны с помощью веб-сервера DREEM. (B) Структурная модель SARS-CoV2 5'UTR, предсказанная на основе значений реактивности, сделанных с помощью VARNA. Основания с высокими показателями реактивности окрашены в красный цвет; основания с низкими значениями реактивности окрашены в белый цвет. (C) Рабочий процесс анализа DMS-MaP, начиная с файлов .fastq, полученных в результате виртуализации. Они могут контролироваться качеством с помощью fastqc; последовательности адаптеров обрезаются с помощью TrimGalore, а затем сопоставляются с эталонной последовательностью с помощью Bowtie2. Из полученных файлов .bam DREEM подсчитывает мутации в каждом чтении, создавая карту мутаций или файл .bitvector.txt. Они сообщают о мутациях каждого прочитанного в зависимости от положения, на основе которого могут быть созданы средние профили реактивности популяции. Альтернативно, битвекторы могут быть сгруппированы с использованием DREEM для поиска альтернативных конформаций РНК. Наконец, полученные структурные модели визуализируются с помощью программного обеспечения (например, VARNA). Сокращения: DMS-MaP = мутационное профилирование с секвенированием с использованием диметилсульфата; SARS-CoV2 = тяжелый острый респираторный синдром-коронавирус 2. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Обсуждение
Протокол здесь описывает, как исследовать РНК in vitro и в клетках с использованием экспериментов по мутационному профилированию DMS. Кроме того, он дает инструкции о том, как подготовить библиотеки для секвенирования Illumina для генерации генно-специфических данных и анализа полученных файлов .fastq. Кроме того, могут использоваться общегеномные библиотечные подходы. Тем не менее, генно-специфическая ОТ-ПЦР дает самые качественные и наиболее надежные данные. Поэтому при сравнении образцов важно убедиться, что они подготовлены с одинаковыми стратегиями секвенирования, так как генерация библиотеки вызывает некоторую предвзятость. Воспроизводимость всегда должна измеряться с помощью реплик.
Несколько мер предосторожности
РНК представляет собой нестабильную молекулу, которая чувствительна к деградации как через повышенные температуры, так и через РНКазы. Поэтому рекомендуются специальные меры — использование средств индивидуальной защиты (СИЗ), материала, не содержащего РНКазы, ингибиторов РНКАЗы. Самое главное, что РНК следует держать на льду, когда это возможно. Особенно это касается метилированной РНК, которая еще более чувствительна к высоким температурам.
Важно подтвердить, что интересующая структура РНК не чувствительна к концентрации ДМС и буферным условиям. Буферы, такие как 100 мМ Tris, 100 мМ MOPS и 100 мM HEPES при рН 7-7,5, дают высокий сигнал, но могут быть недостаточными для поддержания рН во время реакции21. Поскольку DMS гидролизуется в воде, что снижает pH, сильный буфер имеет решающее значение для поддержания нейтрального pH во время реакции модификации. Было показано, что добавление бицина помогает поддерживать рН в качестве слегка основного21 , но приводит к низкой модификации DMS на Gs и Us, что может быть информативным, но должно быть проанализировано отдельно из-за производства гораздо более низкого сигнала, чем As и Cs, и не обсуждается далее в этом протоколе.
В ген-специфической ОТ-ПЦР модифицированная РНК обратно транскрибируется в ДНК и амплифицируется фрагментами с помощью ПЦР. Хотя размер РНК теоретически может быть неограниченным, эти фрагменты ПЦР не должны превышать длину 400-500 пар оснований (bp), чтобы предотвратить смещение во время реакции обратной транскрипции. В идеале фрагменты должны находиться в рамках последовательности (т.е. если секвенирование проводится с использованием программы парного секвенирования 150 x 150 циклов, один фрагмент не должен превышать 300 бит/с). При использовании программ секвенирования с меньшим количеством циклов продукты ПЦР могут быть фрагментированы с помощью dsDNase. Кроме того, поскольку последовательности внутри последовательностей праймера не содержат никакой структурной информации, фрагменты должны перекрываться, когда исследуемая РНК содержит фрагмент >1. Реакции RT могут содержать несколько праймеров RT для разных фрагментов (до 10 различных праймеров RT). В зависимости от последовательностей, объединение праймеров RT может сделать обратную транскрипцию менее эффективной, но обычно работает хорошо. Каждую реакцию ПЦР следует проводить отдельно.
При зондировании РНК с DMS экспериментальные условия играют дополнительную роль, так как многие РНК термодинамически нестабильны и изменяют свою конформацию в зависимости от факторов окружающей среды, таких как температура. Чтобы избежать неровностей, условия эксперимента должны быть как можно более постоянными, в том числе в отношении времени реакции. Буферные условия, по-видимому, в определенной степени взаимозаменяемы 17,20,22,23 при сохранении основных условий — буферной способности и наличия одновалентных (Na) и двухвалентных ионов (Mg) — для обеспечения надлежащего сворачивания РНК24.
Что касается библиотечной подготовки модифицированных РНК, то необходимо учитывать несколько аспектов. Во-первых, как упоминалось ранее, модифицированные РНК менее стабильны, чем их немодифицированные аналоги, что означает, что им может потребоваться оптимизация времени фрагментации для оптимального распределения размеров фрагментов. Кроме того, некоторые наборы для подготовки библиотеки РНК, а также многие другие подходы RNAseq используют случайные праймеры в наборе обратной транскрипции. Это может привести к снижению охвата референта, особенно в 3' гена, и, в конечном счете, к недостаточной глубине охвата. Если охват определенного региона слишком низок, возможно, потребуется удалить эти основания из прогноза структуры. Помимо наборов ОТ-ПЦР и RNAseq для всего генома, могут использоваться и другие подходы к подготовке библиотек. Протоколы, которые включают лигирование 3' и/или 5' адапторов к РНК, выгодны при использовании небольших фрагментов РНК или когда следует избегать потери зондирующей информации в областях праймера.
Наконец, анализ экспериментов по химическому зондированию всегда должен интерпретироваться осторожно. В настоящее время не существует программного обеспечения, которое предсказывает структуру РНК любой РНК только из последовательности с высокой точностью. Хотя ограничения химического зондирования значительно повышают точность, создание хороших моделей для длинных РНК (>500 нт) по-прежнему является сложной задачей. Эти модели должны быть дополнительно проверены другими подходами и/или мутагенезом. Программное обеспечение для прогнозирования РНК оптимизирует максимальное количество пар оснований, тем самым значительно наказывая открытые конформации, которые могут не точно представлять сворачивание РНК5. Таким образом, полученная модель структуры должна быть проверена путем количественной оценки соглашения о прогнозировании с базовыми данными химического зондирования (например, AUROC) и между репликациями (например, mFMI), как показано на примере Lan et al.20.
В идеале для укрепления гипотезы следует использовать несколько экспериментов в разных системах, чтобы оспорить полученную структурную модель. Они могут включать использование in vitro и внутриклеточных подходов, компенсаторных мутаций и различных клеточных линий и видов. Более того, необработанные реактивности часто так же или даже более информативны, чем предсказания структуры, поскольку они записывают снимок «основной истины» складного ансамбля РНК. Таким образом, необработанные реактивности очень подходят и информативны для сравнения изменений структуры между различными условиями. Важно отметить, что структуры с самой низкой свободной энергией, рассчитанные с использованием ограничений химического зондирования с вычислительным прогнозированием, должны использоваться только в качестве начальной гипотезы для полной модели структуры.
Раскрытие информации
У авторов нет конфликта интересов, о которых можно было бы заявить.
Благодарности
Никакой
Материалы
| Name | Company | Catalog Number | Comments |
| 1 Kb Plus DNA Ladder | 10787018 | Thermo | |
| 2-mercaptoethanol | M6250-250ML | Sigma | |
| Acid-Phenol:Chloroform, pH 4.5 | AM9720 | Thermo | |
| Advantage PCR | 639206 | Takara | |
| CloneAmp HiFi PCR Premix | 639298 | Takara | |
| DMS | D186309 | Sigma | |
| dNTPs 10 mM each | U151B | Promega | |
| E-Gel EX Agarose Gels, 2% | G402022 | Thermo | precast agarose gels |
| Ethanol (200 proof) | E7023-4X4L | Sigma | |
| Falcon tubes, 15 mL, 50 mL | |||
| GlycoBlue | co-precipitant | ||
| HCT-8 cells | ATCC #CCL-244 | ||
| Invitrogen MgCl2 (1 M) | AM9530G | fisherscientific | |
| Isopropanol | 278475 | Sigma | |
| Megascript T7 transcription | AM1334 | Thermo | |
| NanoDrop spectrophotometer | |||
| Novex TBE Gels, 8%, 10 well | EC6215BOX | Thermo | |
| OC43 | ATCC #VR-1558 | ||
| RiboRuler Low Range RNA Ladder | SM1831 | Thermo | |
| RNAse H | M0297L | NEB | |
| Sodium Cacodylate, 0.4 M, pH 7.2 | 102090-964 | VWR | |
| Sodium hydroxide solution | S8263-150ML | Sigma | |
| SuperScript II Reverse Transcriptase for FSB and DTT | 18064014 | Thermo | |
| TGIRT-III Enzyme | TGIRT50 | Ingex | |
| The Oligo Clean & Concentrator | D4060 | Genesee | |
| The RNA Clean & Concentrator kits are RNA clean up kits | R1016 | Genesee | |
| TRIzol Reagents | 15596018 | Thermo | RNA isolation reagent |
| Water, (For RNA Work) (DEPC-Treated, DNASE, RNASE free/Mol. Biol.) | BP561-1 | fisherscientific | |
| xGen Broad-range RNA Library Prep 16rxn | 10009865 | IDT | |
| Zymo RNA clean and concentrator columns |
Ссылки
- Kim, S. H., et al. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA. Science. 185 (4149), 435-440 (1974).
- Robertus, J. D., et al. Structure of yeast phenylalanine tRNA at 3 Å resolution. Nature. 250 (467), 546-551 (1974).
- Zaug, A. J., Cech, T. R. In vitro splicing of the ribosomal RNA precursor in nuclei of Tetrahymena. Cell. 19 (2), 331-338 (1980).
- Zhao, Y., et al. NONCODE 2016: An informative and valuable data source of long non-coding RNAs. Nucleic Acids Research. 44, D203-D208 (2016).
- Vandivier, L. E., Anderson, S. J., Foley, S. W., Gregory, B. D. The conservation and function of RNA secondary structure in plants. Annual Review of Plant Biology. 67, 463(2016).
- Jumper, J., et al. Highly accurate protein structure prediction with AlphaFold. Nature. 596 (7873), 583-589 (2021).
- Das, R. RNA structure: A renaissance begins. Nature Methods. 18 (5), 439-439 (2021).
- Smola, M. J., Rice, G. M., Busan, S., Siegfried, N. A., Weeks, K. M. Selective 2′-hydroxyl acylation analyzed by primer extension and mutational profiling (SHAPE-MaP) for direct, versatile and accurate RNA structure analysis. Nature Protocols. 10 (11), 1643-1669 (2015).
- Mathews, D. H., et al. Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure. Proceedings of the National Academy of Sciences of the United States of America. 101 (19), 7287-7292 (2004).
- Zuker, M., Stiegler, P. Optimal computer folding of large RNA sequences using thermodynamics and auxiliary information. Nucleic Acids Research. 9 (1), 133-148 (1981).
- Lorenz, R., et al. ViennaRNA Package 2.0. Algorithms for Molecular Biology. 6, (2011).
- Reuter, J. S., Mathews, D. H. RNAstructure: Software for RNA secondary structure prediction and analysis. BMC Bioinformatics. 11, (2010).
- Wells, S. E., Hughes, J. M. X., Igel, A. H., Ares, M. Use of dimethyl sulfate to probe RNA structure in vivo. Methods in Enzymology. , 479-493 (2000).
- Tomezsko, P. J., et al. Determination of RNA structural diversity and its role in HIV-1 RNA splicing. Nature. 582 (7812), (2020).
- Zubradt, M., et al. DMS-MaPseq for genome-wide or targeted RNA structure probing in vivo. Nature Methods. 14 (1), (2017).
- Woodson, S. A. Compact intermediates in RNA folding. Annual Reviews in Biophysics. 39, (2010).
- Morandi, E., et al. Genome-scale deconvolution of RNA structure ensembles. Nature Methods. 18 (3), 249-252 (2021).
- Olson, S. W., et al. Discovery of a large-scale, cell-state-responsive allosteric switch in the 7SK RNA using DANCE-MaP. Molecular Cell. 82 (9), 1708-1723 (2022).
- Incarnato, D., Morandi, E., Simon, L. M., Oliviero, S. RNA Framework: An all-in-one toolkit for the analysis of RNA structures and post-transcriptional modifications. Nucleic Acids Research. 46 (16), (2018).
- Lan, T. C. T., et al. Secondary structural ensembles of the SARS-CoV-2 RNA genome in infected cells. Nature Communications. 13 (1), 1128(2022).
- Homan, P. J., et al. Single-molecule correlated chemical probing of RNA. Proceedings of the National Academy of Sciences of the United States of America. 111 (38), 13858-13863 (2014).
- Yang, S. L., et al. Comprehensive mapping of SARS-CoV-2 interactions in vivo reveals functional virus-host interactions. Nature Communications. 12 (1), 5113(2021).
- Manfredonia, I., et al. Genome-wide mapping of SARS-CoV-2 RNA structures identifies therapeutically-relevant elements. Nucleic Acids Research. 48 (22), 12436-12452 (2020).
- Fischer, N. M., Polěto, M. D., Steuer, J., vander Spoel, D. Influence of Na+ and Mg2+ ions on RNA structures studied with molecular dynamics simulations. Nucleic Acids Research. 46 (10), 4872-4882 (2018).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены